
Deep-Water Cartilaginous Fishes in the Central Mediterranean Sea: Comparison between Geographic Areas with Two Low Impact Tools for Sampling

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
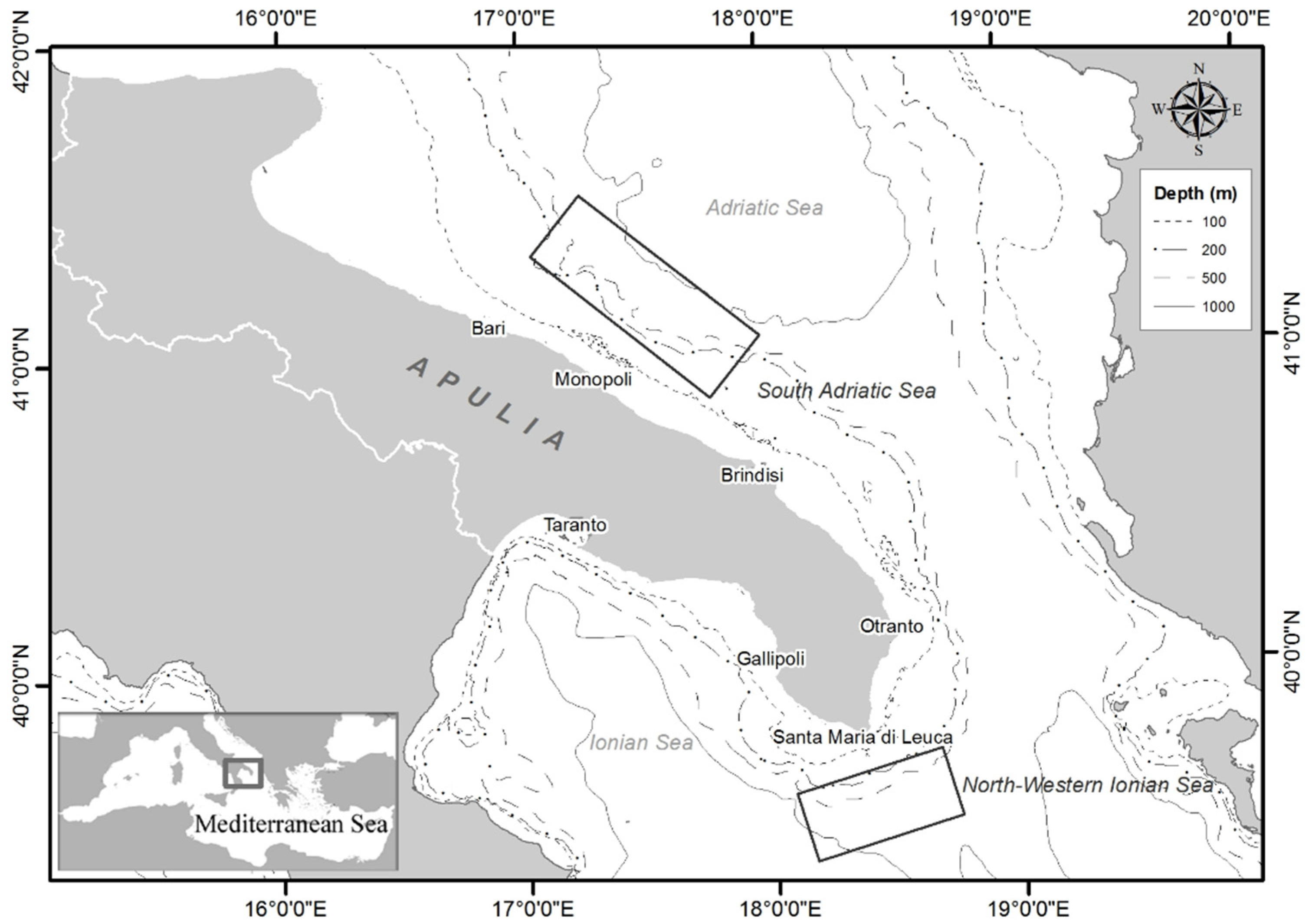
2.1. Area of Study
2.2. Survey Methodology
2.3. Data Analysis
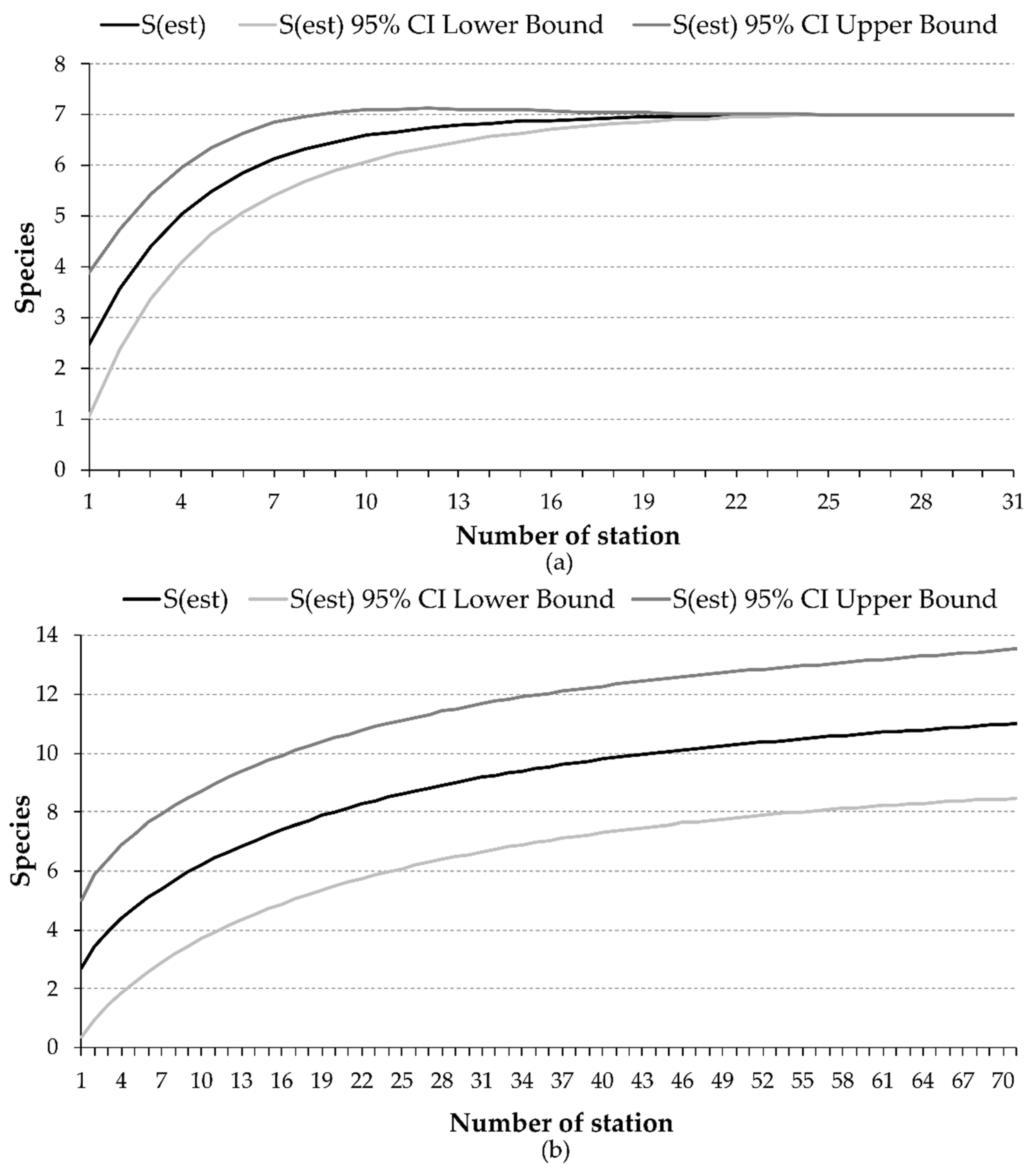
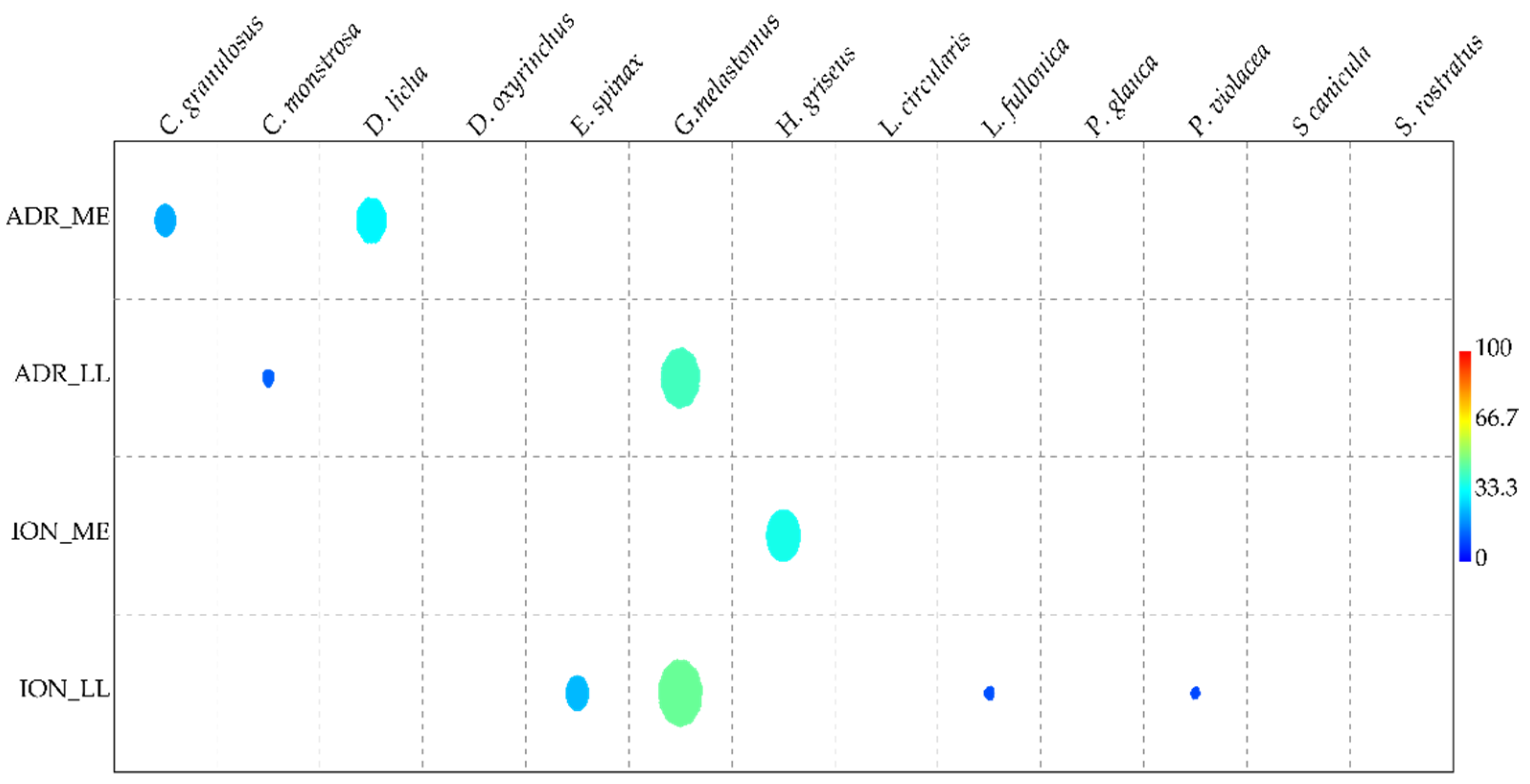
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Serena, F.; Abella, A.J.; Bargnesi, F.; Barone, M.; Colloca, F.; Ferretti, F.; Fiorentino, F.; Jenrette, J.; Moro, S. Species diversity, taxonomy and distribution of chondrichthyes in the Mediterranean and Black Sea. Eur. Zool. J. 2020, 87, 497–536. [Google Scholar] [CrossRef]
- Dulvy, N.K.; Simpfendorfer, C.A.; Davidson, L.N.K.; Fordham, S.V.; Bräutigam, A.; Sant, G.; Welch, D.J. Challenges and priorities in shark and ray conservation. Curr. Biol. 2017, 27, R565–R572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weigmann, S. Annotated checklist of the living sharks, batoids and chimaeras (Chondrichthyes) of the world, with a focus on biogeographical diversity. J. Fish Biol. 2016, 88, 837–1037. [Google Scholar] [CrossRef]
- Fields, A.; Feldheim, K.A.; Poulakis, G.R.; Chapman, D.D. Facultative parthenogenesis in a critically endangered wild vertebrate. Curr. Biol. 2015, 25, R439–R447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferretti, F.; Myers, R.A.; Serena, F.; Lotze, H.K. Loss of large predatory sharks from the Mediterranean Sea. Conserv. Biol. 2008, 22, 952–964. [Google Scholar] [CrossRef] [PubMed]
- Stevens, J.D.; Bonfil, R.; Dulvy, N.K.; Walker, P.A. The effects of fishing on sharks, rays, and chimaeras (chondrichthyans), and the implications for marine ecosystems. ICES J. Mar. Sci. 2000, 57, 476–494. [Google Scholar] [CrossRef]
- Cortes, E. Standardized diet compositions and trophic levels of sharks. ICES J. Mar. Sci. 1999, 56, 707–717. [Google Scholar] [CrossRef]
- Heupel, M.R.; Knip, D.M.; Simpfendorfer, C.A.; Dulvy, N.K. Sizing up the ecological role of sharks as predators. Mar. Ecol. Prog. Ser. 2014, 495, 291–298. [Google Scholar] [CrossRef] [Green Version]
- Rigby, C.; Simpfendorfer, C.A. Patterns in life history traits of deep-water chondrichthyans. Deep Sea Res. Part II Top. Stud. Oceanogr. 2015, 115, 30–40. [Google Scholar] [CrossRef]
- Peristeraki, P.; Tserpes, G.; Kavadas, S.; Kallianiotis, A.; Stergiou, K.I. The Effect of bottom trawl fishery on biomass variations of demersal chondrichthyes in the eastern Mediterranean. Fish. Res. 2020, 221, 105367. [Google Scholar] [CrossRef]
- Follesa, M.C.; Marongiu, M.F.; Zupa, W.; Bellodi, A.; Cau, A.; Cannas, R.; Colloca, F.; Djurovic, M.; Isajlovic, I.; Jadaud, A.; et al. Spatial variability of Chondrichthyes in the northern Mediterranean. Sci. Mar. 2019, 83S1, 81–100. [Google Scholar] [CrossRef] [Green Version]
- Devine, J.A.; Baker, K.D.; Haedrich, R.L. Deep-sea fishes qualify as endangered. Nature 2006, 439, 29. [Google Scholar] [CrossRef] [PubMed]
