
Effect of Humic Substances as Feed Additive on the Growth Performance, Antioxidant Status, and Health Condition of African Catfish (Clarias gariepinus, Burchell 1822)

 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design
- HS0—control diet without HS supplement (0 g/100 g of feed);
- HS1—diet with 1% of HS (1 g/100 g of feed);
- HS3—diet with 3% of HS (3 g/100 g of feed); and
- HS6—diet with 6% of HS additive (6 g/100 g of feed).
2.2. Sampling and Calculations
2.3. Analysis of Blood Parameters
2.3.1. Biochemical Parameters
2.3.2. Antioxidant Status—Reduced (GSH) and Oxidized (GSSG) Glutathione
2.4. Statistical Analysis
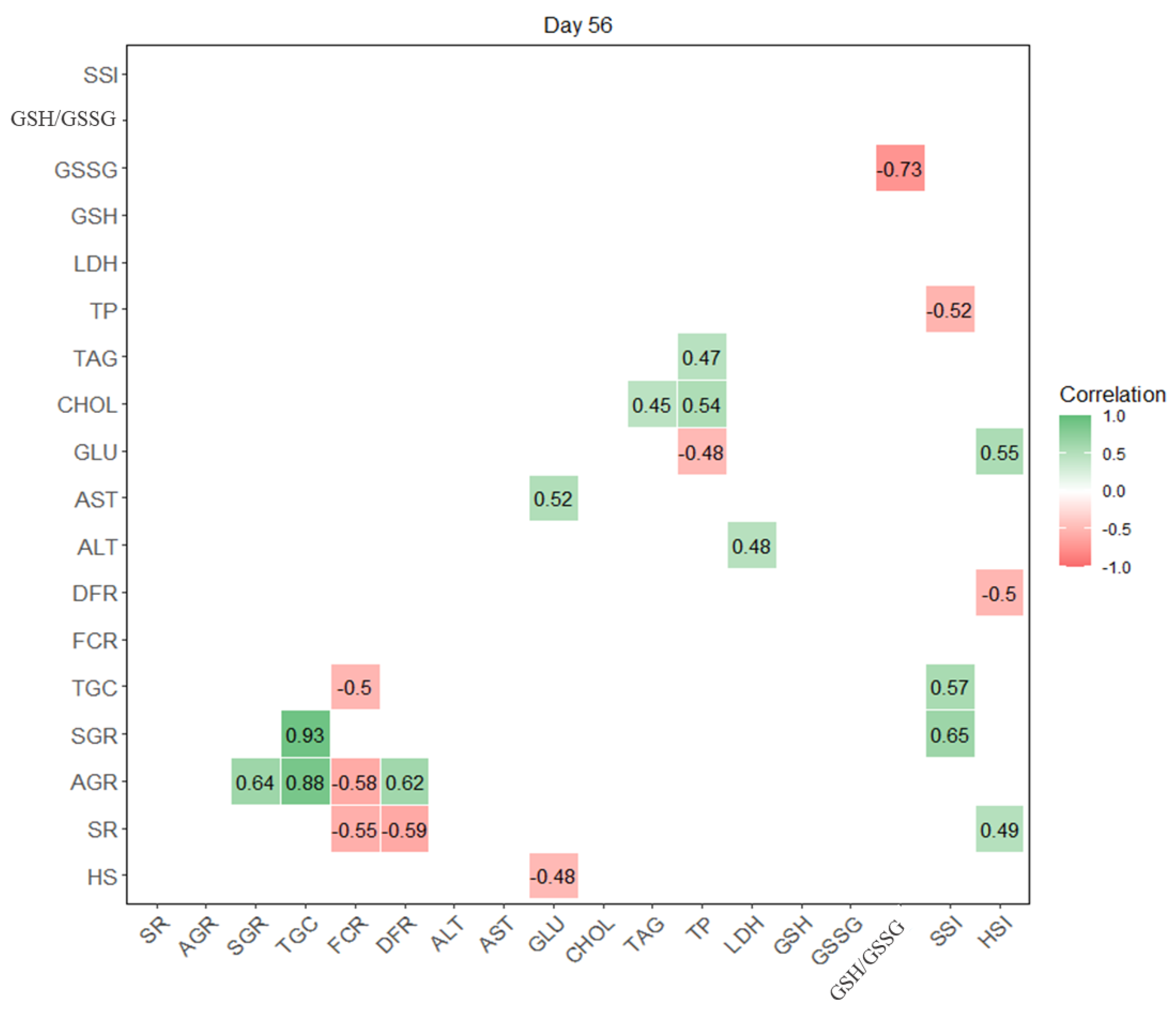
3. Results
4. Discussion
4.1. Humic Substances
4.2. Leonardite Mineraloid
4.3. Growth Performance
4.4. Food Conversion Ratio
4.5. Condition Factor and Somatic Indices
4.6. Biochemical Parameters
4.7. Antioxidant Status, Reduced (GSH) and Oxidized (GSSG) Glutathione
4.8. Overall Mortality
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. The State of World Fisheries and Aquaculture 2020: Sustainability in Action; FAO: Rome, Italy, 2020; 224p. [Google Scholar] [CrossRef]
- Hua, K.; Cobcroft, J.M.; Cole, A.; Condon, K.; Jerry, D.R.; Mangott, A.; Praeger, C.; Vucko, M.J.; Zeng, C.; Zenger, K.; et al. The future of aquatic protein: Implications for protein sources in aquaculture diets. One Earth 2019, 1, 316–329. [Google Scholar] [CrossRef] [Green Version]
- Dawood, A.O.M.; Koshio, S.; Esteban, M.A. Beneficial roles of feed additives as immunostimulants in aquaculture: A review. Rev. Aquacult. 2018, 10, 950–974. [Google Scholar] [CrossRef]
- Van de Nieuwegiessen, P.G.; Olwo, J.; Khong, S.; Verreth, J.A.J.; Schrama, J.W. Effects of age and stocking density on the welfare of African catfish, Clarias gariepinus Burchell. Aquaculture 2009, 288, 69–75. [Google Scholar] [CrossRef]
- Mota, V.C.; Limbu, P.; Martins, C.I.M.; Eding, E.H.; Verreth, J.A.J. The effect of nearly closed RAS on the feed intake and growth of Nile tilapia (Oreochromis niloticus), African catfish (Clarias gariepinus) and European eel (Anguilla anguilla). Aquac. Eng. 2015, 68, 1–5. [Google Scholar] [CrossRef]
- Ajiboye, A.O.; Awogbade, A.A.; Babalola, O.A. Effects of water exchange on water quality parameters, nutrient utilization and growth of African catfish (Clarias gariepinus). Int. J. Livest. Prod. 2015, 6, 57–60. [Google Scholar] [CrossRef]
- Schram, E.; Roques, J.A.C.; Abbink, W.; Spanings, T.; de Vries, P.; Bierman, S.; van de Vis, H.; Flik, G. The impact of elevated water ammonia concentration on physiology, growth and feed intake of African catfish (Clarias gariepinus). Aquaculture 2010, 306, 108–115. [Google Scholar] [CrossRef]
