
Design and Operation of Empirical Manganese-Removing Bioreactors and Integration into a Composite Modular System for Remediating and Recovering Metals from Acidic Mine Waters


Abstract
:1. Introduction
2. Materials and Methods
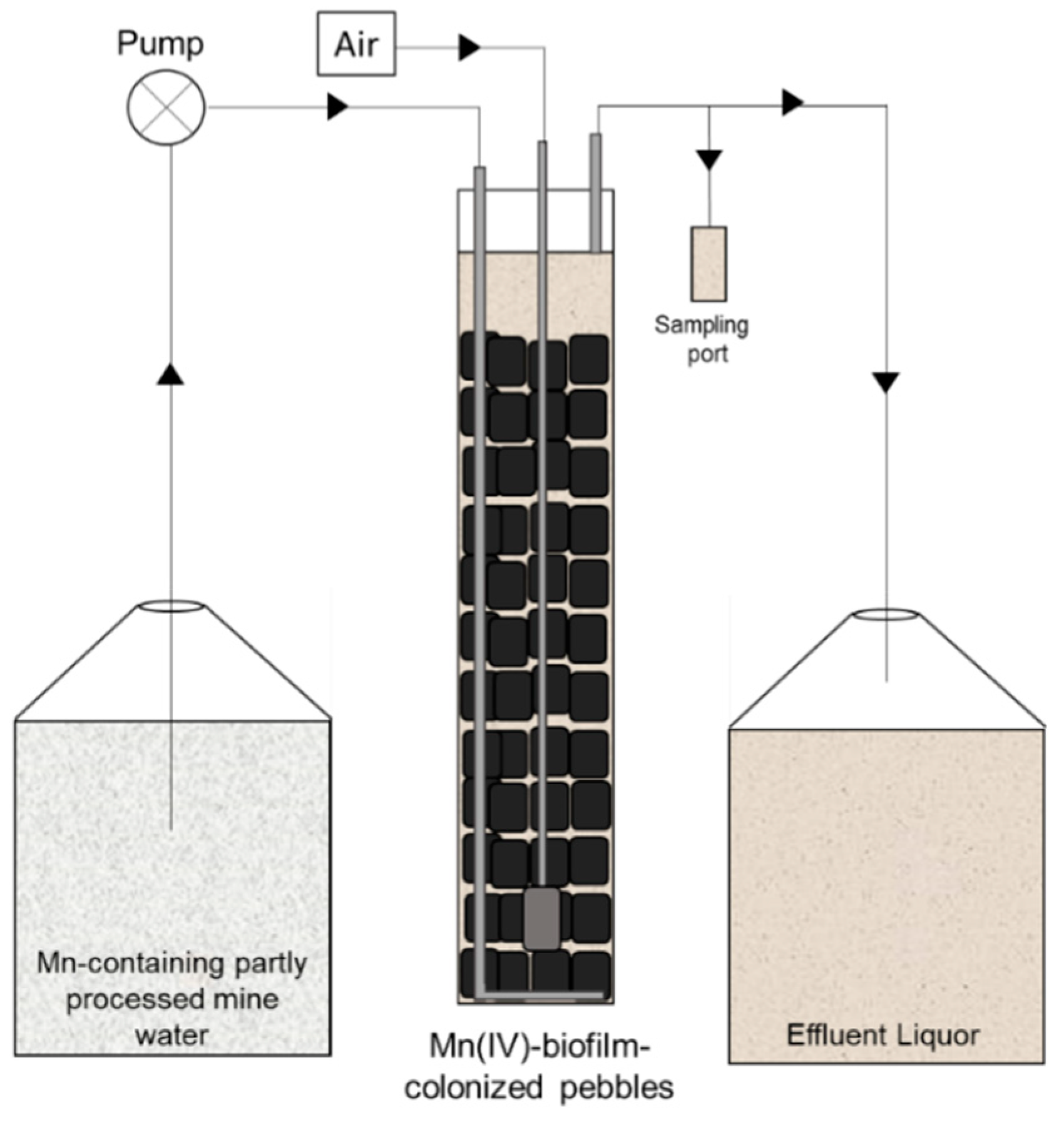
2.1. Commissioning and Operation of Mn(II)-Oxidizing Bioreactors (MnOBs)
Variations in Operational and Set Up Conditions of MnOBs