- Massutí, E.; Moranta, J. Demersal assemblages and depth distribution of elasmobranchs from the continental shelf and slope off the Balearic Islands (Western Mediterranean). ICES J. Mar. Sci. 2003, 60, 753–766. [Google Scholar] [CrossRef] [Green Version]
- Sion, L.; Bozzano, A.; D’Onghia, G.; Capezzuto, F.; Panza, M. Chondrichthyes species in deep waters of the Mediterranean Sea. Sci. Mar. 2004, 68, 153–162. [Google Scholar] [CrossRef] [Green Version]
- Ragonese, S.; Vitale, S.; Dimech, M.; Mazzola, S. Abundances of demersal sharks and chimaera from 1994-2009 scientific surveys in the Central Mediterranean Sea. PLoS ONE 2013, 8, e74865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marongiu, M.F.; Porcu, C.; Bellodi, A.; Cannas, R.; Cau, A.; Cuccu, D.; Mulas, A.; Follesa, M.C. Temporal dynamics of demersal chondrichthyan species in the Central Western Mediterranean Sea: The case study in Sardinia Island. Fish. Res. 2017, 193, 81–94. [Google Scholar] [CrossRef]
- Pikitch, E.K.; Chapman, D.D.; Babcock, E.A.; Shivji, M.S. Habitat use and demographic population structure of elasmobranchs at a Caribbean atoll (Glover’s Reef, Belize). Mar. Ecol. Prog. Ser. 2005, 302, 187–197. [Google Scholar] [CrossRef]
- Worm, B.; Davis, B.; Kettemer, L.; Ward-Paige, C.A.; Chapman, D.; Heithaus, M.R.; Kessel, S.T.; Gruber, S.H. global catches, exploitation rates, and rebuilding options for sharks. Mar. Policy 2013, 40, 194–204. [Google Scholar] [CrossRef]
- Simpfendorfer, C.A.; Hueter, R.E.; Bergman, U.; Connett, S.M.H. Results of a fishery-independent survey for pelagic sharks in the Western North Atlantic, 1977–1994. Fish. Res. 2002, 55, 175–192. [Google Scholar] [CrossRef]
- Baum, J.K.; Blanchard, W. Inferring Shark Population Trends from generalized linear mixed models of pelagic longline catch and effort data. Fish. Res. 2010, 102, 229–239. [Google Scholar] [CrossRef]
- Santana-Garcon, J.; Braccini, M.; Langlois, T.J.; Newman, S.J.; McAuley, R.B.; Harvey, E.S. Calibration of pelagic stereo-bruvs and scientific longline surveys for sampling sharks. Methods Ecol. Evol. 2014, 5, 824–833. [Google Scholar] [CrossRef] [Green Version]
- Andrews, K.S.; Williams, G.D.; Farrer, D.; Tolimieri, N.; Harvey, C.J.; Bargmann, G.; Levin, P.S. Diel Activity patterns of sixgill sharks, Hexanchus griseus: The Ups and Downs of an Apex Predator. Anim. Behav. 2009, 78, 525–536. [Google Scholar] [CrossRef]
- Rodríguez-Cabello, C.; Sánchez, F. Catch and post-release mortalities of deep-water sharks caught by bottom longlines in the Cantabrian Sea (NE Atlantic). J. Sea Res. 2017, 130, 248–255. [Google Scholar] [CrossRef]
- Brooks, E.J.; Mandelman, J.W.; Sloman, K.A.; Liss, S.; Danylchuk, A.J.; Cooke, S.J.; Skomal, G.B.; Philipp, D.P.; Sims, D.W.; Suski, C.D. The Physiological response of the caribbean reef shark (Carcharhinus perezi) to longline capture. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2012, 162, 94–100. [Google Scholar] [CrossRef]
