
Abstract
The Devonian-Carboniferous transition is marked by major perturbations to the Earth’s climate system and marine redox conditions, a first-order mass extinction, and the succeeding marine ecosystem recovery, recording a sequence of environmental and biotic events. In South China, there exist numerous well-preserved and continuous Devonian-Carboniferous boundary successions that were formed in low-latitude passive margin basin, and this presents a unique opportunity to decipher the strata along a proximal to basinal transect using integrated biostratigraphical, event-stratigraphical, and geochemical approaches. Our geochemical data (δ13Ccarb, δ15Nbulk, δ238U and Mo/U enrichments, I/Ca ratio) indicated local and global C-N cycling perturbations and marine anoxia in the main phase of the Hangenberg Crisis interval (Middle Siphonodella praesulcata Zone), which coincided with a major regression in South China. Stromatoporoid biostromes and most typical Devonian faunas did not survive into the interval, and the deep-water black shales yield only opportunistic survivors, such as ammonoid Postclymenia cf. evoluta and bivalve Guerichina, along with miospores belonging to the LN Zone. The upper Hangenberg Crisis interval (Upper Si. praesulcata Zone, equal to the Protognathodus kockeli Zone) is marked by initial post-glacial transgression, a global δ13Ccarb spike, and opportunistic faunal blooms, coincident with conodont biofacies shift to the polygnathid-protognathodid biofacies. The Protognathodus fauna, nearly absent from platform facies, is often found in the condensed upper crisis interval of the basin, slope, and platform margin facies within the Youjiang Basin, although the abundance is extremely low and the phylogenetic lineage of Pr. meischneri-Pr. collinsoni-Pr. kockeli cannot be recognised in most successions. In South China, the first occurrence of Pr. kockeli seems to correspond to lithofacies changes, recorded in the first level of transgression just above the Hangenberg Sandstone event. Multiple lines of evidence, including great spatial heterogeneity of δ13Ccarb and δ15Nbulk records, extremes of redox proxies, and lack of macrofossils, suggest that marine habitats remained in a critical/turbulent state during the Upper Si. praesulcata Zone, triggering the opportunistic blooms and minor extinctions. Following the crisis intervals, radiation/recovery of many fossil groups did not occur until the upper part of the Siphonodella sulcata Zone to the Si. duplicata Zone. In particular, a negative shift of δ13Ccarb to pre-crisis values occurring near the base of the Si. sulcata/Pr. kuehni Zone could be recognised worldwide. The research history, current status, and achievement of Devonian-Carboniferous boundary in China are briefly reviewed.
Similar content being viewed by others

Introduction
During the Devonian and Carboniferous periods, China was separated into several blocks of continents and microplates with independent tectonic activities, scattered near the southeast margin of Siberia, Kazakhstan collage system, and in the peri-Palaeo-Tethys Ocean regions (Fig. 1a). These areas covered a wide range of sedimentary basins in both northern and southern hemispheres, providing a large scale perspective on Earth’ climate-environmental perturbations during the Devonian-Carboniferous transition and associated biotic response, facilitating recognition of the base of the Carboniferous.

a Palaeogeography locations of Chinese plates during the Late Devonian, palaeogeographic maps are from Ron Blakey (www2.nau.edu/rcb7) and b tectonic-stratigraphic regions of the Devonian and well-known Devonian-Carboniferous boundary sections in China (after Qie et al. 2019a) I. Altai-Hinggan; II. Junggar; III. Tarim; IV. North China; V. Qilian-Kunlun; VI. Qinling; VII. Qiangtang-Three Rivers; VIII. South China; IX. Himalaya-western Yunnan
In particular, the Qian (Guizhou)-Gui (Guangxi) area in South China (Fig. 1b) has figured prominently in past discussions of placing the boundary between the Devonian and Carboniferous systems, owing to its diverse faunas and faunal affinity with western Europe (Bai et al. 1994; Bai and Wang 1987; Yu 1988). The well-known Muhua II and Dapoushang sections, southern Guizhou, were recommended as candidate sections for the Devonian-Carboniferous boundary GSSP in the 1980s, as they represent complete deep-water basinal sequences, in which the evolutionary lineage from Si. praesulcata to Si. sulcata has been observed (Hou et al. 1985; Ji et al. 1989; Ziegler and Sandberg 1996). In February 1990, the IUGS ratified a proposal by the International Commission on Stratigraphy that the GSSP at La Serre, France, be used to define the base of the Carboniferous system, with the Hasselbachtal section in Germany and the Nanbiancun section in Guangxi, South China being auxiliary stratotype sections (Paproth et al. 1991; Fig. 1b).
Since the proposal of the GSSP for the Devonian-Carboniferous boundary (DCB), the choice of La Serre has received criticism due to unfavourable lithofacies and lack of a “true” phylogenetic transition from Si. praesulcata to Si. sulcata (Ji et al. 1989; Schönlaub et al. 1992; Ziegler and Sandberg 1996). Meanwhile, there are many difficulties in distinguishing the index conodont Si. sulcata from its ancestor, Si. praesulcata, owing to the existence of abundant transitional forms and morphotypes (Kaiser and Corradini 2011). As a result, the ICS established a task group to propose a new criterion for redefining the DCB and to search for a suitable section for the GSSP (Aretz 2010). In 2016 Montpellier DCB workshop, a new criterion “base of the Protognathodus kockeli Zone, beginning of radiation and top of major regression (top of the Hangenberg Sandstone event) and end of mass extinction” has been proposed by the task group, which need to be tested for global correlation.
Knowledge of bio-, chrono-, and chemo-stratigraphy of DCB in China continues substantially to improve as does our understanding of the Hangenberg events in various palaeogeographic settings (South China, Junggar and Tibet) (Carmichael et al. 2016; Liu et al. 2019a; Qie et al. 2015; Zong et al. 2015, 2020) and sedimentary facies (nearshore and offshore platform/shelf, slope, deep-water rift basin) (Ji 2004; Qie et al. 2016; Zhang et al. 2019a, 2020). This paper aims to (1) briefly review the research history, current status and achievement of DCB in China; (2) to illustrate representative DCB sections in South China; and (3) to test the “Montpellier criterion” for the Devonian-Carboniferous boundary redefinition.
Study history of DCB in China
The type areas for the Chinese regional Upper Devonian and lower Carboniferous chronostratigraphy are in central Hunan and southern Guizhou, South China, respectively (Qie et al. 2019a; Wang et al. 2019), and represented by mixed siliciclastic and carbonate platform successions with diverse benthic faunas (e.g. brachiopods, rugose, and tabulate corals and foraminifers). The traditional lower Carboniferous of China, i.e. the Fengningian System, included the Aikuan and Tatang groups (Ting 1931; Fig. 2), which correspond to the Tournasian and Visean, respectively. The Aikuan Group was further subdivided into the Gelaohe (Kolaohe) and Tangbagou (Tangpakou) series in ascending order (Ting 1931; Ting and Grabau 1936), containing two rugose coral zones (Cysto-phrentis and Uralinia (Pseudouralinia) zones) defined by Yu (1931), giving a much lower boundary for the base of Carboniferous in China (Figs. 2 and 3).


Upper Devonian to Middle Mississippian integrative stratigraphy framework of China. International standard chronostratigraphy units, conodont zones and absolute ages are after Becker et al. (2012) and Percival et al. (2018); Polarity Chron is after Gradstein et al. (2012) and Ogg et al. (2016); Chinese regional chronostratigraphy units (CRS) and Upper Devonian to Middle Mississippian biozonations of important fossil groups in China are after Qie et al. (2019a), Wang and Jin (2003), and Wang et al. (2019) and references therein. Note that the middle and upper Si. praesulcata zones is equivalent to the CKI and Protognathodus kockli zone of Kaiser et al. (2009), respectively; carbon isotopic record of carbonate rocks from the Lali, Naqing, Qilinzhai, Malanbian, and Pengchong sections in South China (data are compiled from Davydov et al. 2012; Yao et al. 2015; Qie et al. 2016; Zhang et al. 2019b and Chen et al. 2021). Solid line and dashed line, Locfit regression with 95% confidence interval
Since the late 1970s, major progresses have been made regarding the DCB in pelagic facies of South China, and important fossils groups that are biostratigraphically significant, including ammonite and conodont, have been constantly reported in deep-water Youjiang Basin (Ruan 1979; Wang and Wang 1978). Palmatolepis- and Siphonodella-based conodont zonations of Upper Devonian to lowermost Carboniferous were well-established (Hou et al. 1985; Ji and Ziegler 1993), and the Muhua II, Dapoushang, and Nanbiancun sections were successively recommended as candidate sections for the DCB GSSP (Hou et al. 1985; Yu 1988; Ji et al. 1989; Figs. 4 and 5). The DCB in pelagic facies of China has been defined to coincide with the first occurrence of Si. sulcata, slightly below the classic base of the Gattendorfia Zone (Hou et al. 1985; Hu et al. 2020; Ji et al. 1989).

The international auxiliary stratotype section of the Devonian-Carboniferous boundary at Nanbiancun, Guilin, China. a Section detail with the DCB at the base of bed 56. b Monument for the international auxiliary DCB stratotype section in the geopark. c Palaeogeography location of the Nanbiancun section. d Lithological log, ranges of important fossil groups, and carbon isotopic records in relation to the DCB at Nanbiancun (after Yu 1988 and this study)

The Muhua II DCB section at Muhua, Changshun, Guizhou. a Palaeogeography reconstruction of South China and locations of DCB sections. b Geological map of the Muhua area and locations of the Muhua sections and the Wangyou section. c Muhua II section detail with the DCB at the base of bed 23. d Lithological log, ranges of important fossil groups, and carbon isotopic records in relation to the DCB in the Muhua II section (Hou et al. 1985; Xu et al. 1986)
In contrast, the Devonian-Carboniferous boundary in shallow-water facies remained inconclusive due to major lithological changes, widespread sedimentary gaps, and lack of/or delayed occurrences of index fossils. A great controversy about the DCB in shallow-water facies at type areas for the Upper Devonian and the Fengningian System (lower Carboniferous) of China erupted in the late 1980s (Ji 1987; Wang 1987a; Wang and Yin 1985; Wu et al. 1981; Yang et al. 1980). By its end, several viewpoints have been widely accepted: (1) the rugose coral Cystophrentis Zone in South China should be assigned to the uppermost Famennian rather than to the Carboniferous, and the DCB should be moved upward around the base of the Tangbagou Formation; (2) an interval zone with rare macrofossils exists between the Cystophrentis and Uralinia (Pseudouralinia) zones following the end-Devonian biotic event; and (3) the DCB in shallow-water facies of China could be marked by the first appearance of the conodont Si. homosimplex or by the extinction of coral Cystophrentis and brachiopod Tenticospirifer, as well as by the disappearance of the foraminifer Quasiendothyra fauna (Qin et al. 1988; Tan 1987; Wang 1987a, b). During this time, an endemic shallow-water Siphonodella zonation has been proposed by Ji and Ziegler (1992), although precise correlation with standard pelagic Siphonodella zonation of Sandberg et al. (1978) is still debatable (Ji 2004; Qie et al. 2014, 2016).
