
Effects of Ecological Restoration Using Non-Native Mangrove Kandelia obovata to Replace Invasive Spartina alterniflora on Intertidal Macrobenthos Community in Maoyan Island (Zhejiang, China)

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Macrobenthos Sampling and Laboratory Procedures
2.3. Data Analyses
3. Results
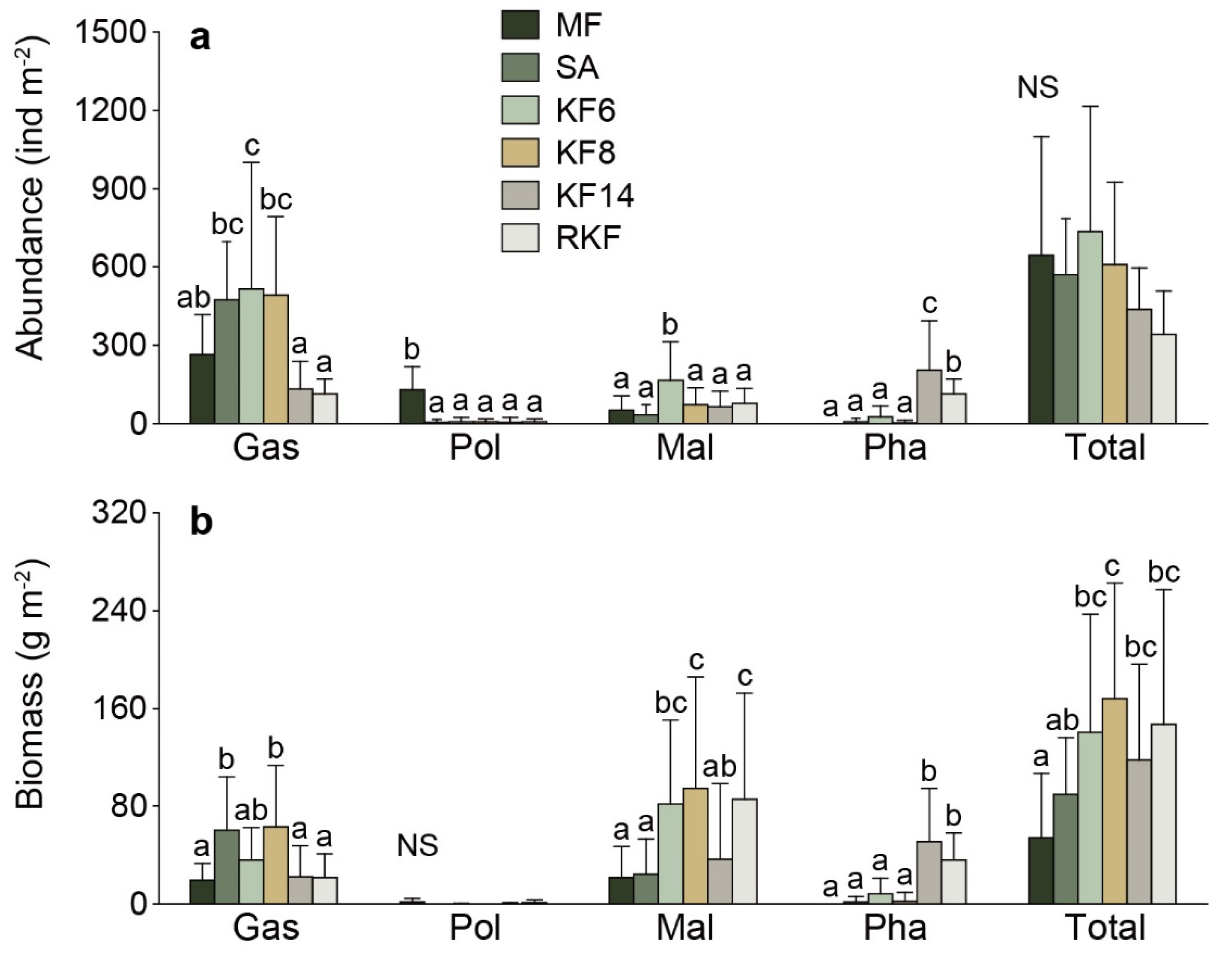
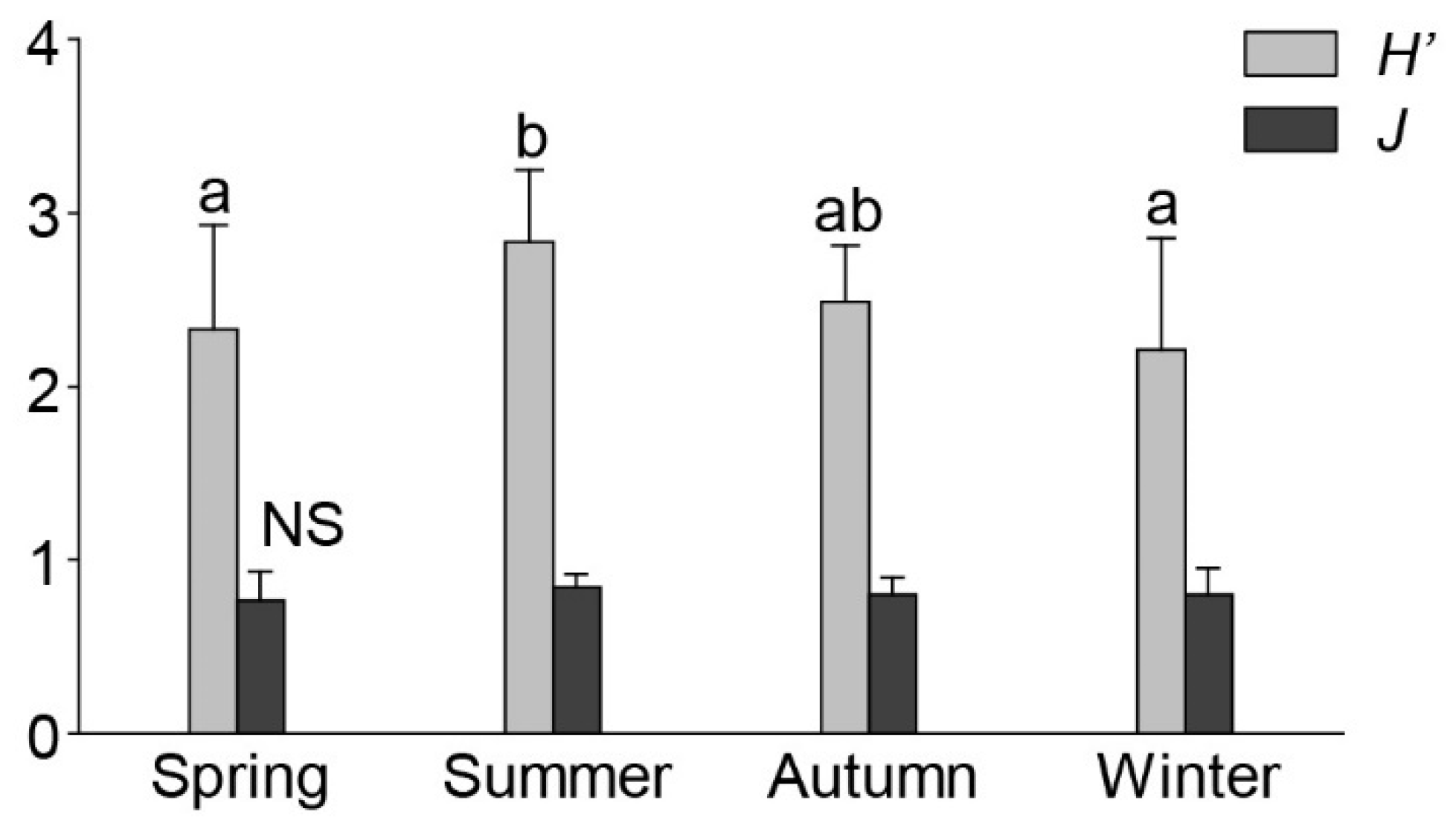
3.1. Species Composition and Biodiversity
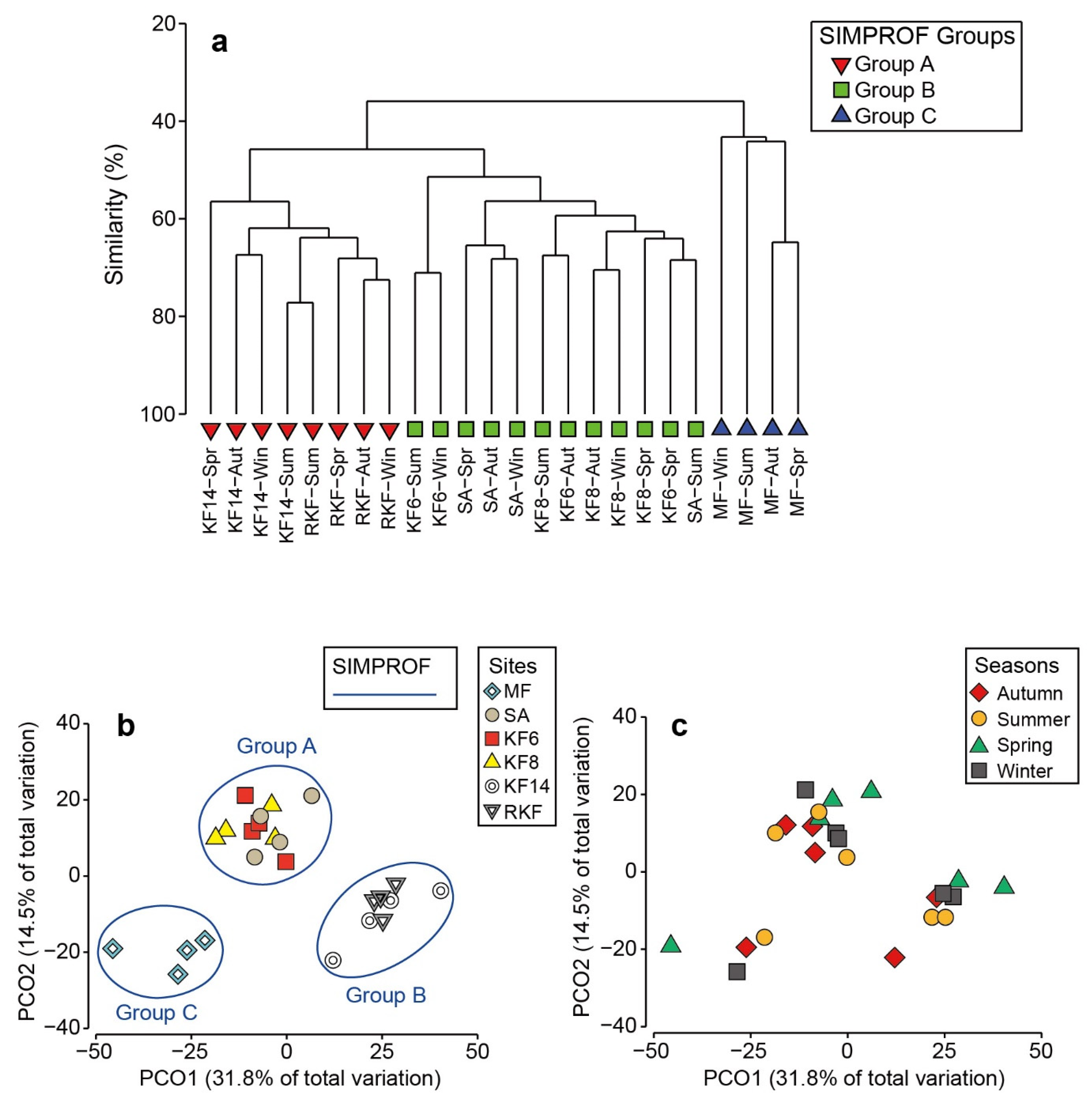
3.2. Macrobenthos Community Structure
3.3. AMBI and M-AMBI
4. Discussion
4.1. Seasonality in Macrobenthos Community
4.2. Influence of Non-Indigenous Mangrove and Spartina on Macrobenthos Communities
4.3. Influence of Mangrove Restoration on Macrobenthos Communities
4.4. Assessment of the Ecological Effectiveness of Replacement Control
4.5. Assessment of Benthic Ecological Status
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Simberloff, D.; Martin, J.L.; Genovesi, P.; Maris, V.; Wardle, D.A.; Aronson, J.; Courchamp, F.; Galil, B.; García-Berthou, E.; Pascal, M.; et al. Impacts of Biological Invasions: What’s What and the Way Forward. Trends Ecol. Evol. 2013, 28, 58–66. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.Z. Invasive Plants in Coastal Wetlands: Patterns and Mechanisms. In Wetlands: Ecosystem Services, Restoration and Wise Use; An, S., Verhoeven, A., Eds.; Ecological Studies; Springer International Publishing: Cham, Switzerland, 2019; Volume 238, pp. 97–128. [Google Scholar]
- Lee, S.Y.; Khim, J. Hard Science Is Essential to Restoring Soft-Sediment Intertidal Habitats in Burgeoning East Asia. Chemosphere 2017, 168, 765–776. [Google Scholar] [CrossRef] [PubMed]
- Wan, S.; Qin, P.; Liu, J.; Zhou, H. The Positive and Negative Effects of Exotic Spartina alterniflora in China. Ecol. Eng. 2009, 35, 444–452. [Google Scholar] [CrossRef]
- Levin, L.A.; Neira, C.; Grosholz, E.D. Invasive Cordgrass Modifies Wetland Trophic Function. Ecology 2006, 87, 419–432. [Google Scholar] [CrossRef]
- Neira, C.; Levin, L.A.; Grosholz, E.D. Benthic Macrofaunal Communities of Three Sites in San Francisco Bay Invaded by Hybrid Spartina, with Comparison to Uninvaded Habitats. Mar. Ecol. Prog. 2005, 292, 111–126. [Google Scholar] [CrossRef]
