
Structure, Diversity, and Environmental Determinants of High-Latitude Threatened Conifer Forests


 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
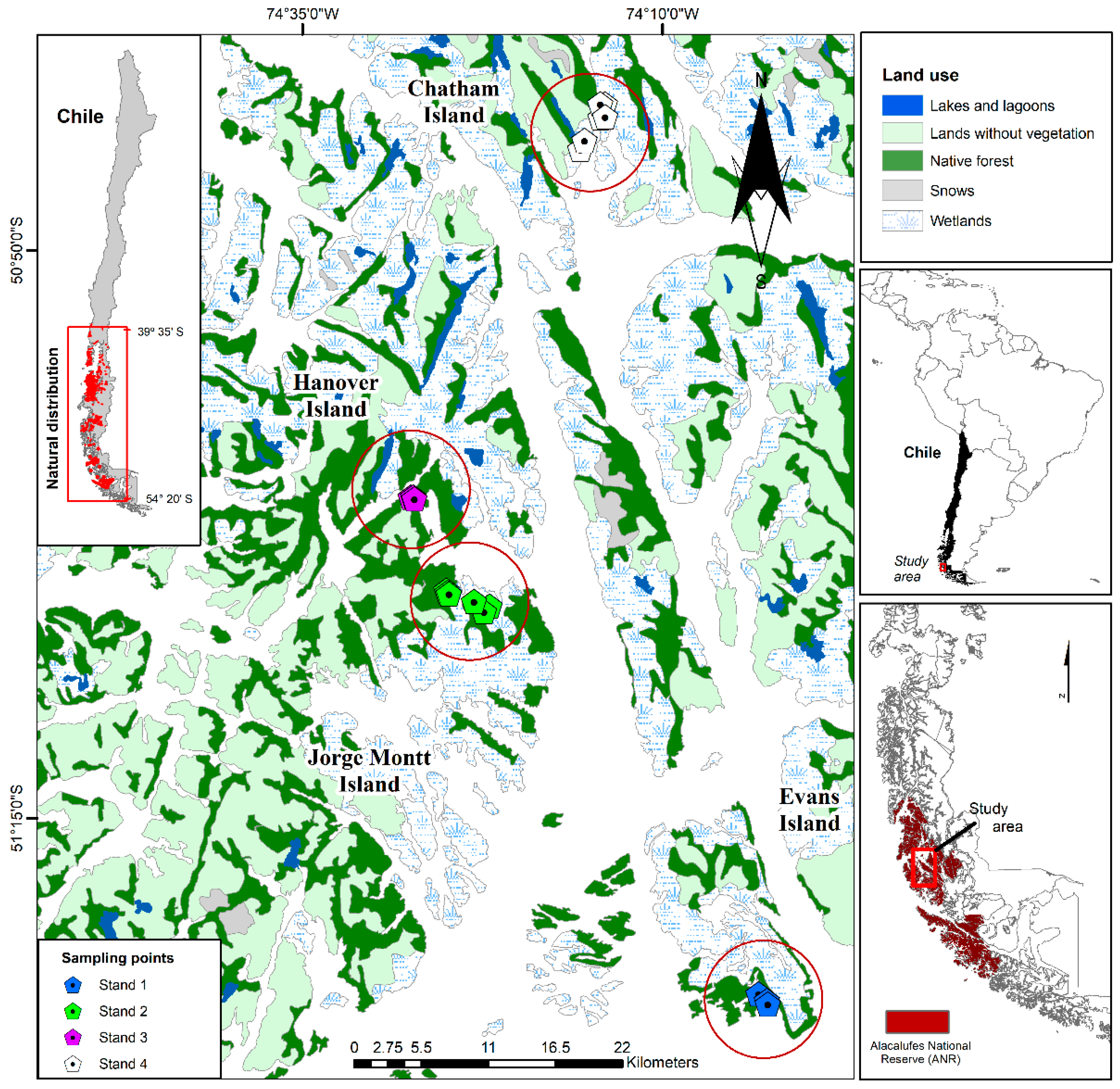
2.1. Study Area
2.2. Sampling and Data Collection
2.3. Diversity
2.4. Environmental Matrix Based on Biophysical Variables
2.5. Statistical Analyses
3. Results
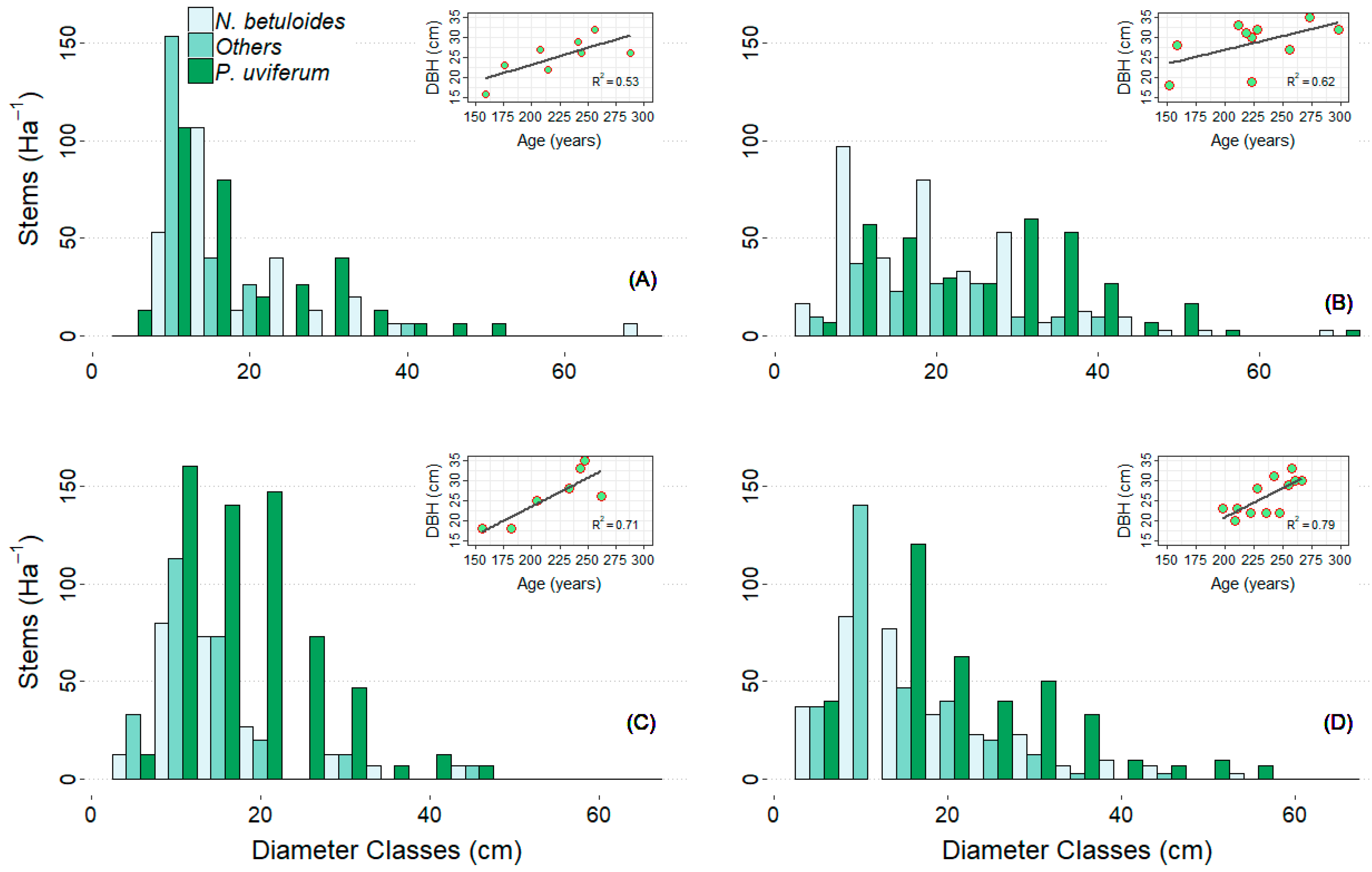
3.1. Quantitative Structure of the Stands
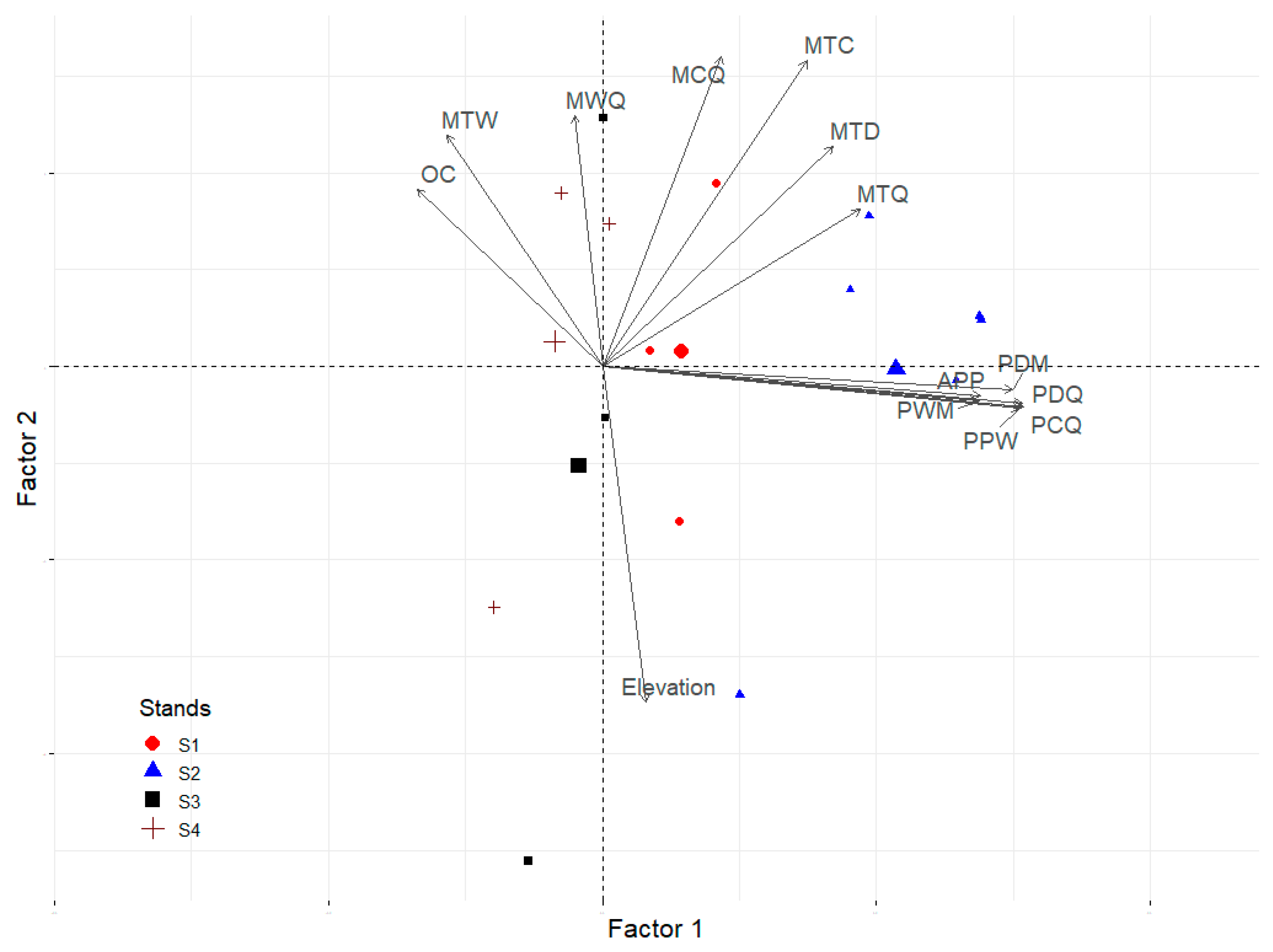
3.2. Analysis of the Environmental Matrix Based on Biophysical Variables
3.3. Composition, Diversity, and Community Structure
4. Discussion
Implication for the Conservation of High-Latitude Conifer Forests
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Myers, N.; Mittermeler, R.A.; Mittermeler, C.G.; da Fonseca, G.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef]
- Armesto, J.; Rozzi, R.; Smith-Ramirez, C.; Arroyo, M.T. Conservation targets in South American temperate forests. Science 1998, 282, 1271–1272. [Google Scholar] [CrossRef] [Green Version]
- Smith-Ramírez, C. The Chilean coastal range: A vanishing center of biodiversity and endemism in South American temperate rainforests. Biodivers. Conserv. 2004, 13, 373–393. [Google Scholar] [CrossRef]
- Veblen, T.T.; Delmastro, R.J.; Schlatter, J.E. The conservation of Fitzroya cupressoides and its environment in southern Chile. Environ. Conserv. 1976, 3, 291–301. [Google Scholar] [CrossRef]
- Walter, K.S.; Gillett, H.J. (Eds.) 1997 IUCN Red List of Threatened Plants; IUCN—The World Conservation Union: Gland, Switzerland, 1998. [Google Scholar]