- Appelbaum, S.; Kamler, E. Survival, growth, metabolism and behaviour of Clarias gariepinus (Burchell 1822) early stages under different light conditions. Aquacult. Eng. 2000, 22, 269–287. [Google Scholar] [CrossRef]
- Kouřil, J.; Drozd, B.; Prokešová, M.; Stejskal, V. Intensive Rearing of African Sharptooth Catfish (Clarias gariepinus); Methodological Edition; FFPW USB: Vodnany, Czech Republic, 2013; Volume 138, 60p, ISBN 978-80-87437-79-7. (In Czech) [Google Scholar]
- FAO. FAO Yearbook. Fishery and Aquaculture Statistics 2009; FAO: Rome, Italy, 2011; Volume B13, pp. 63–72. ISBN 978-92-5-006975-3. [Google Scholar]
- FAO. FAO Yearbook. Fishery and Aquaculture Statistics 2018; FAO: Rome, Italy, 2020; Volume B13, pp. 67–78. [Google Scholar] [CrossRef]
- Islam, K.M.S.; Schuhmacher, A.; Gropp, J.M. Humic Acid substances in animal agriculture humic acid substances in animal agriculture. Pak. J. Nutr. 2005, 4, 126–134. [Google Scholar] [CrossRef] [Green Version]
- Steinberg, C.E.W.; Höss, S.; Kloas, W.; Lutz, I.; Meinelt, T.; Pflugmacher, S.; Wiegand, C. Hormonelike effects of humic substances on fish, amphibians, and invertebrates. Environ. Toxicol. 2004, 70, 409–411. [Google Scholar] [CrossRef]
- Steinberg, C.E.W.; Kamara, S.; Prokhotskaya, V.Y.; Manusadžianas, L.; Karasyova, T.A.; Timofeyev, M.A.; Jie, Z.; Paul, A.; Meinelt, T.; Farjalla, V.F.; et al. Dissolved humic substances—Ecological driving forces from the individual to the ecosystem level? Freshw. Biol. 2006, 51, 1189–1210. [Google Scholar] [CrossRef]
- Mcdonald, S.; Bishop, A.G.; Prenzler, P.D.; Robards, K. Analytical chemistry of freshwater humic substances. Anal. Chim. Acta 2004, 527, 105–124. [Google Scholar] [CrossRef]
- Rupiasih, N.N.; Vidyasagar, P.B. A Review: Compositions, structures, properties and applications of humic substances. JAST 2005, 8, 16–25. [Google Scholar]
- Steinberg, C.E.W.; Meinelt, T.; Timofeyev, M.A.; Bittner, M.; Menzel, R. Humic substances part 2: Interactions with organisms. Environ. Sci. Pollut. Res. 2008, 15, 128–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meinelt, T.; Paul, A.; Phan, T.M.; Zwirnmann, E.; Krüger, A.; Wienke, A.; Steinberg, C.E.W. Reduction in vegetative growth of the water mold Saprolegnia parasitica (Coker) by humic substance of different qualities. Aquat. Toxicol. 2007, 83, 93–103. [Google Scholar] [CrossRef]
- Peña-Méndez, E.M.; Havel, J.; Patočka, J. Humic substances—Compounds of still unknown structure: Applications in agriculture, industry, environment, and biomedicine: A review. J. Appl. Biomed. 2005, 3, 13–24. [Google Scholar] [CrossRef] [Green Version]
- Steinberg, C.E.W.; Paul, A.; Pflugmacher, S.; Meinelt, T.; Klöcking, R.; Wiegand, C. Pure humic substances have the potential to act as xenobiotic chemicals—A review. Fresenius Environ. Bull. 2003, 12, 391–401. [Google Scholar]