- Qiao, P.Y.; Wang, A.D.; Xie, B.H.; Wang, L.; Han, G.X.; Zhang, X.T. Effects of herbicides on invasive Spartina alterniflora in the Yellow River Delta. Acta Ecol. Sin. 2019, 39, 5627–5634. [Google Scholar]
- Gu, Y.F. Technologies and Their Effects on Ecological Control of Spartina alterniflora in Chongming Dongtan. J. Shanghai Jiaotong Univ. 2019, 037, 83–88. [Google Scholar]
- Zhao, X.J.; Liu, X.Y.; Gong, L.; Deng, Z.Z.; Bai, J.D.; Li, J.S.; Zhao, C.Y. Control of Spartina alterniflora by Integrated Technique of Mowing plus Shading. Chin. J. Ecol. 2014, 33, 2714–2719. [Google Scholar]
- Chen, B.B.; Liu, C.E.; Yuan, Y. Harm and Control Measures of Spartina alterniflora Based on Bibliometrics. Acta Agric. Shanghai 2020, 36, 90–95. [Google Scholar]
- An, S.Q.; Gu, B.H.; Zhou, C.F.; Wang, Z.S.; Deng, Z.F.; Zhi, Y.B.; Li, H.L.; Chen, L.; Yu, D.H.; Liu, Y.H. Spartina Invasion in China: Implications for Invasive Species Management and Future Research. Weed Res. 2007, 47, 183–191. [Google Scholar] [CrossRef]
- Grevstad, F.S.; Wecker, M.S.; Strong, D.R. Biological Control of Spartina. In Proceedings of the Third International Conference on Invasive Spartina, San Francisco, CA, USA, 8–9 November 2004. Chapter 4: Spartina Control and Management. [Google Scholar]
- Zhou, T.; Liu, S.; Feng, Z.; Liu, G.; Gan, Q.; Peng, S. Use of Exotic Plants to Control Spartina alterniflora Invasion and Promote Mangrove Restoration. Sci. Rep. 2015, 5, 12980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, J.X.; Guo, J.; Huang, Q.; Jiang, J.; Huang, G.; Yang, Z.; Lin, G. Changes in the Community Structure and Diet of Benthic Macrofauna in Invasive Spartina alterniflora Wetlands Following Restoration with Native Mangroves. Wetlands 2014, 34, 673–683. [Google Scholar] [CrossRef]
- Feng, J.X.; Huang, Q.; Chen, H.; Guo, J.; Lin, G. Restoration of Native Mangrove Wetlands Can Reverse Diet Shifts of Benthic Macrofauna Caused by Invasive Cordgrass. J. Appl. Ecol. 2018, 55, 905–916. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.Z.; Wang, W.; Zhang, Y.; Lin, G. Recent Progresses in Mangrove Conservation, Restoration and Research in China. J. Plant Ecol. 2009, 2, 45–54. [Google Scholar] [CrossRef]