- Lara, A.; Donoso, C.; Escobar, B.; Rovere, A.; Premoli, A.; Soto, D.; Banniester, J. Pilgerodendron uviferum (D.Don) florin. In Las Especies Arbóreas de los Bosques Templados de Chile y Argentina: Autoecología, 1st ed.; Donoso, C., Ed.; Marisa Cuneo Ediciones: Valdivia, Chile, 2006. [Google Scholar]
- Pisano, E. Fitogeografía de Fuego-Patagonia chilena I.- comunidades vegetales entre latitudes 51 y 56° Sur: Punta Arenas, Chile. An. Inst. Patagon. 1977, 8, 121–250. [Google Scholar]
- Cruz, G.; Lara, A. Tipificación, Cambio de Estructura y Normas de Manejo para Ciprés de las Guaytecas (Pilgerodendron uviferum (D.Don) Florin) en la Isla Grande de Chiloé. Tesis de Pregrado, Universidad de Chile, Santiago, Chile, 1981. [Google Scholar]
- Moore, D. Flora of Tierra del Fuego; Anthony Nelson: Shrewsbury, UK, 1983. [Google Scholar]
- Allnutt, T.; Newton, A.C.; Premoli, A.; Lara, A. Genetic variation in the threatened South American conifer Pilgerodendron uviferum (Cupressaceae) detected using RAPD markers. Biol. Conserv. 2003, 10, 9–18. [Google Scholar] [CrossRef]
- Soto, D.P.; Figueroa, H. Efectos de las alteraciones antrópicas sobre la estructura y composición de rodales de Pilgerodendron uviferum en la Cordillera de la Costa de Chile. Ecol. Aust. 2008, 18, 13–25. [Google Scholar]
- Bannister, J.R.; Donoso, P.; Bauhus, J. Persistence of the slow growing conifer Pilgerodendron uviferum in old-growth and fire-disturbed southern bog forests. Ecosystems 2012, 15, 1158–1172. [Google Scholar] [CrossRef]
- Martínez, O.; Muñoz, A.M. Aspectos conservativos de las coníferas chilenas. Bosque 1988, 9, 77–82. [Google Scholar] [CrossRef]
- Díaz-Vaz, J.E. Anatomía de Maderas; Marisa Cúneo Ediciones: Valdivia, Chile, 2003. [Google Scholar]
