
Abstract
Autophagy is a mechanism responsible for the degradation of cellular components to maintain their homeostasis. However, autophagy is commonly altered and compromised in several diseases, including neurodegenerative disorders. Parkinson’s disease (PD) can be considered a multifactorial disease because environmental factors, genetic factors, and aging are involved. Several genes are involved in PD pathology, among which the LRRK2 gene and its mutations, inherited in an autosomal dominant manner, are responsible for most genetic PD cases. The R1441G LRRK2 mutation is, after G2019S, the most important in PD pathogenesis. Our results demonstrate a relationship between the R1441G LRRK2 mutation and a mechanistic dysregulation of autophagy that compromises cell viability. This altered autophagy mechanism is associated with organellar stress including mitochondrial (which induces mitophagy) and endoplasmic reticulum (ER) stress, consistent with the fact that patients with this mutation are more vulnerable to toxins related to PD, such as MPP+.
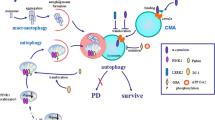
Graphical abstract







Similar content being viewed by others


Availability of data and material
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Code availability
Not applicable.
Abbreviations
- Baf. A1:
-
Bafilomycin A1 (Baf. A1)
- CCCP:
-
Carbonyl cyanide 3-chlorophenylhydrazone
- EBSS:
-
Earle’s balanced salt solution
- ER:
-
Endoplasmic reticulum
- LRRK2:
-
Leucine-rich repeat kinase 2
- LTR:
-
Lysotracker red
- MAMs:
-
Mitochondria-associated ER membranes
- MPP+ :
-
1-Methyl-4-phenylpiridinium
- PD:
-
Parkinson’s disease
- PINK1:
-
PTEN-induced putative kinase 1
- ROS:
-
Reactive oxygen species
- TOMM20:
-
Translocase of outer mitochondrial membrane 20
References
Bezprozvanny I. Calcium signaling and neurodegenerative diseases. Trends Mol Med. 2009;15:89–100.
Bonello F, Hassoun SM, Mouton-Liger F, Shin YS, Muscat A, Tesson C, Lesage S, Beart PM, Brice A, Krupp J, et al. LRRK2 impairs PINK1/Parkin-dependent mitophagy via its kinase activity: pathologic insights into Parkinson’s disease. Hum Mol Genet. 2019;28:1645–60.
Bravo-San Pedro JM, Niso-Santano M, Gomez-Sanchez R, Pizarro-Estrella E, Aiastui-Pujana A, Gorostidi A, Climent V, Lopez de Maturana R, Sanchez-Pernaute R, Lopez de Munain A, et al. The LRRK2 G2019S mutant exacerbates basal autophagy through activation of the MEK/ERK pathway. Cell Mol Life Sci. 2013;70:121–36.
Burte F, Carelli V, Chinnery PF, Yu-Wai-Man P. Disturbed mitochondrial dynamics and neurodegenerative disorders. Nat Rev Neurol. 2015;11:11–24.
Cali T, Ottolini D, Negro A, Brini M. Enhanced parkin levels favor ER-mitochondria crosstalk and guarantee Ca(2+) transfer to sustain cell bioenergetics. Biochim Biophys Acta. 2013;1832:495–508.
Canet-Aviles RM, Wilson MA, Miller DW, Ahmad R, McLendon C, Bandyopadhyay S, Baptista MJ, Ringe D, Petsko GA, Cookson MR. The Parkinson’s disease protein DJ-1 is neuroprotective due to cysteine-sulfinic acid-driven mitochondrial localization. Proc Natl Acad Sci U S A. 2004;101:9103–8.
Cerri S, Blandini F. Role of autophagy in Parkinson’s disease. Curr Med Chem. 2019;26:3702–18.
Cherra SJ 3rd, Steer E, Gusdon AM, Kiselyov K, Chu CT. Mutant LRRK2 elicits calcium imbalance and depletion of dendritic mitochondria in neurons. Am J Pathol. 2013;182:474–84.