According to the newest Carboniferous timescale of China (see Wang et al. 2019; Fig. 3), the lower boundaries of the Chinese regional Fengningian subsystem, Aikuanian series, and Tangbagouan stage coincided with the Devonian-Carboniferous boundary marked by the current GSSP definition, the FAD of conodont Si. sulcata. Due to the lithofacies and biofacies differences, the precise location of the DCB in the type area of the Chinese regional Tangbagouan Stage is still unclear. Qie et al. (2016) presented a high-resolution and integrated conodont biostratigraphy and δ13C stratigraphy framework for the Tangbagou Formation at its hypostratotype section, suggesting that the DCB should be placed near the base of the conodont Polygnathus spicatus Zone in the lower part of the Tangbagou Formation, and within the falling limb after the peak values of the Hangenberg carbon isotope excursion (HICE). This level is ~ 13 m above the base of the Tangbagou Formation at Qilinzhai, within the Cystophrentis-Uralinia Interval Zone, and slightly below the first occurrence of the conodont Si. homosimplex. However, Nie (2019) and Song and Gong (2019) indicated the DCB should correspond to major fauna changeover (e.g. brachiopods and ostracods), which is consistent with extinction of coral Cystophrentis and the boundary between Gelaohe and Tangbagou formations. Further work is needed to facilitate recognition of DCB in different sedimentary facies and regional stratigraphy correlation.
DCB sequences in South China
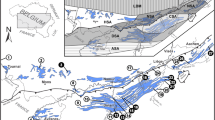
South China is the most important area for the study of the DCB in China, where all the stratotype sections for the Chinese regional stages of the Devonian and Carboniferous systems are situated (Qie et al. 2019a; Wang et al. 2019). The South China Block began to drift away from the north Gondwana margin since 400 Ma (Xian et al. 2019), leading to spreading of the eastern part of the Palaeo-Tethys Ocean and the formation of a passive continental marginal basin in its southwest part. The drift-rifting process is likely to have induced complex rifting-fault systems within the South China Block that, combined with tropical location and suitable climate, resulted in the development of extensive carbonate platforms separated by deep-water interplatform basins (Fig. 5a). As a consequence, numerous well-preserved DCB successions are recorded in various sedimentary settings (Qie et al. 2019a). Detailed studies of diverse fossil groups in the nearly complete marine sequences in South China make it an ideal area for the further improvement of Devonian and Carboniferous chronostratigraphic scale.
Nanbiancun, Guilin, Guangxi
The section at Nanbiancun near Guilin (GPS coordinates: 110°15′38′′ E, 25°19′44′′ N) is one of the international auxiliary stratotype sections for the current Devonian-Carboniferous boundary after a world-wide discussion (Paproth et al. 1991; Figs. 1, 4). In spite of diverse benthic and pelagic faunas and continuous carbonate deposition across the DCB, it was rejected as GSSP due to the unfavourable sedimentary facies and uncertainty about the FO of Si. sulcata (Gong et al. 1991; Ji 2004).
The succession at Nanbiancun was deposited on the north margin of the Guilin isolated carbonate platform during the Devonian-Carboniferous transition, and is characterised by a deepening upward trend as a result of intensified rifting. The Guilin Platform was separated from the Qian-Gui carbonate platform by a narrow deep-water rift basin and, thus, was rarely affected by the terrestrial input (Fig. 4). At this locality, the DCB succession contains predominantly limestones of the Rongxian (Yonghsien), Nanbiancun, and Chuanbutou formations with the best documentation of macrofossils across DCB in the world (Yu 1988). Fourteen fossil groups, conodonts, foraminifers, algae, corals, blastoids, bryozoans, trilobites, ostracods, brachiopods, ammonoids, gastropods, bivalves, holothuroids, and fishes, as well as sedimentological and geochemical characteristics, were well-illustrated by Yu (1988).
The Rongxian Formation (beds 40–48) is mainly represented by greyish white, medium-bedded, massive bioclastic/and intraclastic packetone to grainstone with bindstone intercalations. Abundant fragments of crinoids, corals, brachiopods, bryozoan, and trilobite are present, as well as microfossils such as conodonts, calcispheres, calcareous algae, and ostracods. The facies association and diverse fauna indicate deposition in an open-marine or restricted platform environment above or near fair-weather wave base. The presence of conodont Pa. gracilis expansa and Pa. g. gonioclymeniae points to the latest Devonian age (i.e. the Lower to Upper Pa. g. expansa zones).
The Nanbiancun Formation (beds 49–67) is 1.23 m thick and consists of coquina and bioclastic packstone and grainstone with several very thin clay layers (0.4–5 cm). The significant change in thickness of beds 66 and 67 within a relatively short lateral distance (Fig. 4) and occurrences of reworked fossils (e.g. Pa. g. sigmoidalis) suggested that deposition took place on the upper part of the slope or platform margin with slightly strong hydrodynamic condition. Brachiopods are major contributors to the bioclastic content, which could exceed 30% of the sediment. According to Yu (1988), the DCB is located at the base of the bed 56, marked by the FO of condont Si. sulcata. However, many researchers suggested that the DCB beds 54–57 represent a tempestite sequence, and hence, the boundary is placed in the middle of a bed/event depositing reworked material. A number of diversified faunas that are of correlative value, including conodont Pr. kuehni and ammonite Acutimitoceras sp., existed in the boundary bed (Fig. 4). At Nanbiancun, early siphonodellids are abundant and diverse, indicated by five morphotypes of Si. praesulcata and a great majority of specimens with intermediate features, although the Protognathodus fauna is relatively rare and restricted within the DCB interval (Fig. 4). Proto-gnathodus kockeli, the potential new index fossil for the DCB, enters at the base of bed 52, whereas the first Pr. meischneri and Pr. collinsoni occur in bed 54. Until now, the expected stratigraphic ordering of the first occurrences of early protognathodids (Pr. meischneri-Pr. collinsoni-Pr. kockeli-Pr. kuehni) is not recognisable at Nanbiancun.
The Chuanbutou Formation is 96-m thick at Nanbiancun, and its basal part consists of massive packstone and grainstone with hummocky cross-bedding and normal or reverse graded bedding, indicative of slope setting below the normal wave base and above the storm wave base. The lower Si. duplicata Zone to lower Si. crenulata Zone is presented for this formation (Yu 1988).
At Nanbiancun, the pre-Hangenberg Crisis intervals (beds 48–50) record a baseline δ13Ccarb value of ~ 2‰, and a minor positive δ13Ccarb shift with an amplitude of ~ 1‰ is observed in the Upper Si. praesulcata Zone (=kockeli Zone of Kaiser et al. 2009), which is consistent with the HICE. In the Si. sulcata Zone, δ13Ccarb values fluctuate between 1.7 and 2.8‰ and gradually fall back to and stay stable around 1.7‰ in the Si. duplicata Zone.
DCB sections in Muhua area
The Muhua area is situated at the northwest flank of the Wangyou dome anticline, about 100 km south of Guiyang, the capital city of the Guizhou Province (Fig. 5a, b). During the Devonian to Carboniferous transition, the Muhua area was located at the joint zone between the Qian-Gui Platform in the north and the Youjiang Basin in the south. Upper Devonian to lowest Carboniferous succession is exposed continuously and consists mainly of cephalopod-bearing lime mudstone, argillaceous limestone, and nodular limestone of the Wuzhishan Formation (= previously used Daihua Formation) and Wangyou Formation (Fig. 5). The lithofacies and fossil assemblage (ammonoids, conodonts, ostracods, and gastropods) point to a deep-water setting below the storm-wave base, with only minor inputs of silicilastics and bioclastics from adjacent Qian-Gui Platform.
Several well-known DCB sections, including the Muhua II, Dapoushang, Gedongguan, Muhua III, and Limushan sections, have been described in detail by Hou et al. (1985) and Ji et al. (1989), and equivalent successions can be found elsewhere in this area. A complete zonal succession with a rich fossil content, especially of the index groups of conodonts and ammonoids, exists in the DCB intervals. In the 1980s, both the Muhua II and Dapoushang sections were proposed as candidates for the basal Carboniferous GSSP (Paproth et al. 1991; Ziegler and Sandberg 1996) due to occurrences of complete evolutionary lineage from Si. praesulcata to Si. sulcata. Geochemical work has been published across the boundary at the Muhua II, Muhua III, Dapoushang, and Gedongguan sections (Xu et al. 1986; Ji et al. 1989; Bai et al. 1994; Qie et al. 2015; Fig. 6), and currently other investigations, including element geochemistry, sulphur isotopes, and astronomical cyclostratigraphy, are being undertaken.

In the Muhua area, Clymenia and Wocklumeria faunas were reported from the upper part of the Wuzhishan Formation, which is characterised by thin- to medium-bedded lime mudstone, bioclastic wackestone with occasional packstone and argillaceous limestone (Hou et al. 1985). Ammonoids, conodonts, and microvetebrates dominate the fossil assemblage, although a few ostracods, trilobites, calcispheres, and echinoderm fragments are also present. At the top of the Wuzhishan Formation, the synchronous extinction of typical Devonian faunas, including the ammonite Parawocklumeria paradoxa and the conodonts Po. vogesi, Ps. m. trigonicus, and Pa. gr. gracilis, marks the onset of the Hangenberg Crisis.
The overlying Wangyou Formation forms the uppermost Devonian to lowermost Carboniferous succession from the middle Si. praesulcata to Lower Si. crenulata zones, which is 4.2-m thick at Muhua II and 5.3-m thick at Dapoushang. The Wangyou Formation represents condensed deposition associated with intensified rifting. It consists of alternating lime mudstone, bioclastic wackestone, argillaceous limestone, and nodular limestone with intercalations of very thin (< 1 cm) clay layers, rich in conodonts, ammonoids, ostracods, and gastropods (Hou et al. 1985; Ji et al. 1989; Figs. 5 and 6). Notably, Gattendorfia was reported to occur in the Si. bransoni conodont Zone at both sections.
