
Abstract
Large-sized cricetid rodents (Cricetodotontini) immigrated to Central Europe at the beginning of the Middle Miocene Climate Transition at ca. 15 My and are thus of great value for biostratigraphic and palaeoecologic purposes. An important fossil material belonging to a relatively small species of Mixocricetodon from the North Alpine Foreland Basin is described. It is a rare component in faunas of equivalent age, and the new material confirms the synonymy with the genus Lartetomys. The lineage L. mirabilis-L dehmi is documented in its chronostratigraphic framework, and the origin of the genus is discussed but remains uncertain.
Similar content being viewed by others


Introduction
Cricetodotontini are large cricetid rodents that immigrated to Central Europe at the beginning of the Middle Miocene Climate Transition (e.g. Prieto and Rummel 2016 and references therein), a period marked by a cooling trend. The tribe is therefore of great biostratigraphic value (Rummel 2000, Rummel and Kälin 2003, López Guerrero et al. 2014, Prieto and Rummel 2016), and well-documented evolutionary lines are recognised (López-Guerrero et al. 2015). However, one genus remains rare and enigmatic.
Mein and Freudenthal (1971a, b, 1981) defined the genus Lartetomys with only a few rare teeth discovered in the karst deposits from Vieux-Collonges (France; lower to middle Miocene, see § Material and methods). They defined two species of different sizes but expressed doubts about the homogeneity of the genus. The smallest species, L. zapfei, was then definitively related to democricetodontines by Garapich and Kälin (1999), and then assigned to a new genus by Theocharopoulos (2000) through the study of the rich material of the Greek early Miocene. The larger L. mirabilis, closer to Cricetodon, was only known from two teeth from Vieux-Collonges. Lartetomys mirabilis was also listed among the fauna of the sands of Castelnau-d'Arbieu (France) without being described (Bulot et al. 1992). Finally three molars, partly damaged, were documented in Contres (France; Augé et al. 2002).
Mein (2003) then proposed a compilation of the literature on Eurasian rodents and noted (p: 411) that Lartetomys was found in Germany under the name Mixocricetodon dehmi (proposal followed by López-Guerrero et al. 2015). This very large Cricetodontini species was based on a rich fossil material from the middle Miocene karst fillings of Petersbuch, and very rare specimens from the Bavarian molasse (Rummel 1997). A second species of Mixocricetodon, unnamed and smaller than M. dehmi, was documented at Petersbuch 41 “rechte Spalte [right fissure]”. The only representative of this species, a mandible including the incisor, as well as the m2 and m3, is considered by Rummel (1997) to be more primitive than M. dehmi. The biostratigraphic potential of the genus was confirmed when Seehuber (2009) recognised a M1 and a fragment of m2 of a small-sized Mixocricetodon in Burg-Balzhausen (middle Miocene; Germany). However, the fossil material is still too scarce to study the phylogenetic relationships of the above-mentioned species.
In this context, the locality Höll (German part of the northern Alpine basin, middle Miocene, see § Material and methods) is of primary importance. Indeed, among the many remains initially attributed to Cricetodon (Scholz 1986a), a significant population of small-sized Lartetomys (=Mixocricetodon) could be isolated. Thus, the purpose of this work is, in view of this new material, to describe the evolution of the genus.
Material and methods
The terminology used in the description of the molars mainly follows Freudenthal et al. (1994) and Rummel (1998). Details are in Prieto (2012). The term “prelophid” proposed by Freudenthal (1985) refers to a transversal crest developed halfway between the anteroconid and the protoconid/metaconid pair. The use of the term anteromesoloph follows Heissig (1995). Hypolophulid II is defined in Rummel (1997). The terminology for the molar planation follows Hershkovitz (1967: fig. 4).
The molars were photographed and then digitally measured using the method proposed by Prieto et al. (2016: fig. 3).
The fossils considered herein are housed in the collections of the Naturkundemuseum und Römisches Museum im Zumsteinhaus at Kempten, Germany (possible change of institution in the future; Höll, M-0S-2177 for all specimens originally assigned to Cricetodon; the Lartetomys specimens have been isolated and each fossil labelled), the Bayerische Staatssammlung für Paläontologie und Geologie at Munich, Germany (holotype and paratype of Mixocricetodon dehmi, 1997 I 1 and 2; Laimering 3, 1989 XX; Göttschlag, 1986 XXIII), and the Naturmuseum of the city of Augsburg, Germany (Burg-Balzhausen; 2005-716/1927 and 2005-772/1927; Petersbuch 108, fossils are under study).
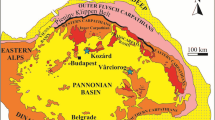
The fossil record of the genus Lartetomys, as well as the geographical position of the localities, is summarised in Fig. 1. The local time scale for Germany follows Prieto and Rummel (2016; See also Rochool et al. 2017 for radioisotopic datings) with comments in Prieto et al. (2018). The use of the MN system is problematic for the time range considered in this paper and allows only for approximate long-distance correlation (e.g. Van der Meulen et al. 2011). We discuss the relative dating of the localities considered herein in the following:

Fossil record of Lartetomys in its geographic and chronostratigraphic context (Prieto and Rummel 2016). Green colour shows the occurrence and the stratigraphic range of L.mirabilis. L. dehmi is presented in red
Germany
In Bavaria, the richest sites come from the karst fillings of Peterbuch. Petersbuch 31 “liegend [lying]” and Petersbuch 39 faunas, for which Mixocriceton material has been described, are correlated to MN 6 by Rummel (1997), as are Petersbuch 32 and 33 (Rummel 2000). These faunas contain Cricetodon aff. aureus and are therefore correlated to OSM F. To avoid any confusion, it is important to note here that the upper part of the Petersbuch 31 fissure is more recent, and characterised, among others, by the presence of C. aff. sansaniensis and Megacricetodon aff. gregarius (Rummel 2000, Prieto 2007). Petersbuch 41 “rechte Spalte [right fissure]” is older, MN 5 for Rummel (1997, 2000) who recognises M. lappi and C. aff. meini. This association allows a correlation of the fauna with OSMF-EBE. Petersbuch 108, on the other hand, is currently under study. Pickford (2017) describes a new species of Suoidea, Choeromorus petersbuchensis, which he dates at about 13.8 Ma (the base of MN 6 in his study). The small mammal fauna contains Cricetodontini related to those of Petersbuch 68. In contrast, it contains Galerix exilis instead of Parasorex socialis (Prieto and Rummel 2009). Thus, Petersbuch 108 is slightly older. This indicates that this fauna is most probably the youngest of those studied for Peterbuch (probably OSM?-KIR).
The German molasse sites generally contain only rare specimens of Lartetomys. A notable exception is Höll whose fossil material will be presented in this work. The locality is localised in the southern part of the North Alpine Foreland Basin, southwest from Isny (Allgäu, Scholz 1986a, b). Scholz (1986a) correlates the fauna to the MN 6. The preliminary studies on the evolutionary level of Cricetodon from Höll indicate that the locality correlates most probably to the German local time scale unit OSM F (see details in Prieto and Scholz 2013, Prieto et al. 2017). Burg-Balzhausen c.a. 1.4 km north-east from the church of Balzhausen is correlated to OSM E by Seehuber (2009), and the presence of Cricetodon meini allows more precise correlation to OSM E-EBE. Heissig (2006) recognises Mixocricetodon dehmi in the clay pit situated 1 km W-NW of the village of Laimering. In Laimering 3, in a 10–15-cm-thick layer situated 20–30 cm under the main bentonite (14.93 Ma after Rocholl et al. 2017), we identified one possible Lartetomys M3. The fauna is correlated to OSM F thanks to the presence of C. aureus. Laimering 4a, a clay pit at the eastern margin of the village, has not been studied herein. Mixocricetodon dehmi is listed in Ziemetshausen 1b by Rummel (1997). This locality, a sand quarry, is located about 1.3 km from the village church. The fauna, found in redeposited sediments dating from the impact of the Ries meteorite, is correlated with OSM F (Heissig 1997). Boon (1991) describes C. aff. meini in Ziemetshausen, a fauna excavated directly from the Brockhorizont (Boulder horizon), equivalent to level 1b. Among these Cricetodon, an illustrated M3 (Boon, 1991: Plate 7, fig. 3) actually belongs to Lartetomys. Similarly, Rummel (1997) lists M. dehmi in Göttschlag (OSM F; Heissig 1997). Prieto and Rummel (2016) followed this dating. Bolliger (1994) studied cricetid rodents and recognised in the faunule Cricetodon cf. sansaniensis and a medium-sized Megacricetodon which is assigned at this time to M. germanicus. This form could be rather close to M. gersii. Thus, Göttschlag is correlated here with OSM-BOH.