- Skomal, G.B. Evaluating the physiological and physical consequences of capture on post-release survivorship in large pelagic fishes. Fish. Manag. Ecol. 2007, 14, 81–89. [Google Scholar] [CrossRef]
- Malcolm, H.A.; Gladstone, W.; Lindfield, S.; Wraith, J.; Lynch, T.P. Spatial and temporal variation in reef fish assemblages of marine parks in New South Wales, Australia—baited video observations. Mar. Ecol. Prog. Ser. 2007, 350, 277–290. [Google Scholar] [CrossRef]
- Brooks, E.J.; Sloman, K.A.; Sims, D.W.; Danylchuk, A.J. Validating the use of baited remote underwater video surveys for assessing the diversity, distribution and abundance of sharks in the Bahamas. Endanger. Species Res. 2011, 13, 231–243. [Google Scholar] [CrossRef]
- White, J.; Simpfendorfer, C.A.; Tobin, A.J.; Heupel, M.R. Application of baited remote underwater video surveys to quantify spatial distribution of elasmobranchs at an ecosystem scale. J. Exp. Mar. Biol. Ecol. 2013, 448, 281–288. [Google Scholar] [CrossRef]
- McLean, D.L. Comparison of baited longlines and baited underwater cameras for assessing the composition of continental slope deepwater fish assemblages off Southeast Australia. Deep Sea Res. Part I Oceanogr. Res. Pap. 2015, 98, 10–20. [Google Scholar] [CrossRef]
- Devine, B.M.; Wheeland, L.J.; Fisher, J.A.D. First Estimates of Greenland shark (Somniosus microcephalus) local abundances in arctic waters. Sci. Rep. 2018, 8, 974. [Google Scholar] [CrossRef] [Green Version]
- Pimentel, C.R.; Andrades, R.; Ferreira, C.E.L.; Gadig, O.B.F.; Harvey, E.S.; Joyeux, J.; Giarrizzo, T. BRUVS Reveal locally extinct shark and the way for shark monitoring in brazilian oceanic islands. J. Fish Biol. 2020, 96, 539–542. [Google Scholar] [CrossRef]
- Osgood, G.J.; McCord, M.E.; Baum, J.K. Using Baited Remote Underwater Videos (BRUVs) to characterize chondrichthyan communities in a global biodiversity hotspot. PLoS ONE 2019, 14, e0225859. [Google Scholar] [CrossRef] [PubMed]
- Söffker, M.; Sloman, K.A.; Hall-Spencer, J.M. In situ observations of fish associated with coral reefs off Ireland. Deep Sea Res. Part I Oceanogr. Res. Pap. 2011, 58, 818–825. [Google Scholar] [CrossRef] [Green Version]
- D’Onghia, G.; Capezzuto, F.; Carluccio, A.; Carlucci, R.; Giove, A.; Mastrototaro, F.; Panza, M.; Sion, L.; Tursi, A.; Maiorano, P. Exploring composition and behaviour of fish fauna by in situ observations in the Bari Canyon (Southern Adriatic Sea, Central Mediterranean). Mar. Ecol. 2015, 36, 541–556. [Google Scholar] [CrossRef]
- D’Onghia, G.; Capezzuto, F.; Cardone, F.; Carlucci, R.; Carluccio, A.; Chimienti, G.; Corriero, G.; Longo, C.; Maiorano, P.; Mastrototaro, F.; et al. Macro- and megafauna recorded in the submarine Bari Canyon (Southern Adriatic, Mediterranean Sea) using different tools. Mediterr. Mar. Sci. 2015, 16, 180. [Google Scholar] [CrossRef]
- Capezzuto, F.; Maiorano, P.; Panza, M.; Indennidate, A.; Sion, L.; D’Onghia, G. Occurrence and behaviour of Paromola cuvieri (Crustacea, Decapoda) in the Santa Maria Di Leuca cold-water coral community (Mediterranean Sea). Deep Sea Res. Part I Oceanogr. Res. Pap. 2012, 59, 1–7. [Google Scholar] [CrossRef]
