
Abstract
In the debate on causal explanation in biology, in the past two decades largely influenced by the new mechanist (NM) approach, the concept of a pathway has recently reemerged as a promising research agenda (see, in particular, Ross Philosophy of Science, 85(4), 551-572, 2018; The British Journal for the Philosophy of Science, 72(1), 131-158, 2021). Ross’ account of biological explanation differentiates several autonomous types of causal structures that play explanatory and other roles across the life sciences. NM, however, prioritizes mechanisms as vehicles of biological explanations. According to this program, the causal architecture of biological pathways and other causal structures, such as cascades and processes, can be interpreted with at least one of the NM’s mechanism concepts. In other words, these alternative causal structures are not sufficiently distinctive to merit the explanatory autonomy with regard to the NM corresponding concepts. We examine the explanatory practice of molecular biology and concur with Ross that there are indeed distinct types of causal structures, not all falling under the concept of a mechanism. Nonetheless, we show that the concept of mechanism is referring to a privileged causal structure, at the center of explanatory efforts in molecular biology. Pathways and other causal concepts, while somewhat distinct from mechanisms themselves, are explanatorily relevant to the degree in which they exhibit mechanistic features, are parts of a mechanistic architecture, or may lead to a mechanistic arrangement. What emerges in that manner is a hierarchy of causal structures with mechanisms at the explanatory top, and lower levels differing in the degree in which they contribute to mechanistic arrangements.




Similar content being viewed by others
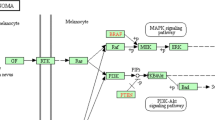


Notes
Differently than Ross (2021), which is specifically targeting the foundational and derivative issues concerned with the pathway concept, Ross (2018) examines the causal concept in question as related to the problem of causal selection. Despite that different focus, Ross’ earlier paper contains some important insights on the issue of features characterizing the causal structure of metabolic pathways.
Ross, at this and other similar points, insists on quite a strong analogy of the pathway causal structures of this kind and manufacturing contexts in ordinary life situations, such as the structure of an assembly line production. In our view, the characteristics and limits of this analogy require a separate analysis, which goes beyond the scope of the present paper.
The most frequently referred NM characterizations of the mechanism concept, at least, the seminal ones, are as follows: (1) “Mechanisms are entities and activities organized such that they are productive of regular changes from start or set-up to finish or termination conditions” (Machamer et al., 2000: 3); (2) “A mechanism for a phenomenon of behavior is: a complex system that produces that behavior by the interaction of a number of parts, where the interactions between parts can be characterized by direct, invariant, change-relating generalizations” (Glennan, 2002: S344); and (3) “A mechanism is a structure performing a function in virtue of its component parts, component operations, and their organization. The orchestrated functioning of the mechanism is responsible for one or more phenomena” (Bechtel & Abrahamsen, 2005: 423). We are here putting them on board in order to confront them to Ross’ characterizations of the main causal concepts in biology. Apart from that, we will be using (1)-(3) and some more recent versions both in moving objections to Ross’ account, and in arguing for our view of causal explanation in molecular biology.
Going up to higher levels, we arrive at the protein's causal contribution to more palpable higher endo-phenotypic and phenotypic effects.
In the next section, we see in more detail how both characteristics interact in providing causal explanations in molecular biology.
It is worth noting here that there is a previous list of features with regard to the causal structure of pathways, in particular in biochemistry, and referring causal concept. This Ross’ list puts forward the following features: (i) causal control, (ii) material continuity, and (iii) fixed order. Although the list only partially overlaps with the current one, both in number and respective characterization, we are not examining in this setting the details of their difference. The main reason for that is contextual; since the current list of features is closely related to the issue of causal explanation in biology, whereas the preceding list is targeting the causal selection problem (see Ross, 2018).
Special thanks to anonymous reviewers for pressing this point in our assessment of Ross’ account.
