
Extremophiles, a Nifty Tool to Face Environmental Pollution: From Exploitation of Metabolism to Genome Engineering

 ,
,  , ,
, , 
Abstract
:1. Introduction
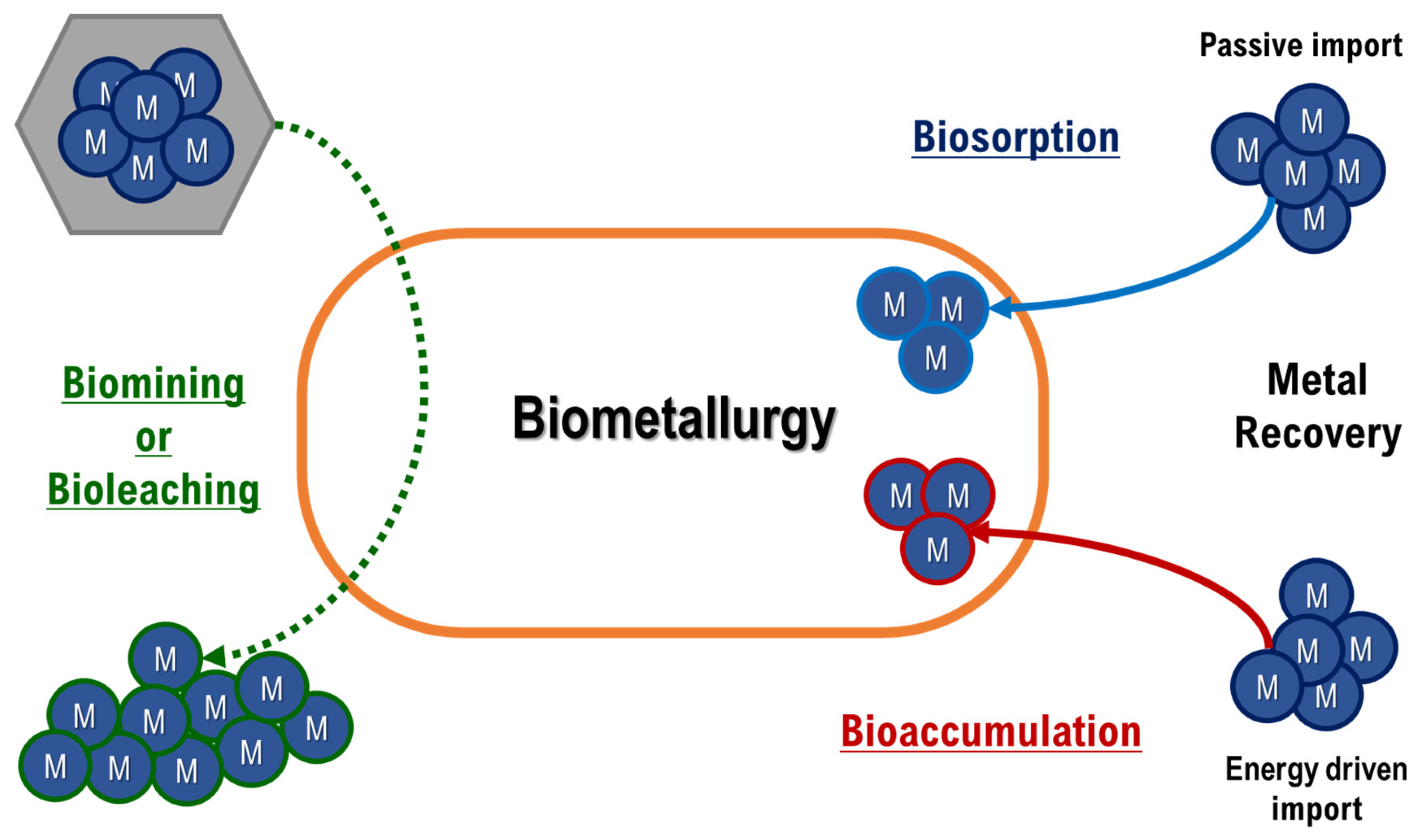
2. Heavy Metals
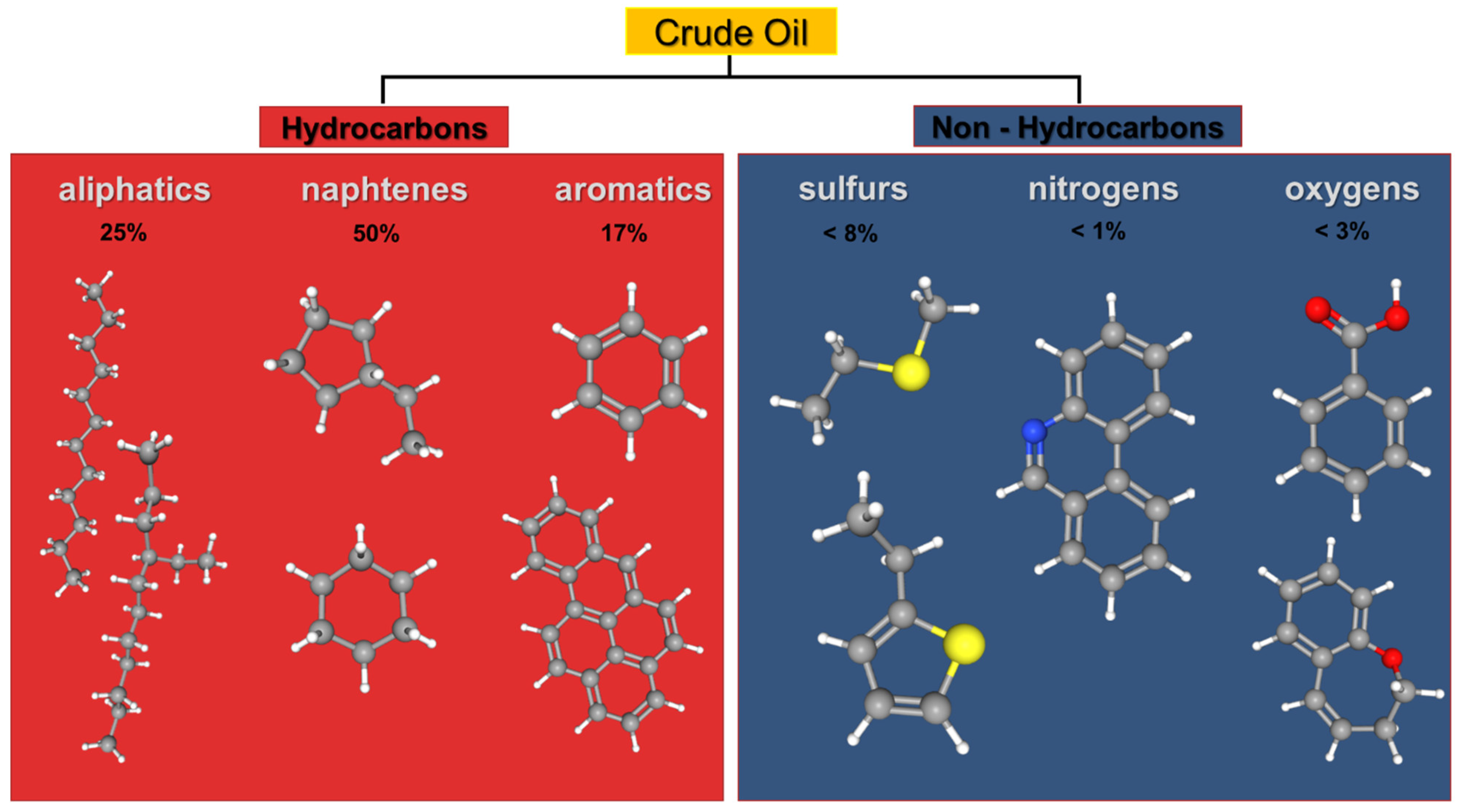
3. Organic Pollutants
Toxic Dyes
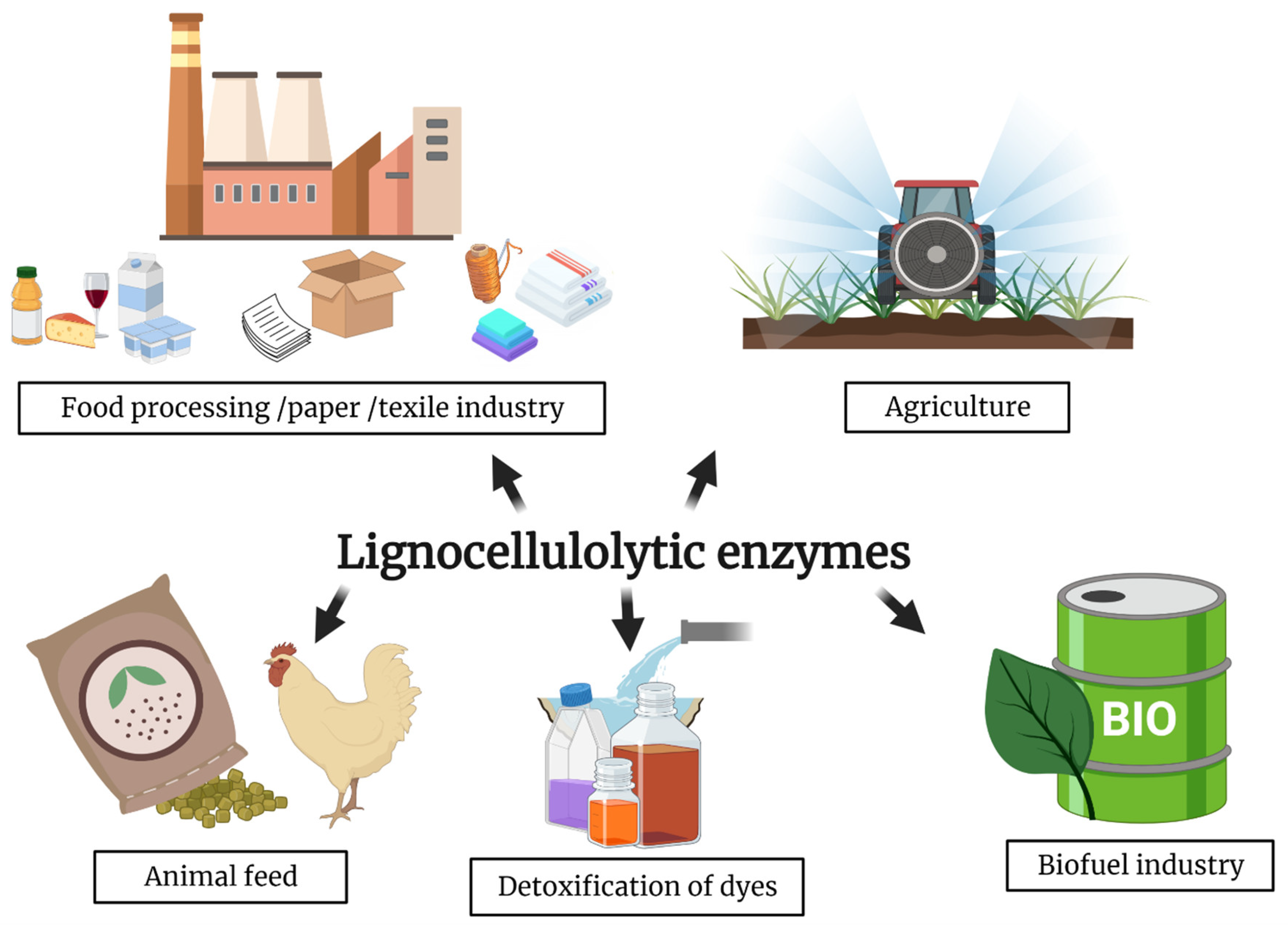
4. Lignocellulosic Biomasses
4.1. Lignin Degrading Thermozymes
4.2. Cellulose and Hemicellulose Degrading Thermozymes
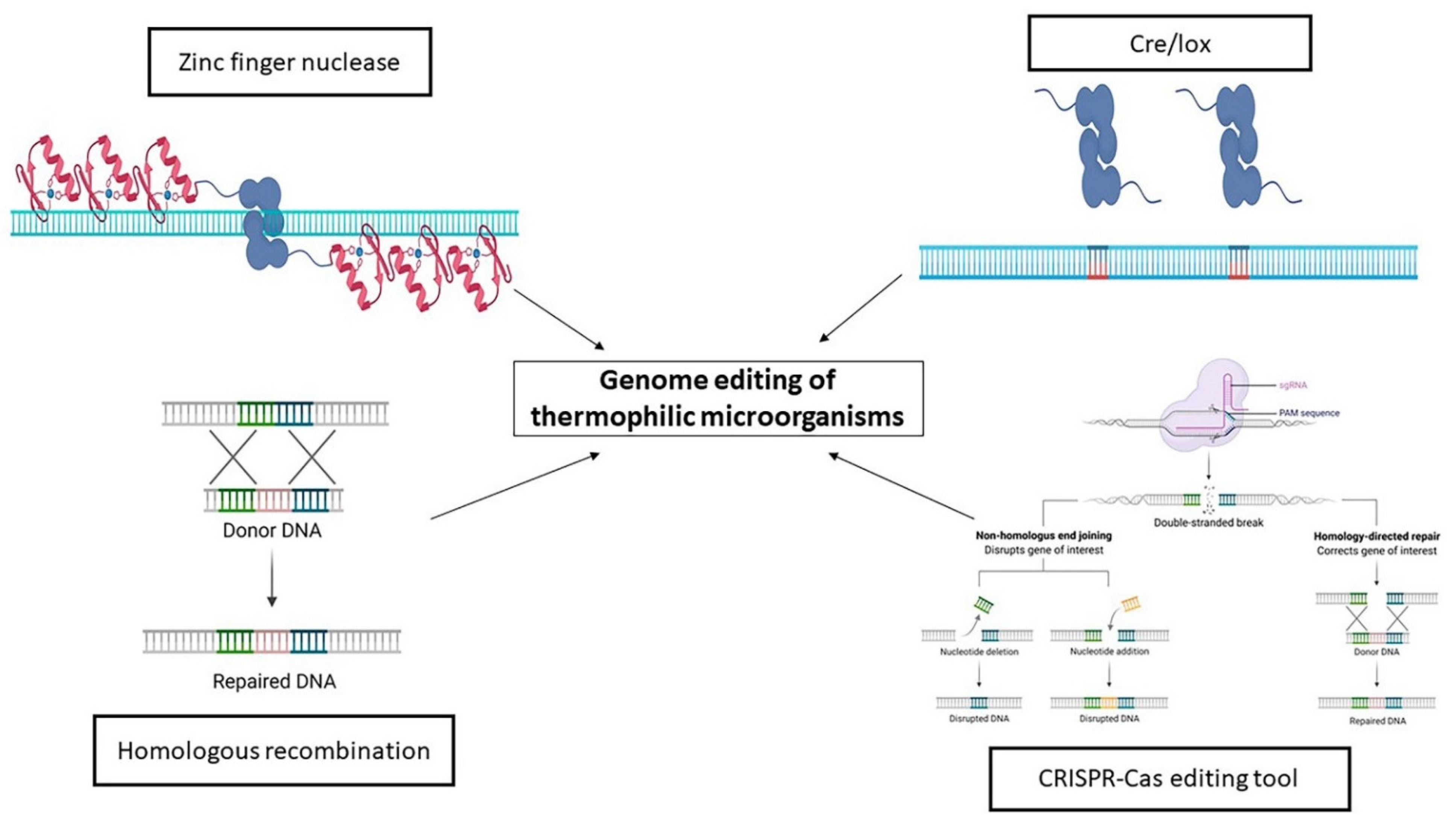
5. Engineering of Thermophiles
The Rise of the CRISPR-Cas Era
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Arora, N.K.; Panosyan, H. Extremophiles: Applications and roles in environmental sustainability. Environ. Sustain. 2019, 2, 217–218. [Google Scholar] [CrossRef] [Green Version]
- Dumorné, K.; Córdova, D.C.; Astorga-Eló, M.; Renganathan, P. Extremozymes: A potential source for industrial applications. J. Microbiol. Biotechnol. 2017, 27, 649–659. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, S.K.; Bhattacharjee, I.; Chandra, G. Biosorption of heavy metals from industrial waste water by Geobacillus thermodenitrificans. J. Hazard. Mater. 2010, 175, 117–125. [Google Scholar] [CrossRef]
- Zheng, C.; He, J.; Wang, Y.; Wang, M.; Huang, Z. Hydrocarbon degradation and bioemulsifier production by thermophilic Geobacillus pallidus strains. Bioresour. Technol. 2011, 102, 9155–9161. [Google Scholar] [CrossRef] [PubMed]
- Finch, A.; Kim, J. Thermophilic Proteins as Versatile Scaffolds for Protein Engineering. Microorganisms 2018, 6, 97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DESA UN (Department of Economic and Social Affairs of United Nations). Transforming Our World: The 2030 Agenda for Sustainable Development. In A New Era in Global Health; Springer: New York, NY, USA, 2015. [Google Scholar] [CrossRef]
- Pakshirajan, K.; Rene, E.R.; Ramesh, A. Biotechnology in Environmental Monitoring and Pollution Abatement 2015. Biomed Res. Int. 2015, 2015. [Google Scholar] [CrossRef] [PubMed]
- Schneider, F.; Kläy, A.; Zimmermann, A.B.; Buser, T.; Ingalls, M.; Messerli, P. How can science support the 2030 Agenda for Sustainable Development? Four tasks to tackle the normative dimension of sustainability. Sustain. Sci. 2019, 14, 1593–1604. [Google Scholar] [CrossRef] [Green Version]
- Barcelos, M.C.S.; Lupki, F.B.; Campolina, G.A.; Nelson, D.L.; Molina, G. The colors of biotechnology: General overview and developments of white, green and blue areas. FEMS Microbiol. Lett. 2018, 365, fny239. [Google Scholar] [CrossRef] [Green Version]
- Yadav, M.; Sehrawat, N.; Singh, M.; Kumar, V.; Sharma, A.K.; Kumar, S. Thermophilic microbes-based fuel cells: An eco-friendly approach for sustainable energy production. In Bioremediation for Environmental Sustainability; Elsevier: Amsterdam, The Netherlands, 2021; pp. 235–246. [Google Scholar]
- Hsieh, M.-C.; Chung, Y.-C. Measurement of biochemical oxygen demand from different wastewater samples using a mediator-less microbial fuel cell biosensor. Environ. Technol. 2014, 35, 2204–2211. [Google Scholar] [CrossRef]
- Straub, C.T.; Counts, J.A.; Nguyen, D.M.N.; Wu, C.-H.; Zeldes, B.M.; Crosby, J.R.; Conway, J.M.; Otten, J.K.; Lipscomb, G.L.; Schut, G.J.; et al. Biotechnology of extremely thermophilic archaea. FEMS Microbiol. Rev. 2018, 42, 543–578. [Google Scholar] [CrossRef] [Green Version]
- Lanzilli, M.; Esercizio, N.; Vastano, M.; Xu, Z.; Nuzzo, G.; Gallo, C.; Manzo, E.; Fontana, A.; D’Ippolito, G. Effect of Cultivation Parameters on Fermentation and Hydrogen Production in the Phylum Thermotogae. Int. J. Mol. Sci. 2020, 22, 341. [Google Scholar] [CrossRef]
- Sar, P.; Kazy, S.K.; Paul, D.; Sarkar, A. Metal Bioremediation by Thermophilic Microorganisms. In Thermophilic Microbes in Environmental and Industrial Biotechnology: Biotechnology of Thermophiles; Satyanarayana, T., Littlechild, J., Kawarabayasi, Y., Eds.; Springer: Dordrecht, The Netherlands, 2013; pp. 171–201. ISBN 978-94-007-5899-5. [Google Scholar]
- Satyanarayana, T.; Littlechild, J.; Kawarabayasi, Y. Thermophilic microbes in environmental and industrial biotechnology. Biotechnol. Thermoph. 2013, 3. [Google Scholar] [CrossRef]
- Rekadwad, B.; Gonzalez, J.M. Multidisciplinary involvement and potential of thermophiles. Folia Microbiol. 2019, 64, 389–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duffus, J.H. “Heavy metals”—A meaningless term? (IUPAC Technical Report). Pure Appl. Chem. 2002, 74, 793–807. [Google Scholar] [CrossRef] [Green Version]
- Pourret, O.; Hursthouse, A. It’s Time to Replace the Term “Heavy Metals” with “Potentially Toxic Elements” When Reporting Environmental Research. Int. J. Environ. Res. Public Health 2019, 16, 4446. [Google Scholar] [CrossRef] [Green Version]
- Mathew, B.B.; Singh, H.; Biju, G.; Krishnamurthy, N.B. Classification, Source, and Effect of Environmental Pollutants and Their Biodegradation. J. Environ. Pathol. Toxicol. Oncol. 2017, 36, 55–71. [Google Scholar] [CrossRef]
- Rensing, C.; Rosen, B.P. Heavy Metals Cycle (Arsenic, Mercury, Selenium, others). In Encyclopedia of Microbiology; Elsevier Inc.: Amsterdam, The Netherlands, 2009; pp. 205–219. ISBN 9780123739445. [Google Scholar]
- Finkelstein, J. Metalloproteins. Nature 2009, 460, 813. [Google Scholar] [CrossRef] [Green Version]
- Boer, J.L.; Mulrooney, S.B.; Hausinger, R.P. Nickel-dependent metalloenzymes. Arch. Biochem. Biophys. 2014, 544, 142–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janusz, G.; Pawlik, A.; Świderska-Burek, U.; Polak, J.; Sulej, J.; Jarosz-Wilkołazka, A.; Paszczyński, A. Laccase properties, physiological functions, and evolution. Int. J. Mol. Sci. 2020, 21, 966. [Google Scholar] [CrossRef] [Green Version]
- Men, C.; Liu, R.; Wang, Q.; Guo, L.; Shen, Z. The impact of seasonal varied human activity on characteristics and sources of heavy metals in metropolitan road dusts. Sci. Total Environ. 2018, 637–638, 844–854. [Google Scholar] [CrossRef]
- Vhahangwele, M.; Khathutshelo, L.M. Environmental Contamination by Heavy Metals. In Heavy Metals; Hosam, S., Ed.; IntechOpen: London, UK, 2018; pp. 115–133. ISBN 978-1-78923-361-2. [Google Scholar]
- Cole, M.; Lindeque, P.; Halsband, C.; Galloway, T.S. Microplastics as contaminants in the marine environment: A review. Mar. Pollut. Bull. 2011, 62, 2588–2597. [Google Scholar] [CrossRef]
