
First Insight into Groundwater Copepods of the Polish Lowland


1
Department of Hydrobiology, Faculty of Biology, University of Białystok, Ciołkowskiego 1J, 15-245 Białystok, Poland
2
Department of Zoology and Genetics, Faculty of Biology, University of Białystok, Ciołkowskiego 1J, 15-245 Białystok, Poland
3
Doctoral School of Exact and Natural Sciences, University of Białystok, Ciołkowskiego 1K, 15-245 Białystok, Poland
*
Author to whom correspondence should be addressed.
Water 2021, 13(15), 2086; https://doi.org/10.3390/w13152086
Submission received: 11 July 2021
/
Revised: 25 July 2021
/
Accepted: 28 July 2021
/
Published: 30 July 2021
(This article belongs to the Special Issue Aquatic Biodiversity: Threats and Conservation)
Abstract
:Our results are the first insight into groundwater copepods of the Polish Lowland. The sampling was conducted in 28 wells in north-eastern Poland, and Copepoda were present in 16 wells. We have identified six Copepoda species and one Cladocera. We have classified four species as stygophiles—Eucyclops serrulatus, Diacyclops bisetosus, Diacyclops crassicaudis, and Cyclops furcifer. These species were frequently found in studied wells of different regions of north-eastern Poland, often in high numbers, and females with egg sacs were observed. We present a detailed morphological description of these species, together with molecular characteristics based on mitochondrial DNA markers (COI gene) for E. serrulatus, D. bisetosus, and D. crassicaudis, and 12S ribosomal RNA for C. furcifer. We also present the development of abnormal structures in one specimen of D. crassicaudis, where the upper part of furcal rami was fused to form a single plate.
1. Introduction
Subterranean habitats fulfill the requirements of experimental model systems to address general questions in ecology and evolution [1]. Groundwaters are populated by a specialized fauna called stygobionts and by accidental taxa (stygoxenes) temporarily imported from the surface. Stygophiles are intermediate between stygobionts and stygoxenes—they spend part of their life below ground and may even be more common in groundwater than in surface waters [2]. It is estimated that about 40% of the European crustacean fauna is represented by stygobiontic species, among which dominate copepods with about 1000 species and subspecies known from continental groundwater [3]. Most of them belong to the Harpacticoida (about 600 species) and Cyclopoida (about 300 species). Moreover, almost half of the newly described copepods are from groundwater habitats [3].
There are some regions with well-recognized groundwater copepods, and there are regions for which there is no information. Generally, groundwater fauna is quite well explored in mountain regions of southern Europe (Alps, Apennines, Pyrenees, Carpathian, and Balkan Mountains), and these regions are considered biodiversity hotspots of stygobiontic Harpacticoida [4]. There is some information about groundwater copepods from the western part of the European lowlands (Netherland, Germany, Denmark), contrary to the central part of the European lowlands. The groundwater of central and northern Poland is quite a forgotten habitat, contrary to the mountain region of Poland. A preliminary study of groundwater copepods from wells in Carpathian flysch (Poland) revealed the presence of four species [5], while the study of different subterranean habitats of southern Poland by Kur (2020) indicated the presence of 22 Cyclopoida species, but only one stygobiont [6]. The preliminary research in the European lowlands revealed two Harpacticoida species new for the Polish fauna in the springs and wells of north-eastern Poland, Elaphoidella elaphoides (Chappuis, 1923) and Bryocamptus (Rheocamptus) spinulosus (Borutzky, 1934) [7]. One problem is that Harpacticoida is the least known group of Copepoda in freshwater habitats in Poland because the last detailed taxonomic studies of Harpacticoida in Polish inland waters were carried out about 100 years ago [7].
The main aim of this research is to give new insights into groundwater copepods (Cyclopoida, Harpacticoida, Calanoida) of the Polish lowlands. We present a detailed morphological description of each species together with molecular characteristics (COI gene or 12S RNA). Currently, international databases (GenBank, BOLD System) contain single barcodes for each genus of copepods that are found in the groundwater. The other problem is that most of the available barcodes did not have sufficient morphological description and, thus, proper taxonomic identification, due to the well-known taxonomy crisis [8]. Therefore, presented in this article are detailed morphological descriptions of each species together with genetics analyzes of mtDNA molecular markers, which will be the cornerstone for future biodiversity analyzes based on eDNA metabarcoding because this method needs reference databases with nucleotide sequences for accurately identified species [9]. The other aim of this study is to determine which species found in the groundwater of Polish lowlands could be considered as stygobiont or stygophiles.
2. Materials and Methods
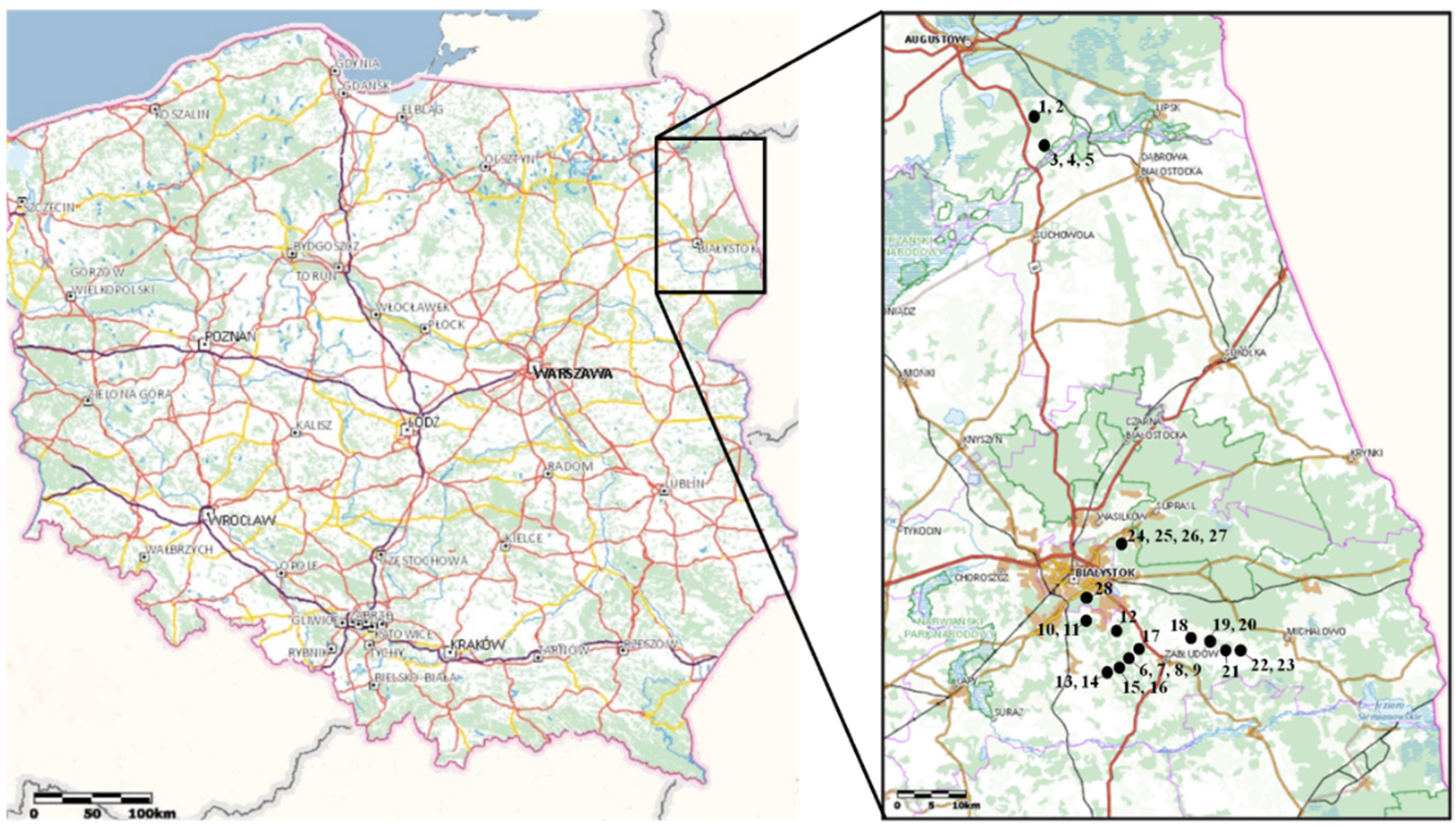
The sampling was conducted in 28 wells in north-eastern Poland in two regions (Figure 1), which are a representation of the glaciated part of the Central European lowlands. Wells 1–9 were sampled on the 2 October 2019, while wells 10–28 were sampled on the 15 July 2020. Wells 1–5 are located in the Biebrza River valley in the area of Vistulian Glaciation. Wells 6–28 are located in rural areas around Bialystok city (Figure 1) on the area of Warta Glaciation. Most of the studied dug wells were in good conditions but mainly unused presently. All of the wells have a cover that isolates them, and examples of the studied wells are shown in Figure 2.