- Abdel-Wahab, A.M.; El-Refaee, A.M.E.; Ammar, A.A. Effects of humic acid as feed additive in improvement of nonspecific immune response and disease resistance in common carp (Cyprinus carpio). Egypt. J. Aquacult. 2012, 2, 83–90. [Google Scholar]
- Sharaf, M.M.; Tag, H.M. Growth performance, gill, liver and kidney histomorphology of common carp (Cyprinus carpio) fingerlings fed humic acid supplemented diets. Egypt. J. Exp. Biol. 2011, 7, 285–294. [Google Scholar]
- Lieke, T.; Steinberg, C.E.W.; Pan, B.; Perminova, I.V.; Meinelt, T.; Knopf, K.; Kloas, W. Phenol-rich fulvic acid as a water additive enhances growth, reduces stress, and stimulates the immune system of fish in aquaculture. Sci. Rep. 2021, 11. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; He, J.; He, Z.; Li, Z.; Zhao, B.; Mu, Y.; Lee, J.-Y.; Chu, Z. Effects of fulvic acid on growth performance and intestinal health of juvenile loach Paramisgurnus dabryanus (Sauvage). Fish Shellfish Immunol. 2017, 62, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Meinelt, T.; Schreckenbach, K.; Knopf, K.; Wienke, A.; Stüber, A.; Steinberg, C.E.W. Humic substances affect physiological condition and sex ratio of swordtail (Xiphophorus helleri Heckel). Aquat. Sci. 2004, 66, 239–245. [Google Scholar] [CrossRef]
- Yilmaz, S.; Ergun, S.; Çelik, E.S.; Yigit, M. Effects of dietary humic acid on growth performance, haemato-immunological and physiological responses and resistance of Rainbow trout, Oncorhynchus mykiss to Yersinia ruckeri. Aquac. Res. 2018, 49, 3338–3349. [Google Scholar] [CrossRef]
- Yamin, G.; Falk, R.; Avtalion, R.R.; Shoshana, N.; Ofek, T.; Smirnov, R.; Rubenstein, G.; van Rijn, J. The protective effect of humic-rich substances on atypical Aeromonas salmonicida subsp. salmonicida infection in common carp (Cyprinus carpio L.). J. Fish Dis. 2017, 40, 1783–1790. [Google Scholar] [CrossRef]
- Yamin, G.; Zilberg, D.; Levy, G.; varn Rijn, J. The protective effect of humic-rich substances from monogenean parasites infecting the guppy (Poecilia reticulata). Aquaculture 2017, 479, 487–489. [Google Scholar] [CrossRef]
- Da Costa, S.T.; Sutili, F.J.; Gressler, L.T.; Loebens, L.; Colpo, C.V.; Lazzari, R.; Baldisserotto, B. Growth of silver catfish (Rhamdia quelen) exposed to acidic pH at different humic acid levels. Ciênc. Rural. 2016, 46, 1094–1099. [Google Scholar] [CrossRef] [Green Version]
- Nakagawa, J.; Iwasaki, T.; Kodama, H. Protection against Flavobacterium psychrophilum infection (cold water disease) in Ayu fish (Plecoglossus altivelis) by oral administration of humus extract. J. Vet. Med. Sci. 2009, 71, 1487–1491. [Google Scholar] [CrossRef] [Green Version]
- Lang, Š.; Kopp, R.; Mareš, J. Verified Technology about Biofilter Starting in Danish Type of Intensive Recirculating Facility for Fish Farming; Verified technology; Mendel University: Brno, Czech Republic, 2015; 23p, ISBN 978-80-7509-375-2. (In Czech) [Google Scholar]