- Horta Arduin, R.; Mathieux, F.; Huisman, J.; Blengini, G.A.; Charbuillet, C.; Wagner, M.; Baldé, C.P.; Perry, N. Novel indicators to better monitor the collection and recovery of (critical) raw materials in WEEE: Focus on screens. Resour. Conserv. Recycl. 2020, 157, 104772. [Google Scholar] [CrossRef] [PubMed]
- European Commission. Raw Materials Demand for Wind and Solar PV Technologies in the Transition towards a Decarbonised Energy System; European Commission: Brussels, Belgium, 2020. [Google Scholar]
- Pedone, E.; Bartolucci, S.; Fiorentino, G. Sensing and adapting to environmental stress: The archaeal tactic. Front. Biosci. 2004, 9, 2909–2926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ranawat, P.; Rawat, S. Metal-tolerant thermophiles: Metals as electron donors and acceptors, toxicity, tolerance and industrial applications. Environ. Sci. Pollut. Res. 2017, 1–29. [Google Scholar] [CrossRef]
- Panyushkina, A.E.; Babenko, V.V.; Nikitina, A.S.; Selezneva, O.V.; Tsaplina, I.A.; Letarova, M.A.; Kostryukova, E.S.; Letarov, A. V Sulfobacillus thermotolerans: New insights into resistance and metabolic capacities of acidophilic chemolithotrophs. Sci. Rep. 2019, 9, 15069. [Google Scholar] [CrossRef] [Green Version]
- Aulitto, M.; Gallo, G.; Puopolo, R.; Mormone, A.; Limauro, D.; Contursi, P.; Piochi, M.; Bartolucci, S.; Fiorentino, G. Genomic Insight of Alicyclobacillus mali FL18 Isolated From an Arsenic-Rich Hot Spring. Front. Microbiol. 2021, 12. [Google Scholar] [CrossRef]
- Bruins, M.R.; Kapil, S.; Oehme, F.W. Microbial resistance to metals in the environment. Ecotoxicol. Environ. Saf. 2000, 45, 198–207. [Google Scholar] [CrossRef] [PubMed]
- Cánovas, D.; Cases, I.; De Lorenzo, V. Heavy metal tolerance and metal homeostasis in Pseudomonas putida as revealed by complete genome analysis. Environ. Microbiol. 2003, 5, 1242–1256. [Google Scholar] [CrossRef]
- Cazorla, F.M.; Arrebola, E.; Sesma, A.; Pérez-García, A.; Codina, J.C.; Murillo, J.; de Vicente, A. Copper Resistance in Pseudomonas syringae Strains Isolated from Mango Is Encoded Mainly by Plasmids. Phytopathology 2002, 92, 909–916. [Google Scholar] [CrossRef] [Green Version]
- Cervantes, C.; Campos-García, J.; Devars, S.; Gutiérrez-Corona, F.; Loza-Tavera, H.; Torres-Guzmán, J.C.; Moreno-Sánchez, R. Interactions of chromium with microorganisms and plants. FEMS Microbiol. Rev. 2001, 25, 335–347. [Google Scholar] [CrossRef]
- Cervantes, C.; Gutierrez-Corona, F. Copper resistance mechanisms in bacteria and fungi. FEMS Microbiol. Rev. 1994, 14, 121–137. [Google Scholar] [CrossRef] [PubMed]
- Cha, J.S.; Cooksey, D.A. Copper resistance in Pseudomonas syringae mediated by periplasmic and outer membrane proteins. Proc. Natl. Acad. Sci. USA 1991, 88, 8915–8919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.-F.; Walmsley, A.R.; Rosen, B.P. An arsenic metallochaperone for an arsenic detoxification pump. Proc. Natl. Acad. Sci. USA 2006, 103, 15617–15622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Helow, E.R.; Sabry, S.A.; Amer, R.M. Cadmium biosorption by a cadmium resistant strain of Bacillus thuringiensis: Regulation and optimization of cell surface affinity for metal cations. Biometals 2000, 13, 273–280. [Google Scholar] [CrossRef]
- Yang, H.-C.; Rosen, B.P. New mechanisms of bacterial arsenic resistance. Biomed. J. 2016, 39, 5–13. [Google Scholar] [CrossRef] [Green Version]
- Schelert, J.; Dixit, V.; Hoang, V.; Simbahan, J.; Drozda, M.; Blum, P. Occurrence and Characterization of Mercury Resistance in the Hyperthermophilic Archaeon Sulfolobus solfataricus by Use of Gene Disruption. J. Bacteriol. 2004, 186, 427–437. [Google Scholar] [CrossRef] [Green Version]
- Smith, A.T.; Smith, K.P.; Rosenzweig, A.C. Diversity of the metal-transporting P 1B-type ATPases. JBIC J. Biol. Inorg. Chem. 2014, 19, 947–960. [Google Scholar] [CrossRef] [Green Version]
- Bondarczuk, K.; Piotrowska-Seget, Z. Molecular basis of active copper resistance mechanisms in Gram-negative bacteria. Cell Biol. Toxicol. 2013, 29, 397–405. [Google Scholar] [CrossRef] [Green Version]
- Bartolucci, S.; Contursi, P.; Fiorentino, G.; Limauro, D.; Pedone, E. Responding to toxic compounds: A genomic and functional overview of Archaea (383 views visite). Front. Biosci. 2013, 18, 165–189. [Google Scholar]
- Ranawat, P.; Rawat, S. Stress response physiology of thermophiles. Arch. Microbiol. 2017, 199, 391–414. [Google Scholar] [CrossRef]
- Gallo, G.; Puopolo, R.; Limauro, D.; Bartolucci, S.; Fiorentino, G. Metal-Tolerant Thermophiles: From the Analysis of Resistance Mechanisms to their Biotechnological Exploitation. Open Biochem. J. 2018, 12, 149–160. [Google Scholar] [CrossRef]
- Vasudevan, N.; Jayshree, A. Extremozymes and Extremoproteins in Biosensor Applications. In Encyclopedia of Marine Biotechnology; Wiley Online Books; Wiley: Hoboken, NJ, USA, 2020; pp. 1711–1736. ISBN 9781119143802. [Google Scholar]
- Zhuang, W.Q.; Fitts, J.P.; Ajo-Franklin, C.M.; Maes, S.; Alvarez-Cohen, L.; Hennebel, T. Recovery of critical metals using biometallurgy. Curr. Opin. Biotechnol. 2015, 33, 327–335. [Google Scholar] [CrossRef] [Green Version]
- Hennebel, T.; Boon, N.; Maes, S.; Lenz, M. Biotechnologies for critical raw material recovery from primary and secondary sources: R&D priorities and future perspectives. New Biotechnol. 2015, 32, 121–127. [Google Scholar] [CrossRef]
- Garole, D.J.; Hossain, R.; Garole, V.J.; Sahajwalla, V.; Nerkar, J.; Dubal, D.P. Recycle, Recover and Repurpose Strategy of Spent Li-ion Batteries and Catalysts: Current Status and Future Opportunities. ChemSusChem 2020, 13, 3079–3100. [Google Scholar] [CrossRef]
- Castro, C.; Urbieta, M.S.; Cazón, J.P.; Donati, E.R. Metal biorecovery and bioremediation: Whether or not thermophilic are better than mesophilic microorganisms. Bioresour. Technol. 2019, 279, 317–326. [Google Scholar] [CrossRef] [PubMed]
- Ilyas, S.; Lee, J.C.; Kim, B.S. Bioremoval of heavy metals from recycling industry electronic waste by a consortium of moderate thermophiles: Process development and optimization. J. Clean. Prod. 2014, 70, 194–202. [Google Scholar] [CrossRef]