Samples were collected using a 10 L bucket and then 50–100 L of water each time (Table 1) were filtered through a 50 µm plankton net and fixed with 96% alcohol. Copepods (Cyclopoida, Harpacticoida, Calanoida) were identified to species and all individuals in the samples were enumerated. Each species was dissected and the most important morphological features (thoracic legs; caudal rami, antennule, coxopodite of P4, etc.) are presented in Figure 3, Figure 4, Figure 5, Figure 6 and Figure 7. We selected few individuals from each species for the genetic analysis. Before DNA extraction, each specimen was photographed using micro imaging software for documentation of its body and distinguishing features. Photography documentation of each species together with its COI or 12S rRNA sequence was uploaded to the BOLD System. We uploaded our sequences also to GenBank. Reference materials are deposited in the Department of Hydrobiology, University of Bialystok as alcohol samples.
Copepoda individuals collected from different wells were stored in sterile plastic test tubes with 70% ethanol. The genomic DNA from individuals belonging to the four different species C. furcifer, D. bisetous, D. crassicaudis, and E. serrulatus were extracted with a DNeasy Blood & Tissue Kit (Qiagen, Hilden, Germany). Amplification of the fragment of the mitochondrial COI gene was performed for E. serrulatus (2 individuals from Zagruszany well), D. bisetosus (8 individuals from Zagruszany well), and D. crassicaudis (3 individuals from Folwarki Wielkie, 2 from Topolany, 1 from Folwarki Małe, and 1 from Żuki wells) using primers dgLCO1490 and dgHCO2198 [10], while for C. furcifer (7 individuals from Folwarki Tylwickie and 1 from Topolany wells) a fragment of 12S ribosomal RNA gene was amplified using primers 12SH13845 and 12SL13337 [11]. PCRs for each primer pair were carried out in 5 μL volumes, and the reaction mixtures consisted of 2 μL of extracted DNA as a template, 1.7 μL of Qiagen Multiplex PCR Master Mix (1x), 0.3 μL mix of primers, and 1 μL of Qiagen nuclease-free water. The thermocycling parameters were as follows: initial denaturation step at 95 °C for 15 min and 42 cycles with denaturation at 94 °C for 30 s, annealing at 48 °C (for COI gene) or at 57 °C (for 12S rRNA gene) for 90 s, extension at 72 °C for 60 s, and final elongation for 30 min at 60 °C. PCRs were performed in a Labcycler SensoQuest thermal cycler. Amplified PCR products were purified with EPPiC Fast (A&A Biotechnology) in an enzymatic reaction following the manufacturer’s protocol. They were then processed for cycle sequencing PCR with the BigDye Terminator v3.1 Cycle Sequencing Kit (Applied Biosystems, Foster City, CA, USA). Unincorporated dideoxynucleotides were eliminated from the sequencing reaction using the ExTerminator Kit (A&A Biotechnology). We carried out the detection of sequencing reaction products on an ABI PRISM 3100 Genetic Analyzer (Applied Biosystems).
Sequencing results of the mtDNA COI gene and 12S rRNA were aligned and revised manually using BioEdit [12]. Photography documentation of each species together with its fragments of the COI gene (E. serrulatus, D. bisetosus, D. crassicaudis) or 12S rRNA gene sequences (C. furcifer) were uploaded to the BOLD System. We also submitted to GenBank obtained in our study sequences of the mtDNA COI and 12S rRNA genes. To test the phylogenetic relationships among our newly obtained COI and 12S rRNA haplotypes and sequences downloaded from GenBank (Table S1), we constructed phylogenetic trees using a maximum likelihood (ML) algorithm in Mega v6.06 [13] using 1000 bootstrap replicates. The GTR+I+G model of substitution was selected as the best fitting model by the AIC test (Akaike Information Criterion) with the jModelTest [14] for the ML trees.
3. Results
The groundwater copepods were found in 16 of the 28 studied wells in north-eastern Poland (Table 1). The highest abundance was recorded in Cisów village, with 27.5 individuals per liter (Table 1). The average abundance of groundwater copepods was 2.5 ± 6.9 ind. L−1. We identified six Copepoda (five Cyclopoida, one Calanoida) and one Cladocera species in the groundwater of north-eastern Poland.
Most studied wells were dominated by one species, nevertheless, two species were found in six wells, and three species were found in two wells (Table 1). The most common species were: Eucyclops serrulatus (eight wells), Diacyclops bisetosus (seven wells), Diacyclops crassicaudis (five wells), and Cyclops furcifer (three wells). We observed abnormal morphology of the caudal rami in D. crassicaudis. Detailed descriptions of the above-mentioned species are presented below, as well as a description of abnormal structures in a single individual of D. crassicaudis.
Other species were single records and represented stygoxenes, which were temporarily imported from the surface. Among stygoxenes, one individual of Eudiaptomus gracilis and Thermocyclops crassus were found in the well in Białystok city (district of Dojlidy) (Figure S1). The analyzed individual of E. gracilis was a male with a flattened body (Figure S1A); the fourth leg (P4) and fifth leg (P5) are shown in Figure S1B,C. Pictures of the Thermocyclops crassus body, furcal branches, and exopodite P4 are presented in the Figure S1D–F. Moreover, the cladoceran Chydorus sphaericus was found in the Zagruszany well (Figure S1G–I) in the number of eleven individuals.
- Cyclops furcifer Claus, 1857
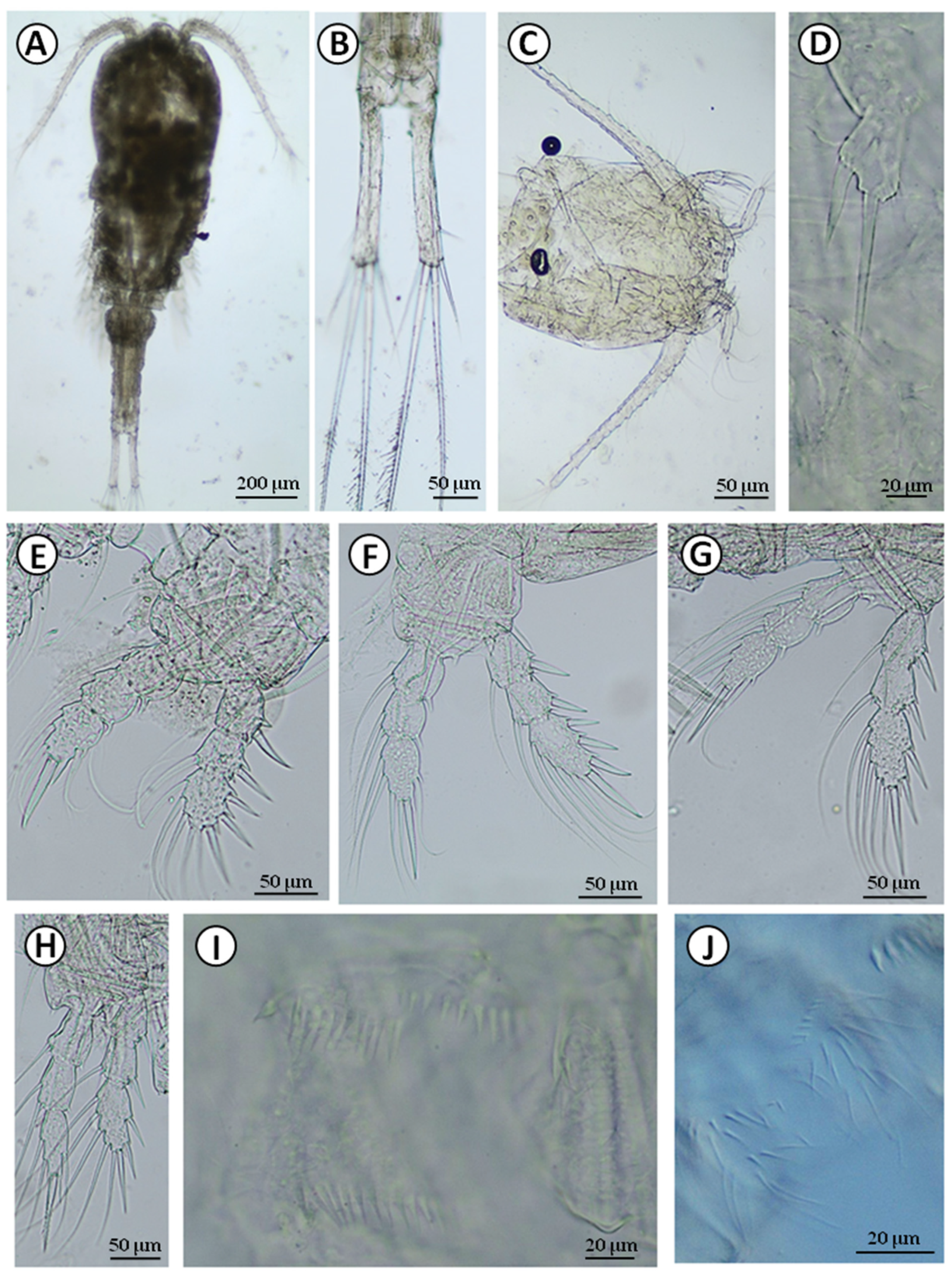
Cyclops furcifer was found in high abundance in the Folwarki Tylwickie village, and in low numbers in two wells in the Topolany village (Table 1). Females with egg sacs were also observed. The body size of the examined adult female was 1578 µm. The ends of the fourth and fifth thoracomer (Th4 and Th5) extended outwards forming ‘wings’ (Figure 3A). The furcal branches were long and slender with a proportion length to width of 6:1 (Figure 3B). The inner margin of furcal branches had hair setae. Terminal internal seta was shorter than the furcal branches (about 0.6) (Figure 3B). Terminal external seta was shorter than the terminal inner seta (Figure 3B). Antennule 17-segmented reaching the middle of Th2 (Figure 3C).