- Yeh, J.; Drazen, J. Baited-Camera observations of deep-sea megafaunal scavenger ecology on the California slope. Mar. Ecol. Prog. Ser. 2011, 424, 145–156. [Google Scholar] [CrossRef] [Green Version]
- Harvey, E.S.; Newman, S.J.; McLean, D.L.; Cappo, M.; Meeuwig, J.J.; Skepper, C.L. Comparison of the relative efficiencies of stereo-BRUVs and traps for sampling tropical continental shelf demersal fishes. Fish. Res. 2012, 125–126, 108–120. [Google Scholar] [CrossRef]
- Capezzuto, F.; Calculli, C.; Carlucci, R.; Carluccio, A.; Maiorano, P.; Pollice, A.; Sion, L.; Tursi, A.; D’Onghia, G. Revealing the Coral habitat effect on benthopelagic fauna diversity in the Santa Maria Di Leuca Cold-water coral province using different devices and bayesian hierarchical modelling. Aquat. Conserv. Mar. Freshw. Ecosyst. 2019, 29, 1608–1622. [Google Scholar] [CrossRef]
- Ayma, A.; Aguzzi, J.; Canals, M.; Lastras, G.; Bahamon, N.; Mecho, A.; Company, J.B. Comparison between ROV video and Agassiz trawl methods for sampling deep water fauna of submarine canyons in the Northwestern Mediterranean Sea with observations on behavioural reactions of target species. Deep Sea Res. Part I Oceanogr. Res. Pap. 2016, 114, 149–159. [Google Scholar] [CrossRef] [Green Version]
- Cappo, M.; Speare, P.; De’ath, G. Comparison of baited remote underwater video stations (BRUVS) and prawn (shrimp) trawls for assessments of fish biodiversity in inter-reefal areas of the Great Barrier Reef Marine Park. J. Exp. Mar. Biol. Ecol. 2004, 302, 123–152. [Google Scholar] [CrossRef]
- Dulvy, N.K.; Allen, D.; Ralph, G.; Walls, R. The Conservation Status of Sharks, Rays and Chimaeras in the Mediterranean Sea; IUCN: Malaga, Spain, 2016; p. 14. [Google Scholar]
- Dulvy, N.K.; Fowler, S.L.; Musick, J.A.; Cavanagh, R.D.; Kyne, M.; Harrison, L.R.; Carlson, J.K.; Davidson, L.N.; Francis, M.P.; Pollock, C.M.; et al. Extinction risk and conservation of the world’s sharks and rays. eLife 2014, 3, e00590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colloca, F.; Enea, M.; Ragonese, S.; Di Loreno, M. A century fishery data documenting the collapse of smooth-hounds (Mustelus spp.) in Mediterranean Sea. Aquat. Conserv. 2017, 27, 1145–1155. [Google Scholar] [CrossRef]
- Pacoureau, N.; Rigby, C.L.; Kyne, P.M.; Sherley, R.B.; Winker, H.; Carlson, J.K.; Fordham, S.V.; Barreto, R.; Fernando, D.; Francis, M.P.; et al. Half a century of global decline in oceanic sharks and rays. Nature 2021, 589, 567–571. [Google Scholar] [CrossRef] [PubMed]
- Barausse, A.; Correale, V.; Curkovic, A.; Finotto, L.; Riginella, E.; Visentin, E.; Mazzoldi, C. The role of fisheries and the environment in driving the decline of elasmobranchs in the Northern Adriatic Sea. ICES J. Mar. Sci. 2014, 71, 1593–1603. [Google Scholar] [CrossRef] [Green Version]
- D’Onghia, G.; Sion, L.; Capezzuto, F. Cold-water coral habitats benefit adjacent fisheries along the Apulian margin (Central Mediterranean). Fish. Res. 2019, 213, 172–179. [Google Scholar] [CrossRef]
