
Resistance to Three Distinct Begomovirus Species in the Agronomical Superior Tropical Pumpkin Line AVPU1426 Developed at the World Vegetable Center

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Screening of Populations by Whitefly Inoculations
2.2. Screening for Reaction to SLCPV-TW Inoculation in Taiwan
2.3. Yield Trial, Fruit Traits Evaluation
2.4. Carotenoids Estimation
3. Results
3.1. Segregation of SLCCNV and ToLCNDV Resistances in Crosses Waltham Butternut × AVPU1426
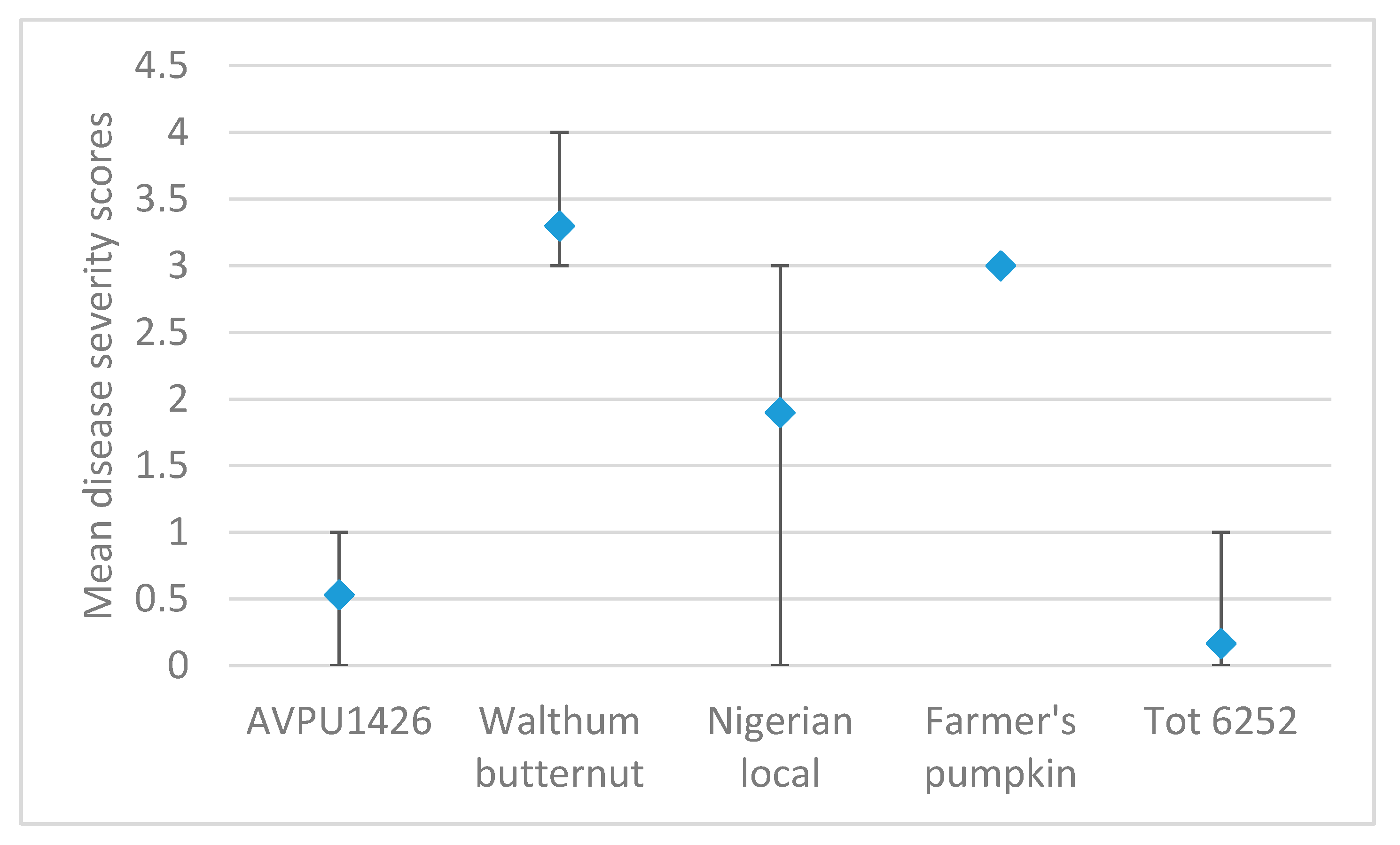
3.2. Screening for Reaction to SLCPV-TW Inoculation in Taiwan
3.3. Horticultural, Nutritional, and Yield Potential of AVPU1426
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- US Department of Agriculture, A.R.S. Nutrient Data Laboratory. USDA National Nutrient Database for Standard Reference, Release 28 (Slightly Revised). Version Current: May 2016. Available online: http://www.ars.usda.gov/nea/bhnrc/mafcl (accessed on 1 March 2020).