France
Vieux-Collonges is a karstic filling known since the second half of the nineteenth century and was excavated in a hamlet located nowadays in Collonges-aux-Mont d'Or, about 10 km northwest of Lyon. This highly diversified fauna (Mein 1958, Maridet 2003) is from the type locality of Lartetomys mirabilis (Mein and Freudenthal 1971a, 1981). The dating of the fauna is controversial, and contradictory ages have resulted from the analysis of the faunal contents (see discussion below). Traditionally, Vieux-Collonges is correlated with the MN 4/MN 5 transition (De Bruijn et al. 1992). Birosse and Le Mouné (Castenau d'Arbieu; Gers) are two faunal locations with L. mirabilis that have been excavated in a sandy horizon, at the same altitude and 700 m apart (Bulot et al. 1992). They are traditionally correlated to MN 5 (Ginsburg and Bulot 2000). The fauna of Contres was discovered 20 km south of Blois (Loir-et-Cher) in the quarry of Château-Gabillon which exposes the marine “Falun” of the Blésois. Augé et al. (2002) recognise early Miocene elements in the fauna but correlate the middle Miocene species with MN 5. L. mirabilis was identified by a fragmentary m2, as well as an m3. Finally, a Lartetomys tooth was found in the karst filling of Four (Isère), about 40 km southeast of Lyon. The filling is stratified and contains fauna ranging from the end of MN 6 to the base of MN 7+8 (Maridet et al. 2000, Maridet 2003).
Correlations between France and Central Europe
The dating of French localities is closely linked to the use of the MN system. This makes accurate correlation of the sites containing Lartetomys problematic. Correlations of localities in the Northern Alpine Foreland Basin with the MN system have been intensively discussed (e.g. Bolliger 1994, 1997, Heissig 1997, Reichenbacher et al. 2013), indicating clear diachronies and/or differences in interpretation between the European regions (e.g. Kälin and Kempf 2009). This problem has been highlighted by Van den Meulen et al. (2011), who proposed to use the MN system in a biochronological framework, and noticed that it only allows a coarse correlation over long distances.
We therefore turn to the evolutionary level of the key taxa present in the localities considered in this work.
In Germany, two evolutionary lines make it possible to date the sites for the considered time window. The first lineage is characterised by an increase in size among the Megacricetodon teeth (mainly m1) of the “M. bavaricus group” (see Prieto and Rummel (2016) for details). This lineage chronologically includes M. aff. collongensis-M. bavaricus-M. lappi. M. bourgeoisi could also belong to this lineage (Berger 2010). Oliver and Peláez-Campomanes (2013) also include in the “M. bavaricus group” the French species M. bezianensis and M. aunayi and the Spanish species M. vandermeuleni.
The second lineage concerns the genus Cricetodon. Cricetodon meini is the first Cricetodontini to migrate to Eastern and Central Europe at about 15 Ma (Boon, 1991, Heissig 1997, Daxner-Höck 2003, Hír 2013), although the tribe is well-established in Greece, Serbia, and Anatolia in the lower Miocene (e.g. De Bruijn et al. 1993, Marković et al., 2016). A rapid increase in size is then observed, leading to forms close to C. aureus (Heissig 1997, Prieto and Rummel 2016, Hír 2013, Hír and Venczel 2018). It is important to note that these species do not currently allow dating OSM F localities relative to each other.
In France, the association M. collongensis-gersii+M. germanicus+M. bavaricus+Cricetodon cf. meini is described in Contres (Augé et al. 2002). In Castelnau d'Arbieu, only one species of Megacricetodon is listed (M. aff. collongensis -gersii), but two Cricetodon, C. meini and C. aureus, are recognised. These two species are also present in Vieux-Collonges, associated with M. bourgeoisi-gersii+M. collongensis+M. lappi.
There is thus a contradiction between the regions, where species from Central Europe included in a lineage are associated at a single locality in France. It is also noted that in the typical locality of MN 5, Pontlevoy-Thenay, Cricetodon aureus is associated with M. lappi and M. collongensis, three taxa never found together in Germany. First of all, a discrepancy in taxonomic identification can be suspected. Thus, M. collongensis, a small species defined in Vieux-Collonges, is very variable and opinions on its homogeneity differ (e.g. Maridet 2003, Oliver Pérez 2015). It is reasonable to follow Oliver and Peláez-Campomanes (2013) who consider M. aff. collongensis from Central Europe as independent of the French species. It is also interesting to note that M. collongensis from Vieux-Collonges is similar in size to many populations assigned to M. minor in Central Europe (Daxner-Höck 2003, Prieto and Rummel 2016). Oliver Pérez (2015) recognises the M. collongensis-M. gersii lineage in France and Spain. She includes this lineage in the “M. primitivus group”. It could be extended in France in M. lemartineli-M. fournasi-M. roussillonensis as proposed by Aguilar (1995). As a result, the use of M. collongensis for long-distance correlation is not recommended at present. Similarly, the boundary between C. meini and C. aureus is not defined for intermediate size populations. Thus, the assignment to one species or the other is subject to interpretation. Indeed, the size of Cricetodon aureus from Pontlevoy-Thenay is within the variation of C. aff. meini from Germany (sensu Boon 1991).
The co-occurence of three species of Megacricetodon and/or two species of Cricetodon is suspect for the period considered herein. Thus, it is necessary to question the homogeneity of the fauna.
First, assignments for the karst filling of Vieux-Collonges appear questionable by the specific diversity of its fauna (see Maridet 2003 for a complete faunal list), as well as by the presence of taxa found elsewhere in different age levels. While it is commonly explained that the fauna covers the MN 4/5 transition, it seems safer to consider Vieux-Collonges as a mixture of species covering a relatively long time (JP, pers. opinion; see also De Bruijn et al. 1993: 206; Bolliger 2000: 9, 16). Excluding older forms such as Protadelomys lugdunensis, Lartetomys mirabilis would be included in a (mixed) fauna whose age ranges from OSMA to OSMF in the Northern Alpine Foreland Basin.
Augé et al. (2002) do not discuss in detail the age of Contres, even if they recognise mixtures due to the presence of Eucricetodon. The presence of the Eomyidae L. florancei, which could be reworked according to these authors, makes it possible to estimate the maximum age equivalent to OSM A. The taxonomy of Megacricetotodon of the locality needs to be revised, but it is interesting to note that only one tooth of Cricetodon has been discovered in Contres, while the genus constitutes a significant part of the fauna of small mammals when it arrived in Central Europe and is abundant in Vieux-Collonges. It can be assumed that little of the Contres material corresponds to this period. In addition, most of the harvested small mammals come from surface-collected material, not from sieving. It then seems logical that the larger species would have been over-represented. This is not the case in Contres. We can propose a minimum probable age of Contres close to OSME-EBE.
The fauna of Castelnau-d'Arbieu is unfortunately not described in detail, so it is difficult to confirm here the presence of two species of Cricetodon. We then propose an OSME-EBE-OSMF correlation.