Cho HJ, Yu J, Xie C, Rudrabhatla P, Chen X, Wu J, Parisiadou L, Liu G, Sun L, Ma B, et al. Leucine-rich repeat kinase 2 regulates Sec16A at ER exit sites to allow ER-Golgi export. EMBO J. 2014;33:2314–31.
Chung SY, Kishinevsky S, Mazzulli JR, Graziotto J, Mrejeru A, Mosharov EV, Puspita L, Valiulahi P, Sulzer D, Milner TA, et al. Parkin and PINK1 patient iPSC-derived midbrain dopamine neurons exhibit mitochondrial dysfunction and alpha-synuclein accumulation. Stem Cell Reports. 2016;7:664–77.
Cuervo AM, Stefanis L, Fredenburg R, Lansbury PT, Sulzer D. Impaired degradation of mutant alpha-synuclein by chaperone-mediated autophagy. Science. 2004;305:1292–5.
de Brito OM, Scorrano L. Mitofusin 2 tethers endoplasmic reticulum to mitochondria. Nature. 2008;456:605–10.
Fonseca TB, Sanchez-Guerrero A, Milosevic I, Raimundo N. Mitochondrial fission requires DRP1 but not dynamins. Nature. 2019;570:E34–42.
Galluzzi L, Bravo-San Pedro JM, Levine B, Green DR, Kroemer G. Pharmacological modulation of autophagy: therapeutic potential and persisting obstacles. Nat Rev Drug Discov. 2017;16:487–511.
Ganguly U, Chakrabarti SS, Kaur U, Mukherjee A, Chakrabarti S. Alpha-synuclein, proteotoxicity and Parkinson’s disease: search for neuroprotective therapy. Curr Neuropharmacol. 2018;16:1086–97.
Giacomello M, Pyakurel A, Glytsou C, Scorrano L. The cell biology of mitochondrial membrane dynamics. Nat Rev Mol Cell Biol. 2020;21:204–24.
Gomez-Suaga P, Luzon-Toro B, Churamani D, Zhang L, Bloor-Young D, Patel S, Woodman PG, Churchill GC, Hilfiker S. Leucine-rich repeat kinase 2 regulates autophagy through a calcium-dependent pathway involving NAADP. Hum Mol Genet. 2012;21:511–25.
Gomez-Suaga P, Rivero-Rios P, Fdez E, Blanca Ramirez M, Ferrer I, Aiastui A, Lopez De Munain A, Hilfiker S. LRRK2 delays degradative receptor trafficking by impeding late endosomal budding through decreasing Rab7 activity. Hum Mol Genet. 2014;23:6779–96.
Gomez-Suaga P, Bravo-San Pedro JM, Gonzalez-Polo RA, Fuentes JM, Niso-Santano M. ER-mitochondria signaling in Parkinson’s disease. Cell Death Dis. 2018;9:337.
Greggio E, Taymans JM, Zhen EY, Ryder J, Vancraenenbroeck R, Beilina A, Sun P, Deng J, Jaffe H, Baekelandt V, et al. The Parkinson’s disease kinase LRRK2 autophosphorylates its GTPase domain at multiple sites. Biochem Biophys Res Commun. 2009;389:449–54.
Hsieh CH, Shaltouki A, Gonzalez AE, Bettencourt da Cruz A, Burbulla LF, St Lawrence E, Schule B, Krainc D, Palmer TD, Wang X. Functional impairment in miro degradation and mitophagy is a shared feature in familial and sporadic Parkinson’s disease. Cell Stem Cell. 2016;19:709–24.
Ivankovic D, Chau KY, Schapira AH, Gegg ME. Mitochondrial and lysosomal biogenesis are activated following PINK1/parkin-mediated mitophagy. J Neurochem. 2016;136:388–402.
Kania E, Pajak B, Orzechowski A. Calcium homeostasis and ER stress in control of autophagy in cancer cells. Biomed Res Int. 2015;2015:352794.