- Trincardi, F.; Foglini, F.; Verdicchio, G.; Asioli, A.; Correggiari, A.; Minisini, D.; Piva, A.; Remia, A.; Ridente, D.; Taviani, M. The Impact of cascading currents on the Bari Canyon system, SW-Adriatic Margin (Central Mediterranean). Mar. Geol. 2007, 246, 208–230. [Google Scholar] [CrossRef]
- Ridente, D.; Foglini, F.; Minisini, D.; Trincardi, F.; Verdicchio, G. Shelf-edge erosion, sediment failure and inception of Bari Canyon on the Southwestern Adriatic Margin (Central Mediterranean). Mar. Geol. 2007, 246, 193–207. [Google Scholar] [CrossRef]
- Chimienti, G.; Bo, M.; Taviani, M.; Mastrototaro, F. Occurrence and biogeography of Mediterranean cold-water corals. In Mediterranean Cold-Water Corals: Past, Present and Future; Orejas, C., Jiménez, C., Eds.; Springer International Publishing AG: Berlin/ Heidelberg, Germany, 2019; Volume 9, pp. 295–333. [Google Scholar]
- D’Onghia, G.; Calculli, C.; Capezzuto, F.; Carlucci, R.; Carluccio, A.; Maiorano, P.; Pollice, A.; Ricci, P.; Sion, L.; Tursi, A. New records of cold-water coral sites and fish fauna characterization of a potential network existing in the Mediterranean Sea. Mar. Ecol. 2016, 37, 1398–1422. [Google Scholar] [CrossRef]
- Taviani, M.; Angeletti, L.; Canese, S.; Cannas, R.; Cardone, F.; Cau, A.; Cau, A.B.; Follesa, M.C.; Marchese, F.; Montagna, P.; et al. The “Sardinian cold-water coral province” in the context of the Mediterranean coral ecosystems. Deep Sea Res. Part II Top. Stud. Oceanogr. 2017, 145, 61–78. [Google Scholar] [CrossRef]
- Freiwald, A.; Beuck, L.; Ruggerberg, A.; Taviani, M.; Hebblen, D. The white coral community in the Central Mediterranean Sea revealed by ROV surveys. Oceanography 2009, 22, 36–52. [Google Scholar] [CrossRef] [Green Version]
- Turchetto, M.; Boldrin, A.; Langone, L.; Miserocchi, S.; Tesi, T.; Foglini, F. Particle transport in the Bari Canyon (Southern Adriatic Sea). Mar. Geol. 2007, 246, 231–247. [Google Scholar] [CrossRef]
- Angeletti, L.; Taviani, M.; Canese, S.; Foglini, F.; Mastrototaro, F.; Argnani, A.; Trincardi, F.; Bakran-Petricioli, T.; Ceregato, A.; Chimienti, G.; et al. New Deep-Water cnidarian sites in the Southern Adriatic Sea. Mediterr. Mar. Sci. 2014, 15, 263. [Google Scholar] [CrossRef] [Green Version]
- Sion, L.; Calculli, C.; Capezzuto, F.; Carlucci, R.; Carluccio, A.; Cornacchia, L.; Maiorano, P.; Pollice, A.; Ricci, P.; Tursi, A.; et al. Does the Bari Canyon (Central Mediterranean) influence the fish distribution and abundance? Prog. Oceanogr. 2019, 170, 81–92. [Google Scholar] [CrossRef]
- Mastrototaro, F.; D’Onghia, G.; Corriero, G.; Matarrese, A.; Maiorano, P.; Panetta, P.; Gherardi, M.; Longo, C.; Rosso, A.; Sciuto, F.; et al. Biodiversity of the white coral bank off Cape Santa Maria Di Leuca (Mediterranean Sea): An update. Deep Sea Res. Part II Top. Stud. Oceanogr. 2010, 57, 412–430. [Google Scholar] [CrossRef]
