
miR-126 Controls the Apoptosis and Proliferation of Immature Porcine Sertoli Cells by Targeting the PIK3R2 Gene through the PI3K/AKT Signaling Pathway

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Statement
2.2. Cell Culture and Transfection
2.3. Cell Cycle Assay
2.4. CCK-8 Assay
2.5. 5-Ethynyl-2′-deoxyuridine (EdU) Assay
2.6. Cell Apoptosis Assay
2.7. Dual-Luciferase Reporter Assay
2.8. Real Time qPCR
2.9. Western Blotting
2.10. Statistical Analysis
3. Results
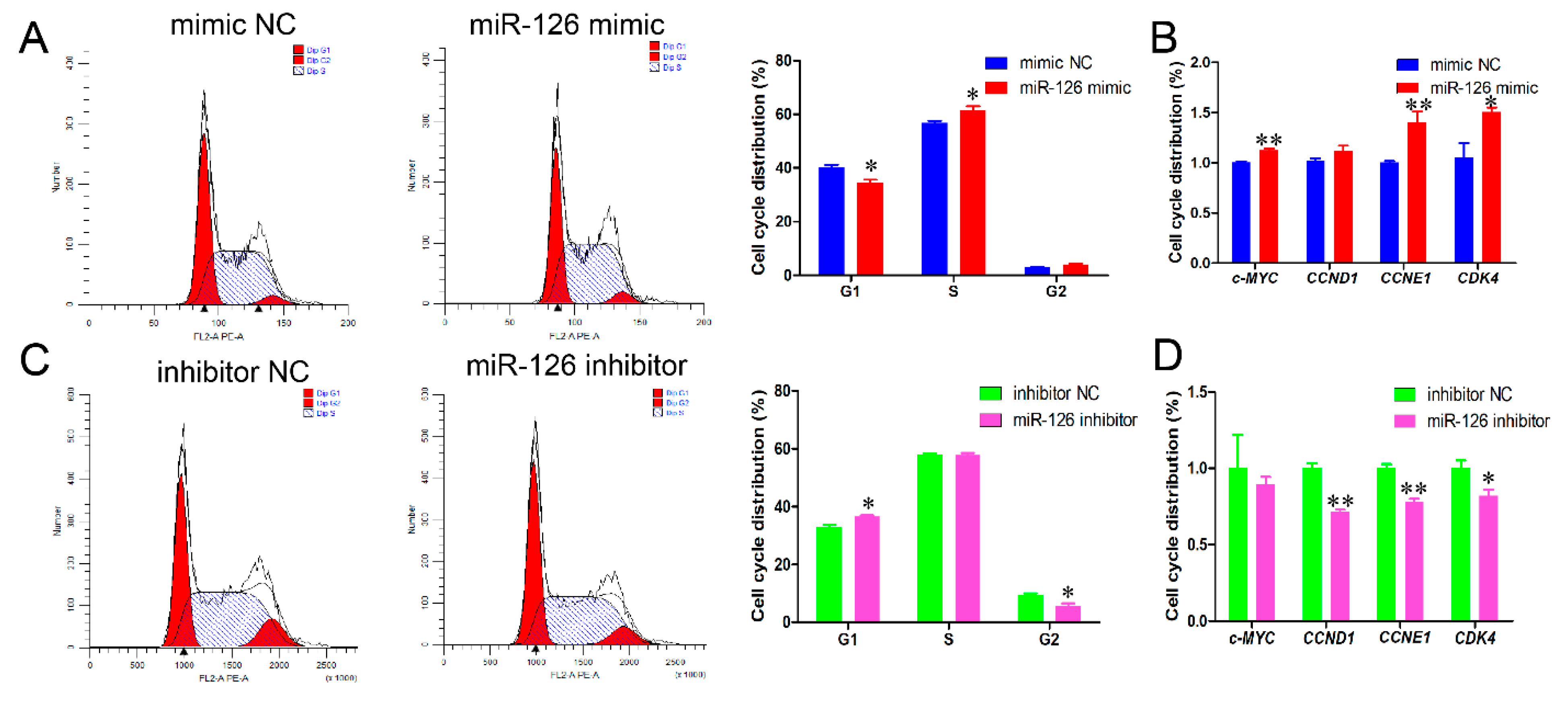
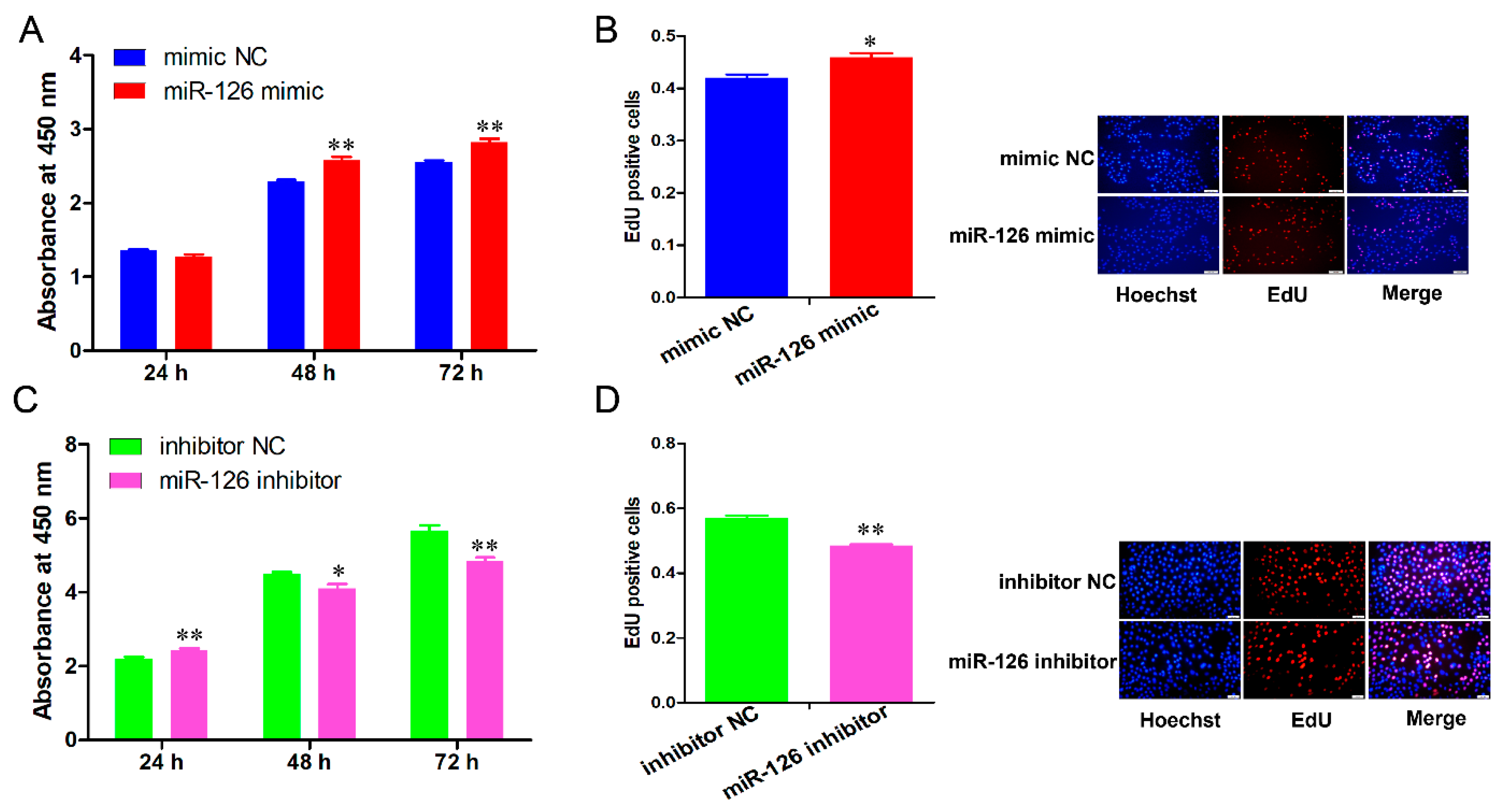
3.1. miR-126 Promotes the Cell Cycle Progression and Proliferation of Immature Porcine Sertoli Cells
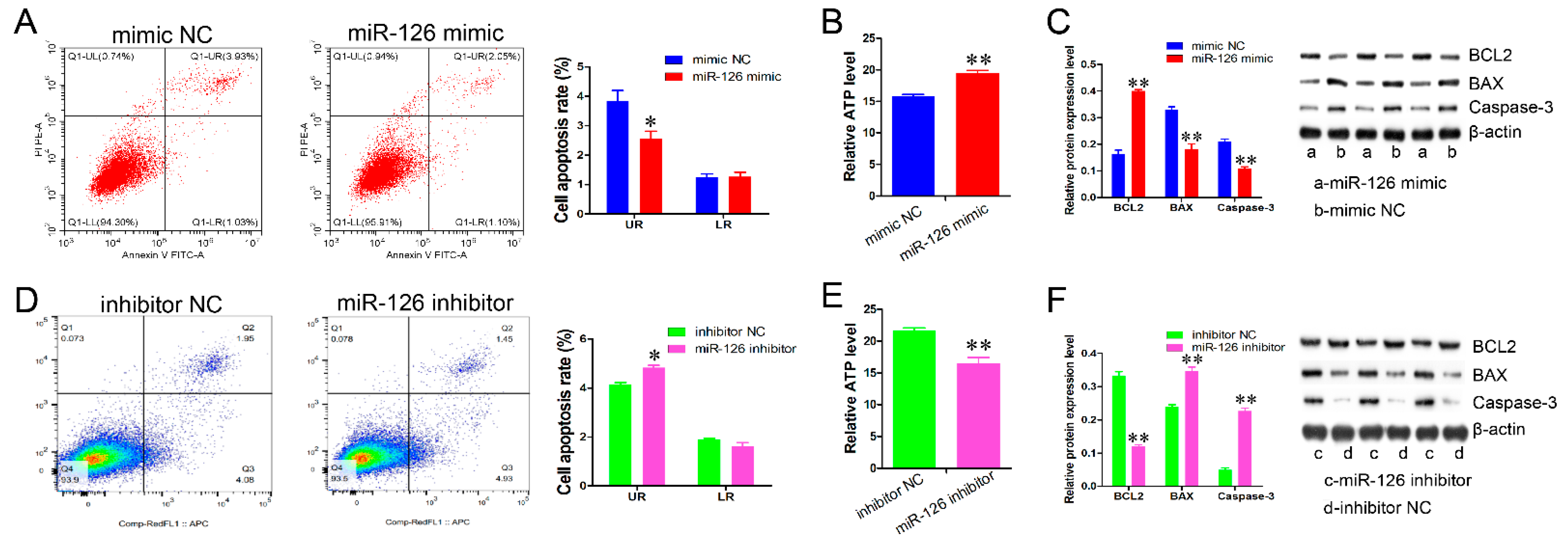
3.2. miR-126 Inhibits Immature Porcine Sertoli Cell Apoptosis
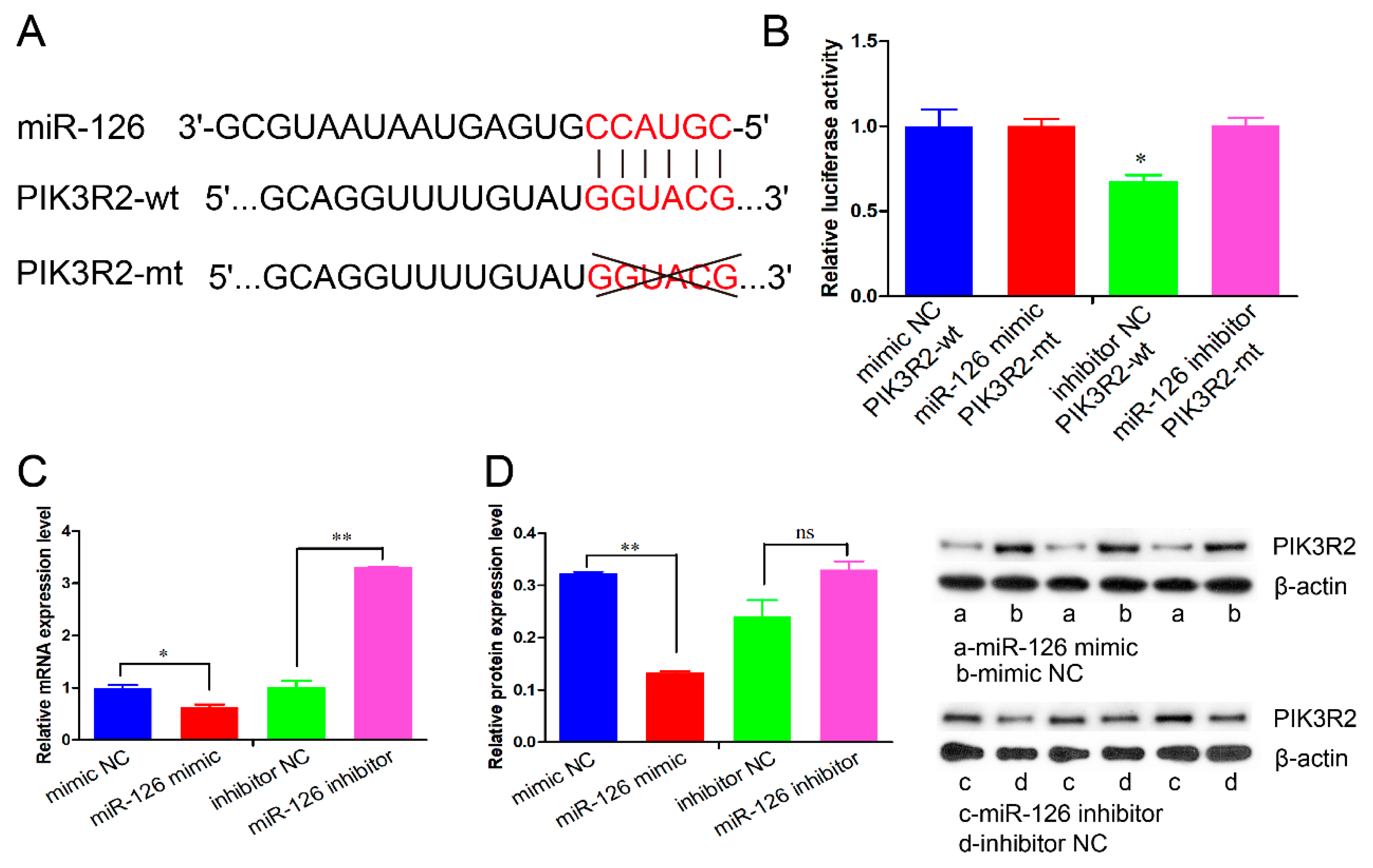
3.3. miR-126 Directly Targets the PIK3R2 Gene
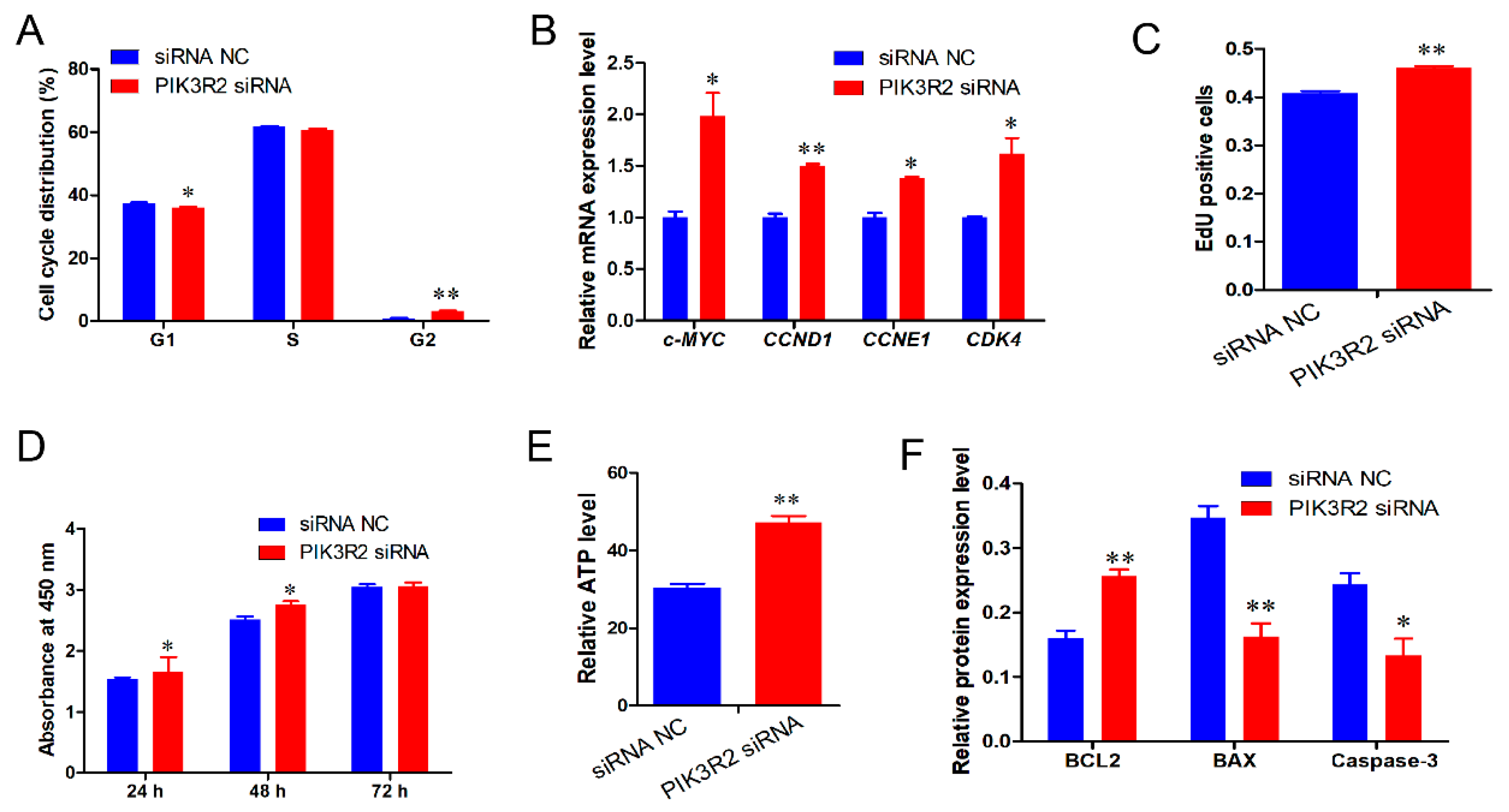
3.4. PIK3R2 Gene Deficiency Promotes Immature Porcine Sertoli Cell Proliferation and Inhibits Apoptosis
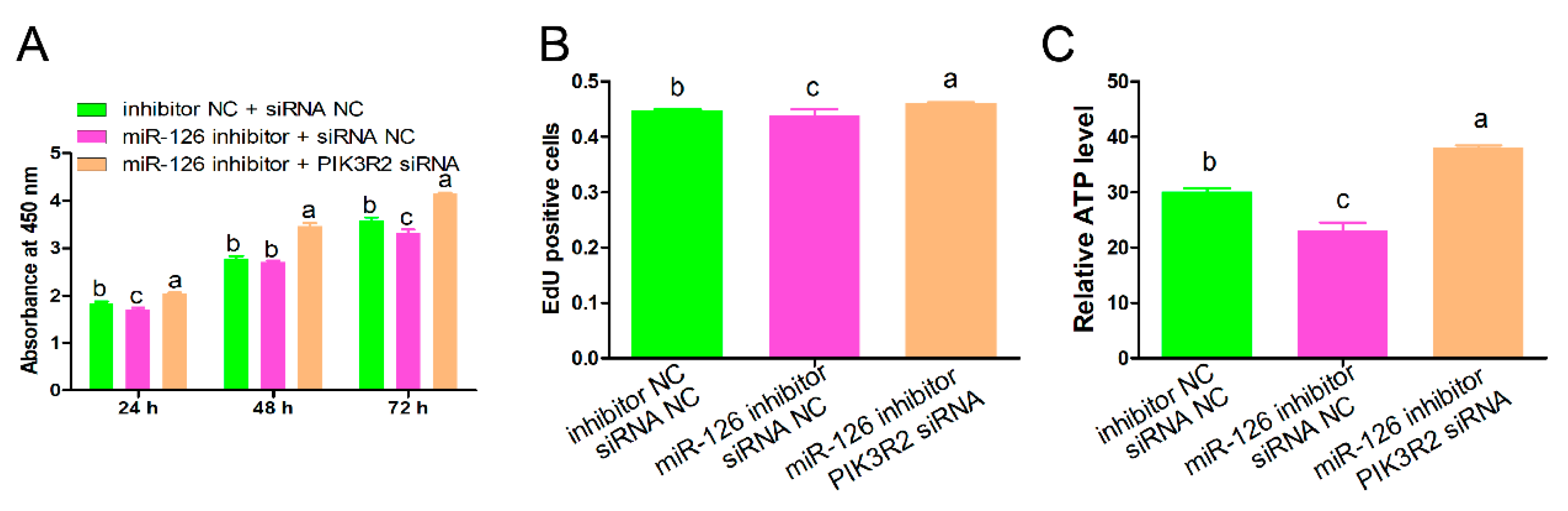
3.5. PIK3R2 Knockdown Attenuated the Regulatory Roles of miR-126 Inhibition
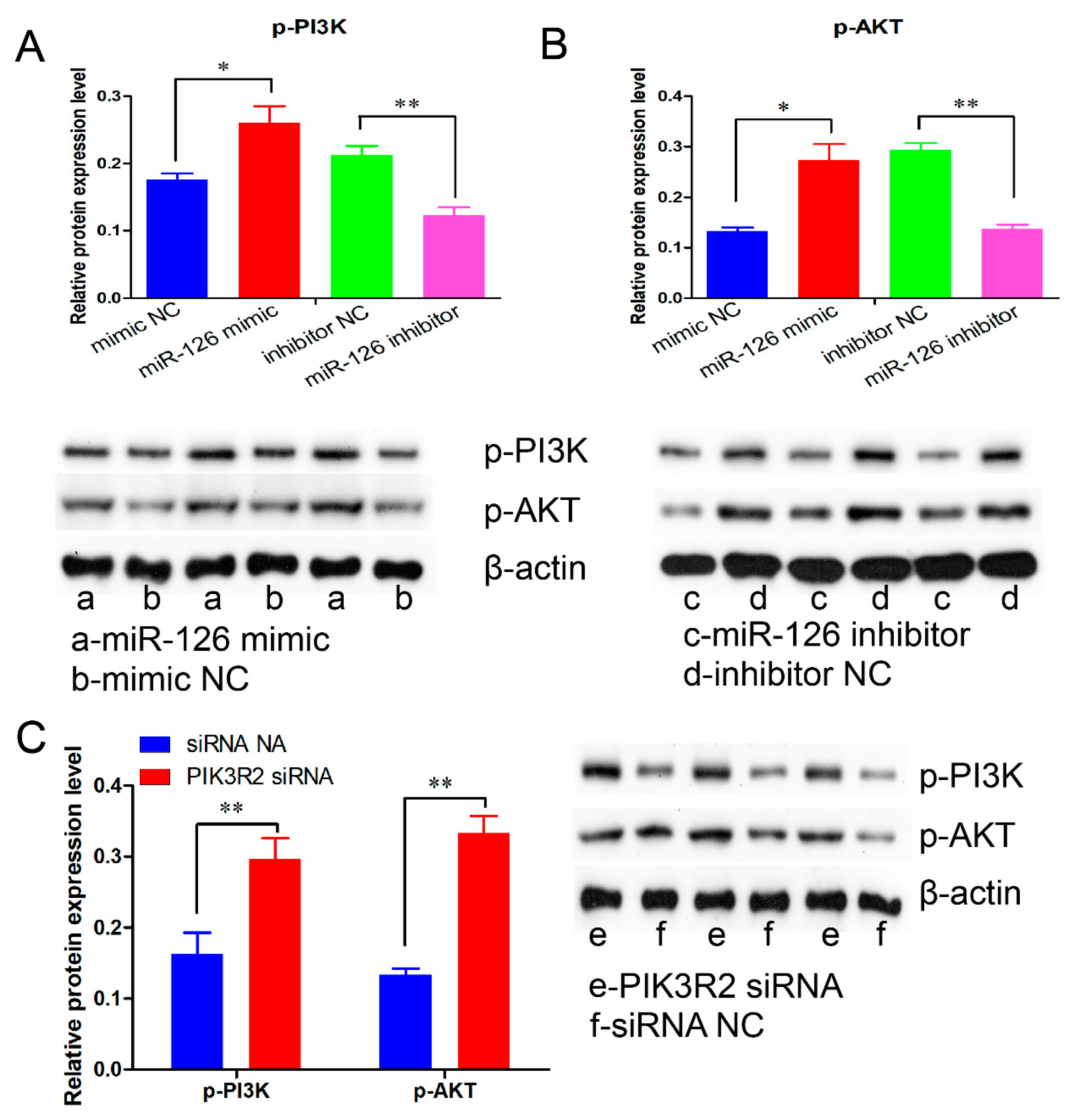
3.6. miR-126 Promotes Immature Porcine Sertoli Cell Growth by Activating the PI3K/AKT Signaling Pathway
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Griswold, M.D. 50 years of spermatogenesis: Sertoli cells and their interactions with germ cells. Biol. Reprod. 2018, 99, 87–100. [Google Scholar] [CrossRef] [PubMed]