The basal 3–10 cm of the Wangyou Formation are occasionally characterised by black shale, and can change laterally into yellow mudstone and marlstone with small size limestone lenses. This level was thought to correspond to the Hangenberg shale in Rhenish Mountains, yielding diverse miospores (such as Vallatisporites pusillites, V. verrucosus, Samarisporites concinnus, Tumulispora dentata, T. rarituberculata, T. turgiduta, T. major, T. macrotuberculata, Archaeozonotriletes variabilis, Verrucosisporites cf. nitidus, V. depressus) and a few conodonts including Po. c. communis, Pr. collinsoni, Pr. meischneri, and Bi. a. aculeatus at Muhua (Hou et al. 1985). The miospore assemblage was thought to be correlated with the Retispora lepidophyta-Verrucosisporites nitidus (LN) Zone of Western Europe (Hou et al. 1985). At Dapoushang, this level (bed E) contains < 3% silt and yields the conodonts Pr. kockeli, Pr. collinsoni, Po. c. communis, Po. purus purus, and Sp. stabilis, which is in contradiction of previous interpretation that Pr. kockeli occurred in the aftermath of HBS and HSS events. The early occurrences of Pr. kockeli could be explained either by condensed sedimentation or by its true first appearance level in the middle Si. praesulcata Zone (=CKI of Kaiser et al. (2009)). Meanwhile, Liu et al. (2012) extracted zircons from the same interval (bed E) at Dapoushang to conduct SHRIMP U-Pb analyses, yielding 20 concordant data that form a cluster with a 206Pb/238U concordia age of 359.6 ± 1.9 Ma, indicating a latest Devonian age.
The lowermost 40–50 cm of the Wangyou Formation, regarded as the Gedongguan Bed by Hou et al. (1985), represents the DCB interval in Muhua area and yields a diverse early siphonodellid and protognathodid fauna (Hou et al. 1985; Ji et al. 1989; Fig. 6). In the Muhua II section, the stratigraphic ordering of the first occurrences of Pr. meischneri (bed 21)-Pr. collinsoni (bed 21 top, basal Wangyou Formation)-Pr. kockeli (bed 22)-Pr. kuehni (bed 22 top) seems to exist, and the DCB was fixed at the base of bed 24 based on the FO of Si. sulcata according to Hou et al. (1985). However, Ziegler and Sandberg (1984) reported the first Si. sulcata in bed 23 and proposed that the DCB should be located between beds 21 and 22 due to the presence of Pr. kuehni and Po. purus in bed 22. The first Si. sulcata in a level corresponding to bed 23 was also corroborated by Bai et al. (1994). In the Dapoushang section, many transitional morphotypes between Si. praesulcata and Si. sulcata occur in the ~ 40-cm-thick bed 0, and the DCB was placed by Ji et al. (1989) at the base of bed 1, yielding conodont association of Si. sulcata, Po. purus subplanus, Po. c. communis, and Bispathodus a. aculeatus. Recently, Nie (2019) restudied the conodonts from the Dapoushang section and reported Si. sulcata at the base of the bed 0, lowering the current DCB to the first limestone right above the HBS and HSS equivalent (bed E).
In Muhua area, the condensed deposits of uppermost Devonian to lowest Carboniferous hinder the precise recognition of the DCB. The equivalent of the HBS and HSS is only 3–10-cm thick, and the occurrences of Pr. kockeli are often documented in the first limestone bed immediately above the black shale. It is apparent that conodont biostratigraphy studies at finer resolution (1–2 cm in height) should be performed in these successions to reveal the phylogeny sequences of the early siphonodellids and protognathodids fauna.
The Qian-Gui Platform to the Youjiang Basin transect
In South China, numerous well-preserved DCB successions, recording various lithofacies and biofacies, have been the subject of careful and detailed studies (Qie et al. 2016; Nie 2019; Song and Gong 2018, 2019; Zhang et al. 2019a; Figs. 7, 8, and 9). By documenting sedimentological, palaeontological, and geochemical evidences at multiple stratigraphic sections spanning a proximal to basinal transect in southern Guizhou and northern Guangxi, we will be able to reconstruct the pattern of end-Devonian mass extinction and climate-environmental perturbations associated with the Hangenberg Crisis, and finally, to find scientifically meaningful as well as practical criteria for the base of the Carboniferous system.

a Geologic map of Upper Devonian and lower Carboniferous and DCB section localities in southern Guizhou and northern Guangxi. b Sedimentary facies and models of the DCB intervals at four sections in a proximal to basinal transect. c Facies and conodont-based correlations of the four DCB sections (Qilinzhai, Xiasi, Jiarantang and Duli). Abbreviations: D3, Upper Devonian; D3-C1z-g-t, the Zhewang, Gelaohe/Etoucun, and Tangbagou formations; C1, Lower Carboniferous

Important shallow-water conodont elements from the DCB intervals in South China characterised by the polygnathid-clydagnathids biofacies of the upper Hangenberg Crisis interval and the endemic siphonodellids biofacies of lower Tournaisian (Qie et al. 2014, 2016 and new collections). ‘a’ for upper view, ‘b’ for lower view, ‘c’ for lateral view. 1–2, 4 Po. co. communis Branson and Mehl, 1934. 1 specimen CQL14020006 from sample 2/1, 2 specimen CQL14020007 from sample 1/1, Qilinzhai, 4 specimen CLA 702302 from sample 2/3, Long’an, 3 Po. co. dentatus Druce, 1969. specimen CQL14020008 from sample 5/1., Qilinzhai, 5 Po. co. cf. dentatus Druce, 1969, specimen CLA 704401 from sample 4/4, Long’an, 6, 8 Cl. gilwernensis Rhodes, Austin and Druce, 1969. 6 specimen CQL14020010 from sample 10/1, Qilinzhai, 8 specimen CLA 706201 from sample 6/2, Long’an, 7. Cl. unicornis Rhodes, Austin and Druce, 1969. 11 specimen CQL14020011 from sample 10/1., Qilinzhai, 9, 14. Po. spicatus E. R. Branson,1934. 9 specimen CQL14020017 from sample10/1, Qilinzhai, 14 specimen CLA 705301 from sample 5/3, Long’an, 10–12 Siphonodella praesulcata Sandberg et al., 1972. 10 specimen CQL14020004 from sample 15/2, 11 specimen CQL14020002 from sample 10/1, Qilinzhai, 12 specimen CLA 705201 from sample 5/2, Long’an, 13. Si. sulcata (Huddle), 1934. specimen CLA 706101 from sample 6/1, Long’an, 15, 19. Si. levis (Ni), 1984. 15 specimen CQL14020021 from sample 10/2, Qilinzhai, 19 specimen XS15.8-001 from sample XST15.8, Xiasi, 16–17 Si. homosimplex Ji and Ziegler 1992. 16 specimen CQL14020022 from sample 10/2, Qilinzhai, 17 specimen CLA708301 from sample 8/3, Long’an, 18 Rhodalepis sp. specimen XS5.6-005 from sample XST5.6, Xiasi

Stratigraphically important deep-water conodont elements from the DCB intervals in Youjiang Basin, South China characterised by the polygnathid-protognathodid biofacies of the upper Hangenberg Crisis interval and the siphonodellids biofacies of lower Tournaisian (Muhua, Hou et al. 1985; Jiarantang, Zhang et al. 2019a; and new collections near Jiarantang and at Duli). ‘a’ for upper view, ‘b’ for lower view. 1-4 Protognathodus kockeli (Bischoff), 1957. (1) specimen DC84420 from sample GMII-28, Muhua, (2) specimen DC84422 from sample GMII-24, Muhua, (3) and (4) from sample XSJ-1, Jiarantang. 5 Protognathodus meischneri Ziegler, 1969. from sample XSJ15-1, Jiarantang, 6-7 Protognathodus collinsoni Ziegler, 1969, (6) specimen SHB03001 from sample SHB-3, near Jiarantang, (7) specimen SHB03002 from sample SHB-3, near Jiarantang, 8-9 Siphonodella praesulcata Sandberg et al., 1972. (8) specimen SHB10001 from sample SHB-10, near Jiarantang, (9) specimen SHB08001 from sample SHB-8, near Jiarantang, 10 Si. sulcata (Huddle), 1934. specimen DCDL1395001 from sample DCDL1395-1400, Duli, 11 Si. praesulcata-Si. sulcata transitional element. specimen SHB21001 from sample SHB-21, near Jiarantang, 12 Si. bransoni. specimen DCDL1400001 from sample DCDL1400-1450, Duli
The four stratigraphic successions examined in this study consist mainly of limestone, mudstone, and shale lithofacies that are grouped into nearshore mixed siliciclastic-carbonate platform, offshore carbonate platform, slope, and basinal facies associations, respectively (Fig. 7). In general, the Duli section contains the largest proportion of deep-water limestone facies with minor black shales, and the shallowest water facies are most abundant in the Qilinzhai section (Fig. 7b, c).
Nearshore mixed siliciclastic-carbonate platform facies
At Qilinzhai, the DCB successions comprise the upper part of the Gelaohe Formation and the lower part of the Tangbagou Formation (Qie et al. 2015). The underlying Gelaohe Formation is mainly composed of calcareous mudstone with minor intercalations of lime mudstone, bioclastic wacke- to pack-stone and black shale, yielding abundant and diverse brachiopods such as Globosochonetes sp., Plichonetes sp., Yanguania dushanensis, Xinshaoproductus quadrata, Schuchertella gelaoheensis, Schuchertella sp., Ripidomella sp., Ptychomaleotoechia kinlingensis, Cleiothyridina submabranacea, Cystospirifer sp., and Crurithyris sp. (Nie, 2019). Rugose coral Cystophrentis, tabulate coral Syringo-pora, as well as many unidentified ostracods, echinoderms, bryozoans, gastropods and foraminifers are also important contributors to this benthic assemblage.
The basal 12.8 m of the overlying Tangbagou Formation, unit A of Qie et al. (2016), consists of nearly pure limestone, and siliciclastic materials are restricted to trivial and thin (3–10 mm) marl partings. This interval, recording peak δ13Ccarb values up to 4.5‰, is devoid of any macrofossils and assigned to the conodont Clydagnathus gilwernensis-Cl. unicornis Assemblage Zone, might be correlated to the upper Si. praesulcata Zone of the pelagic facies (Qie et al. 2015, 2016). It probably represents the survival interval following the main phase of the Hangenberg Crisis and yields conodonts Cl. glilwernensis, Po. c. communis, Po. c. dentatus, Po. i. inornatus, and Si. praesulcata (Fig. 8). This shallow-water facies association contains thin-bedded lime mudstone and skeletal, peloidal wackestone with locally abundant intra-clasts. Skeletal grains include calcispheres, ostracods, and fragments of gastropods and very thin brachiopod shells. The unit B of Qie et al. (2016) is composed of alternating mudstone, marlstone, argillaceous limestone, and limestone, yielding diverse benthic and nektonic faunas (e.g. brachiopods, rugose/and tabulate corals, foraminifers, ostracods and conodonts). Especially, an endemic shallow-water Siphonodella fauna, including Si. homosimplex, Si. levis, Si. sinesis, and Si. eurylobata, associated with a few morphotypes of early siphonodellids (Fig. 8), corresponds to the Si. duplicata Zone to the lower Si. crenulata Zone of the pelagic facies (Qie et al. 2016).