Kitzman HH Jr, McMahon RJ, Aslanian AM, Fadia PM, Frost SC. Differential regulation of GRP78 and GLUT1 expression in 3T3-L1 adipocytes. Mol Cell Biochem. 1996;162:51–8.
Klionsky DJ, Abdalla FC, Abeliovich H, Abraham RT, Acevedo-Arozena A, Adeli K, Agholme L, Agnello M, Agostinis P, Aguirre-Ghiso JA, et al. Guidelines for the use and interpretation of assays for monitoring autophagy. Autophagy. 2012;8:445–544.
Klionsky DJ, Abdelmohsen K, Abe A, Abedin MJ, Abeliovich H, Acevedo Arozena A, Adachi H, Adams CM, Adams PD, Adeli K, et al. Guidelines for the use and interpretation of assays for monitoring autophagy (3rd edition). Autophagy. 2016;12:1–222.
Koentjoro B, Park JS, Sue CM. Nix restores mitophagy and mitochondrial function to protect against PINK1/Parkin-related Parkinson’s disease. Sci Rep. 2017;7:44373.
Larsen SB, Hanss Z, Kruger R. The genetic architecture of mitochondrial dysfunction in Parkinson’s disease. Cell Tissue Res. 2018;373:21–37.
Lavalley NJ, Slone SR, Ding H, West AB, Yacoubian TA. 14-3-3 Proteins regulate mutant LRRK2 kinase activity and neurite shortening. Hum Mol Genet. 2016;25:109–22.
Lee KS, Huh S, Lee S, Wu Z, Kim AK, Kang HY, Lu B. Altered ER-mitochondria contact impacts mitochondria calcium homeostasis and contributes to neurodegeneration in vivo in disease models. Proc Natl Acad Sci U S A. 2018;115:E8844–53.
Lee JH, Han JH, Kim H, Park SM, Joe EH, Jou I. Parkinson’s disease-associated LRRK2-G2019S mutant acts through regulation of SERCA activity to control ER stress in astrocytes. Acta Neuropathol Commun. 2019;7:68.
Lievremont JP, Rizzuto R, Hendershot L, Meldolesi J. BiP, a major chaperone protein of the endoplasmic reticulum lumen, plays a direct and important role in the storage of the rapidly exchanging pool of Ca2+. J Biol Chem. 1997;272:30873–9.
Lin D, Liang Y, Zheng D, Chen Y, Jing X, Lei M, Zeng Z, Zhou T, Wu X, Peng S, et al. Novel biomolecular information in rotenone-induced cellular model of Parkinson’s disease. Gene. 2018;647:244–60.
Liu HF, Lu S, Ho PW, Tse HM, Pang SY, Kung MH, Ho JW, Ramsden DB, Zhou ZJ, Ho SL. LRRK2 R1441G mice are more liable to dopamine depletion and locomotor inactivity. Annals of Clinical and Translational Neurology. 2014;1:199–208.
Lopez de Maturana R, Lang V, Zubiarrain A, Sousa A, Vazquez N, Gorostidi A, Aguila J, Lopez de Munain A, Rodriguez M, Sanchez-Pernaute R. Mutations in LRRK2 impair NF-kappaB pathway in iPSC-derived neurons. J Neuroinflammation. 2016;13:295.
Lynch-Day MA, Mao K, Wang K, Zhao M, Klionsky DJ. The role of autophagy in Parkinson’s disease. Cold Spring Harb Perspect Med. 2012;2:a009357.
Manzoni C. The LRRK2-macroautophagy axis and its relevance to Parkinson’s disease. Biochem Soc Trans. 2017;45:155–62.
Manzoni C, Mamais A, Dihanich S, Abeti R, Soutar MPM, Plun-Favreau H, Giunti P, Tooze SA, Bandopadhyay R, Lewis PA. Inhibition of LRRK2 kinase activity stimulates macroautophagy. Biochim Biophys Acta. 2013a;1833:2900–10.