The undetermined level of the youngest of the French faunas, Four, like all other levels of karst filling, contains the gymnure Parasorex socialis, whose appearance is sudden in Germany (Prieto and Rummel 2009; Petersbuch 68). The oldest correlation would therefore most likely be with OSM?-KIR. A form similar to Megacricetodon gregarius is a characteristic of the Swiss Helsighausen assemblage zone, not named in Germany because it is not now registered in this part of the molasse, but only in fissure fillings which were not used by Prieto and Rummel (2016) for the names and in the Steinheim basin. It seems to us therefore consistent to propose a correlation between these two zones, although it is likely that it can be extended to the base of the OSME-KLE, from which little is in fact known.
Systematic palaeontology
Order Rodentia Bowdich, 1821
Family Cricetidae Fisher von Waldheim, 1817
Subfamily Cricetodontinae Stehlin and Schaub, 1951
Genus Lartetomys Mein and Freudenthal, 1971
pars 1971a Lartetomys nov. genus, in Mein and Freudenthal, 10, 31
v. 1997Mixocricetodon n. gen., Rummel
v. 1999Mixocricetodon, Rummel
v. 2000Mixocricetodon, in Rummel, 165
v. 2003Mixocricetodon in Mein, 411
v. 2011Cricetodon dehmi, in Sen and Erbageva: 268
v. 2013Mixocricetodon, in Masini et al: 57, 60
2015Lartetomys (=Mixocricetodon), López-Guerrero et al. 417
v. 2016Mixocricetodon, in Savorelli and Masini, 4, 7, 10, 11, tables 2, 3
Original diagnosis of Lartetomys: (Mein and Freudenthal 1971a, considering two genera, namely Lartetomys and Karydomys; translated from French): Very large Cricetidae with simple dentition. The M1 has just three roots; the posterosinus of the M1 is not reduced.
Original diagnosis of Mixocricetodon (Rummel 1997; translated from German): Large Cricetodontinae with elongated, relative brachyodont and massive upper and lower molars. m3 not reduced. The anteroconid of the m1 can be deeply and widely split. In the m2, a short hypolophid II is usually developed. The M3 has complete mesoloph and posteroloph. The M1 has an anteromesoloph. All upper molars have a strong mesoloph and three roots. The entomesoloph is always missing. All mandibular molars have a strong mesolophid and a weak ectomesolophid.
Emended diagnosis: Large cricetid-rodent with relative brachyodont and massive upper and lower molars. M1 has a very short to half-length anteromesoloph. M1 and M2 with strong metalophule II which is most often directed backwards. It fuses during the wearing process with the posteroloph. The M3 has complete mesoloph and posteroloph. The m1 has an anteroconid that can be deeply and widely split. Prelophid may be present in younger species. A short and low transversal crest may connect metaconid and protoconid in older species. All upper molars have a strong mesoloph and three roots. The entomesoloph is always missing. In the m2, a short hypolophulid II may be developed. All mandibular molars have a strong mesolophid and a weak ectomesolophid.
Type locality: Fissure filling Vieux-Collonges (France), early-middle Miocene (see details in § Material and Methods).
Species included in Lartetomys: L. mirabilis Mein and Freudenthal, 1971, Mixocricetodon dehmi, Rummel 1997
Species Lartetomys dehmi (Rummel, 1997)
v. pars 1989Cricetodon cf. jotae, in Heissig, 246
v. pars 1991Cricetodon aff. meini, in Boon, fig. VIII 3
v. 1997Mixocricetodon dehmi, in Rummel
v. 1997Mixocricetodon dehmi, in Rummel, tab. 34
v. 2011Mixocricetodon dehmi, in Sen and Erbajeva, 268, fig. 5
v. 2016Mixocricetodon dehmi, in Savorelli and Masini , 4, 7, 10, 11, tables 2, 3
Original diagnosis (Rummel 1997): same as the diagnosis of Mixocricetodon.
Emended diagnosis: Largest Lartetomys species. M1 has a short to half-length anteromesoloph. Some m1 with prelophid. In the m2, a short hypolophid II is usually developed.
Type locality: Fissure filling Petersbuch 39, Germany, middle Miocene.
Species Lartetomys mirabilis Mein and Freudenthal, 1971

Lartetomys dehmi from Petersbuch 39 (a, e) and Lartetomys mirabilis from Burg-Balzhausen (b,d) and Höll (c, f; labelled M-0S-2177). a, c, and f are mirrored. a Holotype of L. dehmi, right mandible with m1-m3, Petersbuch 39, SNSB-BSPG-1997-I 1. b Left damaged m2, NMA 2005-772/1927. c Right mandible with damaged m1 and m2-m3. d Left M1, NMA 2005-716/1927. e Left maxillary with M1-M3, Petersbuch 39, SNSB-BSPG-1997-I 2. f Right maxillary with M1-M3 (f1 occlusal view; f2 labial view; f3 lingual view)

Lartetomys mirabilis from Höll. a, b, and f are mirrored. a Right M1. b Right M1. c Left M2 (maxillary bone not figured). d Left maxillary with M1-M2. e Left M3. f Right m1. g Left m1
1971a Lartetomys mirabilis, in Mein and Freudenthal, 10, 31
1971b Lartetomys mirabilis, in Mein and Freudenthal, 6
1978 Lartetomys, in Baudelot and Collier, 201
1981 Lartetomys mirabilis, in Mein and Freudenthal, 2-3, Pl. 1 figs. 1 and 2
v. pars 1986a Cricetodon sansaniensis, in Scholz, table 1
v. pars 1986b Cricetodon sansaniensis, in Scholz, table 2
v. pars 1991 Cricetodon aff. meini, in Boon, Plate 7 fig. 3.
? 1992 Lartetomys mirabilis, in Bulot et al., 535
v. 1997 Mixocricetodon sp., in Rummel
v. 1999 Mixocricetodon sp., in Rummel, tab. 34
v. 2000 Mixocricetodon [Petersbuch 41], in Rummel: 165
2002 Lartetomys mirabilis, in Augé et al. 836, figs. 14 D and E
v. 2006 Mixocricetodon dehmi, in Heissig, 95-96
v. 2008 Mixocricetodon sp. in Seehuber. 14, 125, 126, 210
v. 2009 Mixocricetodon sp. in Seehuber
2016 Lartetomys mirabilis, in Savorelli and Masini, 4, 7, 10, 11, tables 2, 3
Original diagnosis (Mein and Freudenthal 1981; translated from French): Cricetidae with brachyodont, large-sized and massive teeth with very thick enamel. Upper triradicular M1, showing a large and open posterosinus.
Emended diagnosis: Lartetomys species smaller than L. dehmi. M1 has an emerging to very short anteromesoloph. A short and low crest may transversely connect metaconid and protoconid (m1).
Type locality: Fissure filling Vieux-Collonges (France), early-middle Miocene (see details in § Material and Methods).
Höll
Fossil material: Right maxillary with M1-M3 (Fig. 2f); left maxillary with M1-M2 (Fig. 3d); left maxillary with M1-M2; right maxillary with fragmentary M1-M2; right maxillary with M1; left maxillary with M2 (Fig. 3c); right M1 (Fig. 3b); right fragmentary M1; left worn M3 (Fig. 3e); left mandible with heavily worn m1-m3; left mandible with m2-(fragmentary)-m3; right mandible in sediment with fragmentary m1 + m2-m3 (Fig. 2c); right m1 (Fig. 3f); left m1 (Fig.3g); right fragmentary m3.
Measurements (complete teeth only; see also Fig. 4): M1: 3.35x2.27, 3.39x2.10, 3.51x2.21, 3.72x2.34, 3.77x2.40, 3.79x2.45; M2: 2.87x2.42, 2.50x2.19, 2.60x2.47, 2.76x2.22; M3: 2.41x2.38, 2.37x2.29; m1: 3.21x1.88; 3.30x1.96; 3.40x2.06; m2: 2.77x2.41, 2.81x2.50; m3: 3.00x2.28. The measurements of the mandibles are given in the description.