- Bannister, J.R.; Wagner, S.; Donoso, P.J.; Bauhus, J. The importance of seed trees in the dioecious conifer Pilgerodendron uviferum for passive restoration of fire disturbed southern bog forests. Austral Ecol. 2014, 39, 204–213. [Google Scholar] [CrossRef]
- Farjon, A.; Page, C. (Eds.) Coníferas. Estudio de estado y plan de acción de conservación; Grupo de Especialistas en Coníferas de la CSE/UICN; UICN: Gland, Switerland; Cambridge, UK, 1999. [Google Scholar]
- Hilton-Taylor, C. (Ed.) 2000 IUCN Red List of Threatened Species; IUCN: Gland, Switzerland; Cambridge, UK, 2000. [Google Scholar]
- Holdgate, M.W. Vegetation and soils in the south Chilean Islands. Ecology 1961, 49, 559–580. [Google Scholar] [CrossRef]
- Holz, A.; Veblen, T.T. Pilgerodendron uviferum: The southernmost tree-ring fire recorder species. Écoscience 2009, 16, 322–329. [Google Scholar] [CrossRef]
- Strasburger, E. Tratado de Botánica, 3rd ed.; Ediciones OMEGA: Barcelona, Spain, 2004. [Google Scholar]
- Luebert, F.; Pliscoff, P. Sinopsis Bioclimática y Vegetacional de Chile, 1st ed.; Editorial Universitaria: Santiago, Chile, 2015. [Google Scholar]
- Donoso, C. Bosques Templados de Chile y Argentina: Variación, Estructura y Dinámica, 4th ed.; Editorial Universitaria: Santiago, Chile, 1998. [Google Scholar]
- Sousa, W. The rol of disturbance in natural communities. Annu. Rev. Ecol. Syst. 1984, 15, 353–391. [Google Scholar] [CrossRef]
- Rovere, A.; Premoli, A.; Newton, A.C. Estado de conservación del Ciprés de las Guaitecas (Pilgerodendron uviferum (D.Don) Florin) en la Argentina. Bosque 2002, 23, 11–19. [Google Scholar] [CrossRef]
- Premoli, A.C.; Souto, C.P.; Allnutt, T.R.; Newton, A.C. Effects of population disjunction on isozyme variation in the widespread Pilgerodendron uviferum. Heredity 2001, 87, 337–343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vera, M.; Iglesias, V.; Whitlock, C. Late and postglacial vegetation and fire history from Cordón Serrucho Norte, northern Patagonia. Palaeog. Palaeoclim. Palaeoecol. 2013, 371, 109–118. [Google Scholar]
