The Effect of Low Temperature on the Early Life Stages of the Walleye Pollock, Gadus chalcogrammus—A Laboratory Study

Abstract
:1. Introduction
2. Materials and Methods
2.1. Eggs and Larvae
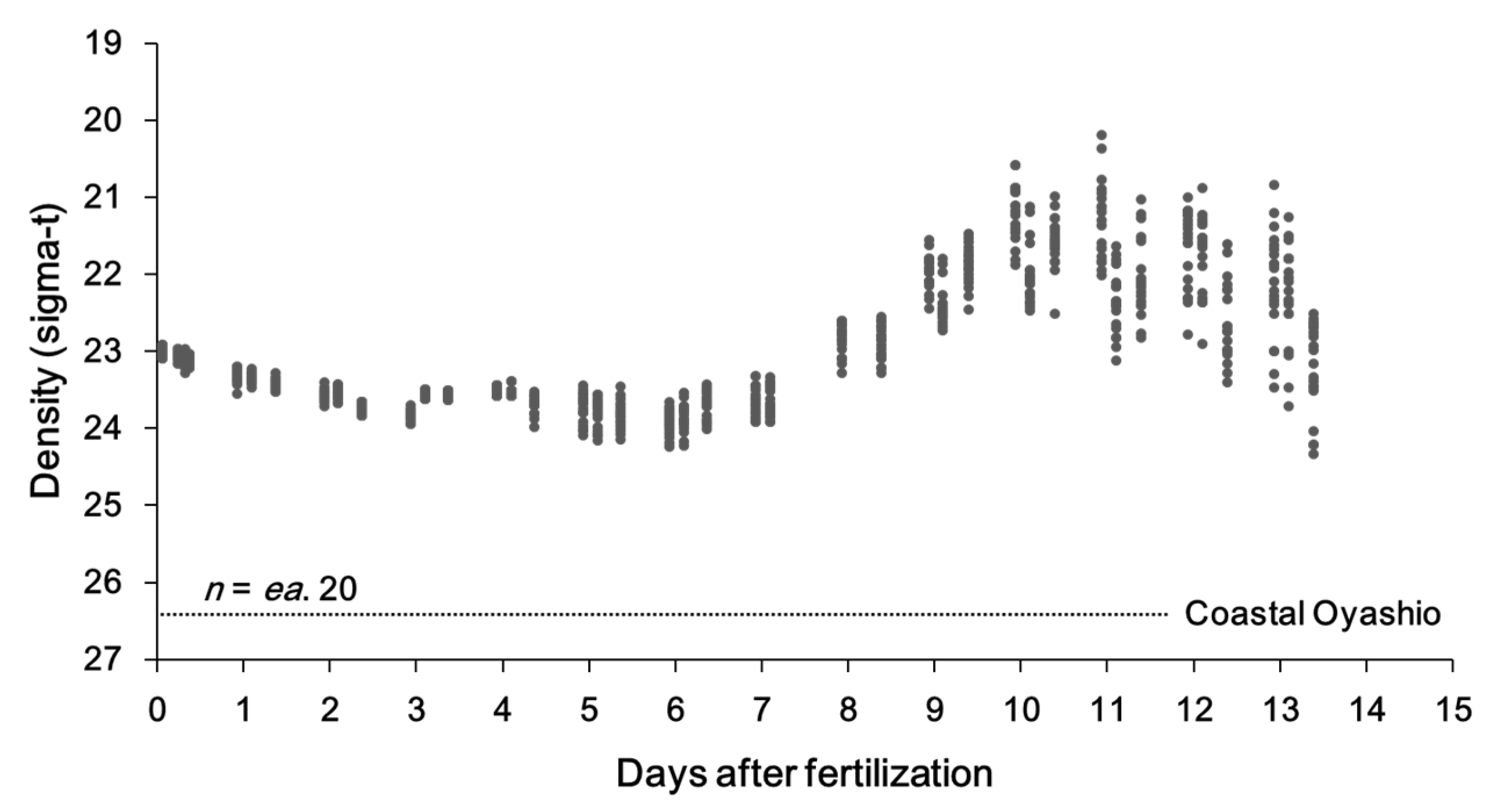
2.2. Specific Density during Early Ontogeny
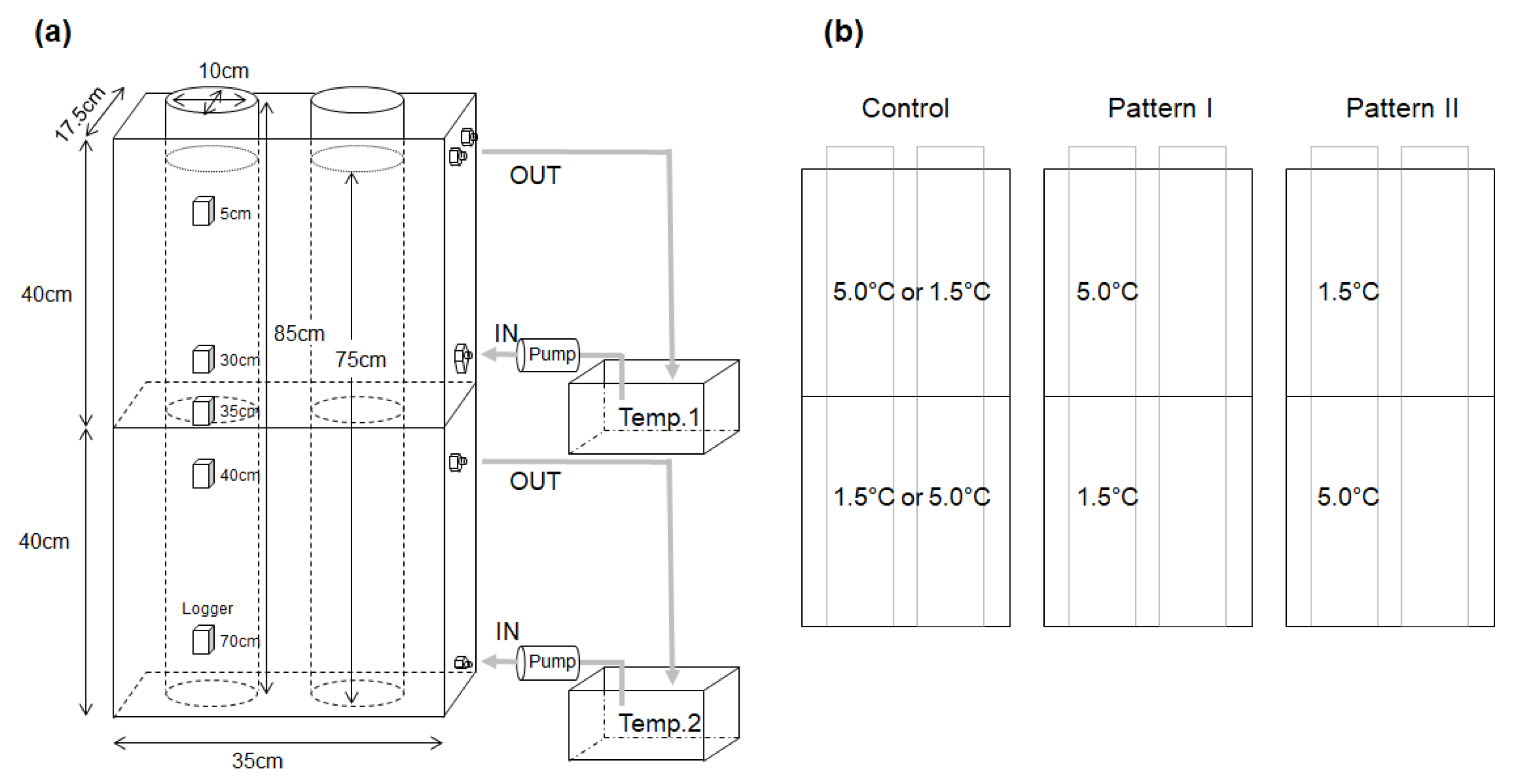
2.3. Egg and Larval Treatment
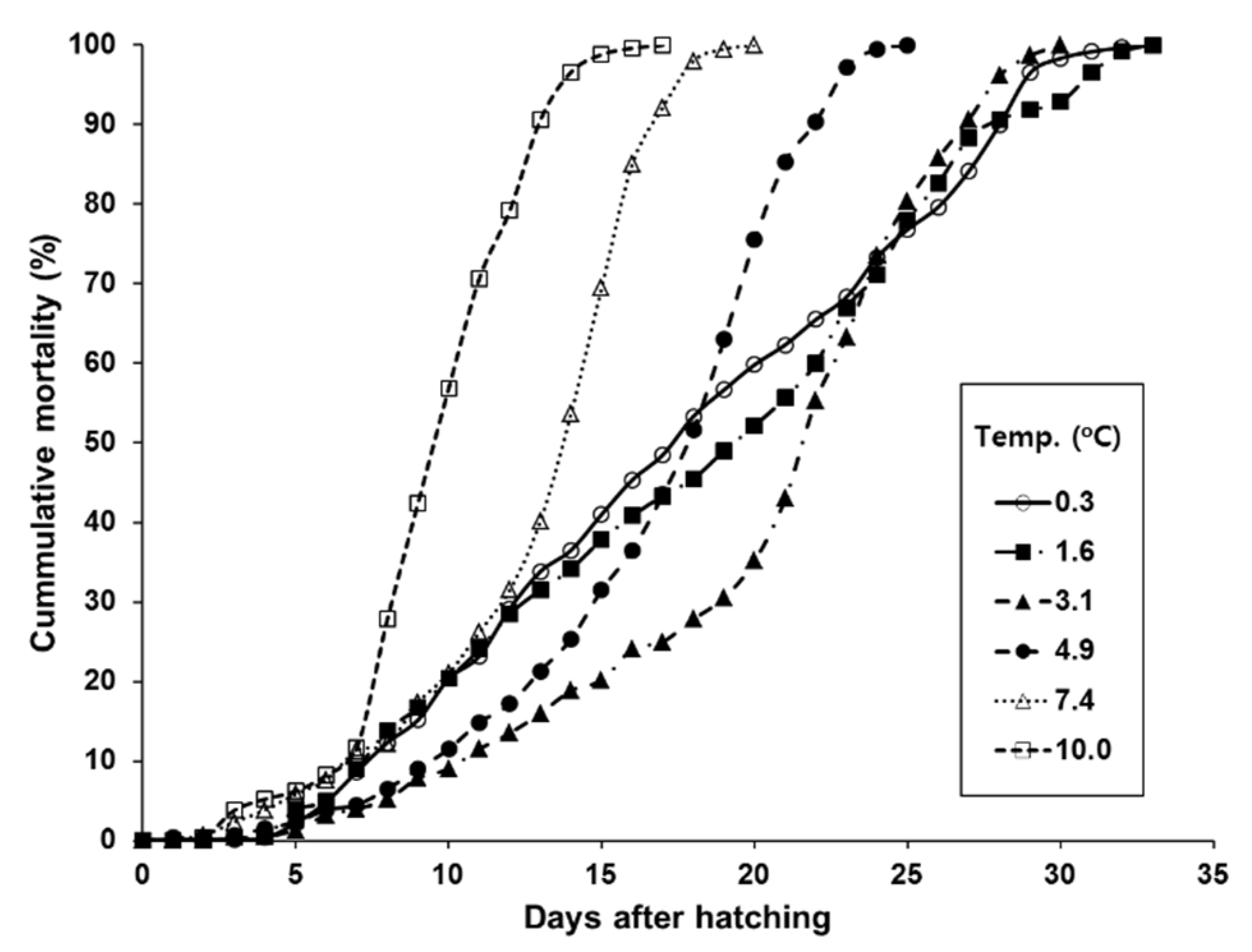
2.4. Mortality of Larvae at Different Temperatures
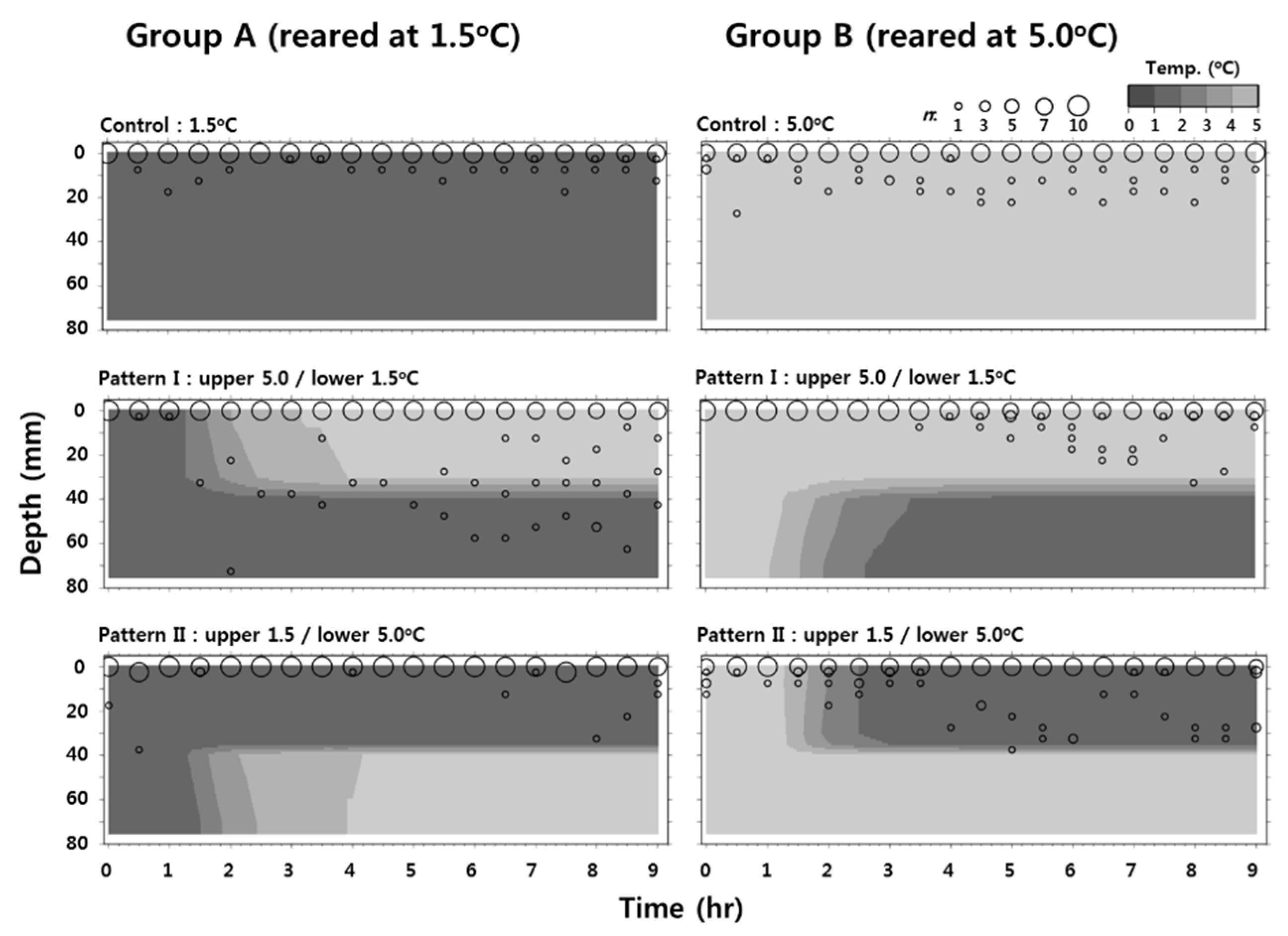
2.5. Behavioral Response of Larvae to Low Temperature
2.6. Statistical Analysis
3. Results
3.1. Specific Density during Early Ontogeny
3.2. Mortality of Larvae at Different Temperatures
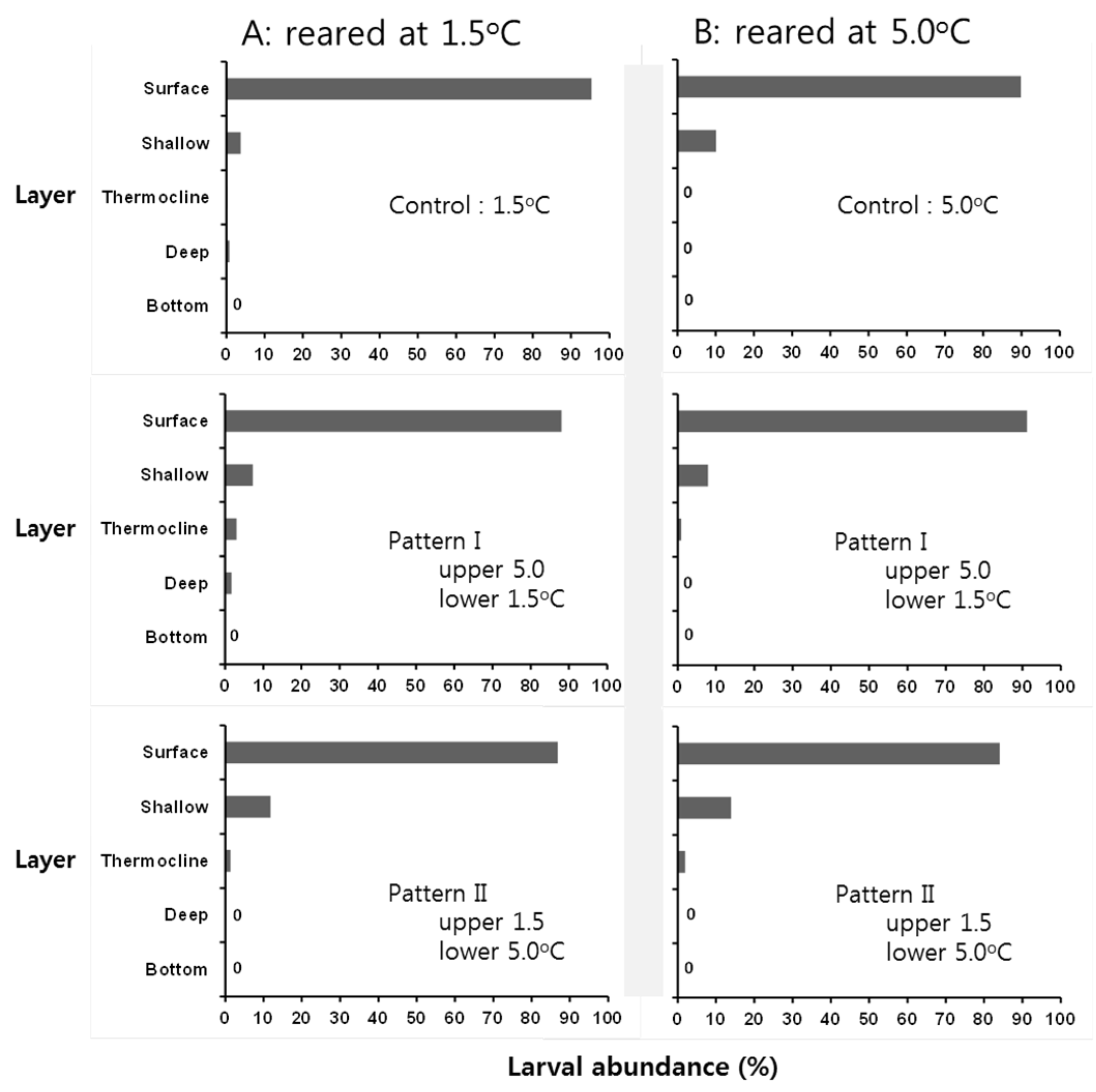
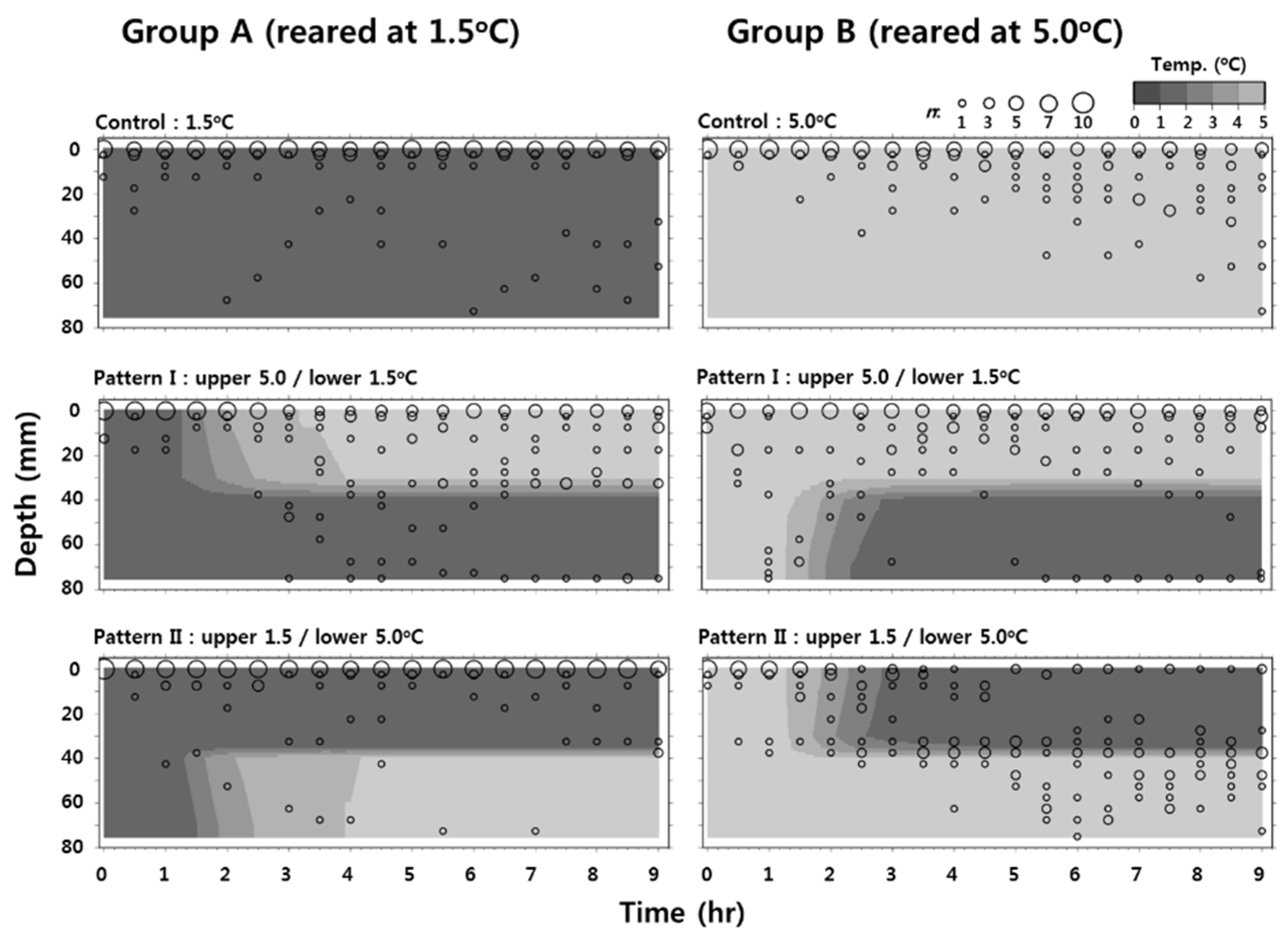
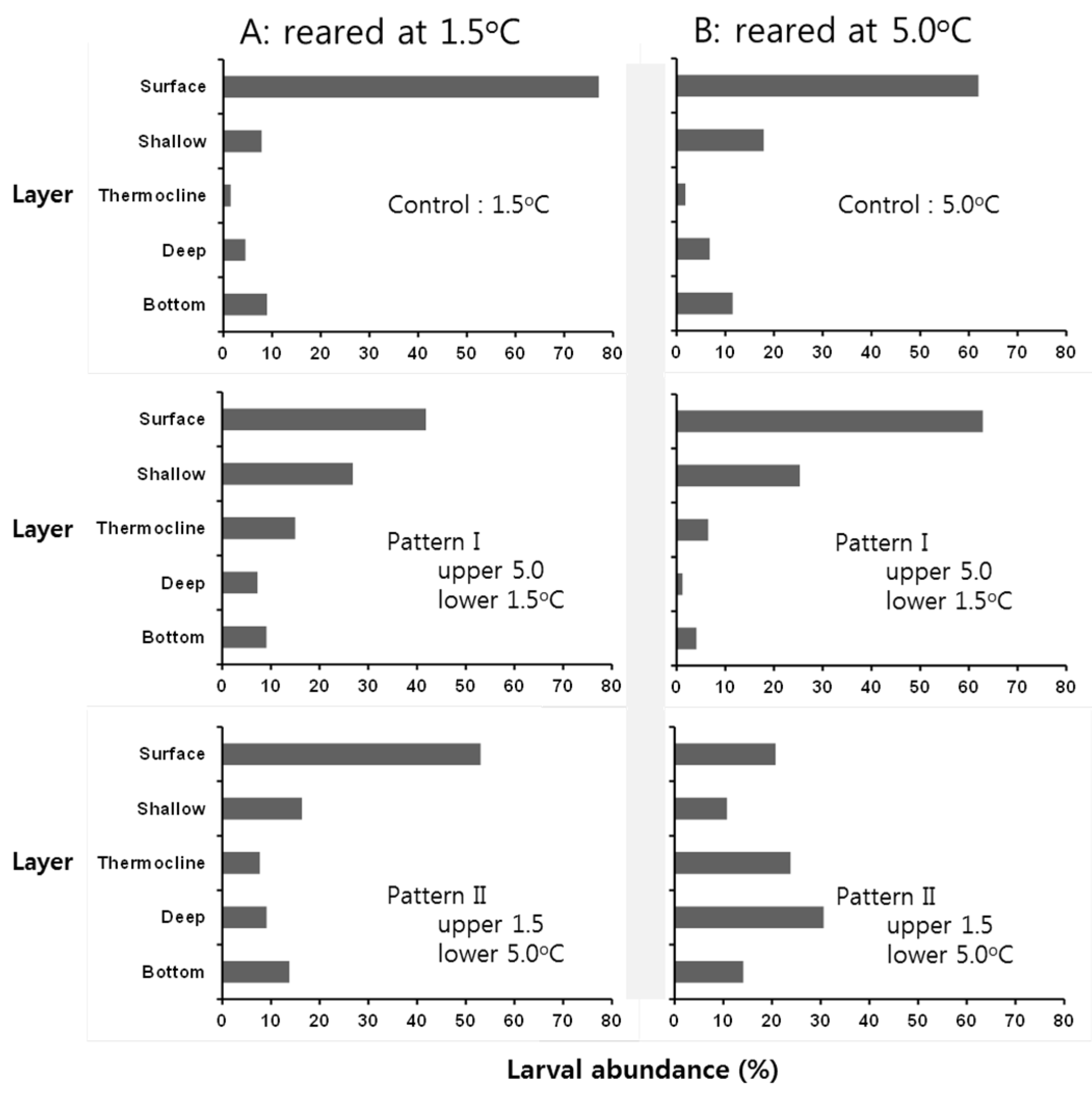
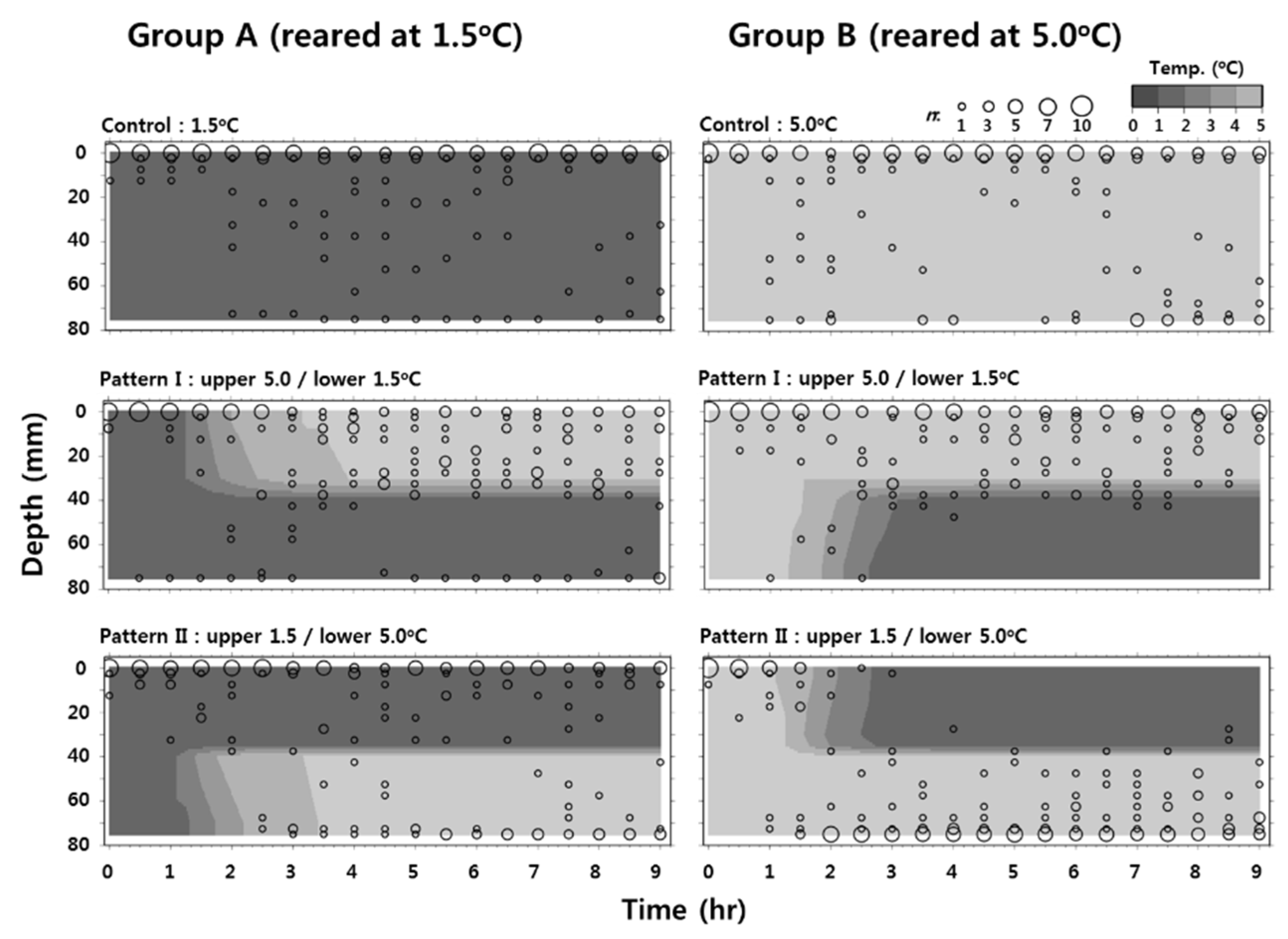
3.3. Behavioral Response of Larvae at Low Temperature
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tsuji, S. Alaska pollack population, Theragra chalcogramma, of Japan and its adjacent waters, I: Japanese fisheries and population studies. Mar. Behav. Physiol. 1989, 15, 147–205. [Google Scholar] [CrossRef]