The abstraction feature of pathways plays an important role in Ross' assessment of why pathways are explanatory. Namely, she argues that there are some explananda for which the pathway information is explanatory, while the mechanistic one is not. According to Ross, if the pathway relations are fixed, lower-level mechanistic information can vary, i.e., lower-level causal information is not explanatory. Note that here we are not just concerned with the issue of abstraction/details that might describe a system at the same level, but lower levels are explicitly invoked. In other words, Ross puts forward an antireductionist argument invoking multiple realizability against the allegedly reductionist mechanist program. The NM program, however, is hardly reductionist in the standard meaning of the term. In her assessment of the overall NM commitments, Franklin-Hall (2016), for instance, identifies (1) somewhat reductive tendencies, i.e., insistence that phenomena are explained in lower-level terms, usually at one level below; and (2) non-reductive tendencies, i.e., resistance to the view that every phenomenon is ultimately explained at the physical level. The fact that mechanisms deployed by etiological explanations depict a sequence of causal steps occurring at the same level points to the conclusion that mechanistic explanatory strategies are not necessarily reductive. Thus, the explanatory non-reductiveness of pathways cannot serve as a ground for establishing a clear distinction between pathways and mechanisms.
One of the most frequent instantiations of the linear type of mechanism is protein synthesis. By that, we are here simply pointing out that there are other, non-linear types of a mechanism, such as the Krebs cycle in biochemistry.
Thagard (2003), for instance, argues that biochemical pathway is a kind of mechanism.
For a considerably long time, the duplication–divergence biological theory was the only available explanatory answer to the crucial ‘how-question’ of a “mystery of the orphans” (Dujon, 1996). Prior to the de novo explanatory model sketched above, researchers in this area took it to be highly unlikely that functional genes could emerge from random, noncoding genomic sequences (see Tautz et al., 2013). This assumption, however, has been overturned by evidence gathered in different taxa (see, e.g., Schmitz et al., 2018; Ruiz-Orera et al., 2018).
There are two leading explanatory models based on the biological theory in question, which respond differently to important difficulties of the above approach to the basic biological phenomenon of new gene production; namely, the “RNA first” and “ORF first” explanatory models (for more details, see McLysaght & Guerzoni, 2015). In our view, we may set aside in the present paper the specifics of this issue and its potential influence on the ordering of different causal structures within molecular explanations in the life sciences. We base our Fig. 3 in Section 4 on the model illustrated by Fig. 2.
It is important to emphasize that when describing different causal structures and assessing the explanatory power of their corresponding concepts throughout the paper we primarily have in mind individual organisms. Thus, causal arrows are representing events in which one activity directly causes the other, rather than different steps in evolutionary process as represented by Fig. 2 (for a comprehensive schematic representation see Fig. 3). The reference to the population level, however, is important because it provides information about robustness of the specific causal process in question. In other words, causal chains shielded by natural selection will more likely proceed in a regular manner.
Similarly, Bechtel and Richardson (2010: 35) talk about mechanisms as “discrete systems in nature”; Andersen (2014: 276) mechanisms as having “precise spatio-temporal boundaries”. Moreover, there is a noteworthy debate on the modularity of mechanisms, which relates to the above insulation characteristic. We are also sympathetic to the following understanding of that characteristic, according to which a mechanism is “a chain of linearly interacting entities that are somewhat independent of the chains that explain other phenomena in the system” (Franklin-Hall, 2008: 212; our emphasis).
This is not to say that the evolutionary causal processes are not explanatory or cannot be captured by the corresponding concepts of mechanism or pathway. Rather, we take it that such cases are complex and require separate analysis, which is beyond the scope of this paper. Here, we have tried to stay close to the type of examples already present in the debate on the explanatory importance of different causal concepts.
Thus, certain aspects or parts of the mechanism of protein synthesis occurring in individual organisms can be described as pathways. We take it that this does not clash with standard scientific use. For instance, in the literature, many references can be found for “the pathway of gene expression”, or for the “signaling pathways for gene expression”. Notice that Ross herself provides the example of gene expression pathways. On our account, they are, however, parts of the mechanistic structure of protein synthesis.
References
Akram, M. (2013). Mini-review on glycolysis and cancer. Journal of Cancer Education. https://doi.org/10.1007/s13187-013-0486-9.