The spine formula of swimming legs was 3.4.3.3 (Figure 3E–H). The last exopodal article was bearing five setae in all swimming legs (Figure 3E–H). The last endopodal article of P4 was almost twice as long as wide, the inner apical spine was longer than the article (Figure 3H). The coupler of P4 had two/three lines of hair-like spinules (Figure 3I). The coupler of P3 was also pilose caudally, which distinguishes it from all the other Cyclops occurring in Poland, except for C. insignis, but this species had 14-segmented antennule and very thick P4 coxopodite seta. The ornamentation of P4 coxopodite is shown in Figure 3J. The P5 had two segments, and the medial spine of second segment was long, reaching far beyond the end of the segment (Figure 3D). At the end of the second segment of P5, there were rows of small spinules at insertion of medial and apical seta (Figure 3D). The spinules were present also on the first segment of P5 at insertion of the lateral seta.
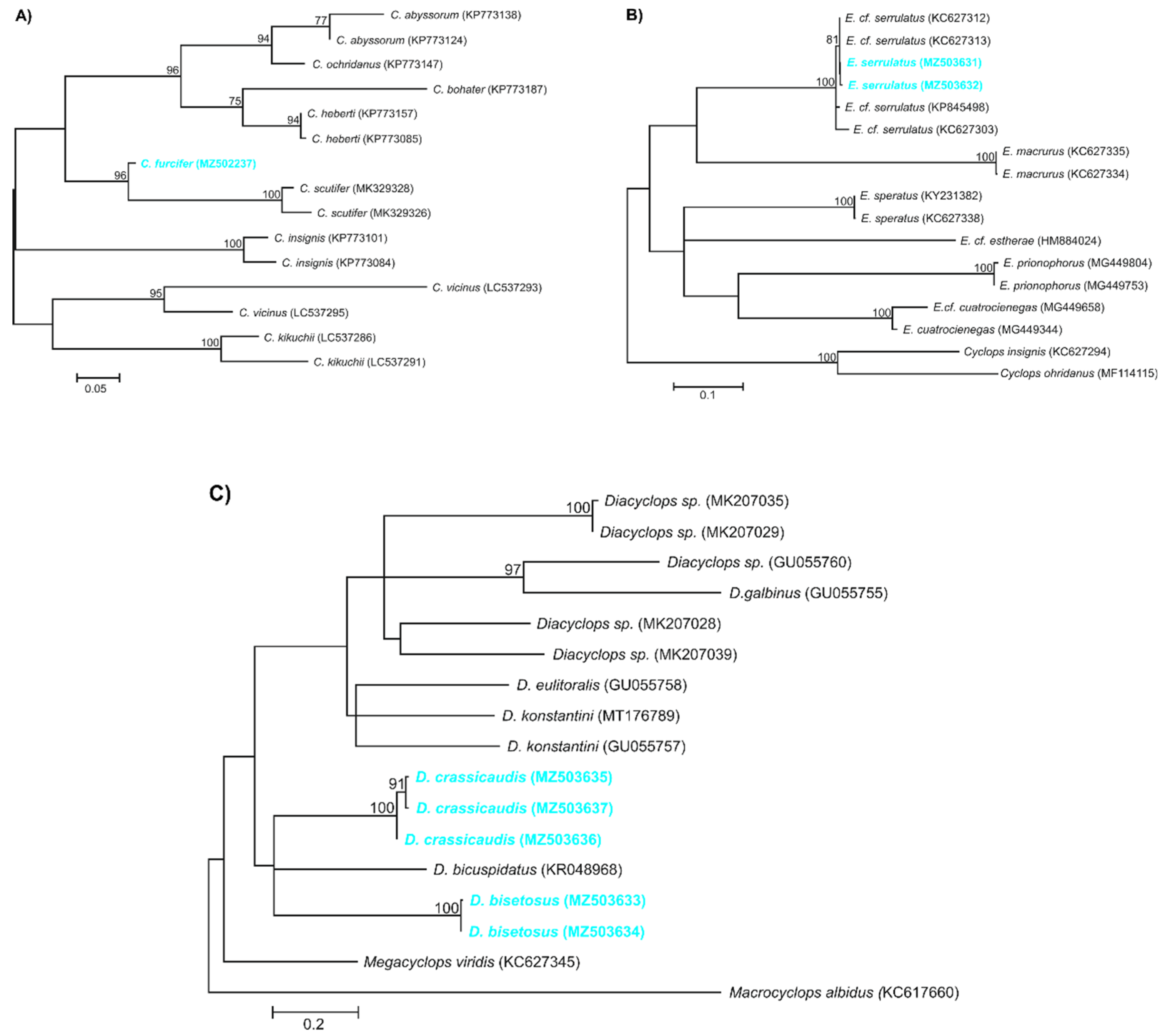
The molecular analysis of a 369-bp of a fragment of the 12S ribosomal RNA gene yielded among studied individuals of C. furcifer from the Folwarki Tylwickie (N = 7) and Topolany (N = 1) wells one new haplotype (GenBank accession no. MZ502237; BOLD sample ID: UwB_4_Cf4). The maximum likelihood phylogenetic reconstructions produced a strong topology (Figure 8A). The ML tree revealed that our 12S rRNA newly discovered haplotype of C. furcifer was grouped with haplotypes described for C. scutifer (MK329326 and MK329328) by Hołyńska and Wyngaard (2019) [15].
- Eucyclops serrulatus s. stricto (Fischer, 1851)
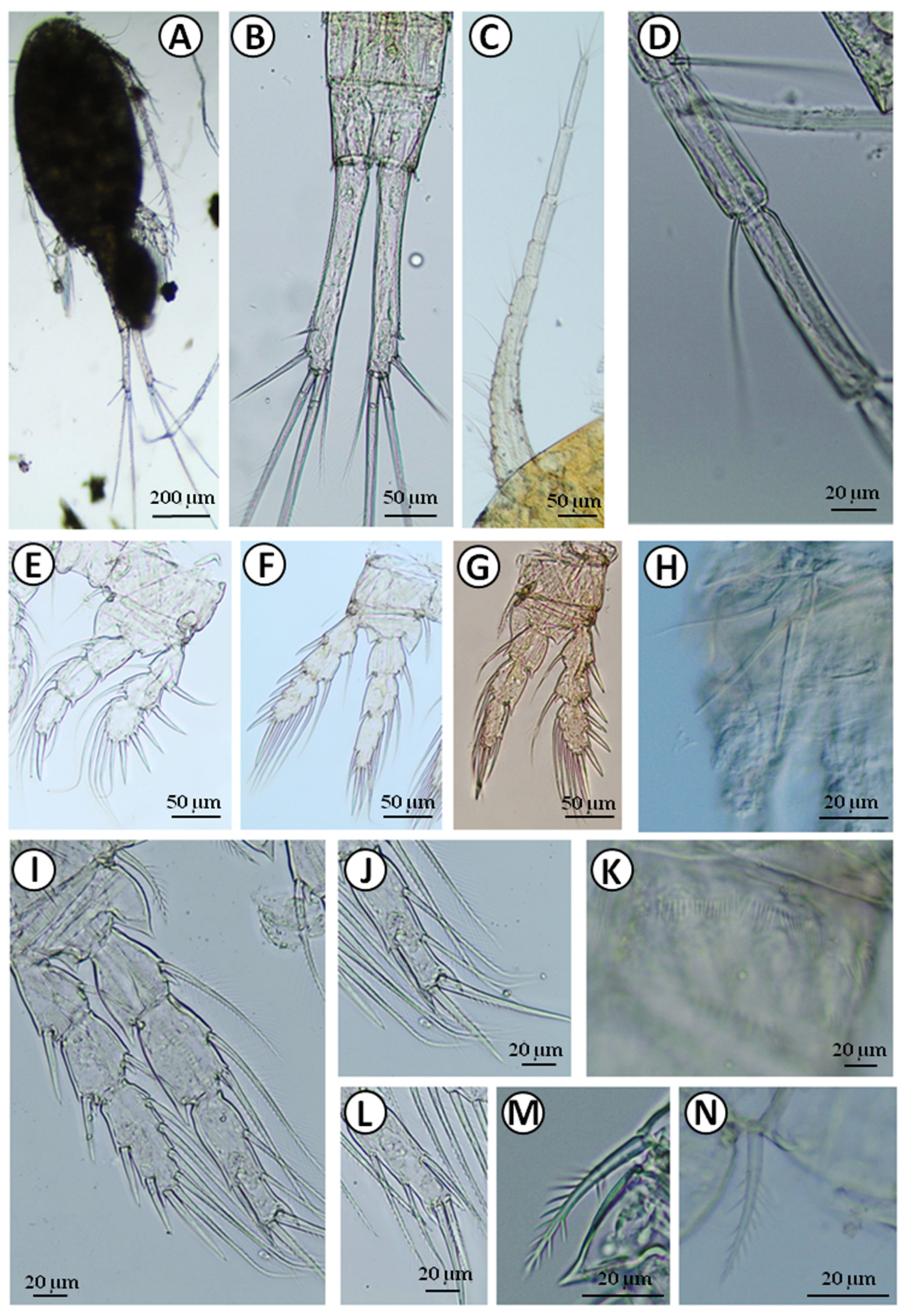
Eucyclops serrulatus was found in eight wells in villages of Cisów, Wolne, Pasynki, and Zagruszany (Table 1). The high abundance of this species was found in well no. 1 in the Cisów village (27.5 ind. L−1) and Pasynki village (Table 1). The adult specimens accounted for about 15–25% of all copepods (Table 1). The body size of the analyzed adult female was 1240 µm. The furcal branches in examined female were slender, six times as long as wide (Figure 4B), but we also observed adult females with shorter furcal branches. The outer margins of the furcal branches serrulated (Figure 4B). The ratio of the innermost caudal seta to the outermost caudal seta was 1.4 (Figure 4B). Antennule (A1) were 12-segmented (Figure 4C), long, and reaching almost the genital segment (Figure 4A), which is quite unusual for this species. The last three segments of A1 had a smooth hyaline membrane (Figure 4C).