- Rebourcet, D.; Darbey, A.; Monteiro, A.; Soffientini, U.; Tsai, Y.T.; Handel, I.; Pitetti, J.L.; Nef, S.; Smith, L.B.; O’Shaughnessy, P.J. Sertoli Cell Number Defines and Predicts Germ and Leydig Cell Population Sizes in the Adult Mouse Testis. Endocrinology 2017, 158, 2955–2969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franca, L.R.; Silva, V.A., Jr.; Chiarini-Garcia, H.; Garcia, S.K.; Debeljuk, L. Cell proliferation and hormonal changes during postnatal development of the testis in the pig. Biol. Reprod. 2000, 63, 1629–1636. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, A.R.; Macheroni, C.; Lucas, T.F.; Porto, C.S.; Lazari, M.F. Crosstalk between FSH and relaxin at the end of the proliferative stage of rat Sertoli cells. Reproduction 2016, 152, 613–628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basu, S.; Arya, S.P.; Usmani, A.; Pradhan, B.S.; Sarkar, R.K.; Ganguli, N.; Shukla, M.; Mandal, K.; Singh, S.; Sarda, K.; et al. Defective Wnt3 expression by testicular Sertoli cells compromise male fertility. Cell Tissue Res. 2018, 371, 351–363. [Google Scholar] [CrossRef]
- Meroni, S.B.; Galardo, M.N.; Rindone, G.; Gorga, A.; Riera, M.F.; Cigorraga, S.B. Molecular Mechanisms and Signaling Pathways Involved in Sertoli Cell Proliferation. Front. Endocrinol. 2019, 10, 224. [Google Scholar] [CrossRef]
- Luo, H.; Chen, B.; Weng, B.; Tang, X.; Chen, Y.; Yang, A.; Chu, D.; Zeng, X.; Ran, M. miR-130a promotes immature porcine Sertoli cell growth by activating SMAD5 through the TGF-beta-PI3K/AKT signaling pathway. FASEB J. 2020, 34, 15164–15179. [Google Scholar] [CrossRef]
- Jansen, F.; Stumpf, T.; Proebsting, S.; Franklin, B.S.; Wenzel, D.; Pfeifer, P.; Flender, A.; Schmitz, T.; Yang, X.; Fleischmann, B.K.; et al. Intercellular transfer of miR-126-3p by endothelial microparticles reduces vascular smooth muscle cell proliferation and limits neointima formation by inhibiting LRP6. J. Mol. Cell. Cardiol. 2017, 104, 43–52. [Google Scholar] [CrossRef]