Andersen, H. (2014). A field guide to mechanisms: Part I. Philosophy Compass, 9(4), 274–283.
Barros, D. B. (2008). Natural selection as a mechanism. Philosophy of Science, 75(3), 306–322.
Bechtel, W., & Abrahamsen, A. (2005). Explanation: A mechanist alternative. Studies in History and Philosophy of Biological and Biomedical Sciences, 36(2), 421–441. https://doi.org/10.1016/j.shpsc.2005.03.010
Bechtel, W., & Richardson, R. (2010). Discovering Complexity: Decomposition and Localization as Strategies in Scientific Research. MIT Press.
Carvunis, A.-R., Rolland, T., Wapinski, I., Calderwood, M. A., Yildirim, M. A., Simonis, N., Charloteaux, B., et al. (2012). Proto genes and de novo gene birth. Nature, 487(7407), 370–374. https://doi.org/10.1038/nature11184
Craver, C, & Tabery, J. (2019). Mechanisms in science. E. N. Zalta (Ed.), The Stanford Encyclopedia of Philosophy (Summer 2019 Edition). https://plato.stanford.edu/archives/sum2019/entries/science-mechanisms/. Accessed 8 Apr 2020.
Craver, F. C. (2001). Role functions, mechanisms, and hierarchy. Philosophy of Science, 68, 53–74.
Craver, F. C. (2007). Explaining the Brain: Mechanisms and the Mosaic Unity of Neuroscience. Clarendon Press.
Craver, F. C., & Darden, L. (2013). In Search of Mechanisms: Discoveries across the Life Sciences. The University of Chicago Press.
Dujon, B. (1996). The yeast genome project: What did we learn? Trends in Genetics: TIG, 12(7), 263–270.
Franklin-Hall, L. (2016). New mechanistic explanation and the need for explanatory constraints. In K. Aizawa & C. Gillett (Eds.), Scientific composition and metaphysical ground. New directions in the philosophy of science. Palgrave Macmillan. https://doi.org/10.1057/978-1-137-56216-6_2
Franklin-Hall, L. (2008). From a Microbiological Point of View. Columbia University.
Frietze, Seth, & Leatherman, Judith. (2014). Examining the process of de novo genebBirth: An Eeucational primer on ‘Integration of New Genes into Cellular Networks, and Their Structural Maturation.’ Genetics, 196(3), 593–99. https://doi.org/10.1534/genetics.113.160895
Garson, J. (2013). The functional sense of mechanism. Philosophy of Science, 80(3), 317–333.
Gill, K., et al. (2016). Glycolysis inhibition as a cancer treatment and its role in an anti-tumour immune response. Biochimica Et Biophysica Acta, 1866(2016), 87–105. https://doi.org/10.1016/j.bbcan.2016.06.005
Glennan, S. (2002). Rethinking mechanistic explanation. Philosophy of Science, 69(S3), S342–S353. https://doi.org/10.1086/341857
Illari, P., & Williamson, J. (2012). What is a mechanism? Thinking about mechanisms across the sciences. European Journal for Philosophy of Science, 2(1), 119–135.
Machamer, P., Darden, L., & Craver, C. F. (2000). Thinking about mechanisms. Philosophy of Science, 67(1), 1–25. https://doi.org/10.1086/392759
McLysaght, A., & Guerzoni, D. (2015). New genes from non-coding sequence: The role of de novo protein-coding genes in Eukaryotic Evolutionary Innovation. Philosophical Transactions of the Royal Society B, 370(1678), 20140332. https://doi.org/10.1098/rstb.2014.0332
Okasha, S. (2019). Philosophy of Biology. Oxford University Press.
Plissonneau, C., Benevenuto, J., Mohd-Assaad , N., Fouché, S., Hartmann, F. E., & Croll, D. (2017). Using Population and Comparative Genomics to Understand the Genetic Basis of Effector-Driven Fungal Pathogen Evolution. Frontiers in Plant Science, 8. https://doi.org/10.3389/fpls.2017.00119
Ross, L. N. (2018). Causal selection and the pathway concept. Philosophy of Science, 85(4), 551–572.