- Savini, A.; Vertino, A.; Marchese, F.; Beuck, L.; Freiwald, A. Mapping cold-water coral habitats at different scales within the Northern Ionian Sea (Central Mediterranean): An assessment of coral coverage and associated vulnerability. PLoS ONE 2014, 9, e87108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tursi, A.; Mastrototaro, F.; Matarrese, A.; Maiorano, P.; D’Onghia, G. Biodiversity of the white coral reefs in the Ionian Sea (Central Mediterranean). Chem. Ecol. 2004, 20, 107–116. [Google Scholar] [CrossRef]
- D’Onghia, G.; Maiorano, P.; Sion, L.; Giove, A.; Capezzuto, F.; Carlucci, R.; Tursi, A. Effects of deep-water coral banks on the abundance and size structure of the megafauna in the Mediterranean Sea. Deep Sea Res. Part II Top. Stud. Oceanogr. 2010, 57, 397–411. [Google Scholar] [CrossRef]
- D’Onghia, G.; Maiorano, P.; Carlucci, R.; Capezzuto, F.; Carluccio, A.; Tursi, A.; Sion, L. Comparing Deep-sea fish fauna between coral and non-coral “megahabitats” in the Santa Maria Di Leuca cold-water coral province (Mediterranean Sea). PLoS ONE 2012, 7, e44509. [Google Scholar] [CrossRef]
- D’Onghia, G.; Capezzuto, F.; Carlucci, R.; Carluccio, A.; Maiorano, P.; Panza, M.; Ricci, P.; Sion, L.; Tursi, A. Using a benthic lander to explore and monitor vulnerable ecosystems in the Mediterranean Sea. ACTA IMEKO 2018, 7, 45. [Google Scholar] [CrossRef]
- Magurran, A. Ecological Diversity and Its Measurement. Princeton University Press: Princeton, NJ, USA, 1991; 178p. [Google Scholar]
- Colwell, R.K.; Chao, A.; Gotelli, N.J.; Lin, S.-Y.; Mao, C.X.; Chazdon, R.L.; Longino, J.T. Models and Estimators Linking Individual-Based and Sample-Based Rarefaction, Extrapolation and Comparison of Assemblages. J. Plant Ecol. 2012, 5, 3–21. [Google Scholar] [CrossRef] [Green Version]
- Brown, S.C.; Bizzarro, J.J.; Cailliet, G.M.; Ebert, D.A. Breaking with Tradition: Redefining Measures for Diet Description with a Case Study of the Aleutian Skate Bathyraja aleutica (Gilbert 1896). Environ. Biol. Fishes 2012, 95, 3–20. [Google Scholar] [CrossRef]
- Clarke, K.R. Non-parametric multivariate analyses of changes in community structure. Austral Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- Anderson, M.J. A New method for non-parametric multivariate analysis of variance: Non-parametric manova for ecology. Austral Ecol. 2001, 26, 32–46. [Google Scholar] [CrossRef]
- Dufrêne, M.; Legendre, P. Species assemblages and indicator species: The need for a flexible asymmetrical approach. Ecol. Monogr. 1997, 67, 345–366. [Google Scholar] [CrossRef]
- Hammer, O.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Clarke, K.; Gorley, R.N. Primer v6: User Manual/Tutorial; PRIMER-E: Plymouth, UK, 2006. [Google Scholar]
- Anderson, M.J.; Gorley, R.N.; Clarke, K.R. PERMANOVA+ for PRIMER: Guide to Software and Statistical Methods; PRIMER-E: Plymouth, UK, 2008. [Google Scholar]
- Spedicato, M.T.; Massutí, E.; Merigot, B.; Tserpes, G.; Jadaud, A.; Relini, G. The MEDITS trawl survey specifications in an ecosystem approach to fishery management. Sci. Mar. 2019, 83, 9–20. [Google Scholar] [CrossRef]