- Li, M.S.; Lee, S.Y. Mangroves of China: A Brief Review. For. Ecol. Manag. 1997, 96, 241–259. [Google Scholar] [CrossRef]
- Huang, X.L.; Peng, X.; Qiu, J.B.; Chen, S.B. Mangrove Status and Development Prospects in Southern Zhejiang Province. J. Zhejiang For. Coll. 2009, 26, 427–433. [Google Scholar]
- Chen, S.B.; Lu, C.Y. Northward Introduction Ecology of Mangrove Plants in Response to Climate Change; China Ocean Press: Beijing, China, 2012. [Google Scholar]
- Kristensen, E. Mangrove Crabs as Ecosystem Engineers; with Emphasis on Sediment Processes. J. Sea Res. 2008, 59, 30–43. [Google Scholar] [CrossRef]
- Lee, S.Y. Mangrove Macrobenthos: Assemblages, Services, and Linkages. J. Sea Res. 2008, 59, 16–29. [Google Scholar] [CrossRef]
- Zhu, Z.C.; Cozzoli, F.; Chu, N.Y.; Salvador, M.; Ysebaert, T.; Zhang, L.Q.; Herman, M.J.; Bouma, T.J. Interactive Effects between Physical Forces and Ecosystem Engineers on Seed Burial: A Case Study Using Spartina anglica. Oikos 2016, 125, 98–106. [Google Scholar] [CrossRef] [Green Version]
- Macintosh, D.J.; Ashton, E.C.; Havanon, S. Mangrove Rehabilitation and Intertidal Biodiversity: A Study in the Ranong Mangrove Ecosystem, Thailand. Estuar. Coast. Shelf Sci. 2002, 55, 331–345. [Google Scholar] [CrossRef] [Green Version]
- Borja, A.; Franco, J.; Perez, V. A Marine Biotic Index to Establish the Ecological Quality of Soft-Bottom Benthos within European Estuarine and Coastal Environments. Mar. Pollut. Bull. 2000, 40, 1100–1114. [Google Scholar] [CrossRef]
- Hu, C.Y.; Shui, Y.Y.; Tian, K.; Zhang, C.C.; Yi, M.M.; Shui, B.N. Community Diversity of Macrobenthos in Different Habitats of Shupaisha Wetland in Oujiang River Estuary. Oceanol. Limnol. Sin. 2016, 47, 133–139. [Google Scholar]
- Lin, Q.L.; Gu, X.X.; Guo, X.D.; Cai, L.Z.; Lin, L. Coastal Wetland Restoration Based on Mangrove Kandelia Obovata Substitution Controlling Spartina Alterniflora Invasion: A Case Study of Aojiang Estuary in Zhejiang Province. Chin. J. Ecol. 2020, 39, 1761–1768. [Google Scholar]
- Yan, J.; Sui, J.; Xu, Y.; Li, X.; Wang, H.; Zhang, B. Major Environmental Variables Related to North-South Differences in the Macrozoobenthic Community in Adjacent Sea Areas of the Yangtze River Estuary, China. Ecol. Indic. 2019, 105, 70–81. [Google Scholar] [CrossRef]
