
Short Communication: Quantification of the Effect of Mycotoxin Binders on the Bioavailability of Fat-Soluble Vitamins In Vitro

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design and Incubations
2.2. Calculations and Statistical Analysis
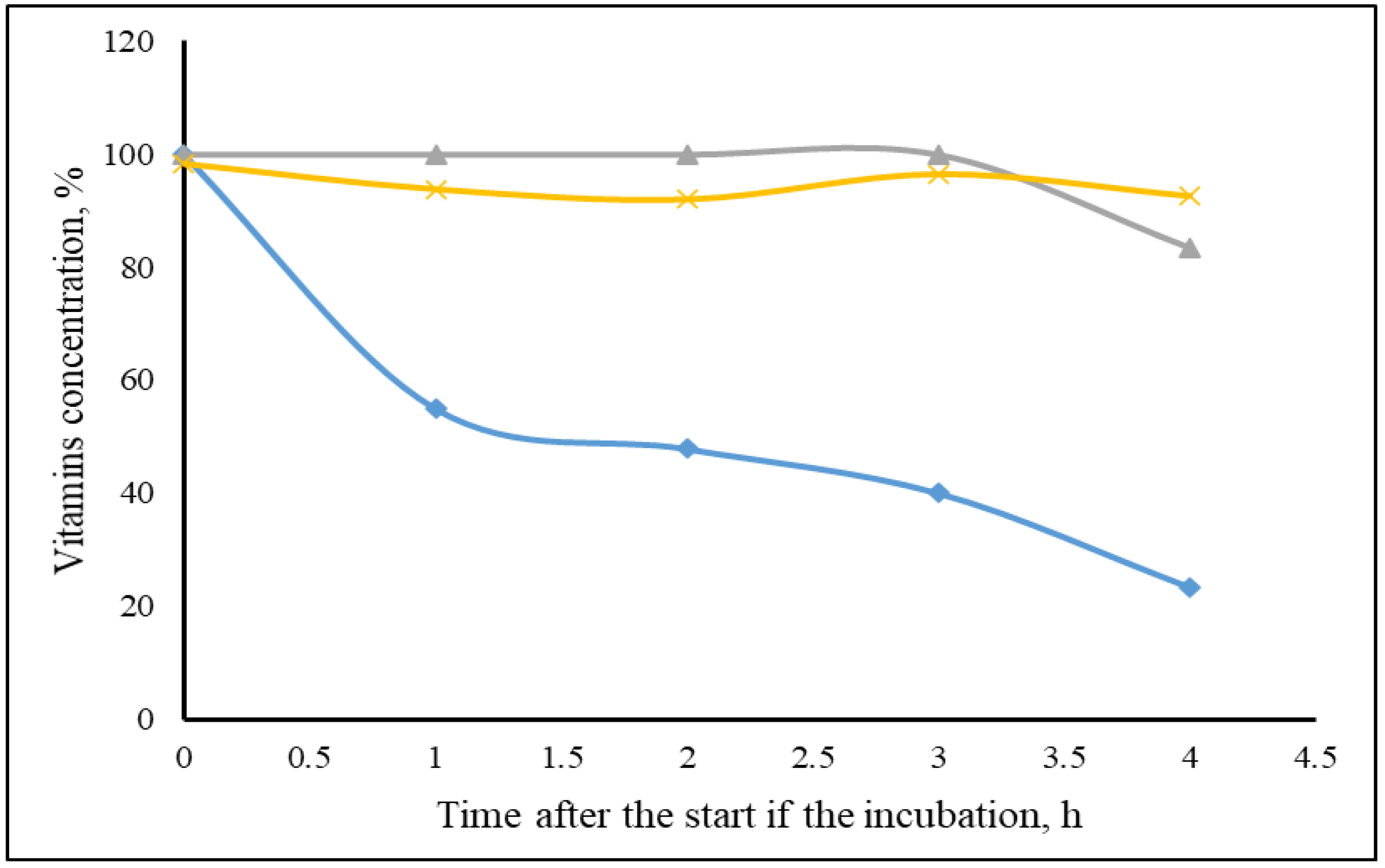
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Moretti, A.; Pascale, M.; Logrieco, A.F. Mycotoxin risks under a climate change scenario in Europe. Trends Food Sci. Technol. 2019, 84, 38–40. [Google Scholar] [CrossRef]
- Biomin®. World Mycotoxin Survey: Impact 2020. Available online: https://www.biomin.net/science-hub/world-mycotoxin-survey-impact-2020/ (accessed on 18 February 2020).