- Bozzano, A.; Murgia, R.; Vallerga, S.; Hirano, J.; Archer, S. The Photoreceptor System in the Retinae of Two Dogfishes, Scyliorhinus canicula and Galeus melastomus: Possible Relationship with Depth Distribution and Predatory Lifestyle. J. Fish Biol. 2001, 59, 1258–1278. [Google Scholar] [CrossRef]
- Navarro, J.; López, L.; Coll, M.; Barría, C.; Saez-Liante, R. Short- and Long- term Importance of Small Sharks in the Diet of the Rare Deep Sea Shark Dalatia licha. Mar. Biol. 2014, 161, 1697–1707. [Google Scholar] [CrossRef]
- Tursi, A.; D’Onghia, G.; Matarrese, A.; Piscitelli, G. Observations on population biology of Galeus melastomus Rafinesque, 1810 (CHONDRICHTHYES, SCYLIORHINIDAE) in the Ionian Sea. Cybium 1993, 17, 187–196. [Google Scholar]
- Ricci, P.; Sion, L.; Capezzuto, F.; Cipriano, G.; D’Onghia, G.; Libralato, S.; Maiorano, P.; Tursi, A.; Carlucci, R. Modelling the trophic roles of the demersal Chondrichthyes in the Northern Ionian Sea (Central Mediterranean Sea). Ecol. Model. 2021, 444, 109468. [Google Scholar] [CrossRef]
- Megalofonou, P.; Chatzispyrou, A. Sexual Maturity and Feeding of the Gulper Shark, Centrophorus granulosus, from the Eastern Mediterranean Sea. Cybium 2006, 30, 67–74. [Google Scholar]
- Guallart, J.; Vicent, J.J. Changes in Composition During Embryo Development of the Gulper Shark, Centrophorus granulosus (Elasmobranchii, Centrophoridae): An Assessment of Maternal-Embryonic Nutritional Relationships. Environ. Biol. Fishes 2001, 61, 135–150. [Google Scholar] [CrossRef]
- Metochis, C.P.; Carmona-Antoñanzas, G.; Kousteni, V.; Damalas, D.; Megalofonou, P. Population Structure and Aspects of the Reproductive Biology of the Blackmouth Catshark, Galeus Melastomus Rafinesque, 1810 (Chondrichthyes: Scyliorhinidae) Caught Accidentally off the Greek Coasts. J. Mar. Biol. Assoc. U. K. 2018, 98, 909–925. [Google Scholar] [CrossRef]
- D’Iglio, C.; Savoca, S.; Rinelli, P.; Spanò, N.; Capillo, G. Diet of the Deep-Sea Shark Galeus Melastomus Rafinesque, 1810, in the Mediterranean Sea: What We Know and What We Should Know. Sustainability 2021, 13, 3962. [Google Scholar] [CrossRef]
- Dulvy, N.K.; Reynolds, J.D. Predicting extinction vulnerability in skates. Conserv. Biol. 2002, 16, 440–450. [Google Scholar] [CrossRef] [Green Version]
- Flowers, K.I.; Ajemian, M.J.; Bassos-Hull, K.; Feldheim, K.A.; Hueter, R.E.; Papastamatiou, Y.P.; Chapman, D.D. A review of batoid philopatry, with implications for future research and population management. Mar. Ecol. Prog. Ser. 2016, 562, 251–261. [Google Scholar] [CrossRef] [Green Version]
- Capezzuto, F.; Ancona, F.; Carlucci, R.; Carluccio, A.; Cornacchia, L.; Maiorano, P.; Ricci, P.; Sion, L.; Tursi, A.; D’Onghia, G. Cold-Water Coral Communities in the Central Mediterranean: Aspects on Megafauna Diversity, Fishery Resources and Conservation Perspectives. Rend. Lincei Sci. Fis. Nat. 2018, 29, 589–597. [Google Scholar] [CrossRef]
- GFCM-RAC/SPA. In Report of the Transversal Workshop on Marine Protected Areas (MPAs); FAO: Salammbo, Tunisia, 2007.