- Springer, A.M. A review: Walleye pollock in the North Pacific? How much difference do they really make? Fish. Oceanogr. 1992, 1, 80–96. [Google Scholar] [CrossRef]
- Hamatsu, T.; Yabuki, K.; Watanabe, K. Decadal changes in reproduction of walleye pollock (Theragra chalcogramma) off the Pacific coast of northern Japan. Fish. Oceanogr. 2004, 13, 74–83. [Google Scholar] [CrossRef]
- Funamoto, T. Temperature-dependent stock-recruitment model for walleye pollock (Theragra chalcogramma) around northern Japan. Fish. Oceanogr. 2007, 16, 515–525. [Google Scholar] [CrossRef]
- Maeda, T.; Takahashi, T.; Ijichi, M.; Hirakawa, H.; Ueno, M. Ecological studies on the Alaska pollack in the adjacent waters of the Funka Bay, Hokkaido [Japan], 2: Spawning season. Bull. Jpn. Soc. Sci. Fish. 1976, 42, 1213–1222. [Google Scholar] [CrossRef]
- Nakatani, T. Studies on the early life history of walleye pollock Theragra chalcogramma in Funka bay and vicinity, Hokkaido [Japan]. Mem. Fac. Fish. Hokkaido Univ. 1988, 35, 1–46. [Google Scholar]
- Nakatani, T.; Maeda, T. Transport Process of the Alaska Pollack Eggs in Funka Bay and the Adjacent Waters, Hokkaido. Nippon Suisan Gakkaishi Jpn. Ed. 1981, 47, 1115–1118. [Google Scholar] [CrossRef] [Green Version]