- Di Castri, F.; Hajek, E. Bioclimatología de Chile, 1st ed.; Editorial Universidad Católica: Santiago, Chile, 1976. [Google Scholar]
- Szeicz, J.; Lara, A.; Díaz, S.; Aravena, J.C. Dendrochronological Studies of Pilgerodendron uviferum in Southwestern South America. In Dendrocronología en América Latina; Roig, F., Ed.; Editorial Universidad del Cuyo: Mendoza, Argentina, 2000. [Google Scholar]
- Soto, D.P.; Le Quesne, C.; Lara, A.; Gardner, M.F. Precarious conservation status of Pilgerodendron uviferum forests in their northern distribution in the Chilean Coastal Range. Bosque 2007, 28, 263–270. [Google Scholar] [CrossRef]
- Soto, D.P.; Bannister, J.R.; Ríos, A.I.; Le Quesne, C. Nuevos registros de poblaciones amenazadas de Pilgerodendron uviferum ((D.Don) Florin) en su límite norte en la Cordillera de la Costa chilena. Gayana Bot. 2010, 67, 120–124. [Google Scholar] [CrossRef]
- Luzio, W.; Seguel, O.; Casanova, M. Suelos de la zona mediterránea húmeda. In Suelos de Chile; Luzio, W., Ed.; Universidad de Chile: Santiago, Chile, 2010; pp. 195–239. [Google Scholar]
- Zuloaga, F.O.; Morrone, O.; Belgrano, M. (Eds.) Monographs in Systematic Botany from the Missouri Botanical Garden 107; Missouri Botanical Garden: St. Louis, MO, USA, 2008. [Google Scholar]
- Marticorena, C.; Quezada, M. Catálogo de la flora vascular de Chile. Gayana Bot. 1985, 42, 1–155. [Google Scholar]
- Quinn, G.; Keough, M. Experimental Design and Data Analysis for Biologists; Cambridge University Press: Cambridge, UK, 2002. [Google Scholar]