- Tao, H.; Wang, M.M.; Zhang, M.; Zhang, S.P.; Wang, C.H.; Yuan, W.J.; Sun, T.; He, L.J.; Hu, Q.K. MiR-126 Suppresses the Glucose-Stimulated Proliferation via IRS-2 in INS-1 beta Cells. PLoS ONE 2016, 11, e0149954. [Google Scholar] [CrossRef]
- Ding, Q.; Wang, Q.; Ren, Y.; Zhu, H.; Huang, Z. miR-126 promotes the growth and proliferation of leukemia stem cells by targeting DNA methyltransferase 1. Int. J. Clin. Exp. Pathol. 2018, 11, 3454–3462. [Google Scholar]
- Xiang, G.; Cheng, Y. MiR-126-3p inhibits ovarian cancer proliferation and invasion via targeting PLXNB2. Reprod. Biol. 2018, 18, 218–224. [Google Scholar] [CrossRef]
- Yuan, X.; Deng, X.; Zhou, X.; Zhang, A.; Xing, Y.; Zhang, Z.; Zhang, H.; Li, J. MiR-126-3p promotes the cell proliferation and inhibits the cell apoptosis by targeting TSC1 in the porcine granulosa cells. Vitr. Cell. Dev. Biol. Anim. 2018, 54, 715–724. [Google Scholar] [CrossRef]
- Gao, J.; Zhou, X.L.; Kong, R.N.; Ji, L.M.; He, L.L.; Zhao, D.B. microRNA-126 targeting PIK3R2 promotes rheumatoid arthritis synovial fibro-blasts proliferation and resistance to apoptosis by regulating PI3K/AKT pathway. Exp. Mol. Pathol. 2016, 100, 192–198. [Google Scholar] [CrossRef]
- Xiao, J.; Lin, H.Y.; Zhu, Y.Y.; Zhu, Y.P.; Chen, L.W. MiR-126 regulates proliferation and invasion in the bladder cancer BLS cell line by targeting the PIK3R2-mediated PI3K/Akt signaling pathway. OncoTargets Ther. 2016, 9, 5181–5193. [Google Scholar]