- Shimizu, M.; Isoda, Y. The transport process of walleye pollock eggs into Funka bay [Hokkaido, Japan] in winter. Bull. Jpn. Soc. Fish. Oceanogr. 1997, 61, 134–143, (In Japanese with English Abstract). [Google Scholar]
- Blood, D.M.; Matarese, A.C.; Yoklavich, M.M. Embryonic development of walleye pollock, Theragra chalcogramma, from Shelikof Strait, Gulf of Alaska. Fish. Bull. 1994, 92, 207–222. [Google Scholar]
- Nakatani, T.; Maeda, T. Thermal Effect on the Development of Walleye Pollock Eggs and Their Upward speed to the Surface. Nippon Suisan Gakkaishi 1984, 50, 937–942. [Google Scholar] [CrossRef]
- Ohtani, K. Studies on the change of the hydrographic conditions in the Funka Bay II. Characteristic of the waters occupying the Funka Bay. Bull. Fac. Fish. Hokkaido Univ. 1971, 22, 58–66, (In Japanese with English Abstract). [Google Scholar]
- Ohtani, K. Water exchange of the Funka Bay. Bull. Coast. Oceanogr. 1979, 17, 50–59. (In Japanese) [Google Scholar]
- Isoda, Y.; Hasegawa, N. Heat budget of Funka Bay. Umi to Sora 1997, 72, 93–101, (In Japanese with English Abstract). [Google Scholar]