- Stokes, M.A.; Smiley, T.L. An Introduction to Tree-Ring Dating; University of Arizona Press: Tucson, AZ, USA, 1996. [Google Scholar]
- Whittaker, R.H. Vegetation of the Siskiyou mountains, Oregon and California. Ecol. Monogr. 1960, 30, 279–338. [Google Scholar] [CrossRef]
- Shannon, C.E. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef] [Green Version]
- Hammer, Ø.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2013, 4, 9. [Google Scholar]
- Kindt, R.; Coe, R. Tree Diversity Analysis: A Manual and Software for Common Statistical Methods Forecological and Biodiversity Studies; World Agroforestry Centre (ICRAF): Nairobi, Kenya, 2005. [Google Scholar]
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high-resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Hengl, T.; Mendes de Jesus, J.; Heuvelink, G.B.M.; Ruiperez Gonzalez, M.; Kilibarda, M.; Blagotić, A.; Shangguan, W.; Wright, M.N.; Geng, X.; Bauer-Marschallinger, B.; et al. SoilGrids250m: Global gridded soil information based on machine learning. PLoS ONE 2017, 12, e0169748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- CONAF. Catastro de los Recursos Vegetacionales Nativos de Chile: Monitoreo de Cambios y Actualizaciones Período 1997–2011; CONAF: Santiago, Chile, 2011. [Google Scholar]
- Holm, S. A Simple Sequentially Rejective Multiple Test Procedure. Scand. J. Stat. 1979, 6, 65–70. [Google Scholar]
- Shaffer, J.P. Modified Sequential Rejective Multiple Test Procedures. J. Am. Stat. Assoc. 1986, 81, 826–831. [Google Scholar] [CrossRef]
- de Mendiburu, F. Agricolae: Statistical Procedures for Agricultural Research. R Package Version 1.3-3. 2020. Available online: https://CRAN.R-project.org/package=agricolae (accessed on 18 January 2021).