- Rahman, M.A.; Salajegheh, A.; Smith, R.A.; Lam, A.K. MicroRNA-126 suppresses proliferation of undifferentiated (BRAF(V600E) and BRAF(WT)) thyroid carcinoma through targeting PIK3R2 gene and repressing PI3K-AKT proliferation-survival signalling pathway. Exp. Cell Res. 2015, 339, 342–350. [Google Scholar] [CrossRef]
- Yang, W.R.; Zhu, F.W.; Zhang, J.J.; Wang, Y.; Zhang, J.H.; Lu, C.; Wang, X.Z. PI3K/Akt Activated by GPR30 and Src Regulates 17beta-Estradiol-Induced Cultured Immature Boar Sertoli Cells Proliferation. Reprod. Sci. 2017, 24, 57–66. [Google Scholar] [CrossRef]
- Sun, Y.; Yang, W.; Luo, H.; Wang, X.; Chen, Z.; Zhang, J.; Wang, Y.; Li, X. Thyroid hormone inhibits the proliferation of piglet Sertoli cell via PI3K signaling pathway. Theriogenology 2015, 83, 86–94. [Google Scholar] [CrossRef]
- Hu, P.; Guan, K.; Feng, Y.; Ma, C.; Song, H.; Li, Y.; Xia, X.; Li, J.; Li, F. miR-638 Inhibits immature Sertoli cell growth by indirectly inactivating PI3K/AKT pathway via SPAG1 gene. Cell Cycle 2017, 16, 2290–2300. [Google Scholar] [CrossRef] [Green Version]
- Ma, C.; Song, H.; Guan, K.; Zhou, J.; Xia, X.; Li, F. Characterization of swine testicular cell line as immature porcine Sertoli cell line. In Vitro Cell. Dev. Biol. Anim. 2016, 52, 427–433. [Google Scholar] [CrossRef]
- Ran, M.L.; Weng, B.; Cao, R.; Li, Z.; Peng, F.Z.; Luo, H.; Gao, H.; Chen, B. miR-26a inhibits proliferation and promotes apoptosis in porcine immature Sertoli cells by targeting the PAK2 gene. Reprod. Domest. Anim. 2018, 53, 1375–1385. [Google Scholar] [CrossRef]
- Luo, Z.; Liu, Y.; Chen, L.; Ellis, M.; Li, M.; Wang, J.; Zhang, Y.; Fu, P.; Wang, K.; Li, X.; et al. microRNA profiling in three main stages during porcine spermatogenesis. J. Assist. Reprod. Genet. 2015, 32, 451–460. [Google Scholar] [CrossRef] [Green Version]