P1–4 had 3-segmented exopod and endopod (Figure 4E–G,I). The first and second segments of exopodites of the swimming legs had one seta each, while the third segment had five setae. The spine formula was 3.4.4.3., respectively (Figure 4E–G,I). The last endopodal segment of P4 had two long apical spines, but we found a different arrangement of these spines (Figure 4J,L). The ornamentation of P4 coxopodite is shown in Figure 4K. The coxal spine of P4 had hair-like setules, but on the outer margin, there was a gap in the setulation (Figure 4M,N). P5 was one segmented, with a large knife-like inner spine and two setae of a similar length to the spine (Figure 4H).
The analysis of an mtDNA COI gene fragment of 631-bp long yielded among two individuals from the Zagruszany well two new haplotypes of E. serrulatus: haplotype H1 (GenBank accession no. MZ503631; BOLD sample ID: UwB_7_Es7) and haplotype H2 (GenBank accession no. MZ503632; BOLD sample ID: UwB_8_Es8), as defined by one polymorphic site being transition. The maximum likelihood phylogenetic reconstructions produced a strong topology (Figure 8B). The ML tree revealed that our two mtDNA COI haplotypes was grouped with haplotypes belonging to the E. serrulatus described for Ukraine, Russia (GenBank accession no. KC627312 and KC627313; unpublished).
Figure 3.
Cyclops furcifer: A—habitus; B—furcal branches; C—cephalothorax with antennule (A1) and antenna (A2); D—P5; E—P1; F—P2; G—P3; H—P4; I—coxopodite of P4; J—coupler of P4.
Figure 3.
Cyclops furcifer: A—habitus; B—furcal branches; C—cephalothorax with antennule (A1) and antenna (A2); D—P5; E—P1; F—P2; G—P3; H—P4; I—coxopodite of P4; J—coupler of P4.

Figure 4.
Eucyclops serrulatus: A—body; B—furcal branches; C—antennule; D—hyaline membrane on the 10th and 11th segments of the antennule; E—P1; F—P2; G—P3; H—P5; I—P4; J, L—last segment of P4 endopodite; K—P4 coxopodite; M, N—coxal spine of P4.
Figure 4.
Eucyclops serrulatus: A—body; B—furcal branches; C—antennule; D—hyaline membrane on the 10th and 11th segments of the antennule; E—P1; F—P2; G—P3; H—P5; I—P4; J, L—last segment of P4 endopodite; K—P4 coxopodite; M, N—coxal spine of P4.

- Diacyclops bisetosus (Rehberg, 1880)
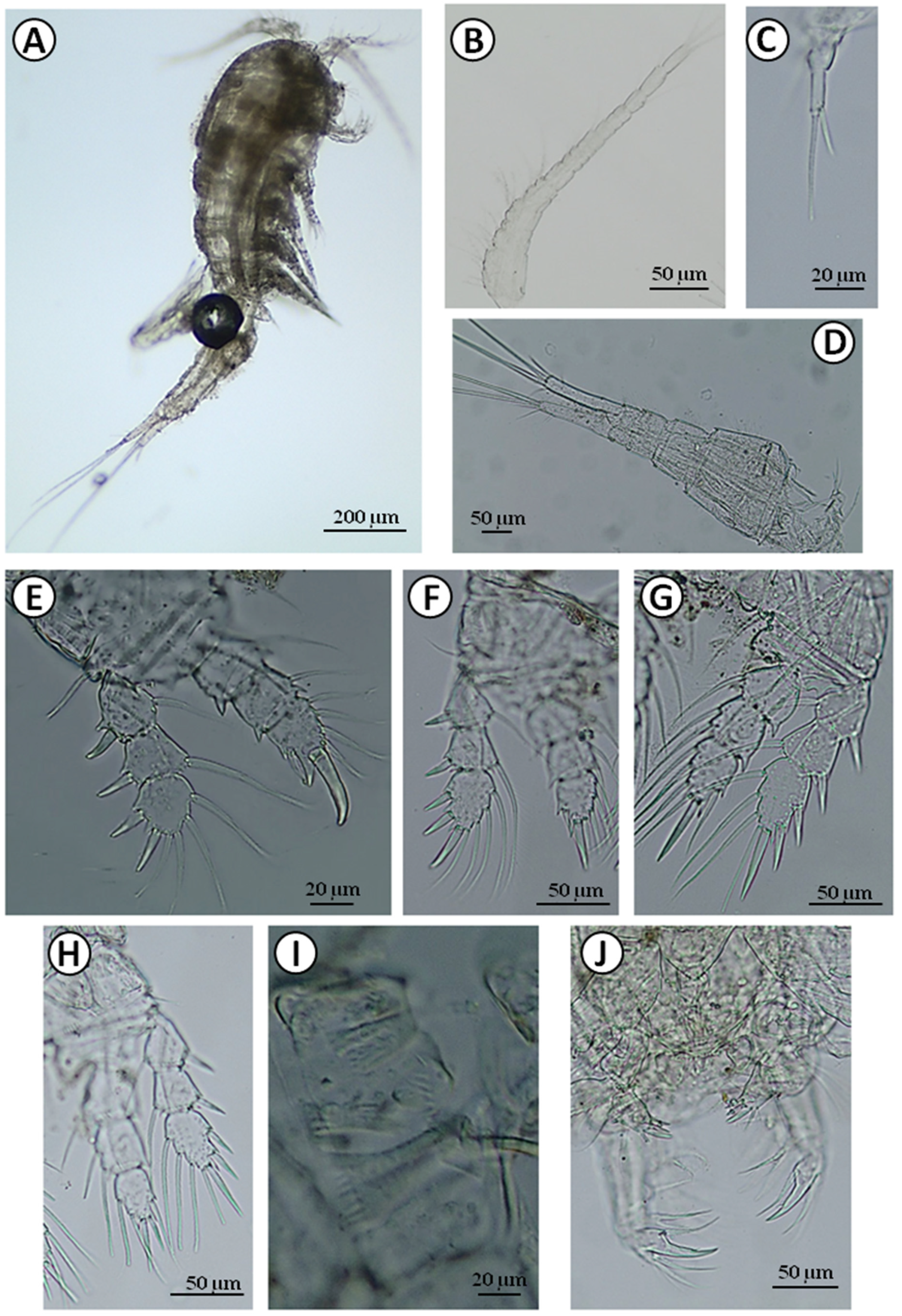
Diacyclops bisetosus was found in low numbers in seven wells in the villages of Wolne, Pasynki, Zagruszany, Topolany (Table 1). The body size of the analyzed adult female was 1074 µm. Antennule were 17-segmented (Figure 5B), relatively short reaching the end of the cephalothorax (Figure 5A). Furcal branches were four times as long as wide in the female measured (Figure 5D), however, we also observed longer furcal branches with a proportion of 7:1 in this species. The innermost and outermost caudal setae were subequal. The swimming legs were three-segmented with a spine formula of 2.3.3.3 (Figure 5E–H). The last exopodal article was bearing four setae in all swimming legs (Figure 5E–H). One of the distinguishing features was the length proportion of the apical spines on the third endopodal segment of P4, where the inner spine was longer than the outer one (Figure 5H). Coxopodite of P4 had ornamentation (Figure 5I). P5 consists of two segments, segment two had a long and slim inner spine (Figure 5C).