Manzoni C, Mamais A, Dihanich S, McGoldrick P, Devine MJ, Zerle J, Kara E, Taanman JW, Healy DG, Marti-Masso JF, et al. Pathogenic Parkinson’s disease mutations across the functional domains of LRRK2 alter the autophagic/lysosomal response to starvation. Biochem Biophys Res Commun. 2013b;441:862–6.
Martin I, Kim JW, Dawson VL, Dawson TM. LRRK2 pathobiology in Parkinson’s disease. J Neurochem. 2014;131:554–65.
Martinez JH, Alaimo A, Gorojod RM, Porte Alcon S, Fuentes F, Coluccio Leskow F, Kotler ML. Drp-1 dependent mitochondrial fragmentation and protective autophagy in dopaminergic SH-SY5Y cells overexpressing alpha-synuclein. Mol Cell Neurosci. 2018;88:107–17.
Mattson MP. Calcium and neurodegeneration. Aging Cell. 2007;6:337–50.
Mauro-Lizcano M, Esteban-Martinez L, Seco E, Serrano-Puebla A, Garcia-Ledo L, Figueiredo-Pereira C, Vieira HL, Boya P. New method to assess mitophagy flux by flow cytometry. Autophagy. 2015;11:833–43.
McCoy MK, Cookson MR. DJ-1 regulation of mitochondrial function and autophagy through oxidative stress. Autophagy. 2011;7:531–2.
Melser S, Lavie J, Benard G. Mitochondrial degradation and energy metabolism. Biochim Biophys Acta. 2015;1853:2812–21.
Mercado G, Castillo V, Soto P, Lopez N, Axten JM, Sardi SP, Hoozemans JJM, Hetz C. Targeting PERK signaling with the small molecule GSK2606414 prevents neurodegeneration in a model of Parkinson’s disease. Neurobiol Dis. 2018;112:136–48.
Michel PP, Hirsch EC, Hunot S. Understanding dopaminergic cell death pathways in Parkinson disease. Neuron. 2016;90:675–91.
Mishra P, Chan DC. Metabolic regulation of mitochondrial dynamics. J Cell Biol. 2016;212:379–87.
Mortiboys H, Johansen KK, Aasly JO, Bandmann O. Mitochondrial impairment in patients with Parkinson disease with the G2019S mutation in LRRK2. Neurology. 2010;75:2017–20.
Niu J, Yu M, Wang C, Xu Z. Leucine-rich repeat kinase 2 disturbs mitochondrial dynamics via Dynamin-like protein. J Neurochem. 2012;122:650–8.
Orenstein SJ, Kuo SH, Tasset I, Arias E, Koga H, Fernandez-Carasa I, Cortes E, Honig LS, Dauer W, Consiglio A, et al. Interplay of LRRK2 with chaperone-mediated autophagy. Nat Neurosci. 2013;16:394–406.
Papkovskaia TD, Chau KY, Inesta-Vaquera F, Papkovsky DB, Healy DG, Nishio K, Staddon J, Duchen MR, Hardy J, Schapira AH, Cooper JM. G2019S leucine-rich repeat kinase 2 causes uncoupling protein-mediated mitochondrial depolarization. Hum Mol Genet. 2012;21:4201–13.
Pfaffl MW. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001;29:e45.
Plowey ED, Cherra SJ 3rd, Liu YJ, Chu CT. Role of autophagy in G2019S-LRRK2-associated neurite shortening in differentiated SH-SY5Y cells. J Neurochem. 2008;105:1048–56.
Rodriguez-Arribas M, Pizarro-Estrella E, Gomez-Sanchez R, Yakhine-Diop SM, Gragera-Hidalgo A, Cristo A, Bravo-San Pedro JM, Gonzalez-Polo RA, Fuentes JM. IFDOTMETER: a new software application for automated immunofluorescence analysis. J Lab Autom. 2016;21:246–59.