- Luo, Z.Y.; Dai, X.L.; Ran, X.Q.; Cen, Y.X.; Niu, X.; Li, S.; Huang, S.H.; Wang, J.F. Identification and profile of microRNAs in Xiang pig testes in four different ages detected by Solexa sequencing. Theriogenology 2018, 117, 61–71. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Li, J.; Fang, C.; Shi, L.; Tan, J.; Xiong, Y.; Bin, F.; Li, C. Genome-wide differential expression of genes and small RNAs in testis of two different porcine breeds and at two different ages. Sci. Rep. 2016, 6, 26852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ran, M.L.; Chen, B.; Wu, M.S.; Liu, X.C.; He, C.Q.; Yang, A.Q.; Li, Z.; Xiang, Y.J.; Li, Z.H.; Zhang, S.W. Integrated analysis of miRNA and mRNA expression profiles in development of porcine testes. RSC Adv. 2015, 5, 63439–63449. [Google Scholar] [CrossRef]
- Ma, C.; Song, H.; Yu, L.; Guan, K.; Hu, P.; Li, Y.; Xia, X.; Li, J.; Jiang, S.; Li, F. miR-762 promotes porcine immature Sertoli cell growth via the ring finger protein 4 (RNF4) gene. Sci. Rep. 2016, 6, 32783. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Zhang, P.; Li, L.; Che, D.; Li, T.; Li, H.; Li, Q.; Jia, H.; Tao, S.; Hua, J.; et al. miRNA editing landscape reveals miR-34c regulated spermatogenesis through structure and target change in pig and mouse. Biochem. Biophys. Res. Commun. 2018, 502, 486–492. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Yang, B.; Pan, P.; Ma, Q.; Wu, Y.; Zhang, Z.; Guo, X.; Ye, J.; Gui, Y. MicroRNA-130a inhibits spermatogenesis by directly targeting androgen receptor in mouse Sertoli cells. Mol. Reprod. Dev. 2018, 85, 768–777. [Google Scholar] [CrossRef]