- Maleke, M.; Valverde, A.; Vermeulen, J.G.; Cason, E.; Gomez-Arias, A.; Moloantoa, K.; Coetsee-Hugo, L.; Swart, H.; Van Heerden, E.; Castillo, J. Biomineralization and bioaccumulation of europium by a thermophilic metal resistant bacterium. Front. Microbiol. 2019, 10, 81. [Google Scholar] [CrossRef]
- Antonucci, I.; Gallo, G.; Limauro, D.; Contursi, P.; Ribeiro, A.L.A.L.; Blesa, A.; Berenguer, J.; Bartolucci, S.; Fiorentino, G. Characterization of a promiscuous cadmium and arsenic resistance mechanism in Thermus thermophilus HB27 and potential application of a novel bioreporter system. Microb. Cell Fact. 2018, 17. [Google Scholar] [CrossRef] [Green Version]
- Poli, A.; Salerno, A.; Laezza, G.; di Donato, P.; Dumontet, S.; Nicolaus, B. Heavy metal resistance of some thermophiles: Potential use of α-amylase from Anoxybacillus amylolyticus as a microbial enzymatic bioassay. Res. Microbiol. 2009, 160, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Özdemir, S.; Kılınç, E.; Poli, A.; Nicolaus, B. Biosorption of Heavy Metals (Cd2+, Cu2+, Co2+, and Mn2+ ) by Thermophilic Bacteria, Geobacillus thermantarcticus and Anoxybacillus amylolyticus: Equilibrium and Kinetic Studies. Bioremediat. J. 2013, 17, 86–96. [Google Scholar] [CrossRef]
- Hetzer, A.; Daughney, C.J.; Morgan, H.W. Cadmium ion biosorption by the thermophilic bacteria Geobacillus stearothermophilus and G. thermocatenulatus. Appl. Environ. Microbiol. 2006, 72, 4020–4027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Özdemir, S.; Kilinc, E.; Poli, A.; Nicolaus, B.; Güven, K. Cd, Cu, Ni, Mn and Zn resistance and bioaccumulation by thermophilic bacteria, Geobacillus toebii subsp. decanicus and Geobacillus thermoleovorans subsp. stromboliensis. World J. Microbiol. Biotechnol. 2012, 28, 155–163. [Google Scholar] [CrossRef] [PubMed]
- Ozdemir, S.; Kılınc, E.; Yalcin, M.S.; Soylak, M.; Sen, F. A new magnetized thermophilic bacteria to preconcentrate uranium and thorium from environmental samples through magnetic solid-phase extraction. J. Pharm. Biomed. Anal. 2020, 186, 113315. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.H.; Cheng, K.C.; Liao, V.H.C. A novel approach for rapidly and cost-effectively assessing toxicity of toxic metals in acidic water using an acidophilic iron-oxidizing biosensor. Chemosphere 2017, 186, 446–452. [Google Scholar] [CrossRef] [PubMed]
- Gherghe, S.L. Considerations about recovery of critical metals using bio-metallurgy. E3S Web Conf. 2017, 18, 01020. [Google Scholar] [CrossRef] [Green Version]
- Hlihor, R.-M.; Apostol, L.-C.; Gavrilescu, M. Environmental Bioremediation by Biosorption and Bioaccumulation: Principles and Applications. In Enhancing Cleanup of Environmental Pollutants: Volume 1: Biological Approaches; Anjum, N.A., Gill, S.S., Tuteja, N., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 289–315. ISBN 978-3-319-55426-6. [Google Scholar]
- Puopolo, R.; Gallo, G.; Mormone, A.; Limauro, D.; Contursi, P.; Piochi, M.; Bartolucci, S.; Fiorentino, G. Identification of a new heavy-metal-resistant strain of Geobacillus stearothermophilus isolated from a hydrothermally active volcanic area in southern Italy. Int. J. Environ. Res. Public Health 2020, 17, 2678. [Google Scholar] [CrossRef] [Green Version]
- Johnson, D.B.; Du Plessis, C.A. Biomining in reverse gear: Using bacteria to extract metals from oxidised ores. Miner. Eng. 2015, 75, 2–5. [Google Scholar] [CrossRef]
- Donati, E.R.; Castro, C.; Urbieta, M.S. Thermophilic microorganisms in biomining. World J. Microbiol. Biotechnol. 2016, 32, 179. [Google Scholar] [CrossRef]
- Dunbar, W.S. Biotechnology and the Mine of Tomorrow. Trends Biotechnol. 2017, 35, 79–89. [Google Scholar] [CrossRef]
- Hawkes, R.B.; Franzmann, P.D.; Plumb, J.J. Moderate thermophiles including “Ferroplasma cupricumulans” sp. nov. dominate an industrial-scale chalcocite heap bioleaching operation. Hydrometallurgy 2006, 83, 229–236. [Google Scholar] [CrossRef]
- Zhou, S.; Gan, M.; Zhu, J.; Liu, X.; Qiu, G. Assessment of Bioleaching Microbial Community Structure and Function Based on Next-Generation Sequencing Technologies. Minerals 2018, 8, 596. [Google Scholar] [CrossRef] [Green Version]
- Jardine, J.L.; Stoychev, S.; Mavumengwana, V.; Ubomba-Jaswa, E. Screening of potential bioremediation enzymes from hot spring bacteria using conventional plate assays and liquid chromatography—Tandem mass spectrometry (Lc-Ms/Ms). J. Environ. Manag. 2018, 223, 787–796. [Google Scholar] [CrossRef] [PubMed]
- Opperman, D.J.; Piater, L.A.; Van Heerden, E. A novel chromate reductase from Thermus scotoductus SA-01 related to old yellow enzyme. J. Bacteriol. 2008, 190, 3076–3082. [Google Scholar] [CrossRef] [Green Version]
- Politi, J.; Spadavecchia, J.; Fiorentino, G.; Antonucci, I.; De Stefano, L. Arsenate reductase from Thermus thermophilus conjugated to polyethylene glycol-stabilized gold nanospheres allow trace sensing and speciation of arsenic ions. J. R. Soc. Interface 2016, 13, 20160629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Politi, J.; Spadavecchia, J.; Fiorentino, G.; Antonucci, I.; Casale, S.; De Stefano, L. Interaction of Thermus thermophilus ArsC enzyme and gold nanoparticles naked-eye assays speciation between As(III) and As(V). Nanotechnology 2015, 26, 435703. [Google Scholar] [CrossRef]
- Puopolo, R.; Sorrentino, I.; Gallo, G.; Piscitelli, A.; Giardina, P.; Le Goff, A.; Fiorentino, G. Self-assembling thermostable chimeras as new platform for arsenic biosensing. Sci. Rep. 2021, 11, 2991. [Google Scholar] [CrossRef] [PubMed]
- Cui, Z.; Tian, W.; Qin, H.; Wang, X.; Zhao, W. Optimal design and control of Eastman organic wastewater treatment process. J. Clean. Prod. 2018, 198, 333–350. [Google Scholar] [CrossRef]
- Rashed, M.N. Adsorption Technique for the Removal of Organic Pollutants from Water and Wastewater. In Organic Pollutants—Monitoring, Risk and Treatment; InTech: West Palm Beach, FL, USA, 2013; pp. 167–194. [Google Scholar]
- Érseková, A.; Hilscherová, K.; Klánová, J.; Giesy, J.P.; Novák, J. Effect-based assessment of passive air samples from four countries in Eastern Europe. Environ. Monit. Assess. 2014, 186, 3905–3916. [Google Scholar] [CrossRef]
- Ashraf, M.A. Persistent organic pollutants (POPs): A global issue, a global challenge. Environ. Sci. Pollut. Res. 2017, 24, 4223–4227. [Google Scholar] [CrossRef]