The lithological boundary between the Gelaohe and Tangbagou formations is consistent with the Hangenberg extinction event, characterised by the sudden demise of the typical Devonian rugose corals, brachiopods and foraminifers. Because of the glacio-eustatic sea-level fall associated with the end-Devonian Gondwanan glaciation, the DCB successions are not well-represented in the shallow-marine sedimentary record, and many DCB sections probably contain hiatuses. A carbon isotopic discontinuity separates the top of the Gelaohe calcareous mudstones from the overlying Tangbagou limestone, suggesting that the middle Si. praesulcata Zone, i.e. the main phases of the Hangenberg Crisis, is probably missing.
Offshore carbonate platform facies
On the shallow-water Qian-Gui Platform, the stratigraphic expression of the DCB successions varies along the distance from the palaeo-shoreline (Fig. 7). Thin-bedded to massive limestone and dolostone with few siliciclastic sediments dominate the offshore platform and platform margin facies, whereas more muddy facies occur in the north. In the Xiasi section, the DCB successions have been well-studied in terms of lithofacies and foraminifer biostratigraphy (Hance et al. 2011), and contains nearly pure limestones of the Etoucun and Tangbagou formations with the most diverse local stromatoporoid fauna just prior to the Hangenberg Crisis. It is situated about 40 km south of the Qilinzhai section, 800 m east of the Xiasi Town (GPS coordinates: 107°26′20.28′′ E, 25°28′0.15′′ N), and represents an open-marine platform setting near the fair-weather wave base.
The uppermost Devonian Etoucun Formation, consisting of thick-bedded to massive peloidal wacke- to grainstones and boundstones, is about 80-m-thick and contains diverse benthic fauna, such as stromatoporoids, rugose/and tabulate corals, gastropods, and foraminifers. Near its top, a 2.3-m-thick stromatoporoid biostrome exists, representing the final resurgence of stromatoporoid reefs in South China prior to the end-Devonian Hangenberg extinction (Fig. 7). Gerronostroma species are the most important reef-forming organisms, accounting for 50–70% of the rock volume.
The basal Tangbagou Formation is composed of greyish thin-bedded lime mudstones and peloidal wackestones devoid of macrofossils. It yields only a few foraminifer Bisphaera irregularis and Earlandia spp. of MFZ1 (Hance et al. 2011), as well as long-ranging, shallow-water conodonts, such as Polygnathus communis communis, Po. c. dentatus, and Po. inornatus. The early Tournaisian shallow-water Siphonodella fauna did not occur until the lower part of the Tangbagou Formation (Fig. 7).
Slope facies
Further south, the Jiarantang section (GPS coordinates: 107°25′35.07′′ E 25°25′12.04′′ N) is located in the transition zone between the Qian-Gui Platform and the deep-water Youjiang Basin. It was first described by Feng (1977), and recently re-studied by Zhang et al. (2019a) concerning the cymaclymeniid ammonoids that was first reported in the Changshun Shale (equivalent of the Hangenberg Black Shale) of South China.
The late Famennian sequence is composed of thin- to thick-bedded argillaceous, and dolomitic limestones of the Rongxian Formation, indicative of platform margin facies (Zhang et al. 2019a). The carbonate sedimentation is interrupted by a 1.5-m-thick black shale (Changshun Shale) yielding Postclymenia cf. evoluta that is closely to forms in contemporaneous Hangenberg Black Shale of the Rhenish Massif, Germany. The bivalve Guerichia sp. and the brachiopod Semicostella? sp. were also recorded in this unit (Zhang et al. 2019a). Above the thin- to thick-bedded bioclastic pack- to grainstone with minor intercalations of black shale of the Wangyou Formation yield a great number of diversified micro- and macro-faunas of stratigraphic value. The conodonts include Pr. kockeli, Pr. meischneri, and Si. praesulcata, which also occur immediately above the Hangenberg Black Shale and its equivalents in various pelagic facies sections from the Muhua area (South China), the Rhenish Massif (Germany), the Carnic Alps (Austria and Italy), and the Moravian Karst (Czech Republic) (Kumpan et al. 2021, this issue). A few specimens of the brachiopod Syringothyris were found by Zhang et al. (2019a) in the bioclastic limestones, which also appears in the DCB successions of western Junggar, representing a newly emergent brachiopod fauna following the Hangenberg Crisis (Zong and Ma 2012). Skeletal grains are fossil fragments that were derived from the Qian-Gui Platform and probably reworked by distal storm/or turbidity currents, and rare reverse-graded packstone and grainstone indicate debris flow deposits.
Deep-water basinal facies
The basinal DCB succession is well-exposed in the Duli section (GPS coordinates: 107°20′ 05.5′′ E 25°23′ 50.1′′ N) and crops out along the X921 road, near the Duli village of Nandan County, Guangxi (Kalvoda et al. 2019; Nie 2019). The stratigraphic sequence is basically the same as its equivalent in Muhua area, with a 10~20 cm-thick black shale separating the cephalopod-bearing Wuzhishan lime mudstone from the overlying Wangyou nodular and banded limestone (Fig. 7). The uppermost part of the Wuzhishan Formation (13.6-m thick) consists mainly of medium- to thick-bedded lime mudstone, bioclastic wackestone with rare argillaceous limestone, developing pelagic conodonts of the lower praesulcata Zone, such as Pa. g. gracilis, Pa. g. sigmoidalis, Pseudopolygnathus marburgensis trigonicus, and Spatho-gnathodus sp.
At the basal Wangyou Formation, the organic-rich (TOC up to 3.37%) black shale with Hg spike contains abundant detrital zircons (> 200 grains per kg) and is correlated with the Hangenberg Black Shale and the Hangenberg Sandstone of the Rhenish Massif (Kalvoda et al. 2019). The Wangyou Formation (5.3-m thick) is mainly composed of thin-bedded argillaceous limestone, marlstone, lime mudstone, and nodular limestone with abundant and diverse conodonts, spanning from the upper Si. praesulcata Zone to the lower Si. crenulata Zone (Fig. 7). Note that the Protognathodus fauna is extremely rare at Duli, and Pr. kockeli, the index fossil for the upper Si. praesulcata Zone, was only recovered in the Si. bransoni Zone together with Pr. kuehni (Nie 2019), and the FO of typical Si. sulcata is at ~ 20 cm above the top of the black shale. Towards the south, the Wangyou Formation interfingers with and is replaced laterally by the Luzhai Formation, which are dominantly black shales and siliceous rocks with abundant pyrites. The observed features of the fine-grained, condensed limestones that make up the basinal facies association point to deposition in quiet water below the storm-wave base.
Geochemical signatures around the DCB in South China
The Devonian-Carboniferous transition was associated with significant perturbations of the geochemical cycles of the ocean, as indicated by time-varying signals of carbon, nitrogen, and uranium isotope and element geochemistry records (Brand et al. 2004; Buggisch and Joachimski 2006; Kaiser et al. 2006, 2016; Kalvoda et al. 2019; Liu et al. 2016, 2019a, b; Qie et al. 2015; Zhang et al. 2020). They allow for high-resolution regional and intercontinental correlation of the DCB intervals.
Stable carbon isotope geochemistry
Carbon isotopic records of whole rock carbonates have been reported from many DCB sections in South China (Figs. 3, 4, 5, 6, 10). However, due to the prevalent development of depositional hiatus, condensed sedimentation, and siliciclastic intercalations, the architecture of the δ13Ccarb curve in the lower Hangenberg Crisis interval (middle Si. praesulcata Zone) is not completely illustrated. At Long’an and Malanbian, a distinct decline in δ13Ccarb occurs near the base of the middle Si. praesulcata Zone, consistent with the start of the Hangenberg extinction and sea-level fall event, and is probably caused by respiration of organic matter contributing abundant 12C-enriched dissolved inorganic carbon to the shallow-water platform settings (Qie et al. 2015; Fig. 10). The deep-water DCB successions at Muhua, Gedongguan, Dapoushang, and Lali, also record a minor decline at the correlative level (Fig.6; Zhang et al. 2019b), which might be related to remineralisation of organic matters from organic carbon-rich sediments. Meanwhile, a similar pattern has been observed from the deep-water successions at Grüne Schneid (Carnic Alps), Trolp (Graz Paleozoic), and Křtiny (Moravian Karst) (Kaiser et al. 2006, 2008; Kumpan et al. 2014).

In the upper Hangenberg Crisis interval (upper Si. praesulcata Zone), all the shallow-water DCB successions (Qilinzhai, Malanbian, Long’an, Nanbiancun) exhibit a distinctive positive δ13Ccarb shift (Figs. 4, 10), indicating a vigorous biological pump existed in the low-latitude South China marginal sea following the latest Devonian glaciation (Qie et al. 2015). The Hangenberg carbon isotope excursion (HICE) was previous reported from Montagne Noire, Carnic Alps, Graz Paleozoic, Holy Cross Mountains, Moravian Karst, Namur-Dinant Basin, and mid-continent of North America, with peak values around 2.5–6‰ (see Kaiser et al. 2016, and references therein). In Iowa, δ13Ccarb excursion reaches 6.8‰, and this shift represents one the of largest Devonian δ13C events (Buggisch and Joachimski 2006; Cramer et al. 2008). In South China, the peak δ13Ccarb values and the magnitude of the HICE vary significantly in different sedimentary settings and are substantially larger and more variable in the shallow-water platform sections relative to the deep-water basin facies (Qie et al. 2015). At Qilinzhai, δ13Ccarb values show a major positive shift increasing by > 4 to 4.5‰, while in the deep-water successions (Muhua, Dapoushang and Gedongguan), δ13Ccarb values fluctuate around 2‰ and show only a minor shift with an amplitude of ~ 1‰ (Figs. 6, 10). Following the HICE peak, δ13Ccarb values start to decline near the base of the Si. sulcata Zone of the pelagic facies and the Po. spicatus Zone of the platform facies. This temporal and spatial pattern of carbon isotope compositions in the South China marginal sea is known for the DCB strata from the Belgian-France Basin and Carnic Alps and has great global significance (Qie et al. 2015, 2016).
Nitrogen isotope geochemistry
Nitrogen is an essential nutrient for all organisms, and its cycling in the ocean is closely related to the microbial community associations, nutrient status of seawater, and marine redox condition (Falkowski 1997; Gruber and Galloway 2008), and thus, perturbations to marine nitrogen isotopes are likely to have significant implications for the end-Devonian mass extinction, microbial processes, and climate changes. Liu et al. (2016) reported nitrogen isotope compositions of three DCB sections of shallow platform facies from South China: Qilinzhai, Malanbian, and Long’an (Fig. 10). These data show that in the middle Si. praesulcata Zone, δ15Nbulk decrease from 4.5 to 2.5‰ at Malanbian and 4 to 0‰ at Long’an, and this perturbation is consistent with a negative shift of δ13Ccarb values and the main phase of the Hangenberg mass extinction. The shift to lower δ15N values suggests intensive denitrification and, thus, nitrogen fixation as the main source of biologically available nitrogen for primary producers in response to globally expanded marine anoxia (Liu et al. 2016). In the upper Si. praesulcata Zone, however, there exists significant spatial and temporal variation in δ15Nbulk values associated with different carbon isotopic compositions. δ15Nbulk values range from 3 to 6‰ at Qilinzhai and from 1 to 3‰ at Long’an and remain relatively constant around 3‰ at Malanbian. This spatial heterogeneity is most likely to be related to restricted water mass circulation, resulting in various nutrient inventories and redox conditions in different sedimentary settings of the South China marginal sea (Liu et al. 2016).