- Funamoto, T.; Yamamura, O.; Kono, T.; Hamatsu, T.; Nishimura, A. Abiotic and biotic factors affecting recruitment variability of walleye pollock (Theragra chalcogramma) off the Pacific coast of Hokkaido, Japan. Fish. Oceanogr. 2013, 22, 193–206. [Google Scholar] [CrossRef]
- Yoo, H.K.; Yamamoto, J.; Saito, T.; Sakurai, Y. Laboratory observations on the vertical swimming behavior of Japanese common squid Todarodes pacificus paralarvae as they ascend into warm surface waters. Fish. Sci. 2014, 80, 925–932. [Google Scholar] [CrossRef]
- Coombs, S.H. A density-gradient column for determining the specific gravity of fish eggs, with particular reference to eggs of the mackerel Scomber scombrus. Mar. Biol. 1981, 63, 101–106. [Google Scholar] [CrossRef]
- Kendall, A.W.; Kim, S. Buoyancy of walleye pollock (Theragra chalcogramma) eggs in relation to water properties and movement in Shelikof Strait, Gulf of Alaska. Eff. Ocean Var. Recruit. Eval. Parameters Used Stock Assess. Model. 1989, 108, 169–180. [Google Scholar]
- Ryan, T.H. Significance tests for multiple comparison of proportions, variance, and other statistics. Psychol. Bull. 1960, 57, 318. [Google Scholar] [CrossRef]