The analysis of an mtDNA COI gene fragment of 578-bp long yielded among eight individuals from the Zagruszany well two new haplotypes of D. bisetosus: haplotype H1 (GenBank accession no. MZ503633; BOLD sample ID: UwB_5_Db5) and haplotype H2 (GenBank accession no. MZ503634; BOLD sample ID: UwB_6_Db6), as defined by one polymorphic site being in transition. Haplotype H1 was detected in 5/8 (62.5%) of the studied individuals, while haplotype H2 was in 3/8 (37.5%). The maximum likelihood phylogenetic reconstructions produced a strong topology (Figure 8C). The ML tree revealed that our two mtDNA COI haplotypes of D. bisetosus created a separated branch among different species of Diacyclops genus, which also grouped haplotype of D. bicuspidatus (GenBank accession no. KR048968) described by Baek et al. (2016) [16] and also our newly described from Poland haplotypes of D. crassicaudis (GenBank accession no. MZ503635–MZ503637).
- Diacyclops crassicaudis (Sars G.O., 1863)
Diacyclops crassicaudis was found in four wells in the villages of Żuki, Folwarki Wielkie, Folwarki Tylwickie, Topolany (Table 1). The species occurred in high abundance in well no. 21 in Folwarki Tylwickie (6.6 ind. L−1), where the adult specimens accounted for about 20%. In Folwarki Wielkie and Topolany, adult specimens dominated over copepodites (Table 1), and females with egg sacs were present. The body length was 1100 µm in the adult female measured, and the two egg sacs had nine eggs each (Figure 6A). Thoracic segments 2, 3, and 4 were rounded and bulging (Figure 6A). Antennule were 12-segmented (Figure 6B), relatively short and reaching the end of the cephalothorax (Figure 6A). The caudal rami were five times as long as wide, with a wide gap between them, especially at the posterior end of the rami (Figure 6A). The spine formula of swimming legs was 2.3.3.3 (Figure 6C–F). P5 consisted of two segments, with quite a large and massive inner spine (Figure 6G).
The analysis of a 525-bp mtDNA COI gene fragment amplified from studied individuals from four different wells (three from Folwarki Wielkie, two from Topolany, one from Folwarki Małe and Żuki) yielded three new haplotypes of D. crassicaudis: haplotype H1 (GenBank accession no. MZ503635; BOLD sample ID: UwB_1_Dc1), haplotype H2 (GenBank accession no. MZ503636; BOLD sample ID: UwB_2_Dc2), and haplotype H3 (GenBank accession no. MZ503637; BOLD sample ID: UwB_3_Dc3). All individuals from the Folwarki Wielkie well possessed haplotype H1 (3/7, 42.9%), which differed from haplotypes H2 and H3 by 12 polymorphic sites (11 transitions, 1 transversion). Haplotype H2 was found in individulas from Folwarki Małe i Żuki (2/7, 28.6%), and differed from haplotype H3 discovered in individuals from Topolany (2/7, 28.6%) by six transitions. The maximum likelihood phylogenetic reconstructions produced a strong topology (Figure 8C). The ML tree revealed that our three COI haplotypes of D. crassicaudis created a separated branch among different species of Diacyclops genus which also grouped haplotype of D. bicuspidatus (GenBank accession no. KR048968) described by Baek et al. (2016) [16] and also our newly described from Poland haplotypes of D. bisetosus (GenBank accession no. MZ503633 and MZ503634).
We noticed developmental abnormality in the caudal rami morphology in one specimen of D. crassicaudis (Figure 7) in well no. 18 in the Folwarki Małe village. The rami were reduced in length and the parts anterior to the insertion of the lateral caudal setae were fused (Figure 7B). The outermost terminal caudal seta was abnormally long (Figure 7A), and other setae showed a slightly different size (Figure 7B). Whereas other parts of the body were properly developed (Figure 7A,G), and the legs had a spine and setae formula typical of the species (Figure 7B–F).
Figure 5.
Diacyclops bisetosus: A—body; B—antennule; C—P5; D—furcal branches; E—P1; F—P2; G—P3; H—P4; I—coxopodite of P4; J—mouthparts.
Figure 5.
Diacyclops bisetosus: A—body; B—antennule; C—P5; D—furcal branches; E—P1; F—P2; G—P3; H—P4; I—coxopodite of P4; J—mouthparts.

Figure 6.
Diacyclops crassicaudis: A—body; B—antennule and antenna; C—P1; D—P2; E—P3; F—P4; G—P5.

Figure 7.
Diacyclops crassicaudis with aberrant development of caudal rami: A—body; B—aberrant development of furcal branches; C—P1; D—P2; E—P3; F—P4; G—P5; H—antennule.
Figure 7.
Diacyclops crassicaudis with aberrant development of caudal rami: A—body; B—aberrant development of furcal branches; C—P1; D—P2; E—P3; F—P4; G—P5; H—antennule.

Figure 8.
Maximum likelihood topology computed with the GTR+I+G model of substitution evolution, representing the phylogenetic relationships among the sequences of the 12S rRNA gene found in Cyclops furcifer (A), and the COI gene found in Eucyclops serrulatus (B), Diacyclops bisetosus and Diacyclops crassicaudis (C). Numbers listed at the nodes represent the percent support for the node from 1000 bootstrap replicates. The haplotypes found in this study are marked in blue and bolded.
Figure 8.
Maximum likelihood topology computed with the GTR+I+G model of substitution evolution, representing the phylogenetic relationships among the sequences of the 12S rRNA gene found in Cyclops furcifer (A), and the COI gene found in Eucyclops serrulatus (B), Diacyclops bisetosus and Diacyclops crassicaudis (C). Numbers listed at the nodes represent the percent support for the node from 1000 bootstrap replicates. The haplotypes found in this study are marked in blue and bolded.

4. Discussion
Our study presents the first data on the groundwater copepods of the Polish lowland. The middle European lowlands were an important center of post-glacial dispersion of the freshwater zooplankton [17]. Furthermore, Quaternary glaciations may have played an important role in the extinction of the surface species and increasing colonization and speciation in groundwater [3]. Currently, some phylogeographic studies have been published on the groundwater copepods in South and Eastern Europe [4], but none of them included the glaciated part of Central Europe. Generally, the groundwater fauna of the mountain region of Europe is much better known than in European lowlands, which seems to be a white spot on the map.
The results of our study indicated the presence of six Copopoda species and one Cladocera in the groundwater of north-eastern Poland. We classified three of these species as stygoxenes, and four as stygophiles. Among stygoxenes were Chydorus sphaericus, Eudiaptomus gracilis, and Thermocyclops crassus. These species were present as a single record or in low numbers in groundwater, but they are very common in different types of surface water bodies in this region [18,19], north-western Poland [20], and Europe [21,22]. Therefore, these species seem to be accidental taxa temporarily imported from the surface. However, E. gracilis has a flattened body which may indicate adaptation for an interstitial environment where the major constraint is the reduced living space [3,23]. We have classified four species as stygophiles—Eucyclops serrulatus, Diacyclops bisetosus, Diacyclops crassicaudis, and Cyclops furcifer. These species show an ecological preference for the groundwater habitats, and a higher frequency and occurrence in groundwater than in surface habitat. However, sometimes it is difficult to define the real ecology of the copepods found in groundwater [3].
Eucyclops serrulatus was found in eight wells from different regions of north-eastern Poland, often with high abundance to even 27.5 ind. L−1. This species is also a very common littoral copepod in Europe but are usually found in low numbers [21,22,24]. E. serrulatus is primarily benthic, swimming near the sediment layers of lakes, river, streams, and springs. It is commonly found in epikarst, alluvial aquifers, and hyporheic zones where it can live from a few centimeters below the streambed up to 1.5 m deep [25,26,27], as a member of the permanent hyporheos [28]. E. serrulatus is frequently the most abundant epigean cyclopoid in meiofaunal communities [21,29,30]. E. serrulatus is a morphological and ecologically variable species and was recently redescribed [31] and further serrulatus-group was redefined [32]. This group includes 17 species and subspecies, which differ from each other in the presence of microcharacters of the antennary basipodite and the P4 coxopodite [33]. Our species was E. serrulatus s. stricto, with three distalmost segments of antennule with a narrow hyaline membrane of regular shape (Figure 4D), and a very characteristic spine of coxopodite of P4 with a gap among the strong teeth-like setules on the outer side (Figure 4N). Furthermore, we have found a difference between populations in the arrangement of spines of the P4 endopodite (Figure 4J,L). Recently, 36 European populations of E. serrulatus were deeply studied molecularly, which confirms its species complex with eight divergent lineages [34], awaiting the formal description. Further study revealed genetic and morphological heterogeneity within E. serrulatus in Europe and Asia [35]. Our molecular analysis of an mtDNA COI gene fragment discovered two new haplotypes of E. serrulatus which grouped with haplotypes described from Ukraine and Russia in the strongly supported phylogenetics branch on the ML tree.