Scatter diagrams comparing the sizes (Length/Width) of Lartetomys species. Data of Höll from this paper, Vieux-Collonges from Mein and Freudenthal (1981), Petersbuch from Rummel (1997), Four from Maridet et al. (2000), Burg-Balzhausen from Seehuber (2009). Colours refer to localities. The cross correspond to the four dots Length (L) maximum (max) × Width (W) mean, Lmin (minimum) × Wmean, Lmean × Wmin and Lmean × Wmax. The intersection of the lines thus gives the Lmean × Wmean
Description
The enamel of the teeth is wrinkled, especially on the lingual (M1-M3) and anterior walls (M1). The adult molars are bi-levelled to terraced.
M1: One molar shows a small cingulum at the anterior wall of the anterocone (Fig. 3d). The anterocone is divided into two sub-equal cusps. The anterolophule connects the lingual cusp which extends lingually into a strong anteroloph. The labial anteroloph arm is less developed, but a small style is present close to the anterocone. A slight budding on the anterolophule indicates the presence of an early onset of the anteromesoloph. The posterior arm of the protocone runs transversally, as well as the protoloplule II. Both connect to the entoloph, forming a T-like structure. The angle between the ridges may be more pronounced, forming a Y-shaped structure. The ridges remain well-differentiated, in a very different way from what is observed in the Cricetodon from the same site, where these ridges merge with the mesoloph. In two, the mesoloph is of medium size, directed backwards, and joins the metacone. The crest is transverse in three molars but still meets the metacone. Three mesolophs are very short. The protocone spur is very short. The anterior arm of the hypocone is very strong. The metalophule II, also strong, is most often directed backwards, and fuses during wear with the posteroloph. One molar has a transversal metalophule. The lingual sinus is closed by a strong cingulum. Its labial counterpart is clearly narrower. As far as it can be observed, three roots are present. It cannot be determined whether or not the broad lingual root is divided at its apical end.
M2: The teeth are almost rectangular. While the outer and inner margins of Cricetodon teeth from Höll show a concavity at the sinus area (thus highlighting the cusps), these margins are convex in Lartetomys. The small lingual anteroloph arm ends on the anterior wall of the protocone (3, Fig. 3c) and thus delimits a small protosinus. This junction may be also achieved somewhat lingually (1, Fig. 3d). The anterolophule is very short and connects to the antero-labial wall of the protocone. The posterior arm of the protocone and the protolophule II is directed slightly backwards or transverse. Similarly to the M1, both crests connect to the entoloph, forming a “T” or open Y-like structure. The long mesoloph is also directed backwards, and connects to the anterior wall of the metacone (three molars). One mesoloph is medium-sized. Finally, one molar has a short mesoloph. Metacone and hypocone are connected by a strong metalophule. The metacone is, with regard to the hypocone, much more transversally elongated and partially fused with the metalophule II. The metalophule is directed backwards and connects to the posterior arm of the hypocone. A short but well-defined posteroloph closes a small posterosinus. Metalophule II, posterior arm of the hypocone and posteroloph, fuses with wear. The sinus is closed by a strong lingual cingulum. Antero- and mesosinus are closed by narrow cinguli. Three roots.
M3: The front part of the M3 is similar to that of the M2. The posterior arm of the protocone and the protolophule II are directed slightly backwards in one case. The moderately worn M3 shows a trace of protolophule I. The heavily worn M3, in contrast, lacks the protolophules. The hypocone is reduced in comparison to the two anterior cusps, but still observable. In contrast, the metacone is fused with the strong metalophule. This crest runs parallel to the complete mesoloph. The posterior arm of the hypocone ends in a clear posteroloph. Three roots.
Mandible: The fragmentary hemimandible preserves the complete tooth row, but the incisor is broken close to the symphyseal area. The morphology of the enamel is indeed observable, the bone missing at the ventral part, below the diatesma. With 7.8 mm, the diatesma is a bit shorter than the molar row (8.95 mm). The mental foramen is rounded, placed in a labial position, very close to the cranial limit of the diastema, at around 2.3 mm from the anterior root of the m1. The mandibular corpus is conserved, although lingual-labially compressed, especially in the area underlying the m2-m3. The depth of the corpus under the anterior part of the m1 is only a bit less than under the anterior part of the m3 (respectively, 6.28 mm and 6.93 mm). The massetic ridges converge under the posterior root of the m1. At this point, they form a bump.
The right hemimandible in sediment with fragmentary m1 + m2-m3 (teeth in Fig. 2c) does not differ basically from the previous one. The bone is not lingual-labially compressed but broken anteriorly to the mental foramen. Only the anterior part of the ramus is preserved. The depth of the corpus under the anterior part of the m3 is 7.16 mm. The depth under the anterior part of the m1 is estimated at 6.65 mm.
Lower incisor: relatively massive (width 1.75 mm), it shows two ridges that run parallel at the labial-ventral margin.
m1: Two of the four m1 are almost useless for a detailed description: one is broken anteriorly to the mesolophid; the second is extremely worn. The m1s have a more elongated outline than the Cricetodon molars from the same site. The anteroconid is wider and the length of the anterior and central sinuses is clearly greater. The anteroconid is bilobed and relatively narrow on one tooth, the lingual cuspid being clearly smaller than its labial counterpart (2, Fig. 3f). On a second molar, both cuspids are similar in size. The lingual anterolophid arm is curved and ends at the anterior wall of the metaconid. The labial anterolophid arm is longitudinal. One molar has a high metalophulid I that connects to the anteroconid between its two lobes (1, Fig. 3f). The same tooth shows an interrupted anterolophulid which is directed to the centre of the anteroconid. A completely different arrangement of the crests is observed in the specimen in Fig. 3g. The anterolophulid connects to the labial lobe of the anteroconid, while the metalophulid I is medium-sized. Both crests run almost parallel. A narrow and transversal crest connects metaconid and protoconid. The short mesolophid is directed forwards. A very short ectomesolophid is observed. The hypolophulid is directed slightly forwards and connects to the anterior arm of the hypoconid. The posterolophid is strong and reaches the base of the anterior wall of the entoconid. A cingulid is found at the posterior part of the tooth, at the base of the hypoconid. Labial valleys are closed. Two roots.
m2: The teeth are almost rectangular, the postero-lingual border being rounded (Fig. 2c). The labial anterolophid descends to the basis of the labial wall of the protoconid. The lingual anterolophid is much shorter and connects to the anterior wall of the metaconid. The anterolophulid is longitudinal. In contrast, the metalophulid I runs almost parallel to the anterolophids and connects to the most anterior part of the anterolophulid. The mesolophid is relatively short, around one-third of the width of the mesosinusid. It runs transversally, or is directed forwards. A small enlargement in the thickness of the enamel of the ectolophid is interpreted as an extremely short ectomesolophid. The hypolophulid runs transversely and connects to the anterior arm of the hypoconid. The strong posterior arm of the hypoconid fuses with the posterolophid which ends free. It does not reach the lingual part of the molar, making the postero-lingual border of the m2 rounder. Like m1, a cingulid delimits a small valley at the back of the hypoconid.
m3: The tooth resembles the m2 in many points and differs primarily by its outline (Fig. 2c). Morphologically, and beside the absence of a cingulid at the posterior part of the hypoconid, the main difference observed is in the course of the mesolophid: it does not connect to the longitudinal crest as in the m2 but directly to the protoconid. It is thus clearly longer but directed backwards. One molar has a long mesolophid which runs close to the entoconid. A second tooth has a shorter crest. The two other m3 are either damaged or heavily worn. The very short ectomesolophid is better developed than in m2.
Burg-Balzhausen
Fossil material and measurements (after Seehuber 2009): 1 M1, 1 fragmentary m2 (NMA 2005-716/1927; NMA 2005-772/1927). M1: 3.63 × 2.38; m2: width = 2.30.