- Gaur, N.; Narasimhulu, K.; PydiSetty, Y. Recent advances in the bio-remediation of persistent organic pollutants and its effect on environment. J. Clean. Prod. 2018, 198, 1602–1631. [Google Scholar] [CrossRef]
- Hassanshahian, M.; Amirinejad, N.; Behzadi, M.A. Crude oil pollution and biodegradation at the Persian Gulf: A comprehensive and review study. J. Environ. Health Sci. Eng. 2020. [Google Scholar] [CrossRef] [PubMed]
- Neamah, A.I. Separation of the Petroleum System. Hilltop Rev. 2014, 7, 11. [Google Scholar]
- Copley, S.D.; Chem, N.; Author, B. Evolution of Efficient Pathways for Degradation of Anthropogenic Chemicals. Nat. Chem. Biol. 2009, 5, 559–566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whitby, C. Microbial Naphthenic Acid Degradation. Adv. Appl. Microbiol. 2010, 70, 93–125. [Google Scholar]
- Fiorentino, G.; Ronca, R.; Cannio, R.; Rossi, M.; Bartolucci, S. MarR-like transcriptional regulator involved in detoxification of aromatic compounds in Sulfolobus solfataricus. J. Bacteriol. 2007, 189, 7351–7360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fiorentino, G.; Del Giudice, I.; Bartolucci, S.; Durante, L.; Martino, L.; Del Vecchio, P. Identification and physicochemical characterization of BldR2 from Sulfolobus solfataricus, a novel archaeal member of the MarR transcription factor family. Biochemistry 2011, 50, 6607–6621. [Google Scholar] [CrossRef] [PubMed]
- Parales, R.E.; Resnick, S.M. Aromatic hydrocarbon dioxygenases. In Biodegradation and Bioremediation; Springer: Berlin, Germany, 2004; pp. 175–195. [Google Scholar]
- Gibson, D.T.; Parales, R.E. Aromatic hydrocarbon dioxygenases in environmental biotechnology. Curr. Opin. Biotechnol. 2000, 11, 236–243. [Google Scholar] [CrossRef]
- Chakraborty, J.; Suzuki-Minakuchi, C.; Okada, K.; Nojiri, H. Thermophilic bacteria are potential sources of novel Rieske non-heme iron oxygenases. AMB Express 2017, 7, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Yemashova, N.A.; Murygina, V.P.; Zhukov, D.V.; Zakharyantz, A.A.; Gladchenko, M.A.; Appanna, V.; Kalyuzhnyi, S.V. Biodeterioration of crude oil and oil derived products: A review. Rev. Environ. Sci. Bio/Technology 2007, 6, 315–337. [Google Scholar] [CrossRef]
- Koshlaf, E.; Ball, A.S. Soil bioremediation approaches for petroleum hydrocarbon polluted environments. AIMS Microbiol. 2017, 3, 25–49. [Google Scholar] [CrossRef]
- Gu, G.; Li, Z.; Zhao, D.; Zhao, C. Isolation and characterization of a thermophilic oil-degrading bacterial consortium. China Pet. Process. Petrochemical Technol. 2013, 15, 82–90. [Google Scholar]
- Yang, K.; Li, L.; Ding, W.; Liu, J.; Xue, S. A full-scale thermophilic biofilter in the treatment of sludge drying exhaust: Performance, microbial characteristics and bioaerosol emission. J. Chem. Technol. Biotechnol. 2018, 93, 2216–2225. [Google Scholar] [CrossRef]
- Nazina, T.N.; Sokolova, D.S.; Shestakova, N.M.; Grigoryan, A.A.; Mikhailova, E.M.; Babich, T.L.; Lysenko, A.M.; Tourova, T.P.; Poltaraus, A.B.; Feng, Q.; et al. The Phylogenetic Diversity of Aerobic Organotrophic Bacteria from the Dagang High-Temperature Oil Field. Microbiology 2005, 74, 343–351. [Google Scholar] [CrossRef]
- Karlapudi, A.P.; Venkateswarulu, T.C.; Tammineedi, J.; Kanumuri, L.; Ravuru, B.K.; Dirisala, V.r.; Kodali, V.P. Role of biosurfactants in bioremediation of oil pollution-a review. Petroleum 2018, 4, 241–249. [Google Scholar] [CrossRef]
- Feitkenhauer, H.; Schnicke, S.; Müller, R.; Märkl, H. Determination of the kinetic parameters of the phenol-degrading thermophile Bacillus themoleovorans sp. A2. Appl. Microbiol. Biotechnol. 2001, 57, 744–750. [Google Scholar] [CrossRef] [PubMed]
- Bachas-Daunert, P.G.; Sellers, Z.P.; Wei, Y. Detection of halogenated organic compounds using immobilized thermophilic dehalogenase. Anal. Bioanal. Chem. 2009, 395, 1173–1178. [Google Scholar] [CrossRef] [PubMed]
- Yien Jun, L.; Sie Yon, L.; Mubarak, N.; Han Bing, C.; Pan, S.; Danquah, M.K.; Abdullah, E.C.; Khalid, M. An overview of immobilized enzyme technologies for dye and phenolic removal from wastewater. J. Environ. Health Sci. Eng. 2019. [Google Scholar] [CrossRef]
- Maier, J.; Kandelbauer, A.; Erlacher, A.; Cavaco-Paulo, A.; Gübitz, G.M. A New Alkali-Thermostable Azoreductase from Bacillus sp. Strain SF. Appl. Environ. Microbiol. 2004, 70, 837–844. [Google Scholar] [CrossRef] [Green Version]
- Pandey, A.; Singh, P.; Iyengar, L. Bacterial decolorization and degradation of azo dyes. Int. Biodeterior. Biodegradation 2007, 59, 73–84. [Google Scholar] [CrossRef]
- El Harfi, S.; El Harfi, A. Classifications, properties and applications of textile dyes: A review. Appl. J. Environ. Eng. Sci. 2017, 3, 3. [Google Scholar]
- Ghoreishi, S.M.; Haghighi, R. Chemical catalytic reaction and biological oxidation for treatment of non-biodegradable textile effluent. Chem. Eng. J. 2003, 95, 163–169. [Google Scholar] [CrossRef]
- Crini, G. Non-conventional low-cost adsorbents for dye removal: A review. Bioresour. Technol. 2006, 97, 1061–1085. [Google Scholar] [CrossRef] [PubMed]
- Lončar, N.; Colpa, D.I.; Fraaije, M.W. Exploring the biocatalytic potential of a DyP-type peroxidase by profiling the substrate acceptance of Thermobifida fusca DyP peroxidase. Tetrahedron 2016, 72, 7276–7281. [Google Scholar] [CrossRef]
- Deive, F.J.; Domínguez, A.; Barrio, T.; Moscoso, F.; Morán, P.; Longo, M.A.; Sanromán, M.A. Decolorization of dye Reactive Black 5 by newly isolated thermophilic microorganisms from geothermal sites in Galicia (Spain). J. Hazard. Mater. 2010, 182, 735–742. [Google Scholar] [CrossRef] [PubMed]
- Al-Balawi, T.H.A.-K.; Wood, A.L.; Solis, A.; Cooper, T.; Barabote, R.D. Anoxybacillus sp. strain UARK-01, a new thermophilic soil bacterium with hyperthermostable alkaline laccase activity. Curr. Microbiol. 2017, 74, 762–771. [Google Scholar] [CrossRef]
- Gianolini, J.E.; Britos, C.N.; Mulreedy, C.B.; Trelles, J.A. Hyperstabilization of a thermophile bacterial laccase and its application for industrial dyes degradation. 3 Biotech 2020, 10, 1–7. [Google Scholar] [CrossRef]