- Akande, K.E.; Abubakar, M.M.; Adegbola, T.A.; Bogoro, S.E. Nutritional and health implications of mycotoxins in animal feeds: A review. Pak. J. Nutr. 2006, 5, 398–403. [Google Scholar]
- Diaz, D.E.; Hagler, W.M.; Blackwelder, J.T.; Eve, J.A.; Hopkins, B.A.; Anderson, K.L. Aflatoxin Binders II: Reduction of aflatoxin M1 in milk by sequestering agents of cows consuming aflatoxin in feed. Mycopathologia 2004, 157, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Avantaggiato, G.; Solfrizzo, M.; Visconti, A. Recent advances on the use of adsorbent materials for detoxification of Fusarium mycotoxins. Pak. J. Nutr. 2006, 5, 398–403. [Google Scholar] [CrossRef] [Green Version]
- Sabater-vilar, M.; Malekinejad, H.; Selman, M.H.J.; Fink-gremmels, J. In vitro assessment of adsorbents aiming to prevent deoxynivalenol and zearalenone mycotoxicoses. Mycopathologia 2007, 163, 81–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- European Food Safety Authority (EFSA). Opinion of the Scientific Panel on Additives and Products or Substances used in Animal Feed (FEEDAP Panel) on the safety and efficacy of bentonite (dioctahedral montmorillonite) as feed additive for all species. EFSA J. 2011, 9, 2007. [Google Scholar] [CrossRef] [Green Version]
- Tomasevic-Canovi, M.C.; Dakovic, A.; Markovic, V.; Radosavljevic-Mihajlovic, A.; Vukicevic, J. Adsorption effects of mineral adsorbents; part III: Adsorption behaviour in the presence of vitamin B6 and microelements. Acta Vet. 2000, 50, 23–30. [Google Scholar]
- Kihal, A.; Rodriguez-Prado, M.; Godoy, C.; Cristofol, C.; Calsamiglia, S. In vitro assessment of the capacity of certain mycotoxin binders to adsorb some amino acids and water-soluble vitamins. J. Dairy Sci. 2020, 103, 3125–3132. [Google Scholar] [CrossRef] [PubMed]
- Barrientos-Velázquez, A.L.; Arteaga, S.; Dixon, J.B.; Deng, Y. The effects of pH, pepsin, exchange cation, and vitamins on aflatoxin adsorption on smectite in simulated gastric fluids. Appl. Clay Sci. 2016, 120, 17–23. [Google Scholar] [CrossRef]
- Lemke, S.L.; Ottinger, S.E.; Mayura, K.; Ake, C.L.; Pimpukdee, K.; Wang, N. Development of a multi-tiered approach to the in vitro prescreening of clay-based enterosorbents. Anim. Feed Sci. Technol. 2001, 93, 17–29. [Google Scholar] [CrossRef]
- Gallo, A.; Masoero, F. In vitro models to evaluate the capacity of different sequestering agents to adsorb aflatoxins In vitro models to evaluate sequestering agents to adsorb aflatoxins. Ital. J. Anim. Sci. 2017, 9, e21. [Google Scholar] [CrossRef] [Green Version]
- Barroeta, A.C.; Baucells, M.D.; Calsamiglia, S.; Casals, R.; Briz, R.C.; Davin, R. Optimum Vitamin Nutrition; 5M Publishing: Sheffield, UK, 2012. [Google Scholar]
- Sánchez, R.C.; Martorell, J.C.; Baldoví, E.C. Neutralización de micotoxinas por sustancias adsorbentes. Revisión del tema. Nereis 2012, 4, 77–88. [Google Scholar]
- Mortland, M.M.; Lawless, J.G. Smectite interaction with riboflavin. Clays. Clay Miner. 1983, 31, 435–439. [Google Scholar] [CrossRef]


{kind=link}
| Vitamins | Mycotoxin Binders | |||||
|---|---|---|---|---|---|---|
| Bentonite | Clinoptilolite | Sepiolite | MMT 1 | AC 2 | YCW 3 | |
| Incubated separately | ||||||
| D | −4.4 z,c | 2.9 z,b | 8.6 b | 5.2 z,b | 8.5 b | 20.2 a |
| E | 54.5 y,a | 34.1 y,b | 16.6 c | 46.3 y,a | 18.5 c | 30.0 b |
| SEM 4 1 | 5.88 | |||||
| Incubated together | ||||||
| D | −2.5 z,b | −2.5 z,b | −0.3 z,b | 7.4 z,a | −2.9 z,b | −1.0 z,b |
| E | 61.8 y,a | 38.3 y,b | 16.6 y,d | 50.7 y,a | 28.5 y,bc | 23.7 y,cd |
| SEM 2 | 3.54 | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kihal, A.; Rodríguez-Prado, M.E.; Cristofol, C.; Calsamiglia, S. Short Communication: Quantification of the Effect of Mycotoxin Binders on the Bioavailability of Fat-Soluble Vitamins In Vitro. Animals 2021, 11, 2251. https://doi.org/10.3390/ani11082251
Kihal A, Rodríguez-Prado ME, Cristofol C, Calsamiglia S. Short Communication: Quantification of the Effect of Mycotoxin Binders on the Bioavailability of Fat-Soluble Vitamins In Vitro. Animals. 2021; 11(8):2251. https://doi.org/10.3390/ani11082251
Chicago/Turabian StyleKihal, Abdelhacib, María Ercilda Rodríguez-Prado, Carles Cristofol, and Sergio Calsamiglia. 2021. "Short Communication: Quantification of the Effect of Mycotoxin Binders on the Bioavailability of Fat-Soluble Vitamins In Vitro" Animals 11, no. 8: 2251. https://doi.org/10.3390/ani11082251