- Okomoda, V.T.; Aminem, W.; Hassan, A.; Martins, C.O. Effects of feeding frequency on fry and fingerlings of African catfish Clarias gariepinus. Aquaculture 2019, 511. [Google Scholar] [CrossRef]
- Baßmann, B.; Brenner, M.; Palm, H.W. Stress and welfare of African catfish (Clarias gariepinus Burchell, 1822) in a coupled aquaponic system. Water 2017, 9, 504. [Google Scholar] [CrossRef] [Green Version]
- Thorarensen, H.; Kubiriza, G.K.; Imsland, A.K. Experimental design and statistical analyses of fish growth studies. Aquaculture 2015, 448, 483–490. [Google Scholar] [CrossRef]
- Arif, M.; Alagawany, M.; Abd El-Hack, M.E.; Saeed, M.; Arain, M.A.; Elnesr, S.S. Humic acid as a feed additive in poultry diets: A review. Iran. J. Vet. Res. 2019, 20, 167–172. [Google Scholar] [PubMed]
- Olk, D.C.; Dinnes, D.L.; Scoresby, J.R.; Callaway, C.R.; Darlington, J.W. Humic products in agriculture: Potential benefits and research challenges—A review. JSSS 2018, 18, 2881–2891. [Google Scholar] [CrossRef]
- Chianese, S.; Fenti, A.; Iovino, P.; Musmarra, D.; Salvestrini, S. Sorption of organic pollutants by humic acids: A review. Molecules 2020, 25, 918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qian, S.; Ding, W.; Li, Y.; Liu, G.; Sun, J.; Ding, Q. Characterization of humic acids derived from leonardite using a solid-state NMR spectroscopy and effects of humic acids on growth and nutrient uptake of snap bean. Chem. Speciat. Bioavailab. 2015, 27, 156–161. [Google Scholar] [CrossRef] [Green Version]
- Tan, K.H. Humic Matter in Soil and the Environment: Principles and Controversies, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2014; 495p. [Google Scholar] [CrossRef]
- Thiel, K.D.; Helbig, B.; Klöcking, R.; Wutzler, P.; Sprössig, M.; Schweizer, H. Comparison of the in vitro activities of ammonium humate and of enzymically oxidized chlorogenic and caffeic acids against type 1 and type 2 human herpes virus. Pharmazie 1981, 36, 50–53. [Google Scholar] [PubMed]
- Boggs, S.; Livermore, D.; Seitz, M.G. Humic Substances in Natural Waters and Their Complexation with Trace Metals and Radionuclides: A Review; Argonne National Lab: Chicago, IL, USA, 1985; 224p. [Google Scholar] [CrossRef] [Green Version]
- Aderolu, A.Z.; Ogun, A.; Sanni, R.A.; Oguntoyinbo, F.A. Growth response of juvenile catfish (Clarias gariepinus) fed diets supplemented with Lactobacillus sp. Inclusion into feeds and cultured water. Nig. Vet. J. 2013, 34, 684–690. [Google Scholar]
- Hossain, M.A.R.; Haylor, G.S.; Beveridge, M.C.M. Effect of feeding time and frequency on the growth and feed utilization of African catfish Clarias gariepinus (Burchell 1822) fingerlings. Aquaculture Research 2001, 32, 999–1004. [Google Scholar] [CrossRef]