- Kassambara, A.; Mundt, F. Extract and Visualize the Results of Multivariate Data Analyses. R Package Version 2017, 1, 337–354. [Google Scholar]
- Lê, S.; Josse, J.; Husson, F. FactoMineR: An R Package for Multivariate Analysis. J. Stat. Soft. 2008, 25, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Clarke, K.R.; Warwick, R.M. Change in Marine Communities: An Approach to Statistical Analysis and Interpretation, 2nd ed.; Primer-E: Plymouth, MA, USA, 2001. [Google Scholar]
- Anderson, M.J. Permutational Multivariate Analysis of Variance (PERMANOVA); Wiley StatsRef: Statistics Reference Online; Wiley: Hoboken, NJ, USA, 2017; pp. 1–15. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. 2017. Available online: https://CRAN.R-project.org/package=vegan (accessed on 7 June 2020).
- Al-Kandari, N.; Jolliffe, I. Variable selection and interpretation of covariance principal components. Commun. Stat. Simul. Comput. 2001, 30, 339–354. [Google Scholar] [CrossRef]
- Hechenleitner, P.; Gardner, M.; Thomas, P.; Echeverría, C.; Escobar, B.; Brownless, P.; Martínez, C. Plantas Amenazadas del Centro-sur de Chile: Distribución, Conservación y Propagación, 1st ed.; Universidad Austral de Chile y Real Jardín Botánico de Edimburgo: Valdivia, Chile, 2005. [Google Scholar]
- Bannister, J.R.; Lara, A.; Le Quesne, C. Estructura y dinámica de bosques de Pilgerodendron uviferum afectados por incendios en la Cordillera de la Costa de la Isla Grande de Chiloé. Bosque 2008, 29, 33–43. [Google Scholar] [CrossRef]
- Soto, D.P.; Jacobs, D.F.; Salas, C.; Donoso, P.J.; Fuentes, C.; Puettmann, K.J. Light and nitrogen interact to influence regeneration in old-growth Nothofagus-dominated forests in southcentral Chile. For. Ecol. Manag. 2017, 384, 303–313. [Google Scholar] [CrossRef]
- Liang, J.; Crowther, T.W.; Picard, N.; Wiser, S.; Zhou, M.; Alberti, G.; Schulze, E.D.; McGuire, A.D.; Bozzato, F.; Pretzsch, H.; et al. Positive biodiversity-productivity relationship predominant in global forests. Science 2016, 354, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szeicz, J.; Haberle, S.; Bennett, K. Dynamics of North Patagonian Rainforests from Fine-Resolution Pollen, Charcoal and Tree-Ring Analysis, Chonos Archipelago, Southern Chile. Austral Ecol. 2003, 28, 413–422. [Google Scholar] [CrossRef]
- Eitzinger, J.; Orlandini, S.; Stefanski, R.; Naylor, R.E.L. Climate change and agriculture: Introductory editorial. J. Agric. Sci. Camb. 2010, 148, 499–500. [Google Scholar] [CrossRef] [Green Version]
- Bagaram, M.; Giuliarelli, D.; Chirici, G.; Giannetti, F.; Barbati, A. UAV Remote Sensing for Biodiversity Monitoring: Are Forest Canopy Gaps Good Covariates? Remote Sens. 2018, 10, 1397. [Google Scholar]
- Souza, C.M.; Siqueira, J.V.; Sales, M.H.; Fonseca, A.V.; Ribeiro, J.G.; Numata, I.; Cochrane, M.A.; Barber, C.P.; Roberts, D.A.; Barlow, J. Ten-year landsat classification of deforestation and forest degradation in the Brazilian amazon. Remote Sens. 2013, 5, 5493–5513. [Google Scholar] [CrossRef] [Green Version]