Redox proxies
In South China, several proxies (e.g. enrichment of Mo and U, I/Ca and δ238U values) have been applied to reconstruct the local and global redox conditions during the Devonian-Carboniferous transition (Kalvoda et al. 2019; Liu et al. 2019b; Zhang et al. 2020). In the Duli section, the trace-metal abundances indicate that the redox conditions varied between oxic and suboxic in the deep-water Youjiang Basin, and the sediments of the Hangenberg Crisis interval did accumulate under oxygen deficiency conditions (Kalvoda et al. 2019).
Carbonate I/Ca ratio is mostly controlled by changes in IO3- concentrations in the local upper water column and has been shown to be a reliable proxy that responds to marine redox variations over a wide range of geological periods (Lu et al. 2010; Hardisty et al. 2017; Lu et al. 2018). At Malanbian, the ratio of I/Ca values decreases abruptly from > 1 to ~ 0 μmol/mol across the transition from the lower to upper Si. praesulcata Zone, stays constant within the upper Hangenberg Crisis interval, and starts to increase close to the base of the overlying Si. homosimplex Zone. At Long’ an, a similar I/Ca profile is observed, showing a major I/Ca decline near the base of the upper Si. praesulcata Zone, nadir values around 0.16 μmol/mol within the upper Hangenberg Crisis interval, and an increase to 0.97 μmol/mol near the base of the Si. sulcata Zone. The decline of I/Ca values observed in the two shallow-water platform sections suggests that deoxygenation was intensified during the upper Si. praesulcata Zone, and the hypoxia might have expanded to shallow-water depths, at least in the South China area during the D-C transition. It is very likely that the intensified oxygen deficiency is a direct factor for the prolonged Hangenberg Crisis, and poor environmental conditions slowed full recovery of marine ecosystems in the post-extinction intervals.
Since the residence time of U (~ 400 kyr) in modern ocean is significantly longer than ocean mixing times (< 1.5 kyr) and U isotopes are sensitive to marine redox conditions, δ238U values from open-marine limestones were commonly used to place quantitative constraints on global marine redox chemistry changes during critical time intervals (Dahl et al. 2014; White et al. 2018; Zhang et al. 2019c). In the Long’an section, δ238U values of nearly pure limestones were measured, and this is the first δ238U profile for the Hangenberg Crisis (Zhang et al. 2020). δ238U values fluctuate between − 0.82 and 0.32‰ and exhibit three negative excursions interrupted by two positive shifts (Fig. 10). A global uranium cycle model was used to reconstruct the timing, duration, and extent of global marine redox changes during the end-Devonian Hangenberg Crisis, suggesting that marine anoxia expanded intensely in the latest Devonian oceans, covering the majority of global continental shelves, and > 5% of total global seafloor extent (Zhang et al. 2020).
DCB sequences in Junggar and Tibet
DCB in western Junggar
In western Junggar, the DCB successions are formed in an island arc complex within the central Asian Orogenic Belt (CAOB, Fig. 1) and mainly characterised by tuffaceous coarse sandstones and calcareous sandstones with minor bioclastic limestone lenses of the Hongguleleng Formation and the overlying calcareous tuffaceous siltstones and tuffs of the Heishantou Formation (Xu et al. 1990; Zong et al. 2020). This interval yields diversified fossil fauna and flora, consisting of conodonts, brachiopods, cephalopods, trilobites, echinoderms, radiolarians, rugose corals, bivalve, gastropods, miospores, acritarchs, and plant remains; however, precise biostratigraphic control for the DCB remains a matter of debate (Zong et al. 2015, 2016, 2020). Meanwhile, observations concerning the Hongguleleng and Heishantou formations as well as their lateral correlation have produced different interpretations (details see Suttner et al. 2014; Zong et al. 2020 and references therein).
Recent studies on ammonoids, brachiopods, trilobites, and geochemistry have provided better constraints for the subdivision of lithological units and precise location of the DCB in this area (Carmichael et al. 2016; Zong et al. 2015, 2016, 2020). Sedimentary petrology and multiple whole rock geochemical proxies revealed that the Hangenberg Event represents a regression/transgression sequence along with varied oxic/dysoxi/anoxic conditions in western Junggar, and the DCB was probably located within the Heishantou Formation at Bulongguoer (Carmichael et al. 2016), which is equal to the uppermost part of the Hongguleleng Formation of Zong et al. (2015) and Zong et al. (2020). Seven ammonoid levels/assemblages and five brachiopod assemblages were recognised from the Famennian to early Tournaisian successions (Zong et al. 2015, 2016), and occurrences of ammonoid Cymaclymenia-“Mimimitoceras” assemblage and few last cymaclymeniids as well as brachiopod Syringothyris fauna indicate the Hangenberg Crisis interval (middle to upper Si. praesulcata Zone) exists around the boundary between the Hongguleleng and Heishantou formations. The presence of conodont Pr. collisoni and Pr. meischneri at the Emuha section has been reported by Xu et al. (1990) from the top of the Hongguleleng Formation, while Tournaisin Siphonodella cooperi was found in the lower part of the Heishantou Formation, although no plates were illustrated. Based on integrated studies on several DCB sections (e.g. Bulongguoer, Gennaren, Wulankeshun, Chawangtaogaile and Yangzhuang West), Zong et al. (2020) suggest that the DCB should be placed between the disappearance level of the trilobite Omegops and the appearance level of the Belgibole-Conophyllpsia assemblage zone, consistent with the boundary between the Hongguleleng Formation and Heishantou Formation. At the top of the Hongguleleng Formation, a suite of coarse-grained clastic rocks was recognised by Zong et al. (2020) as the “Yangzhuang Sandstone” and considered to be equivalent of the Hangberng Sandstone caused by end-Devonian glacio-eutatic fall, which need further corro-boration.
DCB sections in Tibet
Due to remoteness, high altitude, and extensive mountainous area, a high-resolution stratigraphy and biostratigraphy concerning the DCB in Tibet remain lacking. Late Devonian to early Carboniferous conodonts from Himalayan terrane were first reported by Qiu (1984) and Rao and Yu (1985), who recovered the Protognathodus fauna and Si. sulcata from the middle part of the Yali Formation in Nyalam and Tingri. Subsequently, five conodont zones, including the Si. praesulcata Zone, Si. sulcata Zone, Si. duplicata Zone, the Si. isosticha Zone, and the Gnathodus pseudosemiglaber Zone, were recognised in the Yali Formation near the Yali village in Nyalam County, and the DCB was placed at the FO of Si. sulcata within the Golongri Member, ~ 10 m above the base of the Yali Formation (Qiu 1988). Pr. kockeli was also reported from the lower part of the Yali Formation, but it should belong to Pr. collisoni-Pr. kockeli transitional element judging from the plates of Qiu (1988), and currently conodont taxonomic work is being undertaken. At Yali, miospores have been intensively studied by Gao (1988) and Liu et al. (2019a), and most part of the Zhangdong Formation was assigned to the Retispora lepidophyta-Verrucosisporites nitidus (LN) Zone. The Zhangdong Formation is ~ 200 m thick, mainly consists of dark grey, greyish green silty shale, and shale, and represents an equivalent of the Hangenberg Black Shale in northeastern Gondwana.
Environmental changes and correlation events during the Devonian-Carboniferous transition in China
The end-Devonian Hangenberg Crisis eliminated about ~ 21% of marine invertebrate genera, causing major faunal changeovers and demise of stromatoporoid reef ecosystems, marking the end of the Devonian Period (Sepkoski 1996; Walliser 1996). It is well-represented by the latest Famen-nian Hangenberg beds, recording a sequence of environmental and biological events (Kaiser et al. 2008, 2016; Qie et al. 2015, 2019b).
In the pre-Hangenberg Crisis intervals of South China, long-lasting transgressive tendency persisted from the upper Pa. g. expansa Zone to the lower Si. praesulcata Zone associated with well-oxygenated facies regime on the shallow platforms and within the deep-water Youjiang Basin (Kalvoda et al. 2019; Liang et al. 2019; Liu et al. 2019a; Ma et al. 2009). Diverse benthic, nektonic, and planktonic assemblages were found in various habitats of the South China marginal sea, and in particular, several dm- to m-thick, low-relief stromatoporoid reefs are recorded in the Etoucun Formation on the offshore and isolated carbonate platforms (Liang et al., 2019).
In South China, the main phase of the Hangenberg Crisis usually coincided with abrupt lithofacies changes and major biogeochemical cycling perturbations (Liu et al. 2016; Qie et al. 2015). With regard to conodont zonation, it starts at the base of the revised Middle Si. praesulcata Zone, and probably coincides with the boundary between the ammonoid Wocklumeria/Postclymenia zones, with the boundary between foraminifer DFZ7 and MFZ1, and with the miospore zone boundary between the LE and LN (Fig. 3). Within the deep-water Youjiang Basin, the late Famennian cephalopod-bearing limestone is sharply separated from the succeeding black shales, which is usually 2–10-cm thick, and could range up to 1.5 m at Jiarantang in the slope facies (Figs. 6 and 7). Most typical Devonian faunas did not survive into the black shales, and these yield only opportunistic survivors, such as ammonoid Postclymenia cf. evoluta and bivalve Guerichina, along with miospores belonging to the LN Zone (Hou et al. 1985; Zhang et al. 2019a). Contemporaneous black shales can be recognised in many sections around the globe, particularly in western and southeastern Laurussia, northern Africa, Siberia and Himalaya (see Kaiser et al. 2016 and references therein; Liu et al. 2019a), arguing that marine anoxia might play an important role in the Hangenberg mass extinction. This interpretation is supported by the evolution of local and global marine redox chemistry changes using nitrogen isotopes of organic matters and U isotopes of marine limestones from South China (Liu et al. 2016; Zhang et al. 2020). The black shale interval in Youjiang Basin is correlated to the Hangenberg Black Shale and Hangenberg Sandstone in the Rhenish Massif. It contains abundant miospores and detrital zircons from land and was probably due to a short regressive pulse in association with a cooling episode in the southern hemisphere as documented in glaciogenic deposits in basins of Brazil, Bolivia, and Peru and eastern North America (Brezinski et al. 2008; Caputo 1985; Isaacson et al. 2008). On shallow-water platforms of South China, a maximal lowstand is demonstrated by depositional hiatus in nearshore facies at Qilinzhai, and by miospore correlation with prograding siliciclastic sediment at Malanbian, and in more offshore direction to the south, by evidence of condensed sedimentation (Qie et al. 2015). The abrupt changes of lithofacies and sedimentary conditions coincided with benthic faunal turnovers (e.g. foraminfer and rugose coral) and disappearance of framework-building clades (i.e. stromatoporoids and tabulate corals), leading to the demise of latest Devonian low-relief metazoan biostromes. In western Junggar, the glacio-eustatic origin of this regression is indicated by the occurrence of the “Yangzhuang Sandstone” intercalated between the Hongguleleng and Heishantou formations in an island arc context, corresponding to the disappearance of the trilobite Omegops fauna (Zong et al. 2020).