Rodriguez-Arribas M, Yakhine-Diop SM, Gonzalez-Polo RA, Niso-Santano M, Fuentes JM. Turnover of lipidated LC3 and autophagic cargoes in mammalian cells. Methods Enzymol. 2017a;587:55–70.
Rodriguez-Arribas M, Yakhine-Diop SMS, Pedro JMB, Gomez-Suaga P, Gomez-Sanchez R, Martinez-Chacon G, Fuentes JM, Gonzalez-Polo RA, Niso-Santano M. Mitochondria-associated membranes (MAMs): overview and its role in Parkinson’s disease. Mol Neurobiol. 2017b;54:6287–303.
Saez-Atienzar S, Bonet-Ponce L, Blesa JR, Romero FJ, Murphy MP, Jordan J, Galindo MF. The LRRK2 inhibitor GSK2578215A induces protective autophagy in SH-SY5Y cells: involvement of Drp-1-mediated mitochondrial fission and mitochondrial-derived ROS signaling. Cell Death Dis. 2014;5:e1368.
Sahani MH, Itakura E, Mizushima N. Expression of the autophagy substrate SQSTM1/p62 is restored during prolonged starvation depending on transcriptional upregulation and autophagy-derived amino acids. Autophagy. 2014;10:431–41.
Samann J, Hegermann J, von Gromoff E, Eimer S, Baumeister R, Schmidt E. Caenorhabditits elegans LRK-1 and PINK-1 act antagonistically in stress response and neurite outgrowth. J Biol Chem. 2009;284:16482–91.
Schapira AH. Mitochondria in the aetiology and pathogenesis of Parkinson’s disease. Lancet Neurol. 2008;7:97–109.
Schindelin J, Arganda-Carreras I, Frise E, Kaynig V, Longair M, Pietzsch T, Preibisch S, Rueden C, Saalfeld S, Schmid B, et al. Fiji: an open-source platform for biological-image analysis. Nat Methods. 2012;9:676–82.
Schwab AJ, Sison SL, Meade MR, Broniowska KA, Corbett JA, Ebert AD. Decreased sirtuin deacetylase activity in LRRK2 G2019S iPSC-derived dopaminergic neurons. Stem Cell Reports. 2017;9:1839–52.
Somme JH, Molano Salazar A, Gonzalez A, Tijero B, Berganzo K, Lezcano E, Fernandez Martinez M, Zarranz JJ, Gomez-Esteban JC. Cognitive and behavioral symptoms in Parkinson’s disease patients with the G2019S and R1441G mutations of the LRRK2 gene. Parkinsonism Relat Disord. 2015;21:494–9.
Su YC, Qi X. Inhibition of excessive mitochondrial fission reduced aberrant autophagy and neuronal damage caused by LRRK2 G2019S mutation. Hum Mol Genet. 2013;22:4545–61.
Su YC, Guo X, Qi X. Threonine 56 phosphorylation of Bcl-2 is required for LRRK2 G2019S-induced mitochondrial depolarization and autophagy. Biochim Biophys Acta. 2015;1852:12–21.
Tagliaferro P, Kareva T, Oo TF, Yarygina O, Kholodilov N, Burke RE. An early axonopathy in a hLRRK2(R1441G) transgenic model of Parkinson disease. Neurobiol Dis. 2015;82:359–71.
Teves JMY, Bhargava V, Kirwan KR, Corenblum MJ, Justiniano R, Wondrak GT, Anandhan A, Flores AJ, Schipper DA, Khalpey Z, et al. Parkinson’s disease skin fibroblasts display signature alterations in growth, redox homeostasis, mitochondrial function, and autophagy. Front Neurosci. 2017;11:737.