- Bullock, E.; Woodcock, C.E.; Olofsson, P. Monitoring tropical forest degradation using spectral unmixing and Landsat time series analysis. Remote Sens. Environ. 2018, 238, 110968. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factor | Variable | Unit | Description | Reference |
|---|---|---|---|---|
| 1. Climate | AMT | °C | Annual mean temperature | Hijmans et al. [40] |
| MDR | °C | Mean diurnal range (mean of monthly (max temp−min temp)) | ||
| ISO | Index | Isothermality (BIO2/BIO7) (×100) | ||
| TSS | Index | Temperature seasonality (standard deviation × 100) | ||
| MTW | °C | Max temperature of warmest month | ||
| MTC | °C | Min temperature of coldest month | ||
| TAR | °C | Temperature annual range (BIO5–BIO6) | ||
| MTQ | °C | Mean temperature of wettest quarter | ||
| MTD | °C | Mean temperature of driest quarter | ||
| MWQ | °C | Mean temperature of warmest quarter | ||
| MCQ | °C | Mean temperature of coldest quarter | ||
| APP | mm | Annual precipitation | ||
| PWM | mm | Precipitation of wettest month | ||
| PDM | mm | Precipitation of driest month | ||
| PPS | % | Precipitation seasonality (coefficient of variation) | ||
| PWQ | mm | Precipitation of wettest quarter | ||
| PDQ | mm | Precipitation of driest quarter | ||
| PPW | mm | Precipitation of warmest quarter | ||
| PCQ | mm | Precipitation of coldest quarter | ||
| 2. Edaphic | PWP | % | Available soil water capacity (volumetric fraction) until wilting point | SoilGrids Hengl et al. [41] |
| Silt | % | Silt content (2–50 micrometer) mass fraction | ||
| Sand | % | Sand content (50–2000 micrometer) mass fraction | ||
| OC | kg m−3 | Soil organic carbon density | ||
| C | permille | Soil organic carbon content | ||
| pH | scale | Soil pH × 10 in H2O | ||
| CEC | Cmol kg−1 | Cation exchange capacity of soil | ||
| Clay | % | Clay content | ||
| BD | kg m−3 | Bulk density | ||
| SD | cm | Absolute depth to bedrock | ||
| 3. Topographic | Aspect | range | Aspect (rank) | CONAF et al. [42] |
| Slope | % | Slope (degree in %) | ||
| Elevation | m asl | m asl (meter above sea level) |
| Parameters | Stands | p-Value | ||||
|---|---|---|---|---|---|---|
| S1 | S2 | S3 | S4 | |||
| Descriptive | Aspect | SW | FLAT | SW | FLAT | - |
| Area (ha) | 41.83 | 175.71 | 22.31 | 112.23 | - | |
| Age (years) | 214 (1.0) c | 242 (0.5) b | 245 (1.0) a | 201 (1.0) d | 0.001 | |
| Dasometric | Havg (m) | 5.80 (0.2) d | 11.00 (1.1) a | 6.30 (0.2) c | 7.20 (0.2) b | 0.001 |
| MSD (cm) | 18.57 (4.3) ab | 25.13 (5.0) a | 17.40 (4.0) b | 18.20 (2.4) b | 0.044 | |
| D (trees ha−1) | 806 (208.2) | 856 (290.0) | 1087 (477.0) | 1187 (290.0) | 0.237 | |
| BA (m2 ha−1) | 21.27 (6.8) | 40.43 (14.7) | 23.57 (7.1) | 30.10 (5.3) | 0.081 | |
| Variable | PC1 | PC2 | PC3 |
|---|---|---|---|
| Aspect | −0.034 | 0.180 | 0.006 |
| Slope | −0.028 | 0.135 | 0.208 |
| Elevation | −0.002 | 0.348 | −0.117 |
| AMT | −0.023 | −0.257 | 0.296 |
| MDR | 0.205 | 0.169 | 0.199 |
| ISO | 0.160 | 0.120 | 0.013 |
| TSS | 0.201 | 0.132 | 0.234 |