The genus Diacyclops, comprising a large number of stygobiontic species [36,37], was represented by two species in our samples; both were classified as stygophiles. Diacyclops bisetosus was found in seven wells in different regions, whereas in surface water bodies in Poland it is found occasionally in low numbers [19,20]. However, D. bisetosus is a cosmopolitan, but probably cryptic species widely distributed in Europe [22]. This species was frequently found in habitats that could be considered as an ecotone between surface water and groundwater such as semi-permanent ponds, rivers, springs, puddles, marshes, ditches, caves, and wells [22,38]. D. bisetosus was common in groundwater and caves of the Alps [39], while in the Polish mountains another similar species D. bicuspidatus was frequently reported in the groundwater [6]. Gurney (1933) found it even in a tree-hole in England [40]. D. bisetosus was also the most frequent species in the hypersaline samples from the Crimean Peninsula [41]. The international genetic databases (GenBank and BOLD System) do not have sequences for D. bisetosus; therefore, we present a detailed morphological description of this species with its COI gene sequences.
Diacyclops crassicaudis was found in five wells in the rural areas, east of Białystok city, often with high abundance. Females with egg sacs were frequently reported. This species is considered as cold-stenothermic [39,42] and it was reported from Europe from temporary ponds, puddles after rains and snowmelt, and littoral or psammon habitats of small lakes [22]. D. crassicaudis was also frequently reported from groundwater in Italy, where large groundwater endemism was found and seven subspecies were described in the ‘crassicaudis’ complex [43]. The different environmental conditions in groundwater favor radiation which leads to the formation of the cryptic species [3]. The ‘crassicaudis’ species complex was also frequently reported from groundwater in Central Spain [44], Germany [45,46], Austria [39], USA [47,48,49], Japan [50], and Australia [51]. It also occurs in the groundwater of the mountain regions of Poland [6], while it was rarely recorded in the surface water bodies in Poland. Therefore, we consider it as a stygophiles. The international genetic databases (GenBank and BOLD System) do not have sequences for D. crassicaudis; therefore, we present a detailed morphological description of this species with its COI gene sequences.
We have observed the development of abnormal structures in one specimen of D. crassicaudis, where the anterior part of furcal rami were fused together to form a single plate. Similar aberrancies were observed by Gurney (1933) in Cyclops vicinus and Eucyclops serrulatus [40]. We have also noticed differences in the caudal setae morphology, where the outermost terminal caudal seta and dorsal caudal seta were abnormal long. Both specimens described by Gurney (1933) have one large dorsal seta instead of two dorsal setae. In our case, the furcal rami were not completely fused and there are two long dorsal setae. Abnormal structures, such as supernumerary setae, or deformed setae or spine, are not very uncommon but do not seem to have any significance [40]. We have observed abnormal development of whole furcal rami with setae, which could significantly affect the functioning of these organs. The abnormalities in copepod morphology are most often related to the tumor-like anomalies [52].
Cyclops furcifer was found in three wells in rural areas east of the Białystok city. In one well it appeared in large numbers, and females with egg sacs were observed. This species is reported from Europe in small and shallow lakes, temporary waters, swamps [22,39,53], and also was found in the groundwater of the Channel Islands just off the coast of Normandy, France [54]. Our phylogenetics analysis based on 12S rRNA gene sequences of different species of Cyclops genus confirmed topology obtained by Krajíček et al. (2016) which was based on the 2531-bp long alignment consisting of fragments of six different studied molecular markers [55]. According to this topology, C. furcifer belong together with C. scutifer to the separate phylogenetics group, named by Krajíček et al. (2016) Clade 2 [55]. However, Hołyńska and Wyngaard (2019) divided kolensis-clade (C. kolensis, C. furcifer, C. kikuchii, C. vicinus, C. insignis, C. alaskaensis) from scutifer-clade (C. scutifer, C. jashnovi, C. columbianus) based on morphology and phylogenetics, along with the geographic distribution patterns and ecological traits [15].
Interestingly, we have not found any Harpacticoida, which are otherwise common in groundwater. A previous preliminary study of a few wells in this region revealed the presence of a harpacticoid Elaphoidella elaphoides (Chappuis, 1923), which was a species new to the Polish fauna [7]. Harpacticoida dominates the benthos and are rarely found in groundwater plankton. They can be either benthic or, more frequently, interstitial, irrespective of grain size composition. Gravel and sand are the preferred substrate for most harpacticoid species, whereas poorly sorted or clogged sediments show a drastic decrease in species diversity [3]. While Cyclopoida can live either in the plankton of subterranean lakes and pools or occur in benthic or interstitial habitats where they prefer sediment of medium-coarse grain size [3]. Therefore, the lack of harpacticoids could be a result of the sampling method here applied, allowing the collection of mainly open-water species.
5. Conclusions
Our study is the first report on the groundwater copepods of the Polish lowland. Six copepod and one cladoceran species have been identified in the groundwater of north-eastern Poland. We have classified four species as stygophiles—Eucyclops serrulatus, Diacyclops bisetosus, Diacyclops crassicaudis, and Cyclops furcifer. These species are frequently found in wells in different regions of north-eastern Poland, often in high numbers, and with females with egg sacs. In the surface water bodies of Poland, these species were rarely recorded, which may indicate an ecological preference for the groundwater habitats. We have presented detailed morphological diagnoses of these species, accompanied by molecular characteristics (COI gene or 12S rRNA gene). Developmental aberrancy was observed in one specimen of D. crassicaudis, in which the anterior part of the furcal rami was fused together forming a single plate.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/w13152086/s1, Figure S1: photography of species considered as stygoxenes, Eudiaptomus gracilis, Thermocyclops crassus, and Chydorus sphaericus; Table S1: list of species and GenBank accession numbers of their gene sequences used in the phylogenetic analysis based on the maximum likelihood trees (Figure 8A–C).
Author Contributions
M.K.: writing—original draft preparation, conceptualization, methodology, investigation, formal analysis, validation, visualization, and supervision. S.S.: investigation, visualization, writing—review, and editing. M.Ś.: investigation, methodology, visualization, writing—review, and editing. J.M.: investigation. All authors have read and agreed to the published version of the manuscript.
Funding
This article/publication has received financial support from the Polish Ministry of Science and Higher Education under subsidy for maintaining the research potential of the Faculty of Biology, University of Bialystok.
Data Availability Statement
The original contributions presented in the study are included in the article. Further inquiries can be directed to the corresponding authors.