The fossils were originally presented by Seehuber (2008, 2009). We propose here a more extensive description:
M1: The broad anterocone is deeply furrowed anteriorly, and both cusps are approximately the same size (Fig. 2d). At their base, a small cingulum is formed on the anterior flank of the anterocone. The lingual anteroloph arm is short and ends in a distinct style at the lingual tooth margin. The labial anteroloph arm is short, weak, and also ends in a style. The anteromesoloph forms a strong style-like enamel cusp. The short protolophule (II) also ends in a similar hump. The very short posterior arm of the protocone is separated from this hump by a not very deep caesura (which would not have been recognisable with worn process). The anterior arm of the hypocone is strongly developed and continues, clearly narrowed, in the mesoloph. This half-length enamel ridge ends at the anterior metacone flank. The metalophule II is easily recognisable and flows into the posterior arm of the hypocone. At the point of contact, a weak but easily visible posteroloph branches off posteriorly and ends at the metacone flank. Both the lingual and labial sides of the tooth are enclosed by a clearly formed cingulum. Starting from the anterior hypocone flank, a weak enamel ridge runs into the straight lingual main sinus. Basal enamel areas of the anterocone and metacone are etched (by gastric acid?). The tooth roots are no longer present.
m2: The labial anterolophid arm ends at the base of the protoconid (Fig. 2b). The anterolophulid is well-developed and ends at the anteroconid. The lingual anterolophid arm is strongly shortened and weak. Medially, along the longitudinal axis of the tooth, a strong enamel thickening is developed, into which the protoconid posterior arm the hypoconid anterior arm and a metalophulid II merge. The metalophulid II is directed perpendicular to the longitudinal axis. The mesolophid is half-length and ends freely between ento- and metaconid. Starting from the aforementioned central enamel thickening, a medium-sized ectomesolophid extends into the broad lingual main sinus. The hypoconid posterior arm ends deep at the metaconid base. The posterior tooth area is broken off along the hypoconid and the hypoconid posterior arm. The m2 also shows areas with enamel etching. The roots of the tooth are missing.
General remarks
A left Lartetomys mandible with i+m2m3 from Petersbuch 41 “rechte Spalte” was described by Rummel (1997). Furthermore, Boon (1991: Plate VII, fig. 3) assigned an M3 from Ziemetshausen to Cricetodon aff. meini. In fact, this molar is clearly too large (2.35 × 2.05, estimated after table 16 in Boon, 1991) and corresponds to Lartetomys. A worn M3 from Laimering 3 differs from the corresponding Cricetodon teeth by its larger size (2.24 × 2.03) as well as the non-reduction of its posterior part and belongs also to Lartetomys. These finds fit morphometrically with the fossil material from Höll and Burg-Balzhausen. The posterior part of the M1 of Burg-Balzhausen is original, but this morphology is interpreted as abnormal and therefore not representative. From a metrical point of view, the M1 of L. mirabilis from Vieux-Collonges is slightly larger than those studied here, but does not reach the size of L. dehmi. The m1 belongs to the variation range observed in Höll. The morphology of the French lower molar is not different from the corresponding German teeth. The M1, holotype of L. mirabilis, shows a transverse connection of the metalophule to the hypocone. The metalophule is directed backwards in Mixocricetodon dehmi (Rummel 1997), just like in the great majority of molars from Germany. However, one M1 from Höll shows a connection similar to the one observed at Vieux-Collonges. It is therefore appropriate to assign all these populations to a single species: L. mirabilis.
The morphology of Lartetomys mirabilis teeth is very similar to that of Mixocricetodon dehmi. However, there are some differences, one of which is the presence of a prelophid in the m1, relatively common in Mixocricetodon dehmi, absent in L. mirabilis. With the clear difference in size, these factors are not sufficient to separate the species at the genus level. We therefore confirm here the synonymy of the genera proposed by Mein (2003).
A damaged m2 from Göttschlag is in addition tentatively assigned to Mixocricetodon dehmi.
Sen and Erbajeva (2011) stated that Mixocricetodon dehmi would be better assigned to Cricetodon: the authors outlined that Rummel (1997) discriminates Mixocricetodon from Cricetodon mainly based on the presence of three roots and strong mesolophs on the upper molars and thus considered it insufficient for considering Mixocricetodon as a valid genus. We cannot agree herein with this interpretation. The differences with Cricetodon as provided by Rummel (1997: 88) are: the size; the much more massive cusps in Mixocricetodon; the number of roots in M1 and M2; the presence of a hypolophulid II in Mixocricetodon; the more common development of ectolophs on the upper molars of Cricetodon; the divided/fissured anteroconid (m1) and the lack of an anterior arm of the paracone, as well as the stronger posteroloph in Mixocricetodon; the most often lacking, or reduced mesoconid on the lower Cricetodon molars; and finally, the different structure of the labial main sinusid.
In this way, a new Lartetomys mirabilis-L. dehmi lineage is defined. This lineage is characterised by:
-Increase in size of the molars
-Increase in enamel thickness
-Elongation of the anteromesoloph (M1)
-Development of a prelophid in the m1
-Increase in the size of the ectomesolohid in the m1
-Development of a short hypolophulid II (absent in L. mirabilis)
-Disappearance of the transverse crest that connects metaconid and protoconid in the m1
Discussion
The origin and relationship of Lartetomys is discussed below. We propose two hypotheses:
Eastern Mediterranean hypothesis
Lartetomys migrated to South Germany during the OSME-EBE. This new occurrence is accompanied by that of Cricetodon meini and Karydomys. It can therefore be assumed that these three taxa have a common geographic origin. C. meini is recognised in the Eastern Mediterranean (Koufos 2006). It is generally the only Cricetodontini found in the fauna where it is present. However, Koufos (2006) also recognises Turkomys sp. at Thymiana A (~ 15.5 Ma) but without further precision. Greece is also rich in Karydomys (Theocharopoulos 2000, López-Antoñanzas et al. 2019). Cricetodon meini and K. strati are found together only in Chios (pers. obs. JP), and the origin of Lartetomys cannot be found there. Karydomys is also a member of the Anatolian fauna (Theocharopoulos 2000). Furthermore, the early and middle Miocene Cricetodontini from Turkey have been the subject of numerous studies (e.g. Sen and Ünay 1978, 1979, Ünay 1980, 1990, De Bruijn et al. 1993, 2013, Rummel 1998, Çinar 2011, Çinar Durgut and Ünay 2016). Among the Cricetodon species present just before or at the time of the appearance of Lartetomys in Central Europe, no species matches the morphologic characteristics, thus enabling us to recognise the origin of the genus in the area. In fact, the Turkish species C. tobieni, C. fikreti, C. trallesensis, and Cricetodon n. sp.2 from Horlak 2 (sensu De Bruijn et al. 1993) are very different. Similarly, C. aliveriensis is not relevant. Only C. yapintiensis could be discussed, but the specimens as figured by Çinar (2011) and Çinar Durgut and Ünay (2016) do not allow further analysis. Notably, Karydomys is recorded in Yapinti (Ünay et al. 2001).
Asian hypothesis
Sen and Erbajeva (2011) include Mixocricetodon (= Lartetomys dehmi) in a cladistic analysis. Their results concluded that a close relationship among some Cricetodon (the European C. jotae and C. sansaniensis, as well as the Asian C. volkeri), the genera Gobiocricetodon and Plesiodipus, as well as Tsaganocricetus irtyschensis exists. While it is suggested here that a close relationship to the European Cricetodon, Tsaganocricetus, and Plesiodipus is unlikely, the other forms deserve attention.
Cricetodon volkeri (China, middle Tunggurian, middle Miocene; Wu et al. 2009) was based on very few and partially damaged isolated teeth, making any comparison very difficult. C. fengi is close in age to C. volkeri (China, middle Tunggurian, middle Miocene; Qiu and Li 2016). This species is smaller than Lartetomys mirabilis, but both taxa share some morphologic characteristics.