- Yang, Y.; Xue, K.; Li, Z.; Zheng, W.; Dong, W.; Song, J.; Sun, S.; Ma, T.; Li, W. c-Myc regulates the CDK1/cyclin B1 dependentG2/M cell cycle progression by histone H4 acetylation in Raji cells. Int. J. Mol. Med. 2018, 41, 3366–3378. [Google Scholar] [PubMed] [Green Version]
- Bretones, G.; Delgado, M.D.; Leon, J. Myc and cell cycle control. Biochim. Biophys. Acta 2015, 1849, 506–516. [Google Scholar] [CrossRef]
- Swaffer, M.P.; Jones, A.W.; Flynn, H.R.; Snijders, A.P.; Nurse, P. CDK Substrate Phosphorylation and Ordering the Cell Cycle. Cell 2016, 167, 1750–1761.e16. [Google Scholar] [CrossRef] [Green Version]
- Attwooll, C.; Lazzerini Denchi, E.; Helin, K. The E2F family: Specific functions and overlapping interests. EMBO J. 2004, 23, 4709–4716. [Google Scholar] [CrossRef]
- Trimarchi, J.M.; Lees, J.A. Sibling rivalry in the E2F family. Nat. Rev. Mol. Cell Biol. 2002, 3, 11–20. [Google Scholar] [CrossRef]
- Zhou, X.; He, Y.; Jiang, Y.; He, B.; Deng, X.; Zhang, Z.; Yuan, X.; Li, J. MiR-126-3p inhibits apoptosis and promotes proliferation by targeting phosphatidylinositol 3-kinase regulatory subunit 2 in porcine ovarian granulosa cells. Asian-Australas J. Anim. Sci. 2020, 33, 879–887. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Wang, J.; Wang, B.; Yang, J.; Gong, Z.; Zhao, X.; Zhang, C.; Du, K. MiR-126 inhibits vascular endothelial cell apoptosis through targeting PI3K/Akt signaling. Ann. Hematol. 2016, 95, 365–374. [Google Scholar] [CrossRef]
- Vallejo-Diaz, J.; Chagoyen, M.; Olazabal-Moran, M.; Gonzalez-Garcia, A.; Carrera, A.C. The Opposing Roles of PIK3R1/p85alpha and PIK3R2/p85beta in Cancer. Trends Cancer 2019, 5, 233–244. [Google Scholar] [CrossRef] [Green Version]