- Craik, J.C.A.; Harvey, S.M. The causes of buoyancy in eggs of marine teleosts. J. Mar. Biol. Ass. UK 1987, 67, 169–182. [Google Scholar] [CrossRef]
- Yamashita, Y.; Bailey, K.M. A laboratory study of the bioenergetics of larval walleye pollock, Theragra chalcogramma. Fish. Bull. 1989, 87, 525–536. [Google Scholar]
- Scholander, P.F.; Van Dam, L.; Kanwisher, J.W.; Hammel, H.T.; Gordon, M.S. Supercooling and osmoregulation in Arctic fish. J. Cell. Comp. Physiol. 1957, 49, 5–24. [Google Scholar] [CrossRef]
- Gordon, M.S.; Amdus, B.H.; Scholander, P.F. Freezing resistance in some northern fishes. Biol. Bull. 1962, 122, 52–62. [Google Scholar] [CrossRef]
- DeVries, A.L.; Wohlschlag, D.E. Freezing resistance in some Antarctic fishes. Science 1969, 163, 1073–1075. [Google Scholar] [CrossRef]
- DeVries, A.L.; Komatsu, S.K.; Feeney, R.E. Chemical and physical properties of freezing point-depressing glycoproteins from Antarctic fishes. J. Biol. Chem. 1970, 245, 2901–2908. [Google Scholar] [CrossRef]
- Osuga, D.T.; Feeney, R.E. Antifreeze glycoproteins from arctic fish. J. Biol. Chem. 1978, 253, 5338–5343. [Google Scholar] [CrossRef]
- Harding, M.M.; Anderberg, P.I.; Haymet, A.D.J. “Antifreeze” glycoproteins from polar fish. Eur. J. Biochem. 2003, 270, 1381–1392. [Google Scholar] [CrossRef]