Ross, L. N. (2021). Causal concepts in biology: How pathways differ from mechanisms and why it matters. The British Journal for the Philosophy of Science, 72(1), 131–158. https://doi.org/10.1093/bjps/axy078
Ruiz-Orera, J., Verdaguer-Grau, P., Villanueva-Cañas, J. L., Messeguer, X., & Mar Albà, M. (2018). Translation of Neutrally Evolving Peptides Provides a Basis for De Novo Gene Evolution. Nature Ecology & Evolution, 2(5), 890–896. https://doi.org/10.1038/s41559-018-0506-6
Salmon, W. C. (1984). Scientific explanation and the causal structure of the world. Princeton University Press.
Schaffner, K. F. (2008). Etiological models in psychiatry. In K. S. Kendler, & J. Parnas (Eds.), Philosophical Issues in Psychiatry: Explanation, Phenomenology, and Nosology, (pp. 1–49). John Hopkins University.
Schmitz, J. F. & Bornberg-Bauer, E. (2017). Fact or fiction: Updates on how protein-coding genes might emerge de novo from previously non-coding DNA. F1000 Research, 6 (F1000 Faculty Rev), 57. https://doi.org/10.12688/f1000research.10079.1
Schmitz, J. F., Ullrich, K. K., & Bornberg-Bauer, E. (2018). Incipient de novo genes can evolve from frozen accidents that escaped rapid transcript turnover. Nature Ecology & Evolution, 2(10), 1626–1632. https://doi.org/10.1038/s41559-018-0639-7
Skipper, R., & Milstein, R. L. (2005). Thinking about evolutionary mechanisms: Natural selection. Studies in History and Philosophy of Science Part C Studies in History and Philosophy of Biological and Biomedical Sciences, 36(2), 327–347.
Tautz, D. (2014). The discovery of de novo gene evolution. Perspectives in Biology and Medicine, 57(1), 149–161. https://doi.org/10.1353/pbm.2014.0006
Tautz, D., & Domazet-Lošo, T. (2011). The Evolutionary origin of orphan genes. Nature Reviews. Genetics, 12(10), 692–702. https://doi.org/10.1038/nrg3053
Tautz, D., Neme, R., & Domazet-Lošo, T. (2013). Evolutionary Origin of Orphan Genes. Wiley. https://doi.org/10.1002/9780470015902.a0024601
Thagard, P. (2003). Pathways to biomedical discovery. Philosophy of Science, 70(2), 235–254.
Woodward, J. (2006). Sensitive and insensitive causation. The Philosophical Review, 115, 1–50.
Ylikosky, P., & Kuorikoski, J. (2010). Dissecting explanatory power. Philosophical Studies, 148(2), 201–219.
Acknowledgements
Parts of this paper were presented at the following events: 7th Biennial European Philosophy of Science Association Conference (EPSA 19), Geneva 2019 and the International Society for the History, Philosophy and Social Studies of Biology (ISHPSSB) biennial conference, Oslo 2019. We thank the audiences at these events for their helpful questions. Special thanks to two anonymous reviewers for their exceptionally insightful, precise and constructive comments. This paper is an output of the research project “Theoretical Underpinnings of Molecular Biology” (ThUMB), funded by the Croatian Science Foundation, project grant number: HRZZ-IP-2018-01-3378, and doctoral grant number: DOK-2018-09-7078. We would also like to acknowledge the support from the University of Rijeka (project KUBIM: uniri-human-18-265).
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
This article belongs to the Topical Collection: EPSA2019: Selected papers from the biennial conference in Geneva
Guest Editors: Anouk Barberousse, Richard Dawid, Marcel Weber
Rights and permissions
About this article

Cite this article
Brzović, Z., Balorda, V. & Šustar, P. Explanatory hierarchy of causal structures in molecular biology. Euro Jnl Phil Sci 11, 60 (2021). https://doi.org/10.1007/s13194-021-00380-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s13194-021-00380-7