- Kim, S.G.; Park, D.K.; Jang, S.W.; Lee, J.S.; Kim, S.S.; Chung, M.H. Effects of dietary benzo[a]pyrene on growth and hematological parameters in juvenile rockfish, Sebastes schlegeli (Hilgendorf). Bull. Environ. Contam. Toxicol. 2008, 81, 470–474. [Google Scholar] [CrossRef]
- Kolářová, J.; Velíšek, J. Determination and Evaluation of the Biochemical Profile of Fish Blood; Methodological Edition; FFPW USB: Vodnany, Czech Republic, 2012; Volume 135, 54p, ISBN 978-80-87437-58-2. (In Czech) [Google Scholar]
- Abalaka, S.E. Evaluation of the haematology and biochemistry of Clarias gariepinus as biomakers of environmental pollution in Tiga dam, Nigeria. Braz. Arch. Biol. Technol. 2013, 56, 371–376. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Chen, J.; Lin, J.; Wang, Z.; Bian, H.; Cai, X.; Hao, C. Combined experimental and theoretical study of photoinduced toxicity of an antraquinone dye intermediate to Daphnia magna. Environ. Toxicol. Chem. 2009, 28, 846–852. [Google Scholar] [CrossRef]
- Lushchak, O.V.; Kubrak, O.I.; Lozinsky, O.V.; Storey, J.M.; Storey, K.B.; Lushchak, V.I. Chromium (III) induces oxidative stress in goldfish liver and kidney. Aquat. Toxicol. 2009, 93, 45–52. [Google Scholar] [CrossRef]
- Lushchak, V.I. Environmentally induced oxidative stress in aquatic animals. Aquat. Toxicol. 2011, 101, 13–30. [Google Scholar] [CrossRef]
- Borkovic, S.S.; Pavlovic, S.Z.; Kovacevic, T.B.; Stajn, A.S.; Petrovic, V.M.; Saičić, Z.S. Antioxidant defence enzyme activities in hepatopancreas, gills and muscle of spiny cheek crayfish (Orconectes limosus) from the River Danube. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2008, 147, 122–128. [Google Scholar] [CrossRef]
- Ojopagogo, Y.A.; Adewale, I.O.; Adeyemi, J.A.; Afolayan, A. Some novel features of glutathione transferase from juvenile catfish (Clarias gariepinus) exposed to lindane-contaminated water. Perspect. Sci. 2015, 4, 62–65. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Shen, H.; Wang, X.; Wu, J.; Xue, Y. Effects of chronic exposure of 2, 4 dichlorophenol on the antioxidant system in liver of freshwater fish Carassius auratus. Chemosphere 2004, 55, 167–174. [Google Scholar] [CrossRef] [PubMed]
- Kodama, H.; Denso; Nakagawa, T. Protection against atypical Aeromonas salmonicida infection in carp (Cyprinus carpio L.) by oral administration of humus extract. J. Vet. Med. Sci. 2007, 69, 405–408. [Google Scholar] [CrossRef] [Green Version]
- Peuranen, S.; Vuorinen, P.J.; Vuorinen, M.; Hollender, A. The effects of iron, humic acids and low pH on the gills and physiology of brown trout, Salmo trutta. Ann. Zool. Fenn. 1994, 31, 389–396. [Google Scholar]