- Anderson, M.J.; Gorley, R.N.; Clarke, K.R. Guide to Software and Statistical Methods. 2016. Available online: http://updates.primer-e.com/primer7/manuals/PERMANOVA+_manual.pdf (accessed on 21 July 2021).
- Clarke, R.; Gorley, R. PRIMER Version 7: User Manual/Tutorial 2015. Available online: http://updates.primer-e.com/primer7/manuals/User_manual_v7a.pdf (accessed on 21 July 2021).
- Borja, A.; Tunberg, B.G. Assessing Benthic Health in Stressed Subtropical Estuaries, Eastern Florida, USA Using AMBI and M-AMBI. Ecol. Indic. 2011, 11, 295–303. [Google Scholar] [CrossRef]
- Cai, W.; Meng, W.; Zhu, Y.; Zhou, J.; Liu, L. Assessing Benthic Ecological Status in Stressed Liaodong Bay (China) with AMBI and M-AMBI. Chin. J. Ocean. Limnol. 2013, 31, 482–492. [Google Scholar] [CrossRef]
- Luo, X.; Zhang, J.; Yang, J.; Song, W.; Cui, W. Assessing the Benthic Habitat Quality in the Huanghe (Yellow River) Estuary and Its Adjacent Areas Using AMBI and M-AMBI. Acta Oceanol. Sin. 2014, 33, 117–124. [Google Scholar] [CrossRef]
- Satheeshkumar, P.; Khan, A.B. Influence of Environmental Parameters on the Distribution and Diversity of Molluscan Composition in Pondicherry Mangroves, Southeast Coast of India. Ocean Sci. J. 2012, 47, 61–71. [Google Scholar] [CrossRef] [Green Version]
- Hajializadeh, P.; Safaie, M.; Naderloo, R.; Shojaei, M.G.; Gammal, J.; Villnäs, A.; Norkko, A. Species Composition and Functional Traits of Macrofauna in Different Mangrove Habitats in the Persian Gulf. Front. Mar. Sci. 2020, 7, 575480. [Google Scholar] [CrossRef]
- Lana, C.; Guiss, C. Influence of Spartina Alterniflora on Structure and Temporal Variability of Macrobenthic Associations in a Tidal Flat of Paranagua Bay (Southeastern Brazil). Mar. Ecol. Prog. Ser. 1991, 73, 231–244. [Google Scholar] [CrossRef]
- Sueiro, M.C.; Bortolus, A.; Schwindt, E. The Role of the Physical Structure of Spartina densiflora Brong. in Structuring Macroinvertebrate Assemblages. Aquat. Ecol. 2012, 46, 25–36. [Google Scholar] [CrossRef]
- Tang, M.; Kristensen, E. Associations between Macrobenthos and Invasive Cordgrass, Spartina anglica, in the Danish Wadden Sea. Helgol. Mar. Res. 2010, 64, 321–329. [Google Scholar] [CrossRef]