| MTW | 0.141 | −0.053 | 0.354 |
| MTC | −0.152 | −0.288 | 0.103 |
| TAR | 0.208 | 0.157 | 0.193 |
| MTQ | −0.160 | −0.005 | 0.317 |
| MTD | −0.193 | −0.256 | 0.010 |
| MWQ | 0.062 | −0.181 | 0.356 |
| MCQ | −0.118 | −0.297 | 0.160 |
| APP | −0.273 | 0.008 | −0.008 |
| PWM | −0.272 | −0.001 | −0.038 |
| PDM | −0.272 | −0.017 | −0.029 |
| PPS | −0.229 | 0.130 | 0.120 |
| PWQ | −0.273 | 0.003 | −0.014 |
| PDQ | −0.273 | 0.008 | −0.003 |
| PPW | −0.272 | 0.010 | 0.000 |
| PCQ | −0.273 | 0.004 | −0.010 |
| Tex | −0.100 | 0.216 | 0.114 |
| PWP | 0.150 | −0.229 | −0.207 |
| Silt | 0.185 | −0.141 | −0.130 |
| Sand | −0.116 | 0.204 | 0.168 |
| OC | 0.094 | −0.259 | −0.124 |
| C | 0.116 | −0.278 | −0.058 |
| pH | −0.199 | 0.141 | −0.043 |
| CEC | 0.057 | 0.033 | −0.296 |
| Clay | −0.013 | −0.196 | −0.125 |
| BD | −0.063 | 0.068 | −0.244 |
| SD | −0.036 | −0.138 | 0.182 |
| Abundance by Stand | ||||
|---|---|---|---|---|
| Species | Stand 1 | Stand 2 | Stand 3 | Stand 4 |
| P. uviferum | 16.33 (6.89) | 26.68 (10.74) | 35.70 (16.37) | 32.19 (7.33) |
| T. stipularis | 39.53 (22.54) | 1.17 (0.98) | 2.67 (2.19) | 17.41 (11.74) |
| D. winteri | 2.33 (1.45) | 4.17 (2.43) | 10.00 (4.93) | 9.83 (2.30) |
| L. ferruginea | - | - | - | 2.50 (1.23) |
| M. magellanica | 0.33 (0.33) | 1.33 (1.33) | - | 0.67 (0.33) |
| N. betuloides | 13.00 (3.51) | 25.38 (3.74) | 13.35 (1.86) | 17.02 (1.06) |
| P. laetevirens | 2.35 (1.46) | 4.70 (3.73) | 6.38 (5.88) | 3.18 (1.46) |
| N. antarctica | - | - | - | 2.00 (0.86) |
| P. nubigena | - | - | - | 2.33 (1.12) |
| Diversity Index by Stand | p-Values | ||||
|---|---|---|---|---|---|
| Index | Stand 1 | Stand 2 | Stand 3 | Stand 4 | |
| Richness (S′) | 4.67 (0.33) b | 3.83 (0.40) b | 4.33 (0.33) b | 6.33 (0.21) a | 0.005 |
| Abundance (N) | 74.00 (32.13) | 63.33 (10.81) | 68.00 (19.00) | 87.17 (17.47) | 0.870 |
| Shannon index (H′ loge) | 1.21 (0.09) ab | 0.89 (0.14) b | 1.08 (0.22) ab | 1.45 (0.04) a | 0.028 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Esse, C.; Correa-Araneda, F.; Acuña, C.; Santander-Massa, R.; De Los Ríos-Escalante, P.; Saavedra, P.; Jaque-Jaramillo, X.; Moreno, R.; García-Meneses, P.M.; Soto, D.P. Structure, Diversity, and Environmental Determinants of High-Latitude Threatened Conifer Forests. Forests 2021, 12, 775. https://doi.org/10.3390/f12060775
Esse C, Correa-Araneda F, Acuña C, Santander-Massa R, De Los Ríos-Escalante P, Saavedra P, Jaque-Jaramillo X, Moreno R, García-Meneses PM, Soto DP. Structure, Diversity, and Environmental Determinants of High-Latitude Threatened Conifer Forests. Forests. 2021; 12(6):775. https://doi.org/10.3390/f12060775
Chicago/Turabian StyleEsse, Carlos, Francisco Correa-Araneda, Cristian Acuña, Rodrigo Santander-Massa, Patricio De Los Ríos-Escalante, Pablo Saavedra, Ximena Jaque-Jaramillo, Roberto Moreno, Paola Massyel García-Meneses, and Daniel P. Soto. 2021. "Structure, Diversity, and Environmental Determinants of High-Latitude Threatened Conifer Forests" Forests 12, no. 6: 775. https://doi.org/10.3390/f12060775