- FAOSTAT. Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 4 May 2021).
- Nagendran, K.; Mohankumar, S.; Aravintharaj, R.; Balaji, C.G.; Manoranjitham, S.K.; Singh, A.K.; Rai, A.B.; Singh, B.; Karthikeyan, G. The occurrence and distribution of major viruses infecting cucurbits in Tamil Nadu state, India. Crop Prot. 2017, 99, 10–16. [Google Scholar] [CrossRef]
- Chan, Y.L.; Lee, L.M.; Shih, S.L.; Kuo, F.H.; Kenyon, L. Survey of virus diseases affecting squash (Cucurbita moschata) in Taiwan. Acta. Hortic. 2019, 1257, 23–28. [Google Scholar] [CrossRef]
- Rosen, R.; Kanakala, S.; Kliot, A.; Cathrin, P.B.; Farich, B.A.; Santana-Magal, N.; Elimelech, M.; Kontsedalov, S.; Lebedev, G.; Cilia, M.; et al. Persistent, circulative transmission of begomoviruses by whitefly vectors. Curr. Opin. Virol. 2015, 15, 1–8. [Google Scholar] [CrossRef]
- European and Mediterranean Plant Protection Organization. EPPO Alert List—Tomato Leaf Curl New Delhi Virus. 2015. Available online: https://www.eppo.int/QUARANTINE/Alert_List/viruses/ToLCNDV.htm (accessed on 18 November 2020).
- Rodriguez, E.; Tellez, M.M.; Janssen, D. Whitefly control strategies against tomato leaf curl New Delhi virus in greenhouse zucchini. Int. J. Environ. Res. Public Health 2019, 16, 2673. [Google Scholar] [CrossRef] [PubMed]
- Lapidot, M.; Friedmann, M.; Pilowsky, M.; Ben-Joseph, R.; Cohen, S. Effect of host plant resistance to Tomato yellow leaf curl virus (TYLCV) on virus acquisition and transmission by its whitefly vector. Phytopathology 2001, 91, 1209–1213. [Google Scholar] [CrossRef] [PubMed]
- Lopez, C.; Ferriol, M.; Pico, M.B. Mechanical transmission of tomato leaf curl New Delhi virus to cucurbit germplasm: Selection of tolerance sources in Cucumis melo. Euphytica 2015, 204, 679–691. [Google Scholar] [CrossRef]
- Sáez, C.; Martínez, C.; Ferriol, M.; Manzano, S.; Velasco, L.; Jamilena, M.; López, C.; Picó, B. Resistance to Tomato leaf curl New Delhi virus in Cucurbita spp. Ann. Appl. Biol. 2016, 169, 91–105. [Google Scholar] [CrossRef]
- Muhire, B.M.; Varsani, A.; Martin, D.P. SDT: A virus classification tool based on pairwise sequence alignment and identity calculation. PLoS ONE 2014, 9, e108277. [Google Scholar] [CrossRef] [PubMed]
- Zhong, L.; Gustavsson, K.E.; Oredsson, S.; Glab, B.; Yilmaz, J.L.; Olsson, M.E. Determination of free and esterified carotenoid composition in rose hip fruit by HPLC-DAD-APCI+-MS. Food Chem. 2016, 210, 541–550. [Google Scholar] [CrossRef]
- Verlaan, M.G.; Hutton, S.F.; Ibrahe, R.M.; Kormelink, R.; Visser, R.G.F.; Scott, J.W.; Edwards, J.D.; Bai, Y. The tomato yellow leaf curl virus resistance genes Ty-1 and Ty-3 are allelic and code for DFDGD-Class RNA Dependent RNA polymerases. PLoS Genet. 2013, 9, e1003399. [Google Scholar] [CrossRef]
- Brown, J.; Zerbini, F.M.; Navas-Castillo, J.; Moriones, E.; Ramos-Sobrinho, R.; Silva, J.F.; Fiallo-Olivé, E.; Briddon, R.; Hernández-Zepeda, C.; Idris, A.; et al. Revision of Begomovirus taxonomy based on pairwise sequence comparisons. Arch. Virol. 2015, 160, 1593–1619. [Google Scholar] [CrossRef]
- Juárez, M.; Tovar, R.; Fiallo-Olivé, E.; Aranda, M.A.; Gosálvez, B.; Castillo, P.; Moriones, E.; Navas-Castillo, J. First detection of tomato leaf curl New Delhi virus infecting zucchini in Spain. Plant Dis. 2014, 98, 857–858. [Google Scholar] [CrossRef]