- Chen, Q.; Jian, S.; Ma, K.; Chen, P. Differences in Macrobenthic Faunal Communities in Mangrove Wetland Habitats (Zhanjiang, China) Invaded and Non-Invaded by Exotic Cordgrass Spartina alterniflora. Ecol. Res. 2018, 33, 1113–1123. [Google Scholar] [CrossRef]
- Braga, C.F.; Beasley, C.R.; Isaac, V.J. Effects of Plant Cover on the Macrofauna of Spartina Marshes in Northern Brazil. Braz. Arch. Biol. Technol. 2009, 52, 1409–1420. [Google Scholar] [CrossRef]
- Lana, P.C.; Couto, E.C.G.; Almeida, M.V.O. Polychaete Distribution and Abundance in Intertidal Flats of Paranaguá Bay (SE Brazil). Bull. Mar. Sci. Miami 1997, 60, 433–442. [Google Scholar]
- Metcalfe, K.N.; Glasby, C.J. Diversity of Polychaeta (Annelida) and Other Worm Taxa in Mangrove Habitats of Darwin Harbour, Northern Australia. J. Sea Res. 2008, 59, 70–82. [Google Scholar] [CrossRef]
- Koo, B.J.; Kwon, K.K.; Hyun, J.-H. The Sediment-Water Interface Increment Due to the Complex Burrows of Macrofauna in a Tidal Flat. Ocean Sci. J. 2005, 40, 221–227. [Google Scholar] [CrossRef]
- Wang, M.; Gao, X.; Wang, W. Differences in Burrow Morphology of Crabs between Spartina alterniflora Marsh and Mangrove Habitats. Ecol. Eng. 2014, 69, 213–219. [Google Scholar] [CrossRef]
- Morrisey, D.J.; Skilleter, G.A.; Ellis, J.I.; Burns, B.R.; Kemp, C.E.; Burt, K. Differences in Benthic Fauna and Sediment among Mangrove (Avicennia marina Var. Australasica) Stands of Different Ages in New Zealand. Estuar. Coast. Shelf Sci. 2003, 56, 581–592. [Google Scholar] [CrossRef]
- Dunn, G. Chloride and Osmotic Balance in the Euryhaline Sipunculid Phascolosoma Arcuatum from a Malaysian Mangrove Swamp. Biol. Bull. 1976, 150, 211–221. [Google Scholar]
- Ip, Y.K.; Tan, G.Q.; Kuah, S.S.L.; Chew, S.F. Detoxification of Environmental Sulfide to Sulfane Sulfur in the Intertidal Sipunculid Phascolosoma Arcuatum. J. Comp. Physiol. B Biochem. Syst. Environ. Physiol. 1997, 167, 213–220. [Google Scholar] [CrossRef]
- Liao, Y.; Shou, L.; Tang, Y.; Gao, A.; Chen, Q.; Yan, X.; Chen, J. Influence of Two Non-Indigenous Plants on Intertidal Macrobenthic Communities in Ximen Island Special Marine Protected Area, China. Ecol. Eng. 2018, 112, 96–104. [Google Scholar] [CrossRef]