- Stevenson, D.E.; Lauth, R.R. Latitudinal trends and temporal shifts in the catch composition of bottom trawls conducted on the eastern Bering Sea shelf. Deep Sea Res. Part II 2012, 65–70, 251–259. [Google Scholar] [CrossRef]
- Olla, B.L.; Davis, M.W. Effects of physical factors on the vertical distribution of larval walleye Pollock Theragra chalcogramma under controlled laboratory conditions. Mar. Ecol. Prog. Ser. 1990, 63, 10–112. [Google Scholar] [CrossRef]
- Funamoto, T.; Yamamura, O.; Shida, O.; Itaya, K.; Mori, K.; Hiyama, Y.; Sakurai, Y. Comparison of factors affecting recruitment variability of walleye pollock Theragra chalcogramma in the Pacific Ocean and the Sea of Japan off northern Japan. Fish. Sci. 2014, 80, 117–126. [Google Scholar] [CrossRef]
- Porter, S.M. Effects of size and light on respiration and activity of walleye pollock (Theragra chalcogramma) larvae. J. Exp. Mar. Bio. Ecol. 2001, 256, 253–265. [Google Scholar] [CrossRef]
- Paul, A.J. Light, temperature, nauplii concentrations, and prey capture by first feeding pollock larvae Theragra chalcogramma. Mar. Ecol. Prog. Ser. 1983, 13, 175–179. [Google Scholar] [CrossRef]