- Angeletti, L.; D’Onghia, G.; Otero, M.d.M.; Settanni, A.; Spedicato, M.T.; Taviani, M. A Perspective for Best Governance of the Bari Canyon Deep-Sea Ecosystems. Water 2021, 13, 1646. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Longline | MEMO Lander | |||
|---|---|---|---|---|
| Southern Adriatic Sea | North-Western Ionian Sea | Southern Adriatic Sea | North-Western Ionian Sea | |
| Number of stations | 32 | 39 | 19 | 12 |
| Sampling hours (h) | 134 | 152.7 | 120 | 103 |
| Depth range (m) | 314–612 | 418–635 | 427–788 | 547–792 |
| Longline | MEMO Lander | |||
|---|---|---|---|---|
| Species | Southern Adriatic Sea | North-Western Ionian Sea | Southern Adriatic Sea | North-Western Ionian Sea |
| Centrophorus granulosus (Bloch & Schneider, 1801) | 0.01 | 0.04 | 0.08 | 0.01 |
| Chimaera monstrosa Linnaeus, 1758 | 0.05 | |||
| Dalatias licha (Bonnaterre, 1788) | 0.01 | 0.11 | 0.01 | |
| Dipturus oxyrinchus (Linnaeus, 1758) | 0.02 | 0.01 | 0.03 | |
| Etmopterus spinax (Linnaeus, 1758) | 0.14 | 0.32 | 0.06 | 0.02 |
| Galeus melastomus Rafinesque, 1810 | 3.78 | 4.03 | 0.10 | |
| Hexanchus griseus (Bonnaterre, 1788) | 0.03 | 0.09 | ||
| Leucoraja circularis (Couch, 1838) | 0.02 | |||
| Leucoraja fullonica (Linnaeus, 1758) | 0.04 | |||
| Prionace glauca (Linnaeus, 1758) | 0.01 | |||
| Pteroplatytrygon violacea (Bonaparte, 1832) | 0.03 | 0.07 | ||
| Scyliorhinus canicula (Linnaeus, 1758) | 0.02 | |||
| Somniosus rostratus (Risso, 1827) | 0.01 | |||
| Longline | MEMO Lander | |||
|---|---|---|---|---|
| Southern Adriatic Sea | North-Western Ionian Sea | Southern Adriatic Sea | North-Western Ionian Sea | |
| Shannon–Wiener | 0.30 | 0.48 | 1.72 | 1.33 |
| Margalef | 0.96 | 1.08 | 1.61 | 1.52 |
| Pielou | 0.16 | 0.23 | 0.89 | 0.83 |
| df | S | MS | Pseudo-F | p-Value | Perms | ||
|---|---|---|---|---|---|---|---|
| Tools | Tools | 1 | 54721 | 54721 | 37475 | 0.001 | 997 |
| Res | 83 | 1.212 e+5 | 1460.2 | ||||
| Total | 84 | 1.7592 e+5 | |||||
| Area | Area | 1 | 2334.2 | 2334.2 | 11.161 | 0.328 | 998 |
| Res | 83 | 1.7358 e+5 | 2091.4 | ||||
| Total | 84 | 1.7592 e+5 |
| Species | Av. dissim | Contrib. % | Cumulative % |
|---|---|---|---|
| Galeus melastomus | 60.14 | 74.65 | 74.65 |
| Etmopterus spinax | 6.67 | 8.28 | 82.93 |
| Hexanchus griseus | 3.23 | 4.01 | 86.94 |
| Centrophorus granulosus | 3.18 | 3.94 | 90.88 |
| Dalatias licha | 2.89 | 3.59 | 94.47 |
| Pteroplatytrygon violacea | 1.30 | 1.61 | 96.08 |
| Dipturus oxyrinchus | 1.05 | 1.31 | 97.38 |
| Scyliorhinus canicula | 0.89 | 1.10 | 98.49 |
| Leucoraja fullonica | 0.43 | 0.53 | 99.02 |
| Chimaera monstrosa | 0.38 | 0.47 | 99.49 |
| Somniosus rostratus | 0.18 | 0.22 | 99.71 |
| Leucoraja circularis | 0.12 | 0.15 | 99.86 |
| Prionace glauca | 0.11 | 0.14 | 100.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carluccio, A.; Capezzuto, F.; Maiorano, P.; Sion, L.; D’Onghia, G. Deep-Water Cartilaginous Fishes in the Central Mediterranean Sea: Comparison between Geographic Areas with Two Low Impact Tools for Sampling. J. Mar. Sci. Eng. 2021, 9, 686. https://doi.org/10.3390/jmse9070686
Carluccio A, Capezzuto F, Maiorano P, Sion L, D’Onghia G. Deep-Water Cartilaginous Fishes in the Central Mediterranean Sea: Comparison between Geographic Areas with Two Low Impact Tools for Sampling. Journal of Marine Science and Engineering. 2021; 9(7):686. https://doi.org/10.3390/jmse9070686
Chicago/Turabian StyleCarluccio, Angela, Francesca Capezzuto, Porzia Maiorano, Letizia Sion, and Gianfranco D’Onghia. 2021. "Deep-Water Cartilaginous Fishes in the Central Mediterranean Sea: Comparison between Geographic Areas with Two Low Impact Tools for Sampling" Journal of Marine Science and Engineering 9, no. 7: 686. https://doi.org/10.3390/jmse9070686