Acknowledgments
The authors are thankful to Maria Hołyńska for consultation on identified species, valuable comments about the development of abnormal structures in copepods, and her preliminary review which significantly improve our manuscript. We are grateful to Jolanta Ejsmont-Karabin for the final language correction, and to Piotr Rode for the help in preparing Figure 8.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Mammola, S.; Martinez, A. Let research on subterranean habitats resonate! Subterr. Biol. 2020, 36, 63–71. [Google Scholar] [CrossRef]
- Brancelj, A.; Dumont, H. A review of the diversity, adaptations and groundwater colonization pathways in Cladocera and Calanoida (Crustacea), two rare and contrasting groups of stygobionts. Fundam. Appl. Limnol. 2007, 168, 3–17. [Google Scholar] [CrossRef]
- Galassi, D.M.P. Groundwater copepods: Diversity patterns over ecological and evolutionary scales. Hydrobiologia. 2001, 453, 227–253. [Google Scholar] [CrossRef]
- Iannella, M.; Fiasca, B.; Di Lorenzo, T.; Biondi, M.; Di Cicco, M.; Galassi, D.M.P. Spatial distribution of stygobitic crustacean harpacticoids at the boundaries of groundwater habitat types in Europe. Sci. Rep. 2020, 10, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Karpowicz, M.; Pociecha, A.; Hołyńska, M.; Dumnicka, E.; Galas, J. Groundwater copepods from Carpathian flysch in south Poland. In Proceedings of the 3rd Polish Zooplankton Conference, Szczecinek, Poland, 20–22 July 2018. [Google Scholar] [CrossRef]
- Kur, J.; Mioduchowska, M.; Kilikowska, A. Distribution of cyclopoid copepods in different subterranean habitats (southern Poland). Oceanol. Hydrobiol. Stud. 2020, 49, 255–266. [Google Scholar] [CrossRef]
- Karpowicz, M. New data to the knowledge on the Harpacticoida (Crustacea, Copepoda) fauna in Poland. Fragm. Faunist. 2016, 59, 87–98. [Google Scholar] [CrossRef]
- Agnarsson, I.; Kuntner, M. Taxonomy in a changing world: Seeking solutions for a science in crisis. Syst. Biol. 2007, 56, 531–539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bucklin, A.; Lindeque, P.K.; Rodriguez-Ezpeleta, N.; Albaina, A.; Lehtiniemi, M. Metabarcoding of marine zooplankton: Prospects, progress and pitfalls. J. Plankton Res. 2016, 38, 393–400. [Google Scholar] [CrossRef] [Green Version]
- Meyer, C.P. Molecular systematics of cowries (Gastropoda: Cypraeidae) and diversification patterns in the tropics. Biol. J. Linn. Soc. Lond. 2003, 79, 401–459. [Google Scholar] [CrossRef] [Green Version]
- Machida, R.J.; Miya, M.U.; Nishida, M.; Nishida, S. Complete mitochondrial DNA sequence of Tigriopus japonicus (Crustacea: Copepoda). Mar. Biotechnol. 2002, 4, 406–417. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. 1999, 41, 95–98. [Google Scholar]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Posada, D. jModelTest: Phylogenetic model averaging. Mol. Biol. Evol. 2008, 25, 1253–1256. [Google Scholar] [CrossRef] [PubMed]
- Hołyńska, M.; Wyngaard, G.A. Towards a phylogeny of Cyclops (Copepoda): (in)congruences among morphology, molecules and zoogeography. Zool Scr. 2019, 48, 376–398. [Google Scholar] [CrossRef]
- Baek, S.Y.; Jang, K.H.; Choi, E.H.; Ryu, S.H.; Kim, S.K.; Lee, J.H.; Lim, Y.J.; Lee, J.; Jun, J.; Kwak, M.; et al. DNA barcoding of metazoan zooplankton copepods from South Korea. PLoS ONE 2016, 11, e0157307. [Google Scholar] [CrossRef] [Green Version]
- Hołyńska, M. On the morphology and geographical distribution of some problematic South Palearctic Cyclops (Copepoda: Cyclopidae). J. Nat. Hist. 2008, 42, 2011–2039. [Google Scholar] [CrossRef]
- Ejsmont-Karabin, J.; Kalinowska, K.; Karpowicz, M. Structure of ciliate, rotifer, and crustacean communities in lake systems of Northeastern Poland. In Polish River Basins and Lakes—Part II; Springer: Cham, Switzerland, 2020; Volume 87, pp. 77–101. [Google Scholar] [CrossRef]
- Karpowicz, M.; Ejsmont-Karabin, J. Diversity and Structure of Pelagic Zooplankton (Crustacea, Rotifera) in NE Poland. Water 2021, 13, 456. [Google Scholar] [CrossRef]
- Sługocki, Ł.; Czerniawski, R. Trophic state (TSISD) and mixing type significantly influence pelagic zooplankton biodiversity in temperate lakes (NW Poland). PeerJ. 2018, 6, e5731. [Google Scholar] [CrossRef] [Green Version]
- Walseng, B.; Hessen, D.O.; Halvorsen, G.; Schartau, A.K. Major contribution from littoral crustaceans to zooplankton species richness in lakes. Limnol. Oceanogr. 2006, 51, 2600–2606. [Google Scholar] [CrossRef]
- Błędzki, L.A.; Rybak, J.I. Freshwater Crustacean Zooplankton of Europe; Springer: Cham, Switzerland, 2016; pp. 1–918. [Google Scholar]
- Schminke, H.K. Perspectives in the study of the zoogeography of interstitial Crustacea: Bathynellacea (Syncarida) and Parastenocarididae (Copepoda). Int. J. Speleol. 1981, 11, 83–89. [Google Scholar] [CrossRef] [Green Version]
- Karpowicz, M. Biodiversity of microcrustaceans (Cladocera, Copepoda) in a lowland river ecosystem. J. Limnol. 2017, 76, 15–22. [Google Scholar] [CrossRef] [Green Version]
- Galassi, D.M.P.; Stoch, F.; Fiasca, B.; Di Lorenzo, T.; Gattone, E. Groundwater biodiversity patterns in the Lessinian Massif of northern Italy. Freshw. Biol. 2009, 54, 830–847. [Google Scholar] [CrossRef]
- Galassi, D.M.P.; Huys, R.; Reid, J.W. Diversity, ecology and evolution of groundwater copepods. Freshw. Biol. 2009, 54, 691–708. [Google Scholar] [CrossRef] [Green Version]
- Galassi, D.M.P.; Lombardo, P.; Fiasca, B.; Di Cioccio, A.; Di Lorenzo, T.; Petitta, M.; Di Carlo, P. Earthquakes trigger the loss of groundwater biodiversity. Sci. Rep. 2014, 4, 6273. [Google Scholar] [CrossRef]
- Gibert, J.; Stanford, J.A.; Dole-Olivier, M.-J.; Ward, J.V. Basic attributes of groundwater ecosystems and prospects for research. In Groundwater Ecology; Gibert, J., Danielopol, D.L., Stanford, J.A., Eds.; Academic Press, Inc.: San Diego, CA, USA, 1994; pp. 7–40. [Google Scholar]
- Fiasca, B.; Stoch, F.; Olivier, M.-J.; Maazouzi, C.; Petitta, M.; Di Cioccio, A.; Galassi, D.M.P. The dark side of springs: What drives small-scale spatial patterns of subsurface meiofaunal assemblages? J. Limnol. 2014, 73, 71–80. [Google Scholar] [CrossRef]
- Di Lorenzo, T.; Di Marzio, W.D.; Cifoni, M.; Fiasca, B.; Baratti, M.; Sáenz, M.E.; Galassi, D.M.P. Temperature effect on the sensitivity of the copepod Eucyclops serrulatus (Crustacea, Copepoda, Cyclopoida) to agricultural pollutants in the hyporheic zone. Curr. Zool. 2015, 61, 629–640. [Google Scholar] [CrossRef]
- Alekseev, V.; Dumont, H.J.; Pensaert, J.; Baribwegure, D.; Vanfleteren, J.R. A redescription of Eucyclops serrulatus (Fischer, 1851) (Crustacea: Copepoda: Cyclopoida) and some related taxa, with a phylogeny of the E. serrulatus-group. Zool. Scr. 2006, 35, 123–147. [Google Scholar] [CrossRef]
- Alekseev, V.R.; Defaye, D. Taxonomic differentiation and world geographical distribution of the Eucyclops serrulatus group (Copepoda, Cyclopidae, Eucyclopinae). In Studies on Freshwater Copepoda: A Volume in Honour of Bernard Dussart. Crustaceana Monographs; Defaye, D., von Vaupel Klein, J.C., Suárez-Morales, E., Eds.; Koninklijke Brill NV: Leiden, The Netherlands, 2011; Volume 16, pp. 41–72. [Google Scholar] [CrossRef]
- Alekseev, V.R. Revision of the genus Eucyclops (Claus, 1893) and subfamily Eucyclopinae of the world fauna. Arthropoda Sel. 2019, 28, 490–514. [Google Scholar] [CrossRef]
- Hamrova, E.; Krajicek, M.; Karanovic, T.; Cerny, M.; Petrusek, A. Congruent patterns of lineage diversity in two species complexes of planktonic crustaceans, Daphnia longispina (Cladocera) and Eucyclops serrulatus (Copepoda), in East European mountain lakes. Zool. J. Linn. Soc. 2012, 166, 754–767. [Google Scholar] [CrossRef] [Green Version]