- Sífres, A.; Sáez, C.; Ferriol, M.; Selmani, E.A.; Riado, J.; Picó, B.; López, C. First report of tomato leaf curl New Delhi virus infecting zucchini in Morocco. Plant Dis. 2018, 102, 1045. [Google Scholar] [CrossRef]
- Kheireddine, A.; Sifres, A.; Sáez, C.; Picó, B.; López, C. First report of tomato leaf curl New Delhi virus infecting cucurbit plants in Algeria. Plant Dis. 2019, 103, 3291. [Google Scholar] [CrossRef]
- Martín-Hernández, A.M.; Picó, B. Natural resistances to viruses in cucurbits. Agronomy 2021, 11, 23. [Google Scholar] [CrossRef]
- Paris, H.S.; Cohen, S.; Burger, Y.; Yoseph, R. Single-gene resistance to Zucchini yellow mosaic virus in Cucurbita moschata. Euphytica 1988, 37, 27–29. [Google Scholar] [CrossRef]
- Brown, R.N.; Bolanos-Herrera, A.; Myers, J.R.; Miller, J. Inheritance of resistance to four cucurbit viruses in Cucurbita moschata. Euphytica 2003, 129, 253–258. [Google Scholar] [CrossRef]
- Paris, H.S.; Cohen, S. Oligogenic inheritance for resistance to Zucchini yellow mosaic virus in Cucurbita pepo. Ann. Appl. Biol. 2000, 136, 209–214. [Google Scholar] [CrossRef]
- Formisano, G.; Paris, H.S.; Frusciante, L.; Ercolano, M.R. Commercial Cucurbita pepo squash hybrids carrying disease resistance introgressed from C. moschata have high genetic similarity. Plant Genet. Resour. 2010, 8, 198–203. [Google Scholar] [CrossRef]
- McCreight, J.D.; Kishaba, A.N. Reaction of cucurbit species to squash leaf curl virus and sweetpotato whitefly. J. Am. Soc. Hortic. Sci. 1991, 116, 137–141. [Google Scholar] [CrossRef]
- Kesumawati, E.; Okabe, S.; Khalil, M.; Alfan, G.; Bahagia, P.; Pohan, N.; Zakaria, S.; Koeda, S. Molecular Characterization of Begomoviruses Associated with Yellow Leaf Curl Disease in Solanaceae and Cucurbitaceae Crops from Northern Sumatra, Indonesia. Hortic. J. 2020, 89, 410–416. [Google Scholar] [CrossRef]
- Yadav, K.K.; Hegde, G.; Agarwal, P.; Chawda, V.; Kenyon, L.; Dhillon, N.P.S. Multi-location preliminary field screening of World Vegetable Center bitter gourd breeding lines for reaction to Tomato leaf curl new Delhi virus in selected hotspots in India. Acta Hortic. 2019, 1257, 9–14. [Google Scholar] [CrossRef]
- Dhillon, N.P.S.; Masud, M.A.T.; Pruangwitayakun, S.; Natheung, M.; Lertlam, S.; Jarret, R.L. Evaluation of loofah lines for resistance to tomato leaf curl New Delhi virus and downy mildew, as well as key horticultural traits. Agriculture 2020, 10, 298. [Google Scholar] [CrossRef]
- Islam, S.; Munshi, A.D.; Mandal, B.; Kumar, R.; Behera, T.K. Genetics of resistance in Luffa cylindrica Roem against Tomato leaf curl New Delhi virus. Euphytica 2010, 174, 83–89. [Google Scholar] [CrossRef]
- Saez, C.; Esteras, C.; Martinez, C.; Ferriol, M.; Dhillon, N.P.S.; Lopez, C.; Pico, B. Resistance to tomato leaf curl New Delhi virus in melon is controlled by a major QTL located on chromosome 11. Plant Cell Rep. 2017, 36, 1571–1584. [Google Scholar] [CrossRef]
- Sáez, C.; Ambrosio, L.G.M.; Miguel, S.M.; Valcárcel, J.V.; Díez, M.J.; Picó, B.; López, C. Resistant sources and genetic control of resistance to ToLCNDV in cucumber. Microorganisms 2021, 9, 913. [Google Scholar] [CrossRef]