The upper Hangenberg Crisis interval (upper Si. prae-sulcata Zone) is marked by initial post-glacial transgression, a global δ13Ccarb spike, and opportunistic faunal blooms (Kaiser et al. 2016; Qie et al. 2015). Within the Youjiang Basin, the resumption of carbonate sedimentation occurs immediately above the black shale and is coincident with conodont turnover from the palmatolepid-bispathodid biofacies to the polygnathid-protognathodid biofacies (Fig. 6). The Protognathodus fauna is often found in the condensed upper crisis interval of the basin (Muhua area), slope (Jiarantang), and platform margin to upper slope facies (Nanbiancun) (Fig. 9), although the numbers are extremely low and the stratigraphic ordering of Pr. meischneri-Pr. collinsoni-Pr. kockeli cannot be recognised in most successions (Figs. 4, 6, 7). In South China, the first occurrence of Pr. kockeli seems to correspond to lithofacies changes, immediately atop the Hangenberg Black Shale and its equivalents in slope and basin environments, but this might be due to condensed sedimentation. On shallow-water platforms, the Protognathodus fauna is nearly absent, and this event interval yields a fauna dominated by Polygnathus communis communis, Polygnathus communis dentatus, Clydagnathus unicornis, Cl. gilwernensis, Bispathodus aculeatus, and Pseudopolygnathus sp., with several morphotypes of early siphonodellids (Fig. 8) (Ji 2004; Qie et al. 2014, 2016). In Himalaya region and western Junggar, some species of Protognathodus were reported from shallow-water successions (Qiu 1988; Zong et al. 2015). The distribution pattern of Protognathodus could be explained by its inhabiting cooler waters at a certain depth or higher latitude, thereby accounting for its absence from extensive low-latitude shallow-waters from South China and other places. Our data support the interpretation of Kaiser et al. (2016) that the bloom of the Protognathodus fauna in the crisis interval is a signal of opportunistic palaeoecology, in association with climatic perturbations and persisted marine anoxia (Liu et al. 2019b; Zhang et al. 2020; Fig. 10). Following the latest Devonian glaciation, a vigorous carbonate factory renewed as indicated by the resumption of carbonate sedimentation, global HICE, and pronounced vertical gradient of δ13C in the South China marginal sea; however, environmental conditions were still poor during the Upper Si. praesulcata Zone, which led to the delayed recovery of foraminifera and rugose corals in the lower Tournaisian (possibly in the Si. sandbergi Zone). For most adaptable groups, such as conodonts and ammonoids, the early phase of re-radiation was short-lived, and there were further extinctions near the base of the Si. sulcata Zone, and a similar pattern has been well-illustrated by Kaiser et al. (2016).
In post-crisis interval, a continuous sea-level rise from the Si. sulcata Zone through the Si. jii Zone is indicated by the aggradational successions in South China (Qie et al. 2015). Both the conodont polygnathid-protognathodid biofacies in deep-water environment and polygnathid-clydagnathids biofacies in shallow-water environment shifted to the siphonodellid biofacies. In South China, early siphonodellids occur in pelagic limestones with greater numbers and diversity than protognathodids, and also in shallow-water carbonates (Figs. 8, 9), suggesting that they inhabited near-surface waters and have great potential to make precise stratigraphic correlation. However, the taxonomic complexity of early siphonodellids, as revealed by previous works of Ji (2004) and Nie (2019), hinder the precise assignment of conodont zonation around the DCB in South China, and needed to be revaluated.
Summary
A new criterion “base of the Protognathodus kockeli Zone, beginning of radiation and top of major regression (top of the Hangenberg Sandstone event) and end of mass extinction” has been proposed by the majority of ICS task group members to redefine the base of the Carboniferous system. The initial support for this boundary recognises that the Carboniferous must be defined by scientifically meaningful as well as practical criteria.
Numerous studies have been conducted on the bio-, litho-, and isotopic stratigraphy and event stratigraphy of the continuous DCB successions in South China, making it an important place for precise stratigraphic correlation with the standard Hangenberg beds in Rhenish Massif and further improvement of the geologic timescale. The base of the Protognathodus kockeli Zone/Upper Si. praesulcata Zone usually appears in the first limestone level of post-glacial transgression in the basinal, slope, and platform margin facies of the Youjiang Basin, immediately atop the Changshun Black Shale and its equivalents (the HBS and HSS events). This interval hosts the well-known Hangenberg positive carbon isotope excursion that could be used for high-resolution stratigraphic correlation. However, this position was incorrectly thought to represent the end of mass extinction and beginning of radiation, since environment conditions were still poor during the upper crisis interval, extensive marine anoxia and abrupt climate changes led to blooms of opportunistic fauna and several minor extinction events. The emergence of many new groups did not occur until the upper part of the Si. sulcata Zone to the Si. duplicata Zone.
Following the Hangenberg Crisis interval, conodont biofacies shift to the siphonodellids biofacies in both deep- and shallow-water facies in South China, with greater numbers of early siphonodellids including Si. praesulcata, Si. sulcata, and Si. bransoni and many transitional forms and morphotypes. The Si. sulcata Zone could be recognised, but precise placement of its base is difficult and subjective due to the taxonomic complexity and stratigraphic distribution uncertainties. Taxonomic re-evaluation of the early siphonodellids is needed, especially for the morphotypes of Si. sulcata, since they occur conveniently at the recovery transition, consistent with a negative δ13Ccarb shift following the HICE, and have been widely agreed concept of the Carboniferous.
References
Aretz, M. (2010). Report of the Joint Devonian–Carboniferous boundary GSSP reappraisal task group. Newsletter on Carboniferous Stratigraphy, 28, 26–30.
Bai, S. L., & Wang, C. Y. (1987). New proposal for an international Devonian-Carboniferous boundary. In C. Y. Wang (Ed.), Carboniferous boundaries in China (p. 180). Beijing: Science Press.
Bai, S. L., Bai, Z. Q., Ma, X. P., Wang, D. R., & Sun, Y. L. (1994). Devonian events and biostratigraphy of South China. Beijing: Peking University Press.
Becker, R. T., Gradstein, F. M., & Hammer, O. (2012). The Devonian period. In F. M. Gradstein, J. G. Ogg, M. D. Schmitz, & G. M. Ogg (Eds.), The Geologic Time Scale 2012 (Vol. 2, pp. 559–601). Amsterdam: Elsevier.
Brand, U., Legrand-Blain, M., & Streel, M. (2004). Biochemo-stratigraphy of the Devonian− Carboniferous boundary global stratotype section and point, Griotte Formation, La Serre, Montagne Noire, France. Palaeogeography, Palaeoclimatology, Palaeoecology, 205(3), 337–357.
Brezinski, D. K., Cecil, C. B., Skema, V. W., & Stamm, R. (2008). Late Devonian glacial deposits from the eastern United States signal an end of the mid-Paleozoic warm period. Palaeogeography, Palaeoclimatology, Palaeoecology, 268(3-4), 143–151.
Buggisch, W., & Joachimski, M. M. (2006). Carbon isotope stratigraphy of the Devonian of Central and Southern Europe. Palaeogeography, Palaeoclimatology, Palaeoecology, 240(1), 68–88.
Caputo, M. V. (1985). Late Devonian glaciation in South America. Palaeogeography, Palaeoclimatology, Palaeoecology, 51(1-4), 291–317.
Carmichael, S. K., Waters, J. A., Batchelor, C. J., Coleman, D. M., Suttner, T. J., Kido, E., Moore, L. M., & Chadimová, L. (2016). Climate instability and tipping points in the Late Devonian: Detection of the Hangenberg Event in an open oceanic island arc in the Central Asian Orogenic Belt. Gondwana Research, 32, 213–231.
Chen, B., Chen, J.T., Qie, W.K., Huang, P., He, T.C., Joachimski, M.M., Regelous, M., Pogge von Strandmannd, P.A.E., Liu, J.S., Wang, X.D., Montañez, I.P., & Algeo, T.J. (2021). Was climatic cooling during the earliest Carboniferous driven by expansion of seed plants? Earth and Planetary Science Letters, 565, 116953.
Cramer, B., Saltzman, M., Day, J. E., & Witzke, B. J. (2008). Record of the Late Devonian Hangenberg global positive carbon-isotope excursion in an epeiric sea setting: carbonate production, organic-carbon burial and paleoceanography during the Late Famennian. Geological Association of Canada Special Paper, 48,103–118.
Dahl, T. W., Boyle, R. A., Canfield, D. E., Connelly, J. N., Gill, B. C., Lenton, T. M., & Bizzarro, M. (2014). Uranium isotopes distinguish two geochemically distinct stages during the later Cambrian SPICE event. Earth and Planetary Science Letters, 401, 313–326. https://doi.org/10.1016/j.epsl.2014.05.043.
Davydov, V. I., Korn, D., & Schmitz, M. D. (2012). The Devonian period. In F. M. Gradstein, J. G. Ogg, M. D. Schmitz, & G. M. Ogg (Eds.), The Geologic Time Scale 2012, Volume 2 (pp. 603–651). Amsterdam: Elsevier.
Falkowski, P. G. (1997). Evolution of the nitrogen cycle and its influence on the biological sequestration of CO2 in the ocean. Nature, 387(6630), 272–275.
Feng, R. L. (1977). Discovery of Syringothyris from Southern Guizhou and its significance. Acta Palaeontologica Sinica, 16(1), 53–58. [in Chinese]
Gao, L. D. (1988). Late Devonian and Early Carboniferous miospore zones from Nyalam County, Xizang (Tibet) and boundary of the Devonian–Carboniferous. In Chinese Academy of Geological Sciences (Ed.), Tectonic Evolution of Lithosphere in Himalayan Region (Proceeding of Palaeontology in Tibet) (pp. 181–233). Beijing: Geological Publishing House. [in Chinese]
Gong, X. L., Huang, H. D., Zhang, M. L., & Huang, Q. D. (1991). The stratigraphic classification and correlation of carbonate rocks of Upper Devonian and Lower Carboniferous in Guilin karst region. Nanning: Guangxi Science and Technology Publishing House. [in Chinese]
Gradstein, F. M., Ogg, J. G., Schmitz, M., & Ogg, G. M. (2012). The Geologic Time Scale 2012 (Vol. 2). Amsterdam: Elsevier.