- Chen, Q. Changes in the Macrobenthic Faunal Community during Succession of a Mangrove Forest at Zhanjiang, South China. J. Coast. Res. 2015, 31, 315–325. [Google Scholar] [CrossRef]
- Li, Y.F.; Du, F.Y.; Gu, Y.G.; Ning, J.J.; Wang, L.G. Changes of the Macrobenthic Faunal Community with Stand Age of a Non-Native Mangrove Species in Futian Mangrove National Nature Reserve, Guangdong, China. Zool. Stud. 2017, 56, e19. [Google Scholar] [PubMed]
- Kon, K.; Kurokura, H.; Tongnunui, P. Effects of the Physical Structure of Mangrove Vegetation on a Benthic Faunal Community. J. Exp. Mar. Biol. Ecol. 2010, 383, 171–180. [Google Scholar] [CrossRef]
- Liao, Y.; Shou, L.; Tang, Y.; Zeng, J.; Chen, Q.; Yan, X. Effects of Non-Indigenous Plants on Food Sources of Intertidal Macrobenthos in Yueqing Bay, China: Combining Stable Isotope and Fatty Acid Analyses. Estuar. Coast. Shelf Sci. 2020, 241, 106801. [Google Scholar] [CrossRef]
- Kon, K.; Kurokura, H.; Hayashizaki, K. Role of Microhabitats in Food Webs of Benthic Communities in a Mangrove Forest. Mar. Ecol. Prog. Ser. 2007, 340, 55–62. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.C.; Ye, Y. Restoration of Aegiceras Corniculatum Mangroves in Jiulongjiang Estuary Changed Macro-Benthic Faunal Community. Ecol. Eng. 2011, 37, 224–228. [Google Scholar] [CrossRef]
- Chen, G.C.; Ye, Y.; Lu, C. Changes of Macro-Benthic Faunal Community with Stand Age of Rehabilitated Kandelia Candel Mangrove in Jiulongjiang Estuary, China. Ecol. Eng. 2007, 31, 215–224. [Google Scholar] [CrossRef]
- Nobbs, M. Effects of Vegetation Differ among Three Species of Fiddler Crabs (Uca spp.). J. Exp. Mar. Biol. Ecol. 2003, 284, 41–50. [Google Scholar] [CrossRef]
- Xiang, H.; Li, K.; Cao, L.; Zhang, Z.; Yang, H. Impacts of Pollution, Sex, and Tide on the Time Allocations to Behaviours of Uca Arcuata in Mangroves. Sci. Total Environ. 2020, 742, 140609. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.C.; Gao, M.; Pang, B.; Chen, S.; Ye, Y. Top-Meter Soil Organic Carbon Stocks and Sources in Restored Mangrove Forests of Different Ages. For. Ecol. Manag. 2018, 422, 87–94. [Google Scholar] [CrossRef]