- Saez, C.; Martinez, C.; Montero-Pau, J.; Esteras, C.; Sifres, A.; Blanca, J.; Ferriol, M.; Lopez, C.; Pico, B. A major QTL located in chromosome 8 of Cucurbita moschata is responsible for resistance to tomato leaf curl New Delhi virus. Front. Plant Sci. 2020, 11, 207. [Google Scholar] [CrossRef] [PubMed]
- Romero-Masegosa, J.; Martinex, C.; Aguado, E.; Garcia, A.; Cebrian, G.; Iglesias-Moya, J.; Paris, H.S.; Jamilena, M. Response of Cucurbita spp. To tomato leaf curl New Delhi virus inoculation and identification of a dominant source of resistance in Cucurbita moschata. Plant Pathol. 2020, 70, 206. [Google Scholar] [CrossRef]
- Mundt, C.C. Pyramiding for resistance durability: Theory and practice. Phytopathology 2018, 108, 79–802. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Parent or Generation | Total No. of Plants | Resistant Plants a | Susceptible Plants a | Expected Segregation for Single Recessive Resistance Gene (R:S) | Х2 | p-Value |
|---|---|---|---|---|---|---|
| Waltham Butternut | 10 | 0 | 10 | 0:1 | − | − |
| AVPU1426 | 10 | 10 | 0 | 1:0 | − | − |
| F1 | 10 | 0 | 10 | 0:1 | − | − |
| F2 | 100 | 24 | 76 | 1:3 | 0.053 | 0.817 |
| BCAVPU1426 | 48 | 18 | 30 | 1:1 | 2.96 | 0.083 |
| Parent or Cross | No. of Plants | Resistant Plants a | Susceptible Plants a | Expected Segregation for Single Recessive Resistance Gene (R:S) | Х2 | p-Value |
|---|---|---|---|---|---|---|
| Waltham Butternut | 10 | 0 | 10 | 0:1 | − | − |
| AVPU1426 | 10 | 10 | 0 | 1:0 | − | − |
| F1 | 10 | 0 | 10 | 0:1 | − | − |
| F2 | 100 | 27 | 73 | 1:3 | 0.213 | 0.644 |
| BCAVPU1426 | 50 | 23 | 27 | 1:1 | 0.32 | 0.571 |
| Entry | Fruit Weight (kg/Fruit−1) | Fruit Yield (t/ha−1), | TSS (°B) | Fruit Skin Color | Fruit Flesh Color b | Fruit Shape | β-Carotene a (mg/100 g) | Lutein a (µg/100 g) | Zeaxanthin a (µg/100 g) |
|---|---|---|---|---|---|---|---|---|---|
| AVPU1426 | 3.19 | 20.74 | 6.69 | Mottled green | Yellow-orange (YO16A) | Flat round | 1.57 ± 0.26 | 407.25 ± 36.38 | 429.18 ± 15.94 |
| Rajah | 3.21 | 18.61 | 8.29 | Mottled green | Yellow-orange (YO17A) | Flat round | |||
| LSD (0.05) | 0.2 | 3.86 | 1.71 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dhillon, N.P.S.; Srimat, S.; Laenoi, S.; Bhunchoth, A.; Phuangrat, B.; Warin, N.; Deeto, R.; Chatchawankanphanich, O.; Jom, K.N.; Sae-tan, S.; et al. Resistance to Three Distinct Begomovirus Species in the Agronomical Superior Tropical Pumpkin Line AVPU1426 Developed at the World Vegetable Center. Agronomy 2021, 11, 1256. https://doi.org/10.3390/agronomy11061256
Dhillon NPS, Srimat S, Laenoi S, Bhunchoth A, Phuangrat B, Warin N, Deeto R, Chatchawankanphanich O, Jom KN, Sae-tan S, et al. Resistance to Three Distinct Begomovirus Species in the Agronomical Superior Tropical Pumpkin Line AVPU1426 Developed at the World Vegetable Center. Agronomy. 2021; 11(6):1256. https://doi.org/10.3390/agronomy11061256
Chicago/Turabian StyleDhillon, Narinder P. S., Supornpun Srimat, Suwannee Laenoi, Anjana Bhunchoth, Bencharong Phuangrat, Nuchnard Warin, Rungnapa Deeto, Orawan Chatchawankanphanich, Kriskamol Na Jom, Sudathip Sae-tan, and et al. 2021. "Resistance to Three Distinct Begomovirus Species in the Agronomical Superior Tropical Pumpkin Line AVPU1426 Developed at the World Vegetable Center" Agronomy 11, no. 6: 1256. https://doi.org/10.3390/agronomy11061256