- Bailey, K.M.; Stehr, C.L. Laboratory studies on the early life history of the walleye pollock, Theragra chalcogramma (Pallas). J. Exp. Mar. Bio. Ecol. 1986, 99, 233–246. [Google Scholar] [CrossRef]
- Yokota, T.; Nakagawa, T.; Murakami, N.; Chimura, M.; Tanaka, H.; Yamashita, Y.; Funamoto, T. Effects of starvation at the first feeding stage on the survival and growth of walleye Pollock Gadus chalcogrammus larvae. Fish. Sci. 2016, 82, 73–83. [Google Scholar] [CrossRef]
- Kuroda, H.; Takahashi, D.; Mitsudera, H.; Azumaya, T.; Setou, T. A preliminary study to understand the transport process for the eggs and larvae of Japanese Pacific walleye pollock Theragra chalcogramma using particle-tracking experiments based on a high-resolution ocean model. Fish. Sci. 2014, 80, 127–138. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Batch | Mean | SD | Range |
|---|---|---|---|
| 1 | 1.356 | 0.014 | 1.331–1.386 |
| 2 | 1.406 | 0.010 | 1.385–1.429 |
| 3 | 1.414 | 0.009 | 1.398–1.432 |
| 4 | 1.425 | 0.012 | 1.403–1.451 |
| Batch | Mean | SD | Range |
|---|---|---|---|
| 1 | 23.058 | 0.084 | 22.909–23.195 |
| 2 | 22.995 | 0.053 | 22.924–23.103 |
| 3 | 22.870 | 0.089 | 22.719–23.032 |