- Feng, J.X.; Zhou, J.; Wang, L.; Cui, X.; Ning, C.; Wu, H.; Zhu, X.; Lin, G.H. Effects of Short-Term Invasion of Spartina alterniflora and the Subsequent Restoration of Native Mangroves on the Soil Organic Carbon, Nitrogen and Phosphorus Stock. Chemosphere 2017, 184, 774–783. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Liao, B.; Liu, B.; Peng, C.; Zhang, Y.; Guan, W.; Zhu, Q.; Yang, G. Eradicating Invasive Spartina alterniflora with Alien Sonneratia apetala and Its Implications for Invasion Controls. Ecol. Eng. 2014, 73, 367–372. [Google Scholar] [CrossRef]
- Ashton, E.C.; Macintosh, D.J.; Hogarth, P.J. A Baseline Study of the Diversity and Community Ecology of Crab and Molluscan Macrofauna in the Sematan Mangrove Forest, Sarawak, Malaysia. J. Trop. Ecol. 2003, 19, 127–142. [Google Scholar] [CrossRef]
- Hu, C.Y.; Shui, B.N.; Li, W.; Yang, X.; Zhang, X. Assessing the Ecological Quality Status of Transplanted Mangrove Wetland in the Oujiang Estuary, China. Mar. Pollut. Bull. 2018, 133, 1–8. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | MF | SA | KF6 | KF8 | KF14 | RKF | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| N | W | IRI | N | W | IRI | N | W | IRI | N | W | IRI | N | W | IRI | N | W | IRI | |
| Cl | 0.8 | 2.9 | 31.3 | 8.4 | 17.3 | 714.8 | 5.6 | 1.4 | 155.1 | 14.0 | 13.0 | 825.9 | 5.8 | 7.2 | 323.7 | 7.0 | 5.4 | 241.3 |
| Ml | 9.5 | 0.8 | 258.4 | 27.1 | 2.2 | 896.4 | 4.2 | 0.3 | 87.9 | 10.3 | 0.3 | 265.3 | 1.5 | 0.1 | 22.1 | 1.6 | 0.0 | 13.2 |
| Ob | 23.3 | 19.9 | 841.2 | 26.6 | 11.5 | 1271.5 | 36.3 | 3.9 | 1340.7 | 35.0 | 9.2 | 1349.4 | 14.9 | 4.3 | 479.9 | 10.5 | 1.4 | 265.8 |
| Ta | 1.4 | 17.1 | 308.6 | 0.5 | 7.7 | 45.5 | 6.2 | 28.9 | 876.9 | 6.3 | 33.8 | 668.5 | 3.6 | 12.5 | 224.0 | 6.6 | 23.4 | 750.2 |
| Pa | 0 | 0 | 0 | 1.2 | 1.8 | 25.1 | 3.4 | 5.9 | 129.2 | 0.4 | 1.3 | 4.9 | 46.5 | 42.9 | 2733.3 | 33.6 | 24.5 | 1775.8 |
| (a) | ||||||
| Source | df | SS | MS | Pseudo-F | P (perm) | U. perms |
| Sites | 5 | 50,824 | 10,165 | 5.7172 | 0.001 | 999 |
| Seasons | 3 | 10,396 | 3465.5 | 1.9492 | 0.001 | 998 |
| Sites × Seasons | 15 | 28,458 | 1897.2 | 1.0671 | 0.302 | 998 |
| Res | 48 | 85,341 | 1777.9 | |||
| Total | 71 | 1.75 × 105 | ||||
| (b) | ||||||
| Seasons | Spring | Summer | Autumn | Winter | ||
| Spring | 28.38% | 0.001 | 0.004 | 0.046 | ||
| Summer | 68.62% | 37.74% | 0.022 | 0.091 | ||