- Kumari, A.; Kishor, N.; Guptasarma, P. Characterization of a mildly alkalophilic and thermostable recombinant Thermus thermophilus laccase with applications in decolourization of dyes. Biotechnol. Lett. 2018, 40, 285–295. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Cheng, Y.; Du, B.; Tong, C.; Liang, S.; Han, S.; Zheng, S.; Lin, Y. Overexpression of a novel thermostable and chloride-tolerant laccase from Thermus thermophilus SG0. 5JP17-16 in Pichia pastoris and its application in synthetic dye decolorization. PLoS ONE 2015, 10, e0119833. [Google Scholar]
- Navas, L.E.; Carballo, R.; Levin, L.; Berretta, M.F. Fast decolorization of azo dyes in alkaline solutions by a thermostable metal-tolerant bacterial laccase and proposed degradation pathways. Extremophiles 2020, 24, 705–719. [Google Scholar] [CrossRef] [PubMed]
- Jeon, S.-J.; Park, J.-H. Refolding, characterization, and dye decolorization ability of a highly thermostable laccase from Geobacillus sp. JS12. Protein Expr. Purif. 2020, 173, 105646. [Google Scholar] [CrossRef]
- Wang, J.; Chang, F.; Tang, X.; Li, W.; Yin, Q.; Yang, Y.; Hu, Y. Bacterial laccase of Anoxybacillus ayderensis SK3-4 from hot springs showing potential for industrial dye decolorization. Ann. Microbiol. 2020, 70, 1–9. [Google Scholar] [CrossRef]
- Sadeghian, I.; Rezaie, Z.; Rahmatabadi, S.S.; Hemmati, S. Biochemical insights into a novel thermo/organo tolerant bilirubin oxidase from Thermosediminibacter oceani and its application in dye decolorization. Process Biochem. 2020, 88, 38–50. [Google Scholar] [CrossRef]
- Chen, G.; An, X.; Feng, L.; Xia, X.; Zhang, Q. Genome and transcriptome analysis of a newly isolated azo dye degrading thermophilic strain Anoxybacillus sp. Ecotoxicol. Environ. Saf. 2020, 203, 111047. [Google Scholar] [CrossRef]
- Chen, Y.; Feng, L.; Li, H.; Wang, Y.; Chen, G.; Zhang, Q. Biodegradation and detoxification of Direct Black G textile dye by a newly isolated thermophilic microflora. Bioresour. Technol. 2018, 250, 650–657. [Google Scholar] [CrossRef]
- Sarsaiya, S.; Jain, A.; Kumar Awasthi, S.; Duan, Y.; Kumar Awasthi, M.; Shi, J. Microbial dynamics for lignocellulosic waste bioconversion and its importance with modern circular economy, challenges and future perspectives. Bioresour. Technol. 2019, 291, 121905. [Google Scholar] [CrossRef]
- Bilal, M.; Wang, Z.; Cui, J.; Ferreira, L.F.R.; Bharagava, R.N.; Iqbal, H.M.N. Environmental impact of lignocellulosic wastes and their effective exploitation as smart carriers–A drive towards greener and eco-friendlier biocatalytic systems. Sci. Total Environ. 2020, 722, 137903. [Google Scholar] [CrossRef]
- Chandra, R.; Yadav, S.; Kumar, V. Microbial Degradation of Lignocellulosic Waste and Its Metabolic Products. Environ. Waste Manag. 2015, 263–312. [Google Scholar] [CrossRef]
- Houfani, A.A.; Anders, N.; Spiess, A.C.; Baldrian, P.; Benallaoua, S. Insights from enzymatic degradation of cellulose and hemicellulose to fermentable sugars—A review. Biomass Bioenergy 2020, 134, 13. [Google Scholar] [CrossRef]
- Bugg, T.D.H.; Ahmad, M.; Hardiman, E.M.; Rahmanpour, R. Pathways for degradation of lignin in bacteria and fungi. Nat. Prod. Rep. 2011, 28, 1883–1896. [Google Scholar] [CrossRef]
- Jeffries, T.W. Biodegradation of lignin and hemicelluloses. In Biochemistry of Microbial Degradation; Ratledge, C., Ed.; Springer: Dordrecht, The Netherlands, 1994; pp. 233–277. ISBN 978-94-010-4738-8. [Google Scholar]
- Valdivia, M.; Galan, J.L.; Laffarga, J.; Ramos, J. Biofuels 2020: Biorefineries based on lignocellulosic materials. Microb. Biotechnol. 2016, 9, 585–594. [Google Scholar] [CrossRef] [PubMed]
- Nair, R.B.; Lennartsson, P.R.; Taherzadeh, M.J. Bioethanol production from agricultural and municipal wastes. In Current Developments in Biotechnology and Bioengineering; Elsevier: Amsterdam, The Netherlands, 2017; pp. 157–190. [Google Scholar]
- Aditiya, H.B.; Mahlia, T.M.I.; Chong, W.T.; Nur, H.; Sebayang, A.H. Second generation bioethanol production: A critical review. Renew. Sustain. Energy Rev. 2016, 66, 631–653. [Google Scholar] [CrossRef]
- Gomes, I.; Gomes, J.; Steiner, W. Highly thermostable amylase and pullulanase of the extreme thermophilic eubacterium Rhodothermus marinus: Production and partial characterization. Bioresour. Technol. 2003, 90, 207–214. [Google Scholar] [CrossRef]
- Elleuche, S.; Schäfers, C.; Blank, S.; Schröder, C.; Antranikian, G. Exploration of extremophiles for high temperature biotechnological processes. Curr. Opin. Microbiol. 2015, 25, 113–119. [Google Scholar] [CrossRef]
- Bibra, M.; Wang, J.; Squillace, P.; Pinkelman, R.; Papendick, S.; Schneiderman, S.; Wood, V.; Amar, V.; Kumar, S.; Salem, D. Biofuels and value-added products from extremophiles. In Advances in Biotechnology; Nawani, N.N., Khetmalas, M., Razdan, P.N., Pandey, A., Eds.; Elsevier: Amsterdam, The Netherlands, 2014; pp. 17–51. [Google Scholar]
- Thapa, S.; Mishra, J.; Arora, N.; Mishra, P.; Li, H.; O’Hair, J.; Bhatti, S.; Zhou, S. Microbial cellulolytic enzymes: Diversity and biotechnology with reference to lignocellulosic biomass degradation. Rev. Environ. Sci. Bio/Technology 2020, 19, 621–648. [Google Scholar] [CrossRef]
- Chauhan, P.S.; Goradia, B.; Saxena, A. Bacterial laccase: Recent update on production, properties and industrial applications. 3 Biotech 2017, 7, 1–20. [Google Scholar] [CrossRef]
- Finore, I.; Lama, L.; Poli, A.; Di Donato, P.; Nicolaus, B. Biotechnology implications of extremophiles as life pioneers and wellspring of valuable biomolecules. In Microbial Factories; Springer: Berlin/Heidelberg, Germany, 2015; pp. 193–216. [Google Scholar]
- Rastogi, M.; Shrivastava, S. Recent advances in second generation bioethanol production: An insight to pretreatment, saccharification and fermentation processes. Renew. Sustain. Energy Rev. 2017, 80, 330–340. [Google Scholar] [CrossRef]
- Xu, F. Laccase. In Encyclopedia of Bioprocess Technology: Fermentation, Biocatalysis, and Bioseparation; Flickinger, M.C., Drew, S.W., Eds.; Wiley: New York, NY, USA, 1999; pp. 1545–1554. [Google Scholar]
- Sharma, V.; Ayothiraman, S.; Dhakshinamoorthy, V. Production of highly thermo-tolerant laccase from novel thermophilic bacterium Bacillus sp. PC-3 and its application in functionalization of chitosan film. J. Biosci. Bioeng. 2019, 127, 672–678. [Google Scholar] [CrossRef] [PubMed]
- Atalah, J.; Cáceres-Moreno, P.; Espina, G.; Blamey, J.M. Thermophiles and the applications of their enzymes as new biocatalysts. Bioresour. Technol. 2019, 280, 478–488. [Google Scholar] [CrossRef]