Toyofuku T, Okamoto Y, Ishikawa T, Sasawatari S, Kumanogoh A. LRRK2 regulates endoplasmic reticulum-mitochondrial tethering through the PERK-mediated ubiquitination pathway. EMBO J. 2020;39:e100875.
van der Merwe C, Jalali Sefid Dashti Z, Christoffels A, Loos B, Bardien S. Evidence for a common biological pathway linking three Parkinson’s disease-causing genes: parkin, PINK1 and DJ-1. Eur J Neurosci. 2015;41:1113–25.
Wang X, Yan MH, Fujioka H, Liu J, Wilson-Delfosse A, Chen SG, Perry G, Casadesus G, Zhu X. LRRK2 regulates mitochondrial dynamics and function through direct interaction with DLP1. Hum Mol Genet. 2012;21:1931–44.
Weil R, Laplantine E, Curic S, Genin P. Role of optineurin in the mitochondrial dysfunction: potential implications in neurodegenerative diseases and cancer. Front Immunol. 2018;9:1243.
Williams JA, Hou Y, Ni HM, Ding WX. Role of intracellular calcium in proteasome inhibitor-induced endoplasmic reticulum stress, autophagy, and cell death. Pharm Res. 2013;30:2279–89.
Wong DP, Chu JM, Hung VK, Lee DK, Cheng CH, Yung KK, Yue KK. Modulation of endoplasmic reticulum chaperone GRP78 by high glucose in hippocampus of streptozotocin-induced diabetic mice and C6 astrocytic cells. Neurochem Int. 2013;63:551–60.
Yakhine-Diop SM, Bravo-San Pedro JM, Gomez-Sanchez R, Pizarro-Estrella E, Rodriguez-Arribas M, Climent V, Aiastui A, Lopez de Munain A, Fuentes JM, Gonzalez-Polo RA. G2019S LRRK2 mutant fibroblasts from Parkinson’s disease patients show increased sensitivity to neurotoxin 1-methyl-4-phenylpyridinium dependent of autophagy. Toxicology. 2014;324:1–9.
Yakhine-Diop SMS, Niso-Santano M, Rodriguez-Arribas M, Gomez-Sanchez R, Martinez-Chacon G, Uribe-Carretero E, Navarro-Garcia JA, Ruiz-Hurtado G, Aiastui A, Cooper JM, et al. Impaired mitophagy and protein acetylation levels in fibroblasts from Parkinson’s disease patients. Mol Neurobiol. 2019;56:2466–81.
Yang S, Xia C, Li S, Du L, Zhang L, Hu Y. Mitochondrial dysfunction driven by the LRRK2-mediated pathway is associated with loss of Purkinje cells and motor coordination deficits in diabetic rat model. Cell Death Dis. 2014;5:e1217.
Yuan Y, Cao P, Smith MA, Kramp K, Huang Y, Hisamoto N, Matsumoto K, Hatzoglou M, Jin H, and Feng Z. Dysregulated LRRK2 signaling in response to endoplasmic reticulum stress leads to dopaminergic neuron degeneration in C. elegans. PLoS One. 2011;6:e22354.
Zhu Y, Wang C, Yu M, Cui J, Liu L, Xu Z. ULK1 and JNK are involved in mitophagy incurred by LRRK2 G2019S expression. Protein Cell. 2013;4:711–21.
Zurita Rendon O, and Shoubridge EA. LONP1 is required for maturation of a subset of mitochondrial proteins, and its loss elicits an integrated stress response. Mol Cell Biol. 2018;38.
Acknowledgements
The authors thank M.P. Delgado-Luceño and FUNDESALUD for helpful assistance and are grateful to the patients and donors for their participation.