Cricetodon sonidensis (China, upper Aoerban Formation, lower Miocene, Qiu and Li 2016) is older, close in age to L. mirabilis, and is considered to be the ancestor of C. fengi. Many characteristics recall Lartetomys, such as the strong anterior arm of the hypocone, the mesolophs which are near the metacone, the unreduced M3, and the general structure of the lower molars. Apart from a much smaller size, the difference is that the anteroconid is not divided, and the M1 anterocone is less fissured than in the European genus. The rare remains of Cricetodon sp. from locality XJ 200114 (China, MN3, Maridet et al. 2011) also share characteristics with Lartetomys. In particular, the two figured m3 show the long and backward-directed mesolophid. Similarly, the M2 resembles Lartetomys by the strong anterior arm of the hypocone, which gives rise to the long mesoloph, and the arrangement of the posterior arm of the protocone and the protolophule II. But it differs by the presence of a well-developed posterosinus. In contrast, C. wanhei from the lower Miocene of China (Qiu 2010) shares few key morphological characteristics with Lartetomys.
As mentioned above, Sen and Erbajeva (2011) find a relationship to the Asian genus Gobicricetodon. In that work, they describe G. fillipovi from the locality Aya Cave (Baikal Lake). The fauna interpreted as MN 6 thus probably contains one of the oldest Gobicricetodon. The figured type specimen of G. fillipovi, a jaw with M1-M3, is striking by its similarity to Lartetomys. Some morphological aspects, such as the general shape of the teeth, the division of the M1 anterocone, and the structure of the posterior part of the M2 and the M3 are found in the European genus. The specimens drawn in their fig. 3 do not prevent us from suspecting a phylogenetic relationship with Gobicricetodon, the only clear difference with Lartetomys being the single-cusped anteroconid of the m1. Unfortunately, no m3 is known from the Aya Cave. In contrast, Maridet et al. (2014) note the presence of a few teeth of Gobicricetodon sp. in the late Early Miocene from Central Mongolia. While their m3 lacks the mesolophid, a characteristic crest of Lartetomys, this crest might be present and directed backwards in certain Gobiocricetodon species.
Conclusions
It seems therefore that an Asian origin of Lartetomys (=Mixocricetodon) is plausible. Karydomys, which appears in Europe almost contemporaneously, is also known from the lower Miocene of Kazakstan (Kordikova and Bruijn, 2001) and China (Maridet et al. 2011). In contrast, Cricetodon meini is not known further east than Georgia (C. aff. meini from Belometchetskaya, Pickford et al. 2000). It is therefore at present not possible to determine whether these three taxa found in Germany have their origin in a common geographical area, or whether they originate separately.
Lartetomys mirabilis migrated to central Europe and France about 15 Ma ago, at the beginning of the Middle Miocene Climate Transition. It is a rare component of the faunas; only the Bavarian locality Höll provided a representative sample of the species. It then increased rapidly in size, underwent morphological changes for about 1 million years, and disappeared from the fossil record with L. dehmi. Lartetomys may have continued to evolve in an insular environment with the endemic Cricetidae Mystemys from Gargano (South Italy, Masini et al. 2013). This species is highly derived, and unfortunately, no definitive conclusion can be drawn (Savorelli and Masini 2016).
References
Aguilar, J.-P. (1995). Evolution de la lignée Megacricetodon collongensis- Megacricetodon roussillonensis (Cricetidae, Rodentia, Mammalia) au cours du Miocène inférieur et Moyen dans le Sud de la France. Palaeovertebrata, 24, 1–45.
Augé, M., Ginsburg, L., Lapparent de Broin, F.d., Makinsky, M., Mourer, C., Pouit, D., & Sen, S. (2002). Les vertébrés du Miocène moyen de Contres (Loir-et-Cher, France). Revue Paléobiologie, 21, 819–852.
Baudelot, S., & Collier, A. (1978). Les faunes miocènes du Haut Armagnac (Gers, France) 1. Les gisements. Bulletin de la Société géologique de France, 114, 194–206.
Berger, G. (2010). Die miozäne Flora und Fauna (MN 5) der historischen Fossil-Lagerstätte Georgensgmünd (Mfr.) unter Berücksichtung der Ablagerungen des Urmaintals zwischen Roth und Treuchtlingen. Naturhistorische Gesellschaft Nürnberg e.V. Abhandlungen, 46, 1–191.
Bolliger, T. (1994). Die Obere Süßwassermolasse in Bayern und der Ostschweiz: bio-und lithostratigraphische Korrelationen. Mitteilungen der bayerischen Staatsammlung für Paläontologie und historische Geologie, 34, 109–144.
Bolliger, T. (1997). The current knowledge of the biozonation with small mammals in the Upper Freshwater Molasse in Switzerland, especially the Hörnli-Fan. Mémoires et Travaux de l’E.P.H.E., institut de Montpellier, 21, 501–513.
Bolliger, T. (2000). Wiesholz (canton of Schaffhausen, Switzerland), A pecular mammal fauna from mica-rich sands (Upper Freshwater Molasse, Miocene, early MN6). Revue de Paléobiologie, 19, 1–18.
Boon, E. (1991). Die Cricetiden und Sciuriden der oberen Süßwassermolasse von bayerisch-Schwaben und ihre stratigraphische Bedeutung. Dissertation zur Erlangung des Doktorgrades an der Falkultät für Geowissenschaften der ludwig-Maximilians-Universität München. 158 p.
Bruijn, H. de, Daams, R., Daxner-Höck, G., Fahlbusch, F., Ginsburg, L., Mein, P., & Morales, J. (1992). Report of the RCMNS working group on fossil mammals, Reisenburg 1990. Newsletters on Stratigraphy, 36, 65–118.
Bruijn, H. de, Fahlbusch, V., Saraç, G., & Ünay, E. (1993). Early Miocene rodent faunas from the eastern mediterranean area Part III. The genera Deperetomys and Cricetodon with a discussion of the evolutionary history of the Cricetodontini. Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen, 96, 151–216.
Bruijn, H. de, Ünay, E., & Hordijk, K. (2013). Chapter 26: A review of the Neogene succession of the Muridae and Dipodidae from Anatolia, with special reference to taxa known from Asia and/or Europe. In Wang, X., Flynn, L. and Fortelius, M. (Eds.), Fossil mammals of Asia, Columbia, pp. 566–582.
Bulot, C., Ginsburg, L., & Tassy, P. (1992). Le gisement à mammifères miocènes de Castelnau-d'Arbieu (Gers). Données nouvelles et implications biostratigraphiques. Comptes rendus de l'Académie des Sciences de Paris, sciences de la terre et des planètes, 314(Serie II), 533–537.
Çinar, N. (2011). The cricetodontini from the early miocene of the eastern mediterranean, Master thesis (pp.1-199). Ankara: Department of Geological Engineering, Ankara University.
Çinar Durgut, N., & Ünay, E. (2016). Cricetodontini from the Early Miocene of Anatolia. Bulletin of the Mineral research and exploration, 152, 85–119.
Daxner-Höck, G. (2003). Cricetodon meini and other rodents from Mühlbach and Grund, Lower Austria (Middle Miocene, late MN5). Annalen des Naturhistorischen Museums in Wien, 104(A), 267–291.
Freudenthal, M. (1985). Cricetidae (Rodentia) from the Neogene of Gargano (Prov. of Foggia, Italy). Scripta Geologica, 77, 29–76.
Freudenthal, M., Hugueney, M., & Moissonet, E. (1994). The genus Pseudocricetodon (Cricetidae, Mammalia) in the Upper Oligocene of the province of Teruel (Spain). Scripta Geologica, 104, 57–114.
Ginsburg, L., & Bulot, C. (2000). Le cadre stratigraphique du site de Sansan. Bulletin du Museum National d'Histoire naturelle Paris, 83, 39–67.