- Haque, R.U.; Paradisi, F.; Allers, T. Haloferax volcanii for biotechnology applications: Challenges, current state and perspectives. Appl. Microbiol. Biotechnol. 2020, 104, 1371–1382. [Google Scholar] [CrossRef] [Green Version]
- Navas, L.E.; Martínez, F.D.; Taverna, M.E.; Fetherolf, M.M.; Eltis, L.D.; Nicolau, V.; Estenoz, D.; Campos, E.; Benintende, G.B.; Berretta, M.F. A thermostable laccase from Thermus sp. 2.9 and its potential for delignification of Eucalyptus biomass. AMB Express 2019, 9, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.-W.; Lee, S.-Y.; Park, H.; Jeon, S.-J. Expression, refolding, and characterization of a small laccase from Thermus thermophilus HJ6. Protein Expr. Purif. 2015, 114, 37–43. [Google Scholar] [CrossRef]
- Fernandes, A.T.; Soares, C.M.; Pereira, M.M.; Huber, R.; Grass, G.; Martins, L.O. A robust metallo-oxidase from the hyperthermophilic bacterium Aquifex aeolicus. FEBS J. 2007, 274, 2683–2694. [Google Scholar] [CrossRef]
- Brander, S.; Mikkelsen, J.D.; Kepp, K.P. TtMCO: A highly thermostable laccase-like multicopper oxidase from the thermophilic Thermobaculum terrenum. J. Mol. Catal. B Enzym. 2015, 112, 59–65. [Google Scholar] [CrossRef]
- Aulitto, M.; Fusco, S.; Limauro, D.; Fiorentino, G.; Bartolucci, S.; Contursi, P. Galactomannan degradation by thermophilic enzymes: A hot topic for biotechnological applications. World J. Microbiol. Biotechnol. 2019, 35, 32. [Google Scholar] [CrossRef] [Green Version]
- Berger, E.; Ferreras, E.; Taylor, M.P.; Cowan, D.A. Extremophiles and their use in biofuel synthesis. In Industrial Biocatalysis; Pan Stanford Publishing Pte Ltd.: Singapore, 2014; pp. 239–282. [Google Scholar]
- Shahid, S.; Tajwar, R.; Akhtar, M.W. A novel trifunctional, family GH10 enzyme from Acidothermus cellulolyticus 11B, exhibiting endo-xylanase, arabinofuranosidase and acetyl xylan esterase activities. Extremophiles 2018, 22, 109–119. [Google Scholar] [CrossRef]
- Yadav, P.; Maharjan, J.; Korpole, S.; Prasad, G.S.; Sahni, G.; Bhattarai, T.; Sreerama, L. Production, purification, and characterization of thermostable alkaline xylanase from Anoxybacillus kamchatkensis NASTPD13. Front. Bioeng. Biotechnol. 2018, 6, 65. [Google Scholar] [CrossRef] [PubMed]
- Marcolongo, L.; La Cara, F.; Del Monaco, G.; Paixão, S.M.; Alves, L.; Marques, I.P.; Ionata, E. A novel β-xylosidase from Anoxybacillus sp. 3M towards an improved agro-industrial residues saccharification. Int. J. Biol. Macromol. 2019, 122, 1224–1234. [Google Scholar] [CrossRef] [Green Version]
- Aulitto, M.; Fusco, S.; Bartolucci, S.; Franzén, C.J.; Contursi, P. Bacillus coagulans MA-13: A promising thermophilic and cellulolytic strain for the production of lactic acid from lignocellulosic hydrolysate. Biotechnol. Biofuels 2017, 10, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aulitto, M.; Fusco, S.; Nickel, D.B.; Bartolucci, S.; Contursi, P.; Franzén, C.J. Seed culture pre-adaptation of Bacillus coagulans MA-13 improves lactic acid production in simultaneous saccharification and fermentation. Biotechnol. Biofuels 2019, 12, 45. [Google Scholar] [CrossRef] [Green Version]
- Aulitto, M.; Strazzulli, A.; Sansone, F.; Cozzolino, F.; Monti, M.; Moracci, M.; Fiorentino, G.; Limauro, D.; Bartolucci, S.; Contursi, P. Prebiotic properties of Bacillus coagulans MA-13: Production of galactoside hydrolyzing enzymes and characterization of the transglycosylation properties of a GH42 β-galactosidase. Microb. Cell Fact. 2021, 20, 1–17. [Google Scholar] [CrossRef]
- Tian, L.; Papanek, B.; Olson, D.G.; Rydzak, T.; Holwerda, E.K.; Zheng, T.; Zhou, J.; Maloney, M.; Jiang, N.; Giannone, R.J. Simultaneous achievement of high ethanol yield and titer in Clostridium thermocellum. Biotechnol. Biofuels 2016, 9, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, H.; Miao, R.; Liu, T.; Yang, L.; Yang, Y.; Chen, C.; Lei, J.; Li, Y.; He, J.; Sun, Q. A bifunctional cellulase–xylanase of a new Chryseobacterium strain isolated from the dung of a straw-fed cattle. Microb. Biotechnol. 2018, 11, 381–398. [Google Scholar] [CrossRef] [Green Version]
- Widyasti, E.; Shikata, A.; Hashim, R.; Sulaiman, O.; Sudesh, K.; Wahjono, E.; Kosugi, A. Biodegradation of fibrillated oil palm trunk fiber by a novel thermophilic, anaerobic, xylanolytic bacterium Caldicoprobacter sp. CL-2 isolated from compost. Enzyme Microb. Technol. 2018, 111, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Pereira, J.H.; Chen, Z.; McAndrew, R.P.; Sapra, R.; Chhabra, S.R.; Sale, K.L.; Simmons, B.A.; Adams, P.D. Biochemical characterization and crystal structure of endoglucanase Cel5A from the hyperthermophilic Thermotoga maritima. J. Struct. Biol. 2010, 172, 372–379. [Google Scholar] [CrossRef]
- Brumm, P.J.; Gowda, K.; Robb, F.T.; Mead, D.A. The complete genome sequence of hyperthermophile Dictyoglomus turgidum DSM 6724TM reveals a specialized carbohydrate fermentor. Front. Microbiol. 2016, 7, 1979. [Google Scholar] [CrossRef] [Green Version]
- Fusco, F.A.; Ronca, R.; Fiorentino, G.; Pedone, E.; Contursi, P.; Bartolucci, S.; Limauro, D. Biochemical characterization of a thermostable endomannanase/endoglucanase from Dictyoglomus turgidum. Extremophiles 2018, 22, 131–140. [Google Scholar] [CrossRef]
- Aulitto, M.; Fusco, F.A.; Fiorentino, G.; Bartolucci, S.; Contursi, P.; Limauro, D. A thermophilic enzymatic cocktail for galactomannans degradation. Enzyme Microb. Technol. 2018, 111, 7–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCutchen, C.M.; Duffaud, G.D.; Leduc, P.; Petersen, A.R.H.; Tayal, A.; Khan, S.A.; Kelly, R.M. Characterization of extremely thermostable enzymatic breakers (α-1, 6-galactosidase and β-1, 4-mannanase) from the hyperthermophilic bacterium Thermotoga neapolitana 5068 for hydrolysis of guar gum. Biotechnol. Bioeng. 1996, 52, 332–339. [Google Scholar] [CrossRef]
- Aulitto, M.; Fusco, S.; Fiorentino, G.; Limauro, D.; Pedone, E.; Bartolucci, S.; Contursi, P. Thermus thermophilus as source of thermozymes for biotechnological applications: Homologous expression and biochemical characterization of an α-galactosidase. Microb. Cell Fact. 2017, 16, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bala, A.; Singh, B. Cellulolytic and xylanolytic enzymes of thermophiles for the production of renewable biofuels. Renew. Energy 2019, 136, 1231–1244. [Google Scholar] [CrossRef]