- Dobyns, W.B.; Mirzaa, G.M. Megalencephaly syndromes associated with mutations of core components of the PI3K-AKT-MTOR pathway: PIK3CA, PIK3R2, AKT3, and MTOR. Am. J. Med. Genet. C Semin. Med. Genet. 2019, 181, 582–590. [Google Scholar] [CrossRef]
- Qu, Y.; Zhang, Y.P.; Wu, J.; Jie, L.G.; Deng, J.X.; Zhao, D.B.; Yu, Q.H. Downregulated microRNA-135a ameliorates rheumatoid arthritis by inactivation of the phosphatidylinositol 3-kinase/AKT signaling pathway via phosphatidylinositol 3-kinase regulatory subunit 2. J. Cell. Physiol. 2019, 234, 17663–17676. [Google Scholar] [CrossRef] [PubMed]
- Nie, Z.C.; Weng, W.H.; Shang, Y.S.; Long, Y.; Li, J.; Xu, Y.T.; Li, Z. MicroRNA-126 is down-regulated in human esophageal squamous cell carcinoma and inhibits the proliferation and migration in EC109 cell via PI3K/AKT signaling pathway. Int. J. Clin. Exp. Pathol. 2015, 8, 4745–4754. [Google Scholar] [PubMed]
- Riera, M.F.; Regueira, M.; Galardo, M.N.; Pellizzari, E.H.; Meroni, S.B.; Cigorraga, S.B. Signal transduction pathways in FSH regulation of rat Sertoli cell proliferation. Am. J. Physiol. Endocrinol. Metab. 2012, 302, 914–923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, H.; Ran, M.; Luo, H.; Weng, B.; Tang, X.; Chen, Y.; Yang, A.; Chen, B. miR-499 promotes immature porcine Sertoli cell growth by the PI3K/AKT pathway by targeting the PTEN gene. Reproduction 2019, 159, 145–157. [Google Scholar] [CrossRef] [PubMed]
- Weng, B.; Ran, M.L.; Chen, B.; Wu, M.S.; Peng, F.Z.; Dong, L.H.; He, C.Q.; Zhang, S.W.; Li, Z.H. Systematic identification and characterization of miRNAs and piRNAs from porcine testes. Genes Genom. 2017, 39, 1047–1057. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer | Sequence (5′→3′) |