Garapich, A., & Kälin, D. (1999). New findings on the rare and peculiar genus Lartetomys (Cricetidae, Rodentia, Mammalia). Eclogae Geologicae Helvetiae, 92, 495–502.
Heissig, K. (1989). Neue Ergebnisse zur Stratigraphie der mittleren Serie der oberen Süßwassermolasse Bayerns. Geologica Bavarica, 94, 239–257.
Heissig, K. (1995). Die Entwicklung der großen Democricetodon-Arten und die Gattung Collimys (Cricetidae, Mamm.) im späten Mittelmiozän. Mitteilungen der bayerischen Staatsammlung für Paläontologie und historische Geologie, 35, 87–108.
Heissig, K. (1997). Mammal faunas intermediate between the reference faunas of MN4 and MN6 from the Upper freshwater Molasse of Bavaria. Actes du Congrès BiochroM’97. Mémoires et Travaux de l’Ecole pratique des Hautes Etudes, Institut de Montpellier, 21, 537–546.
Heissig, K. (2006). Biostratigraphy of the "main bentonite horizon" of the Upper Freshwater Molasse in Bavaria. Palaeontographica, A, 277, 93–102.
Hershkovitz, P. (1967). Dynamics of Rodent Molar Evolution: A Study Based on New World Cricetinae, Family Muridae. Journal of Dental Research, 46, 829–842.
Hír, J. (2013). Early and Middle Miocene (MN5-MN6) transitional rodent fauna from Litke (North Hungary, Nógrád County). Fragmenta Palaeontologica Hungarica, 30, 101–137.
Hír, J., & Venczel, M. (2018). A preliminary report on the first results of the reexcavation of the middle Miocene palaeovertebrate locality Szentendre, Cseresznyés-árok (Hungary, Pest County). Nymphaea, Folia naturae Bihariae, 45, 35–80.
Kälin, D., & Kempf, O. (2009). High-resolution stratigraphy from the continental record of the Middle Miocene northern Alpine Foreland Basin of Switzerland. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, 254, 177–235.
Kordikova, E. G., & Bruijn, d. H. (2001). Early Miocene Rodents from the Aktau Mountains (South-Eastern Kazakhstan). Senckenbergiana lethaea, 81, 391–405.
Koufos, G. D. (2006). The Neogene mammal localities of Greece: Faunas, chronology and biostratigraphy. Hellenic Journal of Geosciences, 41, 183–214.
López-Antoñanzas, R., Peláez-Campomanes, P., Prieto, J., Knoll, F., & Hautier, L. (2019). New species of Karydomys (Rodentia) from the Miocene of Chios Island (Greece) and phylogenetic relationships of this rare democricetodontine genus. Papers in Palaeontology, 5, 33–45.
López Guerrero, P., Álvarez Sierra, M. A., García-Paredes, I., & Peláez-Campomanes, P. (2014). New Cricetodontini from the middle Miocene of Europe: an example of mosaic evolution. Bulletin of Geosciences, 89(3), 573–592.
López-Guerrero, P., García-Paredes, I., Prieto, J., López-Antoñanzas, R. & Álvarez-Sierra, M. A. (2015). Palaeodiversity of Cricetodontini during the late Aragonian (middle Miocene) from the European basins. In L. W. van den Hoek Ostende, P. Peláez-Campomanes, & W. Wessels (Eds.) Old worlds, new ideas. A tribute to Albert van der Meulen. Palaeobiodiversity and Palaeoenvironments, 95(3), 415–430.
Maridet, O., (2003). Révision du genre Democricetodon (Mammalia, Rodentia, Cricetinae) et dynamique des faunes de rongeurs du Néogène d’Europe occidentale: évolution, paléodiversité et paléogéographie. Thèse de doctorat (pp. 1-252). Lyon: Université Claude-Bernard Lyon 1.
Maridet, O., Berthet, D., & Mein, P. (2000). Un nouveau gisement karstique polyphasé miocène moyen de Four (Isère): étude des Cricétidés (Mammalia, Rodentia). Géologie de la France, 2, 71–79.
Maridet, O., Wu, W. Y., Ye, J., Bi, S.-D., Ni, X.-J., & Meng, J. (2011). Early Miocene cricetids (Rodentia) from the Junggar basin (Xinjiang, China) and their biochronological implications. Geobios, 44, 445–459.
Maridet, O., Daxner-Höck, G., Badamgarav, D., & Göhlich, U. B. (2014). Cricetidae (Rodentia, Mammalia) from the Valley of Lakes (Central Mongolia): focus on the Miocene record. Annalen des Naturhistorischen Museums in Wien, A, 116, 247–269.
Marković, Z., Bruijn, d. H., & Wessels, W. (2016). A revision of the new rodent collections from the Early Miocene of Sibnica, Serbia. In Z. Marković & M. Milivojević (Eds.), Life on the shore – geological and paleontological research in the Neogene of Sibnica and vicinity (Levač basin, Central Serbia). Part 1 (pp. 63–117). Belgrade: Special Issue of the Natural History Museum in Belgrade.
Masini, F., Rinaldi, P.M., Savorelli, A. & Pavia,M. (2013). A new small mammal assemblage from theM013 fissure filling (Gargano, South-Eastern Italy). Geobios, 46, 49-61.
Mein, P. (1958). Les mammifères de la faune sidérolithique de Vieux-Collonges. Nouvelles Archives du Muséum d'Histoire naturelle Lyon, 5, 122.
Mein, P. (2003). On Neogene rodents of Eurasia: distribution and migrations. In J. W. F. Reumer & W. Wessels (Eds.), Distribution and migration of Tertiary mammals in Eurasia. A volume in honour of Hans de Bruijn (Vol. 10, pp. 407–418). Rotterdam: Deinsea.
Mein, P., & Freudenthal, M. (1971a). Une nouvelle classification des Cricetidae (Mammalia, Rodentia) du Tertiaire de l'Europe. Scripta Geologica, 2, 1–37.
Mein, P., & Freudenthal, M. (1971b). Les Cricetidae (Mammalia, Rodentia) du Néogène Moyen de vieux-Collonges Partie 1: Le genre Cricetodon Lartet, 1851. Scripta Geologica, 5, 49.
Mein, P., & Freudenthal, M. (1981). Les Cricetidae (Mammalia, Rodentia) du Néogène Moyen de Vieux-Collonges Partie 2 Cricetodontinae incertae sedis, Melissiodontinae, Platacanthomyinae, et Anomalomyidae. Scripta Geologica, 60, 11.
Meulen, A. J. van der, Garciá-Peredes, I., Álvarez-Sierra, M. A., Van den Hoek Ostende, L. W., Hordijk, K., Oliver, A., López-Guerrero, P., Hernández-Ballarín, V., & Peláez-Campomanes, P. (2011). Biostratigraphy or biochronology? Lessons from the Early and Middle Miocene small Mammal Events in Europe. Geobios, 44, 309–321.
Oliver, A., & Peláez-Campomanes, P. (2013). Megacricetodon vandermeuleni, sp. nov. (Rodentia, Mammalia), from the Spanish Miocene: a new evolutionary framework for Megacricetodon. Journal of Vertebrate Palaeontology, 33, 943–955.
Oliver Pérez, A. (2015). Evolution of Megacricetodon from the Aragonian and Vallesian (middle and upper Miocene) of the Iberian Peninsula. Memoria de tesis doctoral, Faculty of geological sciences, Universidad Complutense de Madrid. 298 p.
Pickford, M. (2017). Revision of "peccary-like" Suoidea (Artiodactyla: Mammalia) from the Neogene of the Old World. Münchner Geowissenschaftliche Abhandlungen, A, 46, 1–144.
Pickford, M., Gabuna, L., Mein, P., Morales, J., & Azanza, B. (2000). The Middle Miocene mammalian site of Belometchetskaya, North Caucasus: an important biostratigraphic link between Europe and China. Geobios, 33, 257–267.