- Mayer, J.; Kranz, B.; Fischer, L. Continuous production of lactulose by immobilized thermostable β-glycosidase from Pyrococcus furiosus. J. Biotechnol. 2010, 145, 387–393. [Google Scholar] [CrossRef] [PubMed]
- Suleiman, M.; Krüger, A.; Antranikian, G. Biomass-degrading glycoside hydrolases of archaeal origin. Biotechnol. Biofuels 2020, 13, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Ceballos, S.J.; Yu, C.; Claypool, J.T.; Singer, S.W.; Simmons, B.A.; Thelen, M.P.; Simmons, C.W.; VanderGheynst, J.S. Development and characterization of a thermophilic, lignin degrading microbiota. Process Biochem. 2017, 63, 193–203. [Google Scholar] [CrossRef]
- Uthandi, S.; Saad, B.; Humbard, M.A.; Maupin-Furlow, J.A. LccA, an archaeal laccase secreted as a highly stable glycoprotein into the extracellular medium by Haloferax volcanii. Appl. Environ. Microbiol. 2010, 76, 733–743. [Google Scholar] [CrossRef] [Green Version]
- Aulitto, M.; Tom, L.M.; Ceja-Navarro, J.A.; Simmons, B.A.; Singer, S.W. Whole-Genome Sequence of Brevibacillus borstelensis SDM, Isolated from a Sorghum-Adapted Microbial Community. Microbiol. Resource Announc. 2020. [Google Scholar] [CrossRef]
- Benedetti, M.; Vecchi, V.; Betterle, N.; Natali, A.; Bassi, R.; Dall’Osto, L. Design of a highly thermostable hemicellulose-degrading blend from Thermotoga neapolitana for the treatment of lignocellulosic biomass. J. Biotechnol. 2019, 296, 42–52. [Google Scholar] [CrossRef]
- Lin, L.; Xu, J. Dissecting and engineering metabolic and regulatory networks of thermophilic bacteria for biofuel production. Biotechnol. Adv. 2013, 31, 827–837. [Google Scholar] [CrossRef]
- Farkas, J.A.; Picking, J.W.; Santangelo, T.J. Genetic techniques for the archaea. Annu. Rev. Genet. 2013, 47, 539–561. [Google Scholar] [CrossRef]
- Farkas, J.; Stirrett, K.; Lipscomb, G.L.; Nixon, W.; Scott, R.A.; Adams, M.W.W.; Westpheling, J. Recombinogenic properties of Pyrococcus furiosus strain COM1 enable rapid selection of targeted mutants. Appl. Environ. Microbiol. 2012, 78, 4669–4676. [Google Scholar] [CrossRef] [Green Version]
- Maezato, Y.; Johnson, T.; McCarthy, S.; Dana, K.; Blum, P. Metal resistance and lithoautotrophy in the extreme thermoacidophile Metallosphaera sedula. J. Bacteriol. 2012, 194, 6856–6863. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Cooper, T.E.; Krause, D.J.; Whitaker, R.J. Augmenting the genetic toolbox for Sulfolobus islandicus with a stringent positive selectable marker for agmatine prototrophy. Appl. Environ. Microbiol. 2013, 79, 5539–5549. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, H.; Murakami, A.; Yoshida, K. Counterselection system for Geobacillus kaustophilus HTA426 through disruption of pyrF and pyrR. Appl. Environ. Microbiol. 2012, 78, 7376–7383. [Google Scholar] [CrossRef] [Green Version]
- Nagy, A. Cre recombinase: The universal reagent for genome tailoring. Genesis 2000, 26, 99–109. [Google Scholar] [CrossRef]
- Togawa, Y.; Nunoshiba, T.; Hiratsu, K. Cre/lox-based multiple markerless gene disruption in the genome of the extreme thermophile Thermus thermophilus. Mol. Genet. Genomics 2018, 293, 277–291. [Google Scholar] [CrossRef]
- Hidaka, Y.; Hasegawa, M.; Nakahara, T.; Hoshino, T. The entire population of Thermus thermophilus cells is always competent at any growth phase. Biosci. Biotechnol. Biochem. 1994, 58, 1338–1339. [Google Scholar] [CrossRef]
- Blesa, A.; Baquedano, I.; Quintáns, N.G.; Mata, C.P.; Castón, J.R.; Berenguer, J. The transjugation machinery of Thermus thermophilus: Identification of TdtA, an ATPase involved in DNA donation. PLoS Genet. 2017, 13, e1006669. [Google Scholar] [CrossRef] [PubMed]
- Swarts, D.C.; Jore, M.M.; Westra, E.R.; Zhu, Y.; Janssen, J.H.; Snijders, A.P.; Wang, Y.; Patel, D.J.; Berenguer, J.; Brouns, S.J.J. DNA-guided DNA interference by a prokaryotic Argonaute. Nature 2014, 507, 258–261. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.G.; Cha, J.; Chandrasegaran, S. Hybrid restriction enzymes: Zinc finger fusions to Fok I cleavage domain. Proc. Natl. Acad. Sci. USA 1996, 93, 1156–1160. [Google Scholar] [CrossRef] [Green Version]
- Li, T.; Liu, B.; Spalding, M.H.; Weeks, D.P.; Yang, B. High-efficiency TALEN-based gene editing produces disease-resistant rice. Nat. Biotechnol. 2012, 30, 390–392. [Google Scholar] [CrossRef] [PubMed]
- Cava, F.; Laptenko, O.; Borukhov, S.; Chahlafi, Z.; Blas-Galindo, E.; Gómez-Puertas, P.; Berenguer, J. Control of the respiratory metabolism of Thermus thermophilus by the nitrate respiration conjugative element NCE. Mol. Microbiol. 2007, 64, 630–646. [Google Scholar] [CrossRef] [PubMed]
- Ishino, Y.; Shinagawa, H.; Makino, K.; Amemura, M.; Nakata, A. Nucleotide sequence of the iap gene, responsible for alkaline phosphatase isozyme conversion in Escherichia coli, and identification of the gene product. J. Bacteriol. 1987, 169, 5429–5433. [Google Scholar] [CrossRef] [Green Version]
- Pourcel, C.; Salvignol, G.; Vergnaud, G. CRISPR elements in Yersinia pestis acquire new repeats by preferential uptake of bacteriophage DNA, and provide additional tools for evolutionary studies. Microbiology 2005, 151, 653–663. [Google Scholar] [CrossRef] [Green Version]
- Mohanraju, P.; Makarova, K.S.; Zetsche, B.; Zhang, F.; Koonin, E.V.; Van der Oost, J. Diverse evolutionary roots and mechanistic variations of the CRISPR-Cas systems. Science 2016, 353, aad5147. [Google Scholar] [CrossRef] [Green Version]
- Barrangou, R.; Fremaux, C.; Deveau, H.; Richards, M.; Boyaval, P.; Moineau, S.; Romero, D.A.; Horvath, P. CRISPR provides acquired resistance against viruses in prokaryotes. Science 2007, 315, 1709–1712. [Google Scholar] [CrossRef] [PubMed]
- Jinek, M.; Chylinski, K.; Fonfara, I.; Hauer, M.; Doudna, J.A.; Charpentier, E. A Programmable Dual-RNA–Guided DNA Endonuclease in Adaptive Bacterial Immunity. Science 2012, 337, 816–821. [Google Scholar] [CrossRef] [PubMed]
- Pennisi, E. The CRISPR Craze. Science 2013, 341, 833–836. [Google Scholar] [CrossRef] [PubMed]