|---|---|---|
| CCND1 | F | TACACCGACAACTCCATCCG |
| R | GCCGCCAGGTTCCACTT | |
| CCNE1 | F | CCTGCTGAAGATGCCCATAAC |
| R | TGCTCTGCTTCTTACTGCTCG | |
| CDK-4 | F | GTGGCCCTCAAGAGCGTAAG |
| R | CAGACATCCATCAGCCGGAC | |
| C-MYC | F | AACCCTTGGCTCTCCACGAG |
| R | ATTCCGACCTTTTGGCAGGG | |
| PIK3R2 | F | ACTTAGGAAAGGCGGGAACAACAAG |
| R | ACGACAGAGCAGAAGGTGAGAGG | |
| miR-126 | F | CGCGTCGTACCGTGAGTAAT |
| R | AGTGCAGGGTCCGAGGTATT | |
| Pig-TBP | F | GCGATTTGCTGCTGTAATCA |
| R | CCCCACCATGTTCTGAATCT | |
| U6 | F | CTCGCTTCGGCAGCACA |
| R | AACGCTTCACGAATTTGCGT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tang, X.; Chen, Y.; Luo, H.; Bian, Q.; Weng, B.; Yang, A.; Chu, D.; Ran, M.; Chen, B. miR-126 Controls the Apoptosis and Proliferation of Immature Porcine Sertoli Cells by Targeting the PIK3R2 Gene through the PI3K/AKT Signaling Pathway. Animals 2021, 11, 2260. https://doi.org/10.3390/ani11082260
Tang X, Chen Y, Luo H, Bian Q, Weng B, Yang A, Chu D, Ran M, Chen B. miR-126 Controls the Apoptosis and Proliferation of Immature Porcine Sertoli Cells by Targeting the PIK3R2 Gene through the PI3K/AKT Signaling Pathway. Animals. 2021; 11(8):2260. https://doi.org/10.3390/ani11082260
Chicago/Turabian StyleTang, Xiangwei, Yao Chen, Hui Luo, Qiao Bian, Bo Weng, Anqi Yang, Dan Chu, Maoliang Ran, and Bin Chen. 2021. "miR-126 Controls the Apoptosis and Proliferation of Immature Porcine Sertoli Cells by Targeting the PIK3R2 Gene through the PI3K/AKT Signaling Pathway" Animals 11, no. 8: 2260. https://doi.org/10.3390/ani11082260