Funding
This research was supported by the “Instituto de Salud Carlos” III CIBERNED (CB06/05/0041 and PI14/00170) and partially supported by the “Fondo Europeo de Desarrollo Regional” (FEDER) from the European Union. S.M.S.Y-D was supported by CIBERNED. S. C-C and E.U-C are supported by a FPU fellowship (FPU19/04435 and FPU16/00684, respectively) from the Ministerio de Ciencia, Innovación y Universidades, Spain. G. M-C is supported by University of Extremadura (ONCE Foundation). M. B-B is supported by a collaboration grant from the Ministerio de Educación y Formación Profesional, Spain. G. D-G is supported by the Consejería de Educación y Empleo-SEXPE-Fondo Social Europeo (TE-0031-19). M. P-B is a recipient of a fellowship from the “Plan Propio de Iniciación a la Investigación, Desarrollo Tecnológico e Innovación (University of Extremadura)”. E.A-C is supported by a grant (IB18048) from the Junta de Extremadura, Spain. M.N-S and J.M-B. S-P were funded by the “Ramon y Cajal” Program (RYC-2016–20883 and RYC-2018–025099, respectively), Spain.
Author information
Authors and Affiliations
Contributions
RAGP, JMF, and MNS designed the study and wrote the publication with help of JMBSP, MRA, and SMSYD. SMSYD, MRA, and SCC carried out most the lab experiments. GMC, EUC, MBB, GDG, MPB, and EAC provided help with the experiments. VC provided technical assistance with electron microscopy studies. AA and ALM provided the fibroblast model from patient biopsies. All authors have read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval
All patients gave written consent, and all procedures were approved by the “Comité Ético de Investigación Clínica del Área Sanitaria de Gipuzkoa,” Spain.
Consent to participate
Not applicable.
Consent for publication
Not applicable.
Conflict of interest
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Mireia Niso-Santano, José M. Fuentes, and Rosa A. González-Polo are senior co-authors
Supplementary Information
Below is the link to the electronic supplementary material.
10565_2021_9617_MOESM1_ESM.pdf
Supplementary Figure S1 Characterization and pools selection. (a) PCR products of the different cell lines after digestion with the endonuclease Bsh 1236 I. Line 1: negative control (Neg1) of the restriction analysis, using water instead of enzyme, line 2: negative control 2 (Neg2), using water instead of PCR product, lines 3-6: healthy controls without LRRK2 R1441G mutation (C1-4), lines 7-10: PD patients controls without LRRK2 R1441G mutation (Cα1-4 ), lines 11-13: Healthy subjects with LRRK2 R1441G mutation (Rα1-3), lines 14-17: PD patients heterozygous for LRRK2 R1441G (RG1-4). (b, c) Autophagy characterization of cell lines from subjects with or without R1441G LRRK2. The lysates were obtained from fibroblasts of healthy subjects (C), idiopathic PD patients (Cα), healthy subjects with R1441G LRRK2 mutation (Rα) and PD patients with R1441G LRRK2 mutation. (a) Shows the representative blot of LC3 protein. GAPDH is the loading control. (b) represents the densitometry of LC3-II/GAPDH, in arbitrary units (PDF 1213 KB)
10565_2021_9617_MOESM2_ESM.pdf
Supplementary Figure S2 Analysis of lysosomal and autophagic proteins. (a) LAMP2 determination by immunofluorescence. Control and R1441G human fibroblasts were treated or not with Baf. A1 (100 nM) or EBSS for 4 hours. Next, the cells were fixed and labeled against the LAMP2 protein as described in Materials and Methods. Panel a shows representative images from three independent experiments of each cell line for the conditions studied. The scale bar represents the length of 10 μm. (b-g) Characterization of lysosomal proteins by western blot. Cell lysates were obtained from control (Co) and R1441G fibroblasts (RG) treated or not with Baf. A1 (100 nM) or EBSS for 4 hours and equal amounts of proteins were separated in 12% polyacrylamide gels as described in the Materials and Methods section. Panels (b, d, e) show representative western blots targeting LAMP1 and LAMP2 (b), CTSB (d) and CTSC (e) proteins in three independent experiments. β-Actin was used as a loading control. Panels (c, f, g) show the densitometry histograms associated with the experiments represented in b, d and e, respectively. The asterisks show significant