- Sukhikh, N.; Alekseev, V. Genetic and morphological heterogeneity within Eucyclops serrulatus (Fischer, 1851) (Crustacea: Copepoda: Cyclopidae). J. Nat. Hist. 2015, 49, 2929–2953. [Google Scholar] [CrossRef]
- Pesce, G.L.; Galassi, D.P. New or rare species of Diacyclops Kiefer, 1927 (Copepoda, Cyclopoida) from different groundwater habitats in Italy. Hydrobiologia. 1987, 148, 103–114. [Google Scholar] [CrossRef]
- Pipan, T. Epikarst—A Promising Habitat: Copepod Fauna, Its Diversity and Ecology: A Case Study from Slovenia (Europe); Karst Research Institute at ZRC SAZU, ZRC Publishing: Ljubljana, Slovenia, 2005; pp. 1–101. [Google Scholar]
- Kur, J.; Wojtasik, B. Widłonogi Cyclopoida wybranych jezior zlewni górnej Raduni. In Jeziora Kaszubskiego Parku Krajobrazowego; Borowiak, D., Ed.; Uniwersytet Gdański: Gdańsk, Poland, 2007; pp. 145–163. (In Polish) [Google Scholar]
- Gaviria, S. Checklist and distribution of the free-living copepods (Arthropoda: Crustacea) from Austria. Ann. Naturhist. Mus. Wien. 1998, 100B, 539–594. [Google Scholar]
- Gurney, R. British freshwater Copepoda; The Ray Society: London, UK, 1933; pp. 76–78. [Google Scholar]
- Anufriieva, E.; Hołyńska, M.; Shadrin, N. Current Invasions of Asian Cyclopid Species (Copepoda: Cyclopidae) in Crimea, with Taxonomical and Zoogeographical Remarks on the Hypersaline and Freshwater Fauna. Annal Zool. 2014, 64, 109–130. [Google Scholar] [CrossRef] [Green Version]
- Lewis, J.J.; Reid, J.W. Patterns and Processes of Groundwater Invasion by Copepods in the Interior Low Plateaus of the United States. Acta Carsologica 2007, 36, 279–289. [Google Scholar] [CrossRef]
- Pesce, G.L.; Galassi, D.M.P. Copepodi di acque sotterranee della Sicilia. Animalia 1987, 14, 193–235. [Google Scholar]
- Iepure, S.; Rasines-Ladero, R.; Meffe, R.; Carreno, F.; Mostaza, D.; Sundberg, A.; Di Lorenzo, T.; Barroso, J.L. The role of groundwater crustaceans in disentangling aquifer type features—A case study of the Upper Tagus Basin, central Spain. Ecohydrology 2017, 10, e1876. [Google Scholar] [CrossRef]
- Hahn, H.J.; Fuschs, A. Distribution patterns of groundwater communities across aquifer types in south-western Germany. Freshw. Biol. 2009, 54, 848–860. [Google Scholar] [CrossRef]
- Gutjahr, S.; Schmidt, S.; Hahn, H. A proposal for a groundwater habitat classification at local scale. Subterr. Biol. 2014, 14, 25–49. [Google Scholar] [CrossRef] [Green Version]
- Yeatman, H.C. Rare cyclopoid copepods from wells in North Carolina. J. Elisha Mitchell Sci. Soc. 1943, 59, 27–36. [Google Scholar]
- Reid, J.W. Redescription of Diacyclops nearcticus (Kiefer, 1934) and description of four similar new congeners from North America, with comments on D. crassicaudis (G.O. Sars, 1863) and D. crassicaudis var. brachycercus (Kiefer, 1927) (Crustacea: Copepoda). Canad. J. Zool. 1992, 70, 1445–1469. [Google Scholar] [CrossRef]
- Bruno, M.; Cunningham, K.; Perry, S. Copepod Communities from Surface and Ground Waters in the Everglades, South Florida. Southeast. Nat. 2003, 2, 523–546. [Google Scholar] [CrossRef]
- Ito, T. Groundwater Copepods from South-western Japan. Hydrobiologia 1957, 11, 1. [Google Scholar] [CrossRef]
- Pesce, G.; De Laurentiis, P. Copepods from Ground Waters of Western Australia. III. Diacyclops humphreysi n. sp., and Comments on the Diacyclops crassicaudis-Complex (Copepoda, Cyclopidae). Crustaceana 1996, 69, 524–531. [Google Scholar]
- Jagadeesan, L.; Jyothibabu, R. Tumour-like anomaly of copepods-an evaluation of the possible causes in Indian marine waters. Environ. Monit. Assess. 2016, 188, 244. [Google Scholar] [CrossRef]
- Einsle, U. Cyclops heberti n.sp. and Cyclops singularis n.sp., two new species within the genus Cyclops (‘strenuus-subgroup’) (Crust. Copepoda) from ephemeral ponds in southern Germany. Hydrobiologia 1996, 319, 167–177. [Google Scholar] [CrossRef]
- Knight, L.; Brancelj, A.; Hänfling, B.; Cheney, C. The groundwater invertebrate fauna of the Channel Islands. Subterr. Biol. 2015, 15, 69–94. [Google Scholar] [CrossRef] [Green Version]
- Krajíček, M.; Fott, J.; Miracle, M.R.; Ventura, M.; Sommaruga, R.; Kirschner, P.; Černý, M. The genus Cyclops (Copepoda, Cyclopoida) in Europe. Zool. Scr. 2016, 45, 671–682. [Google Scholar] [CrossRef]
Figure 1.
Map of the studied 28 wells in north-eastern Poland. Numbers 1–28 are according to Table 1.
Figure 1.
Map of the studied 28 wells in north-eastern Poland. Numbers 1–28 are according to Table 1.

Figure 2.
An example of the studied wells. A—Karakule; B and C—Żuki; D—Zagruszany; E—Olmonty; F—Topolany.
Figure 2.
An example of the studied wells. A—Karakule; B and C—Żuki; D—Zagruszany; E—Olmonty; F—Topolany.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
The list of studied wells with GPS locations, the total number of copepods, and the Copepoda species (Es—Eucyclops serrulatus; Db—Diacyclops bisetosus; Dc—Diacyclops crassicaudis; Eg—Eudiaptomus gracilis; Tc—Thermocyclops crassus).
Table 1.
The list of studied wells with GPS locations, the total number of copepods, and the Copepoda species (Es—Eucyclops serrulatus; Db—Diacyclops bisetosus; Dc—Diacyclops crassicaudis; Eg—Eudiaptomus gracilis; Tc—Thermocyclops crassus).
| No. | Village | Latitude (N) | Longitude (E) | Sample Volume [l] | Total No. of Adult | Total No. of Copepodites | Copepoda Species |
|---|---|---|---|---|---|---|---|
| 1 | Cisów | 53°45′07.8″ | 23°05′25.0″ | 100 | 575 | 2177 | Es |
| 2 | Cisów | 53°44′52.7″ | 23°05′40.6″ | 100 | 0 | 0 | - |
| 3 | Wolne | 53°42′37.4″ | 23°09′18.9″ | 100 | 0 | 8 | Es, Db |
| 4 | Wolne | 53°42′43.8″ | 23°09′15.9″ | 100 | 0 | 4 | Es |
| 5 | Wolne | 53°42′37.1″ | 23°09′29.9″ | 100 | 0 | 3 | Es |
| 6 | Pasynki | 53°01′36.4″ | 23°16′08.8″ | 100 | 2 | 26 | Db |
| 7 | Pasynki | 53°01′37.2″ | 23°16′09.8″ | 100 | 0 | 6 | Es, Db |
| 8 | Pasynki | 53°01′32.0″ | 23°16′09.0″ | 100 | 13 | 63 | Es, Db |
| 9 | Pasynki | 53°01′33.2″ | 23°16′08.4″ | 100 | 14 | 98 | Es |
| 10 | Olmonty | 53°5′2.22″ | 23°9′50.6″ | 70 | 0 | 0 | - |
| 11 | Olmonty | 53°5′2.42″ | 23°10′18.83″ | 50 | 0 | 0 | - |
| 12 | Halickie | 53°3′54.53″ | 23°14′0.09″ | 80 | 0 | 0 | - |
| 13 | Nowosady | 53°1′15.03″ | 23°13′16.38″ | 85 | 0 | 0 | - |
| 14 | Nowosady | 53°0′16.11″ | 23°13′21.54″ | 80 | 0 | 0 | - |
| 15 | Żuki | 53°0′41.82″ | 23°14′54.19″ | 60 | 0 | 0 | - |
| 16 | Żuki | 53°0′49.19″ | 23°14′58.24″ | 55 | 2 | 1 | Dc |
| 17 | Zagruszany | 53°2′13.58″ | 23°17′14.33″ | 60 | 12 | 66 | Db, Es |
| 18 | Folwarki Małe | 53°2′12.80″ | 23°23′21.44″ | 80 | 5 | 13 | Dc |
| 19 | Folwarki Wielkie | 53°1′44.61″ | 23°24′22.41″ | 55 | 0 | 0 | - |
| 20 | Folwarki Wielkie | 53°1′33.27″ | 23°24′40.37″ | 60 | 29 | 2 | Dc |
| 21 | Folwarki Tylwickie | 53°1′6.97″ | 23°25′34.42″ | 60 | 68 | 331 | Dc, Cf |
| 22 | Topolany | 53°1′9.46″ | 23°30′0.22″ | 55 | 34 | 16 | Dc, Db, Cf |
| 23 | Topolany | 53°1′11.49″ | 23°30′17.94″ | 50 | 7 | 4 | Db, Cf |
| 24 | Karakule | 53°10′21.29″ | 23°15′34.54″ | 60 | 0 | 0 | - |
| 25 | Karakule | 53°10′21.89″ | 23°15′38.48″ | 60 | 0 | 0 | - |
| 26 | Karakule | 53°10′57.65″ | 23°15′47.74″ | 60 | 0 | 0 | - |
| 27 | Karakule | 53°10′33.61″ | 23°15′42.82″ | 60 | 0 | 0 | - |
| 28 | Białystok city | 53°06′28.26″ | 23°12′01.02″ | 50 | 0 | 2 | Eg, Tc |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Karpowicz, M.; Smolska, S.; Świsłocka, M.; Moroz, J. First Insight into Groundwater Copepods of the Polish Lowland. Water 2021, 13, 2086. https://doi.org/10.3390/w13152086
AMA Style
Karpowicz M, Smolska S, Świsłocka M, Moroz J. First Insight into Groundwater Copepods of the Polish Lowland. Water. 2021; 13(15):2086. https://doi.org/10.3390/w13152086
Chicago/Turabian StyleKarpowicz, Maciej, Sabina Smolska, Magdalena Świsłocka, and Joanna Moroz. 2021. "First Insight into Groundwater Copepods of the Polish Lowland" Water 13, no. 15: 2086. https://doi.org/10.3390/w13152086
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.