- Hammock, D.; Huang, C.C.; Mort, G.; Swinehart, J.H. The effect of humic acid on the uptake of mercury(II), cadmium(II), and zinc(II) by chinook salmon (Oncorhynchus tshawytscha) eggs. Arch. Environ. Contam. Toxicol. 2003, 44, 83–88. [Google Scholar] [CrossRef]
- Meinelt, T.; Kroupova, H.; Stüber, A.; Rennerta, B.; Wienke, A.; Steinberg, C.E.W. Can dissolved aquatic humic substances reduce the toxicity of ammonia and nitrite in recirculating aquaculture systems? Aquaculture 2010, 306, 378–383. [Google Scholar] [CrossRef]
- Holland, A.; Duivenvoorden, L.J.; Kinnear, S.H.W. The double-edged sword of humic substances: Contrasting their effect on respiratory stress in eastern rainbow fish exposed to low pH. Environ. Sci. Pollut. Res. 2014, 21, 1701–1707. [Google Scholar] [CrossRef]
- Lieke, T.; Meinelt, T.; Hoseinifar, S.H.; Pan, B.; Straus, D.L.; Steinberg, C.E.W. Sustainable aquaculture requires environmental-friendly treatment strategies for fish diseases. Rev. Aquacult. 2019, 12, 943–965. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Concentration | Elements |
|---|---|
| 25–40% | C, K |
| 1–2% | H, Ca, Si, Fe |
| 0.1–1% | N, Al, S, Cl, Sr |
| 0.01–0.1% | Na, Mg, Ti, V, Cr, Ni, Zr, Sb, Cs |
| 0.001–0.01% | Zn, Ga, Br, Rb, Y, Nb, Cd, In, Sn, Te, Hf, Hg |
| <0.001% | P, Mn, Co, Cu, Ge, As, Se, Mo, Ag, I, Ba, La, Ce, Ta, W, Tl, Pb, Bi, Th, U |
| Components | HS0 | HS1 | HS3 | HS6 |
|---|---|---|---|---|
| Dry matter (% g of feed) | 90.39 | 90.43 | 90.52 | 90.64 |
| Crude protein (% of feed) | 42.00 | 42.00 | 42.01 | 42.02 |
| Crude lipid (% of feed) | 12.00 | 12.00 | 12.00 | 12.00 |
| Ash (% of feed) | 6.80 | 6.82 | 6.87 | 6.95 |
| Parameters | Initial | HS0 | HS1 | HS3 | HS6 | Ftest | n | p |
|---|---|---|---|---|---|---|---|---|
| Period (d) | Day 0 | Day 56 | Day 56 | Day 56 | Day 56 | Day 56 | Day 56 | Day 56 |
| B (kg) | 2.94 ± 0.09 | 10.71 ± 0.51 | 10.47 ± 0.86 | 11.14 ± 0.51 | 10.81 ± 0.84 | 0.82 1WA | 20 | 0.50 |
| BW (g) | 32.38 ± 1.96 | 173.85 ± 6.16 | 169.42 ± 9.27 | 174.53 ± 2.00 | 176.36 ± 12.63 | 1.83KW | 20 | 0.61 |
| TL (mm) | 162.45 ± 3.36 | 272.35 ±4.02 | 271.37 ± 6.95 | 275.05 ± 2.07 | 274.01 ± 7.17 | 0.45 1WA | 20 | 0.72 |
| SL (mm) | 144.31 ± 3.33 | 242.12 ± 4.26 | 238.43 ± 6.24 | 247.10 ± 1.84 | 246.22 ± 6.63 | 3.07 1WA | 20 | 0.06 |
| SGR (% d−1) | - | 2.52 ± 0.87 | 2.52 ± 0.59 | 2.40 ± 0.40 | 2.92 ± 0.70 | 0.57 1WA | 20 | 0.64 |
| AGR (% d−1) | - | 2.53 ± 0.13 | 2.46 ± 0.16 | 2.53 ± 0.03 | 2.56 ± 0.21 | 0.37 1WA | 20 | 0.77 |
| FCR | - | 1.11 ± 0.16 | 1.17 ± 0.30 | 1.06 ± 0.11 | 1.03 ± 0.14 | 3.72 KW | 20 | 0.29 |
| DFR (feed fish−1day−1) | - | 2.40 ± 0.07 | 2.35 ± 0.11 | 2.38 ± 0.06 | 2.42 ± 0.14 | 0.47 1WA | 20 | 0.71 |
| CV (%) | 25.60 ± 2.58 | 47.76 ± 4.62 | 42.60 ± 5.30 | 42.41 ± 3.00 | 40.55 ± 3.31 | 2.76 1WA | 20 | 0.08 |
| K | 0.75 ± 0.01 | 0.86 ± 0.02 | 0.85 ± 0.02 | 0.84 ± 0.01 | 0.86 ± 0.02 | 1.34 1WA | 20 | 0.30 |