| Autumn | 69.87% | 64.38% | 35.16% | 0.428 | ||
| Winter | 73.11% | 68.48% | 68.64% | 27.14% | ||
| Values on the diagonal are percent similarity within season (PERMANOVA analysis); values below the diagonal are percent dissimilarity between seasons (PERMANOVA analysis). | ||||||
| (c) | ||||||
| Sites/ Habitats | MF | SA | KF6 | KF8 | KF14 | RKF |
| MF | 28.52% | 0.001 | 0.001 | 0.001 | 0.001 | 0.001 |
| SA | 73.05% | 46.30% | 0.001 | 0.037 | 0.002 | 0.001 |
| KF6 | 74.21% | 61.61% | 46.61% | 0.002 | 0.001 | 0.001 |
| KF8 | 73.62% | 58.11% | 62.78% | 41.56% | 0.001 | 0.001 |
| KF14 | 83.14% | 69.05% | 69.81% | 70.95% | 38.71% | 0.247 |
| RKF | 83.85% | 72.57% | 67.79% | 79.18% | 60.68% | 41.31% |
| Values on the diagonal are percentage of similarity within habitat (PERMANOVA analysis); values below the diagonal are percent dissimilarity between habitats (PERMANOVA analysis). MF: unvegetated mudflat, SA: S. alterniflora marsh, KF6: 6-year-old restored K. obovata forest, KF8: 8-year-old restored K. obovata forest, KF14: 14-year-old restored K. obovata forest, RKF: 11-year-old reference K. obovata forest. | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Q.; Duarte, C.; Song, L.; Christakos, G.; Agusti, S.; Wu, J. Effects of Ecological Restoration Using Non-Native Mangrove Kandelia obovata to Replace Invasive Spartina alterniflora on Intertidal Macrobenthos Community in Maoyan Island (Zhejiang, China). J. Mar. Sci. Eng. 2021, 9, 788. https://doi.org/10.3390/jmse9080788
Wang Q, Duarte C, Song L, Christakos G, Agusti S, Wu J. Effects of Ecological Restoration Using Non-Native Mangrove Kandelia obovata to Replace Invasive Spartina alterniflora on Intertidal Macrobenthos Community in Maoyan Island (Zhejiang, China). Journal of Marine Science and Engineering. 2021; 9(8):788. https://doi.org/10.3390/jmse9080788
Chicago/Turabian StyleWang, Qiuxuan, Carlos Duarte, Li Song, George Christakos, Susana Agusti, and Jiaping Wu. 2021. "Effects of Ecological Restoration Using Non-Native Mangrove Kandelia obovata to Replace Invasive Spartina alterniflora on Intertidal Macrobenthos Community in Maoyan Island (Zhejiang, China)" Journal of Marine Science and Engineering 9, no. 8: 788. https://doi.org/10.3390/jmse9080788