Gruber, N., & Galloway, J. N. (2008). An Earth-system perspective of the global nitrogen cycle. Nature, 451(7176), 293–296.
Hance, L., Hou, H., & Vachard, D. (2011). Upper Famennian to Visean Foraminifers and some carbonate Microproblematica from South China. Beijing: Geological Publishing House.
Hardisty, D. S., Lu, Z., Bekker, A., Diamond, C. W., Gill, B. C., Jiang, G., Kah, L. C., Knoll, A. H., Loyd, S. J., Osburn, M. R., Planavsky, N. J., Wang, C., Zhou, X., & Lyons, T. W. (2017). Perspectives on Proterozoic surface ocean redox from iodine contents in ancient and recent carbonate. Earth and Planetary Science Letters, 463, 159–170.
Hou, H. F., & Ma, X. P. (2005). Devonian GSSPs and division of the Devonian System in South China. Journal of Stratigraphy, 29, 154–159.
Hou, H. F., Ji, Q., Wu, X. H., Xiong, J. F., Wang, S. T., Gao, L. D., et al. (1985). Muhua Sections of Devonian-Carboniferous Boundary Beds. Beijing: Geological Publishing House. [in Chinese]
Hou, H. F., Wang, S. T., et al. (1988). The Devonian System of China (Vol. 7, Stratigraphy of China). Beijing: Geological Publishing House.
Hu, K., Qi, Y., Qie, W., & Wang, Q. (2020). Carboniferous conodont zonation of China. Newsletters on Stratigraphy, 53(2), 141–190.
Isaacson, P., Diaz-Martinez, E., Grader, G., Kalvoda, J., Babek, O., & Devuyst, F. (2008). Late Devonian-earliest Mississippian glaciation in Gondwanaland and its biogeographic consequences. Palaeo-geography, Palaeoclimatology, Palaeoecology, 268(3-4), 126–142.
Ji, Q. (1987). The boundary between the Devonian and Carboniferous systems of shallow-water facies as viewed in the light of conodont studies. Acta Geologica Sinica, (1), 10–20. [in Chinese]
Ji, Q. (2004). On the change of conodonts near the Devonian-Carboniferous boundary. In E. B. o. P. P. o. S. a. Palaeontology (Ed.), Professional Papers of Stratigraphy and Palaeontology (Vol. 28, pp. 111–123). Beijing: Geological Publishing House. [in Chinese]
Ji, Q., & Ziegler, W. (1992). Phylogeny, speciation and zonation of Siphonodella of shallow water facies (Conodonta, Early Carboniferous). Courier Forschungsinstitut Senckenberg, 154, 223–251.
Ji, Q., & Ziegler, W. (1993). The Lali section: An excellent reference section for Upper Devonian in South China. Courier Forschungsinstitut Senckenberg, 157, 1–183.
Ji, Q., Wei, J., Wang, Z., Wang, S., Sheng, H., Wang, H., et al. (1989). The Dapoushang Section: An excellent Section for the Devonian-Carboniferous Boundary Stratotype in China. Beijing: Science Press.
Kaiser, S. I., & Corradini, C. (2011). The early siphonodellids (Conodonta, Late Devonian-Early Carboniferous): overview and taxonomic state. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, 261(1), 19–35.
Kaiser, S. I., Steuber, T., Becker, R. T., & Joachimski, M. M. (2006). Geochemical evidence for major environmental change at the Devonian-Carboniferous boundary in the Carnic Alps and the Rhenish Massif. Palaeogeography, Palaeoclimatology, Palaeo-ecology, 240(1-2), 146–160.
Kaiser, S. I., Steuber, T., & Becker, R. T. (2008). Environmental change during the Late Famennian and Early Tournaisian (Late Devonian-Early Carboniferous): Implications from stable isotopes and conodont biofacies in southern Europe. Geological Journal, 43(23), 241–260.
Kaiser, S. I., Becker, R., Spalletta, C., & Steuber, T. (2009). High-resolution conodont stratigraphy, biofacies, and extinctions around the Hangenberg Event in pelagic successions from Austria, Italy, and France. In Studies in Devonian Stratigraphy: Proceedings of the 2007 International Meeting of the Subcommission on Devonian Stratigraphy and IGCP, 2009. Paleontological Americana, 63, 99–143.
Kaiser, S. I., Aretz, M., & Becker, R. T. (2016). The global Hangenberg Crisis (Devonian–Carboniferous transition): Review of a first-order mass extinction (Vol. 423, pp. 387–437). London, Special Publications: Geological Society.
Kalvoda, J., Kumpan, T., Qie, W., Frýda, J., & Bábek, O. (2019). Mercury spikes at the Devonian-Carboniferous boundary in the eastern part of the Rhenohercynian Zone (central Europe) and in the South China Block. Palaeogeography, Palaeoclimatology, Palaeoecology, 531, 109221.
Kumpan, T., Bábek, O., Kalvoda, J., Matys Grygar, T., & Frýda, J. (2014). Sea-level and environmental changes around the Devonian–Carboniferous boundary in the Namur–Dinant Basin (S Belgium, NE France): A multi-proxy stratigraphic analysis of carbonate ramp archives and its use in regional and interregional correlations. Sedimentary Geology, 311(0), 43-59.
Kumpan, T., Kalvoda, J., Bábek, O., Matys Grygar, T., & Frýda, J. (2021). The Devonian-Carboniferous boundary in the Moravian Karst (Czech Republic). In M. Aretz & C. Corradini (Eds.), Global review of the Devonian-Carboniferous boundary. Palaeobiodiversity and Palaeoenvironments 101(2). https://doi.org/10.1007/s12549-019-00409-z. [this issue]
Liang, K., Qie, W., Pan, L., & Yin, B. (2019). Morphometrics and palaeo-ecology of syringoporoid tabulate corals from the upper Famennian (Devonian) Etoucun Formation, Huilong, South China. In W. Qie, K. Liang, & P. Königshof (Eds.) Devonian palaeoecosystems and palaeoenvironments of South China. Palaeobiodiversity and Palaeoenvironments, 99(1), 101-115.
Liu, Y. Q., Ji, Q., Kuang, H. W., Jiang, X. J., Xu, H., & Peng, N. (2012). U-Pb Zircon age, sedimentary facies, and sequence stratigraphy of the Devonian–Carboniferous boundary, Daposhang Section, Guizhou, China. Palaeoworld, 21, 100–107.
Liu, J., Qie, W., Algeo, T. J., Yao, L., Huang, J., & Luo, G. (2016). Changes in marine nitrogen fixation and denitrification rates during the end-Devonian mass extinction. Palaeogeography, Palaeoclimatology, Palaeoecology, 448, 195–206.
Liu, F., Kerp, H., Peng, H., Zhu, H., & Peng, J. (2019a). Palynostratigraphy of the Devonian–Carboniferous transition in the Tulong section in South Tibet: A Hangenberg event sequence analogue in the Himalaya-Tethys zone. Palaeogeography, Palaeoclimatology, Palaeoecology, 531, 108704.
Liu, J., Luo, G., Lu, Z., Lu, W., Qie, W., Zhang, F., Wang, X., & Xie, S. (2019b). Intensified Ocean Deoxygenation During the end Devonian Mass Extinction. Geochemistry, Geophysics, Geosystems, 20, 6187–6198.
Lu, W., Ridgwell, A., Thomas, E., Hardisty, D. S., Luo, G., Algeo, T. J., Saltzman, M. R., Gill, B. C., Shen, Y., Ling, H. F., Edwards, C. T., Whalen, M. T., Zhou, X., Gutchess, K. M., Jin, L., Rickaby, R. E. M., Jenkyns, H. C., Lyons, T. W., Lenton, T. M., Kump, L. R., & Lu, Z. (2018). Late inception of a resiliently oxygenated upper ocean. Science, 361(6398), 174–177.
Lu, Z. L., Jenkyns, H. C., & Rickaby, R. E. M. (2010). Iodine to calcium ratios in marine carbonate as a paleo-redox proxy during oceanic anoxic events. Geology, 38(12), 1107-1110.
Ma, X. P., Liao, W. H., & Wang, D. M. (2009). The Devonian system of China, with a discussion on sea-level change in South China. In P. Königshof (Ed.), Devonian change: Case studies in palaeo-geography and palaeoecology (pp. 241–262). London: Geological Society of London Special Publication, 314.
Nie, T. (2019). Famennian (Upper Devonian)-Tournaisian (Lower Carboniferous) conodont biostratigraphy and brachiopod fauna of South China. Ph. D. thesis (pp. 1-124). Beijing: Peking University.
Ogg, J. G., Ogg, G. M., & Gradstein, F. M. (2016). A concise Geologic Time Scale. Amsterdam: Elsevier.
Paproth, E., Feist, R., & Flajs, G. (1991). Decision on the Devonian-Carboniferous boundary stratotype. Episodes, 14(4), 331–335.
Percival, L. M. E., Davies, J., Schaltegger, U., De Vleeschouwer, D., Da Silva, A. C., & Follmi, K. B. (2018). Precisely dating the Frasnian-Famennian boundary: implications for the cause of the Late Devonian mass extinction. Scientific Reports, 8(1), 9578.
Qie, W., Zhang, X., Du, Y., Yang, B., Ji, W., & Luo, G. (2014). Conodont biostratigraphy of Tournaisian shallow-water carbonates in central Guangxi, South China. Geobios, 47(6), 389–401.
Qie, W., Liu, J., Chen, J., Wang, X., Mii, H.-S., Zhang, X., et al. (2015). Local overprints on the global carbonate δ13C signal in Devonian–Carboniferous boundary successions of South China. Palaeo-geography, Palaeoclimatology, Palaeoecology, 418, 290–303.
Qie, W., Wang, X. D., Zhang, X., Ji, W., Grossman, E. L., Huang, X., Liu, J., & Luo, G. (2016). Latest Devonian to earliest Carboniferous conodont and carbon isotope stratigraphy of a shallow-water sequence in South China. Geological Journal, 51(6), 915–935.
Qie, W., Ma, X., Xu, H., Qiao, L., Liang, K., Guo, W., Song, J., Chen, B., & Lu, J. (2019a). Devonian integrative stratigraphy and timescale of China. Science China Earth Sciences, 62(1), 112–134.
Qie, W., Algeo, T. J., Luo, G., & Herrmann, A. (2019b). Global events of the Late Paleozoic (Early Devonian to Middle Permian): A review. Palaeogeography, Palaeoclimatology, Palaeoecology, 531, 109259.
Qin, G. R., Zhao, R. X., & Ji, Q. (1988). Late Devonian and early Carboniferous conodonts from northern Guangdong and their stratigraphic significance. Acta Micropalaeontologica Sinica, 5(1), 57–71. [in Chinese]
Qiu, H. (1984). Paleozoic and Triassic conodont faunas in Xizang (Tibet). Sino-French Cooperative Investigation in Himalayas, 1984, 85–107.