| TGC | 1.59 ± 0.08 | 1.57 ± 0.08 | 1.58 ± 0.03 | 1.58 ± 0.07 | 0.06 1WA | 20 | 0.98 | |
| HSI (%) | - | 1.33 ± 0.38 | 1.23 ± 0.04 | 1.28 ± 0.04 | 1.19 ± 0.04 | 0.34 1WA | 20 | 0.80 |
| SSI (%) | - | 0.04 ± 0.02 | 0.04 ± 0.02 | 0.04 ± 0.02 | 0.04 ± 0.01 | 0.56 1WA | 20 | 0.65 |
| OM (%) | - | 31.56 ± 1.49 | 31.33 ± 3.88 | 29.11 ± 2.77 | 31.11 ± 5.03 | 0.50 1WA | 20 | 0.68 |
| Parameters | Initial | HS0 | HS1 | HS3 | HS6 | F | n | p |
|---|---|---|---|---|---|---|---|---|
| Period (d) | Day 0 | Day 56 | Day 56 | Day 56 | Day 56 | Day 56 | Day 56 | Day 56 |
| TP (g L−1) | 27.23 ± 4.95 | 30.14 ± 1.80 | 26.87± 8.88 | 32.29 ± 5.08 | 30.80 ± 4.43 | 1.90 KW | 40 | 0.59 |
| ALT (ukat L−1) | 3.36 ± 1.41 | 0.49 ± 0.27 | 0.35 ± 0.18 | 0.51 ± 0.20 | 0.53 ± 0.12 | 1.59 1WA | 40 | 0.21 |
| AST (ukat L−1) | 2.75 ± 0.82 | 2.26 ± 2.34 | 1.31 ± 0.46 | 1.40 ± 0.34 | 1.20 ± 0.29 | 3.54 KW | 40 | 0.32 |
| CHOL (mmol L−1) | 3.25 ± 0.65 | 2.93 ± 0.27 | 2.68 ± 0.34 | 2.82 ± 0.43 | 2.82 ± 0.51 | 0.62 1WA | 40 | 0.60 |
| TAG (mmol L−1) | 1.49 ± 1.30 | 1.34 ± 1.19 | 0.95 ± 0.22 | 1.71 ± 1.63 | 0.91 ± 0.32 | 3.13 KW | 40 | 0.37 |
| LDH (ukat L−1) | 6.03 ± 2.52 | 3.76 ± 1.76 | 2.73 ± 1.28 | 3.76 ± 1.64 | 2.69 ± 1.25 | 4.81 KW | 40 | 0.19 |
| GLU (mmol L−1) | 4.83 ± 3.85 | 4.73 ± 1.68a | 4.78 ± 1.58a | 4.59 ± 0.66a | 3.20 ± 1.37b | 2.99 1WA | 40 | 0.04 |
| GSH (µM) | 204.65 ± 41.28 | 187.67 ± 68.42 | 174.60 ± 51.89 | 170.78 ± 43.52 | 171.23 ± 29.77 | 0.25 1WA | 40 | 0.86 |
| GSSG (µM) | 43.89 ± 15.65 | 8.74 ± 4.20 | 8.18 ± 4.12 | 6.99 ± 4.00 | 8.61 ± 3.32 | 0.41 1WA | 40 | 0.74 |
| GSH/GSSG (µM) | 2.98 ± 1.26 | 24.90 ± 19.12 | 25.45 ± 16.14 | 30.90 ± 24.05 | 23.99 ± 19.02 | 1.39 KW | 40 | 0.71 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Prokešová, M.; Bušová, M.; Zare, M.; Tran, H.Q.; Kučerová, E.; Ivanova, A.P.; Gebauer, T.; Stejskal, V. Effect of Humic Substances as Feed Additive on the Growth Performance, Antioxidant Status, and Health Condition of African Catfish (Clarias gariepinus, Burchell 1822). Animals 2021, 11, 2266. https://doi.org/10.3390/ani11082266
Prokešová M, Bušová M, Zare M, Tran HQ, Kučerová E, Ivanova AP, Gebauer T, Stejskal V. Effect of Humic Substances as Feed Additive on the Growth Performance, Antioxidant Status, and Health Condition of African Catfish (Clarias gariepinus, Burchell 1822). Animals. 2021; 11(8):2266. https://doi.org/10.3390/ani11082266
Chicago/Turabian StyleProkešová, Markéta, Milena Bušová, Mahyar Zare, Hung Quang Tran, Eliška Kučerová, Anna Pavlovna Ivanova, Tatyana Gebauer, and Vlastimil Stejskal. 2021. "Effect of Humic Substances as Feed Additive on the Growth Performance, Antioxidant Status, and Health Condition of African Catfish (Clarias gariepinus, Burchell 1822)" Animals 11, no. 8: 2266. https://doi.org/10.3390/ani11082266