- Mougiakos, I.; Mohanraju, P.; Bosma, E.F.; Vrouwe, V.; Bou, M.F.; Naduthodi, M.I.S.; Gussak, A.; Brinkman, R.B.L.; Van Kranenburg, R.; Van Der Oost, J. Characterizing a thermostable Cas9 for bacterial genome editing and silencing. Nat. Commun. 2017, 8, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Mohanraju, P.; Mougiakos, I.; Albers, J.; Mabuchi, M.; Fuchs, R.T.; Curcuru, J.L.; Van Kranenburg, R.; Robb, G.B.; Van Der Oost, J. Development of a Cas12a-Based Genome Editing Tool for Moderate Thermophiles. Cris. J. 2021, 4, 82–91. [Google Scholar] [CrossRef]
- Swarts, D.C.; Jinek, M. Cas9 versus Cas12a/Cpf1: Structure–function comparisons and implications for genome editing. Wiley Interdiscip. Rev. RNA 2018, 9, e1481. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Heavy Metals Tolerant Thermophiles | ||||
|---|---|---|---|---|
| Application | Target | Microorganism | Temperature | Ref. |
| Bioleaching | Cu, Zn, Ni, Cd, Al, Cr, Pb | Consortium of Sulfobacillus thermosulfidooxidans and Thermoplasma acidophilum | 45 °C | [53] |
| Biosorption | Ag, Cd, Co, Cr, Cu, Fe, Pb, Zn | Geobacillus thermodenitrificans | 60 °C | [3] |
| Biomineralization and Bioaccumulation | Eu | T. scotoductus SA-01 | 65 °C | [54] |
| Biosensing | Cd, As | T. thermophilus HB27 | 70 °C | [55] |
| Biosensing | Ni, Zn, Co, Hg, Mn, Cr, Cu, Fe, Cd | A. amylolyticus | 60 °C | [56] |
| Biosorption | Cd, Cu, Co, Mn | Geobacillus thermodenitrificans and A. amylolyticus | 60 °C | [57] |
| Biosorption | Cd | Geobacillus stearothemophilus | 60 °C | [58] |
| Bioaccumulation | Cd, Cu, Ni, Mn, Zn | Geobacillus toebii subsp. Decanicus Geobacillus thermoleovorans subsp. stromboliensis | 60 °C | [59] |
| Biosorption | U, Th | B. cereus SO-14 | 65 °C | [60] |
| Biosensing | As, Cd, Hg, Pb | Acidibacillus ferrooxidans | 45 °C | [61] |
| Organic Compounds Degrading Thermophiles | ||||
|---|---|---|---|---|
| Bioprocess | Target | Microorganisms | Temperature | Ref. |
| Biodegradation | Crude oil | Consortium of Bacillus, Geobacillus and Clostridium | 55 °C | [91] |
| Biodegradation | Hydrocarbons | Geobacillus pallidus | 30–70 °C | [4] |
| Biofilter | Volatile Organic Compounds (VOCs) | Consortium of 25 genera belonging to Alphaproteobacteria, Betaproteobacteria, Gammaproteobacteria, Deltaproteobacteria, Flavobacteriia, Sphingobacteriia, and Bacilli classes | 50–60 °C | [92] |
| Biodegradation | Phenolic compounds | Bacillus thermoleovorans sp. A2 | 65 °C | [95] |
| Biodegradation | Hydrocarbons | Consortium of Geobacillus and Thermoattinomyces spp. | 60 °C | [93] |
| Microorganisms | Substrates | Ref. |
|---|---|---|
| Anoxybacillus pushchinoensis, Anoxybacillus kamchatkensis and Anoxybacillus flavithermus | Reactive Black 5 | [104] |
| Anoxybacillus sp. | Congo red | [105] |
| G. stearothermophilus | Remazol Brilliant Blue R, Methyl Orange, Malachite Green (MG) and Indigo Carmine | [106] |
| T. thermophilus HB27 | Dye orange, Acid red dye, green dye, naphthol brilliant blue, Remazol brilliant blue, congo red | [107] |
| T. thermophilus SG0.5JP17-16 | Congo Red, Reactive Black B and Reactive Black WNN, and Remazol Brilliant Blue R | [108] |
| Thermus sp. 2.9 | Xylidine, RBBR, Gentian Violet, Methyl Orange | [109] |
| Geobacillus sp. JS12 | Congo red, Malachite green | [110] |
| Anoxybacillus ayderensis SK3-4 | Direct blue 6, acid black 1, direct green 6, direct black 19, and acid blue 93 | [111] |
| Thermosediminibacter oceani | Malachite green (MG) and Congo red | [112] |
| Lignocellulosic Wastes | Lignin (%) | Hemicellulose (%) | Cellulose (%) |
|---|---|---|---|
| Softwood stems | 25–35 | 25–35 | 45–50 |
| Hardwood stems | 18–25 | 24–40 | 40–55 |
| Miscellaneous Corn stover | 19 | 22 | 39 |
| Wheat straw | 15 | 50 | 30 |
| Rice straw | 18 | 24 | 33 |
| Nutshells | 30–40 | 25–30 | 25–30 |
| Peels | 14–20 | 11 | 4 |
| Shells | 26–30 | 20–25 | 40–45 |
| Sorted refuse | 20 | 20 | 60 |
| Swine waste | n/a | 28 | 6 |
| Solid cattle manure | 2.7–5.7 | 1.4–3.3 | 1.6–4.7 |
| Grass | 10–25 | 35–50 | 30–40 |
| Cotton seed hairs | 0 | 5–20 | 80–95 |
| Leaves | 0 | 80–85 | 15–20 |
| Sawdust | 14–34 | 71–89 | 31–64 |
| Paper | 0–15 | 0 | 85–99 |
| Newspaper | 14–19 | 25–40 | 40–55 |
| Wastepaper from chemical pulps | 5–10 | 10–20 | 60–70 |
| Primary wastewater solids | 24–29 | n/a | 8–15 |
| Lignocellulosic Component | Microorganism | Temperature | Ref. |
|---|---|---|---|
| Lignin | A. aeolicus | 89 °C | [137] |
| Bacillus sp. PC-3 | 55–92 °C | [132] | |
| Bacillus sp. FNT | 50–55 °C | [133] | |
| H. volcanii | 50 °C | [160] | |
| Thermus sp. 2.9 | 65 °C | [135] | |
| T. thermophilus HJ6 | 80 °C | [136] | |
| T. thermophilus HB27 | 70 °C | [107] | |
| T. terrenum | 67 °C | [138] | |
| Fungal and bacterial consortium | 55 °C | [159] | |
| Cellulose and hemicellulose | A. cellulolyticus 11B | 70 °C | [141] |
| A. kamchatkensis NASTPD13 | 60 °C | [142] | |
| Anoxybacillus sp. 3M | 55 °C | [143] | |
| B. coagulans MA-13 | 55 °C | [144] | |
| Brevibacillus borstelensis SDM | 50 °C | [161] | |
| C. thermocellum | 60 °C | [147] | |
| Chryseobacterium sp. | 55 °C | [148] | |
| Caldicoprobacter sp. CL-2 | 60–75 °C | [149] | |
| D. turgidum | 75–80 °C | [151] | |
| T. maritima | 80 °C | [150] | |
| T. neapolitana 5068 | 70–80 °C | [162] | |
| T. thermophilus HB27 | 70 °C | [107] | |
| P. furiosus | 100 °C | [158] | |
| S. shibatae | 80 °C | [156] | |
| S. solfataricus | 80 °C | [158] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gallo, G.; Puopolo, R.; Carbonaro, M.; Maresca, E.; Fiorentino, G. Extremophiles, a Nifty Tool to Face Environmental Pollution: From Exploitation of Metabolism to Genome Engineering. Int. J. Environ. Res. Public Health 2021, 18, 5228. https://doi.org/10.3390/ijerph18105228
Gallo G, Puopolo R, Carbonaro M, Maresca E, Fiorentino G. Extremophiles, a Nifty Tool to Face Environmental Pollution: From Exploitation of Metabolism to Genome Engineering. International Journal of Environmental Research and Public Health. 2021; 18(10):5228. https://doi.org/10.3390/ijerph18105228
Chicago/Turabian StyleGallo, Giovanni, Rosanna Puopolo, Miriam Carbonaro, Emanuela Maresca, and Gabriella Fiorentino. 2021. "Extremophiles, a Nifty Tool to Face Environmental Pollution: From Exploitation of Metabolism to Genome Engineering" International Journal of Environmental Research and Public Health 18, no. 10: 5228. https://doi.org/10.3390/ijerph18105228