Prieto, J. (2007). Kleinsäuger-Biostratigraphie und Paläoökologie des höheren Mittelmiozäns (MN 8) Bayerns: Spaltenfüllungen der Fränkischen Alb und Lokalitäten der Oberen Süßwassermolasse im Vergleich (pp. 1–213). Dissertation zur Erlangung des Doktorgrades an der Falkultät für Geowissenschaften der Ludwig-Maximilians-Universität München.
Prieto, J. (2012). Comments on the morphologic and metric variability in the cricetid rodent Deperetomys hagni (Fahlbusch, 1964) from the Middle Miocene of South Germany. Zitteliana, A, 52, 71–77.
Prieto, J., & Rummel, M. (2009). Erinaceidae (Mammalia, Erinaceomorpha) from the Middle Miocene fissure filling Petersbuch 68 (southern Germany). Zitteliana, A, 48(49), 103–111.
Prieto, J., & Scholz, H. (2013). First record of Karydomys (Rodentia, Mammalia) from the German part of the North Alpine Foreland Basin. Swiss Journal of Geosciences, 106, 303–307.
Prieto, J., & Rummel, M. (2016). Some considerations about the small-mammal evolution in South Germany, with emphasis on late Burdigalian-earliest Tortonian (Miocene) cricetid rodents. Comptes Rendus Palevol, 15, 837–854.
Prieto, J., Hassler, A., Prinz, G., Gross, M., & Böhme, M. (2016). Die miozänen Kleinsäuger von Schönweg-“Brüchl” (Kärnten). Carinthia II, 126, 503–524.
Prieto, J., Peláez-Campomanes, P., & Scholz, H. (2017). Sciurids (Rodentia, Mammalia) from Höll (Middle Miocene, Bavaria). Zitteliana, A, 89, 351–354.
Prieto, J., Becker, D., Rauber, G., & Pirkenseer, C. M. (2018). New biostratigraphical data for the Burdigalian Montchaibeux Member at the locality Courrendlin-Solé (Canton of Jura, Switzerland). Swiss Journal of Geosciences, 111, 1–11.
Qiu, Z.-D. (2010). Cricetid rodents from the Early Miocene Xiacaowan Formation, Sihong, Jiangsu. Vertebrata PalAsiatica, 48, 7–47.
Qiu, Z., & Li, Q. (2016). Neogene Rodents from Central Nei Mongol, China. Palaeontologia Sinica, C, 198(30), 684.
Reichenbacher, B., Krijgsman, W., Lataster, Y., Pippèr, M., Baak, C. G. C.v., Chang, L., Kälin, D., Jost, J., Doppler, G., Jung, D., Prieto, J., Abdul Aziz, H., Böhme, M., Garnish, J., Kirscher, U., & Bachtadse, V. (2013). A new magnetostratigraphic framework for the Lower Miocene (Burdigalian/Ottnangian, Karpatian) in the North Alpine Foreland Basin. Swiss Journal of Geosciences, 106, 309–334.
Rocholl, A., Schaltegger, U., Gilg, H. A., Wijbrans, J., & Böhme, M. (2017). The age of volcanic tuffs from the Upper Freshwater Molasse (North Alpine Foreland Basin) and their possible use for tephrostratigraphic correlations across Europe for the Middle Miocene. International Journal of Earth Sciences, 107, 387–407.
Rummel, M. (1997). Mixocricetodon dehmi n. gen., n. sp., ein neuer Cricetide (Rodentia, Mammalia) aus dem Mittelmiozän von Süddeutschland. Mitteilungen der bayerischen Staatsammlung für Paläontologie und historische Geologie, 37, 75–93.
Rummel, M. (1998). Die Cricetiden aus dem Mittel- und Obermiozän der Türkey. Documenta naturae, 123, 1–300.
Rummel, M. (1999). Tribe Cricetodontini. In G. Rössner & K. Heissig (Eds.), The Miocene land mammals of Europe (pp. 359–362). Munich: Verlag Dr. Friedrich Pfeil.
Rummel, M. (2000). Die Cricetodontini aus dem Miozän von Petersbuch bei Eichstätt. Die Gattung Cricetodon. Senckenbergiana lethaea, 80, 149–171.
Rummel, M., & Kälin, D. (2003). Die Gattung Cricetodon (Mammalia, Rodentia) aus dem Mittelmiozän der Schweizer Molasse. Zitteliana, A, 43, 123–141.
Savorelli, A., & Masini, F. (2016). Mystemys giganteus n. gen. et sp.: an enigmatic and rare cricetid from the Terre Rosse M013 fissure filling (Gargano, Southeastern Italy). Palaeontographica Abteilung, A, 306, 1–23.
Scholz, H. (1986a). Ein mittelmiozäne Wirbeltierfundstätte: Höll bei Gestratz. Natur und Museum, 116, 65–80.
Scholz, H. (1986b). Beiträge zur Sedimentologie und Paläontologie der Oberen Süßwassermolasse im Allgäu. Jahrbuch der geologischen Bundesanstalt, A, 129, 99–127.
Seehuber, U. (2008). Litho- und biostratigraphische Untersuchungen in der Oberen Süßwassermolasse in der Umgebung von Kirschheim in Schwaben (pp. 1-276). Dissertation zur Erlangung des Doktorgrades an der Falkultät für Geowissenschaften der Ludwig-Maximilians-Universität München.
Seehuber, U. (2009). Litho- und biostratigraphische Untersuchungen in der Oberen Süßwassermolasse in der Umgebung von Kirschheim in Schwaben. Documenta naturae, 175, 1–355.
Sen, S., & Ünay, E. (1978). Cricetodontini (Rodentia, Mammalia) miocènes de Turquie. Évolution et biostratigraphie. Bulletin de la Société géologique de France, 20, 837–840.
Sen, S., & Ünay, E. (1979). Sur quelques Cricetodontini (Rodentia) du Miocène moyen d'Anatolie. Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen, 82, 293–301.
Sen, S., & Erbajeva, M. A. (2011). A new species of Gobicricetodon Qiu, 1996 (Mammalia, Rodentia, Cricetidae) from the Middle Miocene Aya Cave, Lake Baikal. Vertebrata PalAsiatica, 49, 257–274.
Theocharopoulos, C. D. (2000). Late Oligocene-Middle Miocene Democricetodon, Spanocricetodon and Karydomys n. gen. from the eastern mediterranean area. Gaia, 8, 1–92.
Ünay, E. (1980). The Cricetodontini (Rodentia) from the Bayraktepe section (Çanakkale, Turkey). Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen, 83, 399–418.
Ünay, E. (1990). Turkomys pasalarensis Tobien, its range of variation in the type locality at Paşalar, Turkey. Journal of Vertebrate Palaeontology, 19, 437–443.
Ünay, E., Atabey, E., & Saraç, G. (2001). Small mammals and foraminifera from the Anatolian (Central Taurus) Early Miocene. Annals of the Carnegie Museum, 70(4), 247–256.
Wu, W., Meng, J., Ye, J., Ni, X.-J., Bi, S.-D., & Wei, Y.-P. (2009). The Miocene mammals from Dingshanyanchi Formation of North Junggar Basin, Xinjiang. Vertebrata PalAsiatica, 47, 208–233.
Acknowledgements
A large part of the fossils were excavated by Udo Scholz. We warmly thank Lawrence Flynn and one anonymous reviewer for their constructive comments. We thank Sinje Weber for her help in editing the article.
Funding
Open Access funding enabled and organized by Projekt DEAL.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Prieto, J., Rummel, M., Scholz, H. et al. A new middle Miocene lineage based on taxonomic revision of the large and rare cricetid-rodent genus Lartetomys. Palaeobio Palaeoenv 102, 223–236 (2022). https://doi.org/10.1007/s12549-021-00485-0
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12549-021-00485-0