differences between cell types, while the pound signs reflect differences related to the treatment (* or #, p < 0.05; ** or ##, p < 0.01). (h, i) Analysis of p-S6 protein. Cell lysates were obtained from control (Co) and R1441G fibroblasts (RG) treated or not with Baf. A1 (100 nM) or EBSS for 4 hours and equal amounts of proteins were separated in 12% polyacrylamide gels as described in the Materials and Methods section. Panel h shows representative western blot and panel i shows the densitometry histogram. β-Actin was used as a loading control. The pound sign reflect differences related to the treatment (###, p < 0.001). (j-m) Characterization of autophagic proteins by western blot. Cell lysates from control (Co) and R1441G fibroblasts (RG) treated or not with Baf. A1 (100 nM) or EBSS for 4 hours were obtained and equal amounts of proteins were loaded in 12% polyacrylamide gels as described in the Materials and Methods section. Panel j shows representative western blots of p-mTOR, ATG5 and Beclin1 proteins of three independent experiments. β-Actin was used as a loading control. The panels show the densitometry histograms associated for p-mTOR (k), ATG5 (l) and BECN1 (m) proteins of three independent experiments. The asterisks show significant differences between cell types, while the pound signs reflect differences related to the treatment (** p < 0.01; #, p < 0.05) (PDF 101 KB)
10565_2021_9617_MOESM3_ESM.pdf
Supplementary Figure S3 Determination of mitochondrial status and mitochondrial proteins as well as mitolysosome formation in the presence of CCCP. (a-e) Densitometric analyses of selected mitochondrial proteins. The control (Co) and R1441G fibroblasts (RG) were treated and collected as in Fig. 2 (c-e). The panels show the densitometries of the representative western blot shown in Fig. 2 (c-e) for the TOMM20 (a), PHB1 (b), VDAC1 (c), TIMM23 (d) and COXIV (e) proteins, respectively. The asterisks show significant differences (* p < 0.05; ** p < 0.01). (f) Electron microscopy (EM) analysis of mitochondrial content. The cell pellet from the control and R1441G fibroblasts was fixed and processed for EM as described in the Materials and Methods section. The arrows and colored structures in the image point to the recognizable mitochondria. (g) Analysis of mitochondrial reactive oxygen species (ROS) through MitoSOX labeling. The control and R1441G fibroblasts were treated or not with rotenone (1 μM) for 4 hours. Next, the cells were labeled with the MitoSOX staining and processed according to the Materials and Methods section. Panel shows representative immunofluorescence images. h, Study of mitochondria and lysosome colocalization by fluorescence microscopy. The control (Co) and R1441G fibroblasts (RG) were treated for four hours with CCCP (10 μM). Next, the cells were stained and processed as in Fig. 2 (h, i). Panel (h) shows representative images of each of the cell groups under the conditions studied. The scale bar represents a length of 10 μm. (i) The graphs represent the average of the Mander coefficient for the proportion of LTR on the TOMM20 label plus the standard deviation after the treatment with CCCP. (j, k). Effect of Baf. A1 in TOMM20 and TIMM23 proteins levels. Cell lysates were obtained from control (Co) and R1441G fibroblasts (RG) treated or not with Baf. A1 (100 nM) for 4 hours and equal amounts of proteins were separated in 12% polyacrylamide gels as described in the Materials and Methods section. Panel j shows the densitometry histogram associated with the representative western blot targeting TIMM23 and TOMM20 proteins in three independent experiments showed in panel k. β-Actin was used as a loading control. The asterisks show significant differences between cell types, while the pound signs reflect differences related to the treatment (* or #, p < 0.05) (PDF 88 KB)
Rights and permissions
About this article

Cite this article
Yakhine-Diop, S.M.S., Rodríguez-Arribas, M., Canales-Cortés, S. et al. The parkinsonian LRRK2 R1441G mutation shows macroautophagy-mitophagy dysregulation concomitant with endoplasmic reticulum stress. Cell Biol Toxicol 38, 889–911 (2022). https://doi.org/10.1007/s10565-021-09617-w
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10565-021-09617-w