| 4 | 22.549 | 0.077 | 22.403–22.637 |
| Batch | Mean | SD | Range |
|---|---|---|---|
| 1 | 23.121 | 0.983 ab | 21.135–24.493 |
| 2 | 23.268 | 0.439 a | 22.623–24.289 |
| 3 | 22.935 | 0.965 abc | 21.739–24.614 |
| 4 | 22.353 | 1.006 c | 20.6791–24.162 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yoo, H.-K.; Kim, W.-J.; Lim, H.-J.; Byun, S.-G.; Yamamoto, J.; Sakurai, Y. The Effect of Low Temperature on the Early Life Stages of the Walleye Pollock, Gadus chalcogrammus—A Laboratory Study. J. Mar. Sci. Eng. 2021, 9, 818. https://doi.org/10.3390/jmse9080818
Yoo H-K, Kim W-J, Lim H-J, Byun S-G, Yamamoto J, Sakurai Y. The Effect of Low Temperature on the Early Life Stages of the Walleye Pollock, Gadus chalcogrammus—A Laboratory Study. Journal of Marine Science and Engineering. 2021; 9(8):818. https://doi.org/10.3390/jmse9080818
Chicago/Turabian StyleYoo, Hae-Kyun, Woo-Jin Kim, Hyung-Jun Lim, Soon-Gyu Byun, Jun Yamamoto, and Yasunori Sakurai. 2021. "The Effect of Low Temperature on the Early Life Stages of the Walleye Pollock, Gadus chalcogrammus—A Laboratory Study" Journal of Marine Science and Engineering 9, no. 8: 818. https://doi.org/10.3390/jmse9080818