Qiu, H. R. (1988). The Late Devonian–Early Carboniferous conodonts of the Yali section from Nyalam, Qomolangma, South Xizang (Tibet). In Chinese Academy of Geological Sciences (Ed.), Tectonic Evolution of Lithosphere in Himalayan Region (Proceeding of Palaeontology in Tibet) (pp. 272–302). Beijing: Geological Publishing House. [in Chinese]
Rao, J. G., & Yu, H. J. (1985). The Devonian system in south Xizang (Tibet). In CGQXP editorial Committee, Ministry of Geology and Mineral Resources PRC (Ed.), Contribution to the Geology of Qinghai-Xizang (Tibet) Plateau (Vol. 16, pp. 51–74). Beijing: Geological Publishing House. [in Chinese]
Ruan, Y. P. (1979). Devonian ammonoid zonation of China. Acta Stratigraphica Sinica, 3, 134–137.
Sandberg, C. A., Ziegler, W., Leuteritz, K., & Brill, S. M. (1978). Phylogeny, speciation, and zonation of Siphonodella (Conodonta, upper Devonian and lower Carboniferous). Newsletters on Stratigraphy, 7(2), 102–120.
Schönlaub, H. P., Attrep, M., Boeckelmann, K., Dreesen, R., Feist, R., Fenninger, A., et al. (1992). The Devonian/Carboniferous boundary in the Carnic Alps (Austria)-a multidisciplinary approach. Jahrbuch der Geologischen Bundesanstalt, 135(1), 57–98.
Sepkoski, J. J. J. (1996). Patterns of Phanerozoic extinction: a perspective from global data bases. In O. H. Walliser (Ed.), Global events and event stratigraphy in the Phanerozoic (pp. 35–51). Berlin: Springer-Verlag.
Song, J. J., & Gong, Y. M. (2018). Wangshangkia, a new Devonian ostracod genus from Dushan of Guizhou. Journal of Micro-palaeontology, 37, 341–346.
Song, J. J., & Gong, Y. M. (2019). Ostracods from the Devonian-Carboniferous transition in Dushan of Guizhou, South China. In W. Qie, K. Liang, & P. Königshof (Eds.) Devonian palaeo-ecosystems and palaeoenvironments of South China. Palaeobiodiversity and Palaeoenvironments, 99(1), 117-127.
Suttner, T. J., Kido, E., Chen, X., Mawson, R., Waters, J. A., Frýda, J., Mathieson, D., Molloy, P. D., Pickett, J., Webster, G. D., & Frýdová, B. (2014). Stratigraphy and facies development of the marine Late Devonian near the Boulongour Reservoir, northwest Xinjiang, China. Journal of Asian Earth Sciences, 80, 101–118.
Tan, Z. X. (1987). Stratigraphy. In Regional Geological Surveying Party, Bureau of Geology and Mineral Resources of Hunan Province (Ed.), The late Devonian and early Carboniferous strata and palaeobiocoenosis of Hunan (pp. 2–65). Beijing: Geological Publishing House. [in Chinese]
Tien, C. C. (1938). The Devonian of China. Geological Review, (13), 355-404. [in Chinese]
Ting, V. K. (1931). On the stratigraphy of the Fengninian System. Acta Geologica Sinica, 10(1), 31–48.
Ting, V. K., & Grabau, A. W. (1936). The Carboniferous of China and its bearing on the classification of the Mississippian and Pennsyl-vanian (pp. 555–571). Paper presented at the XVI International Geological Congress.
Walliser, O. H. (1996). Global events in the Devonian and Carboniferous. In O. H.Walliser(Ed.), Global events and event stratigraphy in the Phanerozoic (pp. 225–250). Berlin: Springer-Verlag.
Wang, C. Y. (1987a). The age of the Cystophrentis Zone. Journal of Stratigraphy, 11(2), 120–125. [in Chinese]
Wang, K. L. (1987b). On the Devonian-Carboniferous boundary based on foraminiferal fauna from South China. Acta Micro-palaeontologica Sinica, 4(2), 161–173. [in Chinese]
Wang, X. D., & Jin, Y. G. (2003). Carboniferous biostratigraphy of China. In W. T. Zhang, P. J. Chen, & A. R. Palmer (Eds.), Biostratigraphy in China (pp. 281–330). Beijing: Science Press.
Wang, C. Y., & Wang, Z. H. (1978). Upper Devonian and Lower Carboniferous conodonts from southern Guizhou. Memoirs of Nanjing Institute of Geology and Palaeontology, Academia Sinica, 11, 51–96. [in Chinese]
Wang, C. Y., & Yin, B. A. (1985). An important Devonian-Carboniferous boundary stratotype in neritic facies of Yishan County, Guangxi. Acta Micropalaeontologica Sinica, 2(1), 28–48. [in Chinese]
Wang, Z. J., et al. (1990). The Carboniferous system of China (Vol. 8, Stratigraphy of China). Beijing: Geological Publishing House. [in Chinese]
Wang, X., Hu, K., Qie, W., Sheng, Q., Chen, B., Lin, W., Yao, L., Wang, Q., Qi, Y., Chen, J., Liao, Z., & Song, J. (2019). Carboniferous integrative stratigraphy and time scale of China. Science China Earth Sciences, 62(1), 135–153.
White, D. A., Elrick, M., Romaniello, S., & Zhang, F. (2018). Global seawater redox trends during the Late Devonian mass extinction detected using U isotopes of marine limestones. Earth and Planetary Science Letters, 503, 68–77.
Wu, W. S., Zhao, J. M., & Jiang, S. G. (1981). Corals from the Shaodong Formation (Etroeungt) of South China. Acta Palaeontologica Sinica, 20(1), 1–14. [in Chinese]
Xian, H., Zhang, S., Li, H., Xiao, Q., Chang, L., Yang, T., & Wu, H. (2019). How did South China connect to and separate from Gondwana? New paleomagnetic constraints from the Middle Devonian red beds in South China. Geophysical Research Letters, 46(13), 7371–7378.
Xu, D. Y., Zheng, Y., Zhang, Q. W., Shen, Z. D., Sun, Y. Y., & Ye, L. F. (1986). Significance of a δ13C anomaly near the Devonian/Carboniferous boundary at the Muhua section, South China. Nature, 321, 854–855.
Xu, H. K., Cai, C. Y., Liao, W. H., & Lu, L. C. (1990). Hongguleleng Formation in Western Junggar and the boundary between Devonian and Carboniferous. Journal of Stratigraphy, 14(4), 292–301. [in Chinese with English abstract]
Yang, S. P., Hou, H. F., Gao, L. D., Wang, Z. J., & Wu, X. H. (1980). The Carboniferous system of China. Acta Geologica Sinica, (3), 167–175. [in Chinese]
Yao, L., Qie, W., Luo, G., Liu, J., Algeo, T. J., Bai, X., Yang B., Wang X. (2015). The TICE event: Perturbation of carbon–nitrogen cycles during the mid-Tournaisian (Early Carboniferous) greenhouse–icehouse transition. Chemical Geology, 401(0), 1-14.
Yu, C. C. (1931). The correlation of the Fengninian system, the Chinese Lower Carboniferous, as based on coral zones. Bulletin of the Geological Society of China, 10(1), 1-30.
Yu, C. M. (1988). Devonian-Carboniferous boundary in Nanbiancun, Guilin, China: Aspects and Records. Beijing: Science Press.
Zhang, M., Becker, R. T., Ma, X., Zhang, Y., & Zong, P. (2019a). Hangenberg black shale with cymaclymeniid ammonoids in the terminal Devonian of South China. In W. Qie, K. Liang, & P. Königshof (Eds.) Devonian palaeoecosystems and palaeoenvironments of South China. Palaeobiodiversity and Palaeoenvironments, 99(1), 129–142.
Zhang, X., Joachimski, M. M., Over, D. J., Ma, K., Huang, C., & Gong, Y. (2019b). Late Devonian carbon isotope chemostratigraphy: A new record from the offshore facies of South China. Global and Planetary Change, 182, 103024.
Zhang, F., Xiao, S., Romaniello, S. J., Hardisty, D., Li, C., Melezhik, V., Pokrovsky, B., Cheng, M., Shi, W., Lenton, T. M., & Anbar, A. D. (2019c). Global marine redox changes drove the rise and fall of the Ediacara biota. Geobiology, 17(6), 594–610.
Zhang, F., Dahl, T. W., Lenton, T. M., Luo, G., & Shen, S.-z., Algeo, T. J., et al. (2020). Extensive marine anoxia associated with the Late Devonian Hangenberg crisis. Earth and Planetary Science Letters, 533, 115976.
Ziegler, W., & Sandberg, C. A. (1984). Important candidate sections for stratotype of conodont based Devonian-Carboniferous boundary. Courier Forschungsinstitut Senckenberg, 67, 231–239.
Ziegler, W., & Sandberg, C. A. (1996). Reflexions on Frasnian and Famennian stage boundary decisions as a guide to future deliberations. Newsletters on Stratigraphy, 33(3), 157–180.
Zong, P., & Ma, X. P. (2012). Spiriferide and spiriferinide brachiopods across the Devonian and Carboniferous boundary in western Junggar, Xinjiang. Acta Palaeontologica Sinica, 51(2), 157–175. [in Chinese with English abstract]
Zong, P., Becker, R. T., & Ma, X. (2015). Upper Devonian (Famennian) and Lower Carboniferous (Tournaisian) ammonoids from western Junggar, Xinjiang, northwestern China—stratigraphy, taxonomy and palaeobiogeography. Palaeobiodiversity and Palaeoenvironments, 95(2), 159–202.
Zong, P., Ma, X., Xue, J., & Jin, X. (2016). Comparative study of Late Devonian (Famennian) brachiopod assemblages, sea level changes, and geo-events in northwestern and southern China. Palaeogeography, Palaeoclimatology, Palaeoecology, 448, 298–316.
Zong, R., Wang, Z., Fan, R., Song, J., Zhang, X., Shen, Z., et al. (2020). New knowledge on the Hongguleleng Formation and Devonian-Carboniferous boundary in western Junggar, Xinjiang. Acta Geologicca Sinica, 94(8), 2460–2475. [in Chinese with English abstract]
Acknowledgements
We thank M. Aretz and C. Corradini for organising this special issue and their valuable comments.
Funding
This study was financially supported by Strategic Priority Research Program (B) of Chinese Academy of Sciences (XDB26000000) and NSFC grant (91955201, 41772004).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Informed consent
The authors all agree with the content of the manuscript and are aware of the submission.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
This article is a contribution to the special issue “Global review of the Devonian-Carboniferous Boundary”
Rights and permissions
About this article

Cite this article
Qie, W., Sun, Y., Guo, W. et al. Devonian-Carboniferous boundary in China. Palaeobio Palaeoenv 101, 589–611 (2021). https://doi.org/10.1007/s12549-021-00494-z
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12549-021-00494-z