2.2. Biomolecular Analysis of the Microbial Biofilm Communities
2.3. Identification and Characterization of a Putative Mn (II)-Oxidizing Fungus
2.3.1. Biomolecular Analysis of Isolate MnI1
2.3.2. Physiological Characteristics of Isolate MnI1
Mn (II) Oxidation
Effects of pH and Temperature on Growth of Isolate MnI1
Growth on Various Organic Substrates
2.4. Miscellaneous Analytical Techniques
3. Results
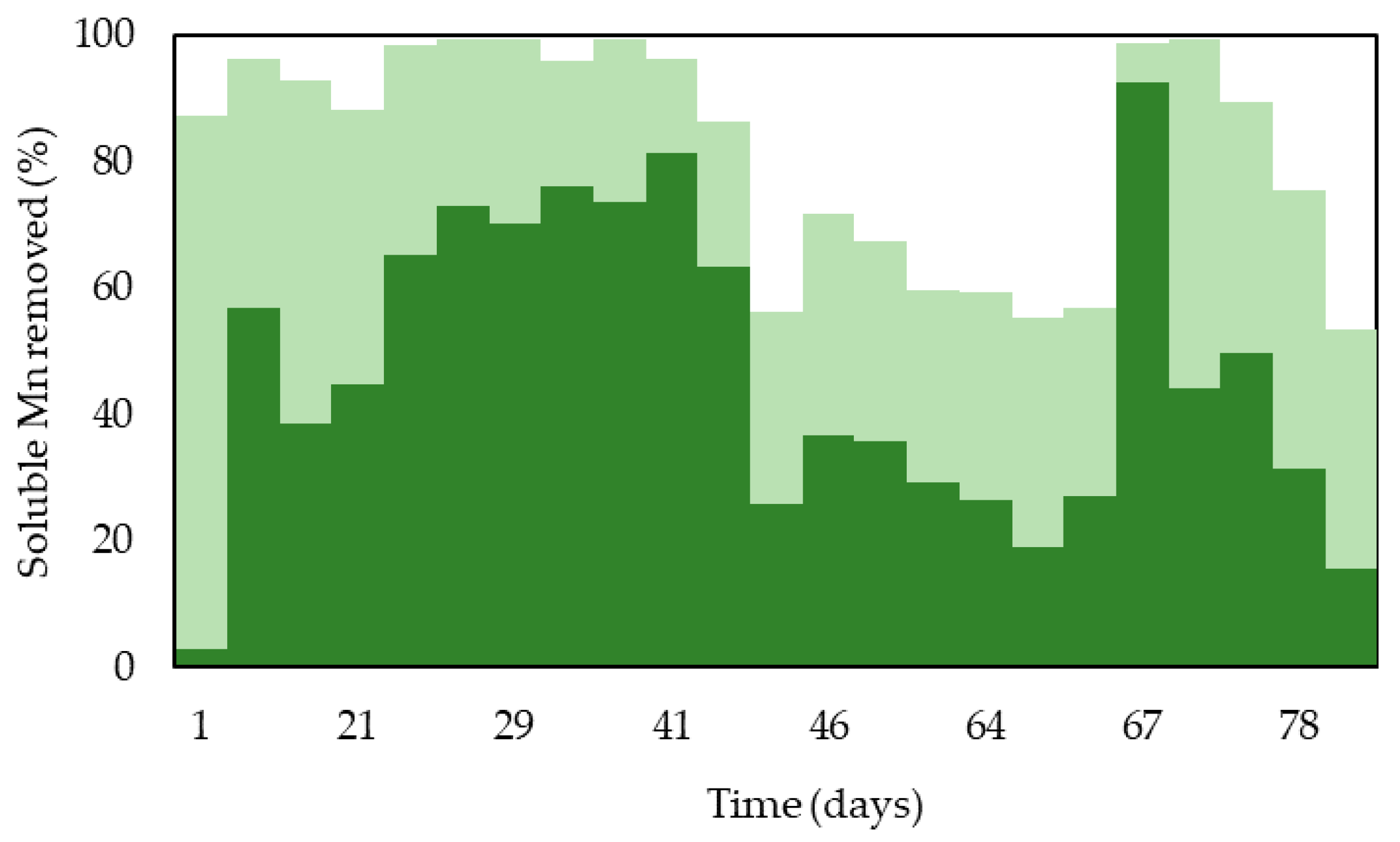
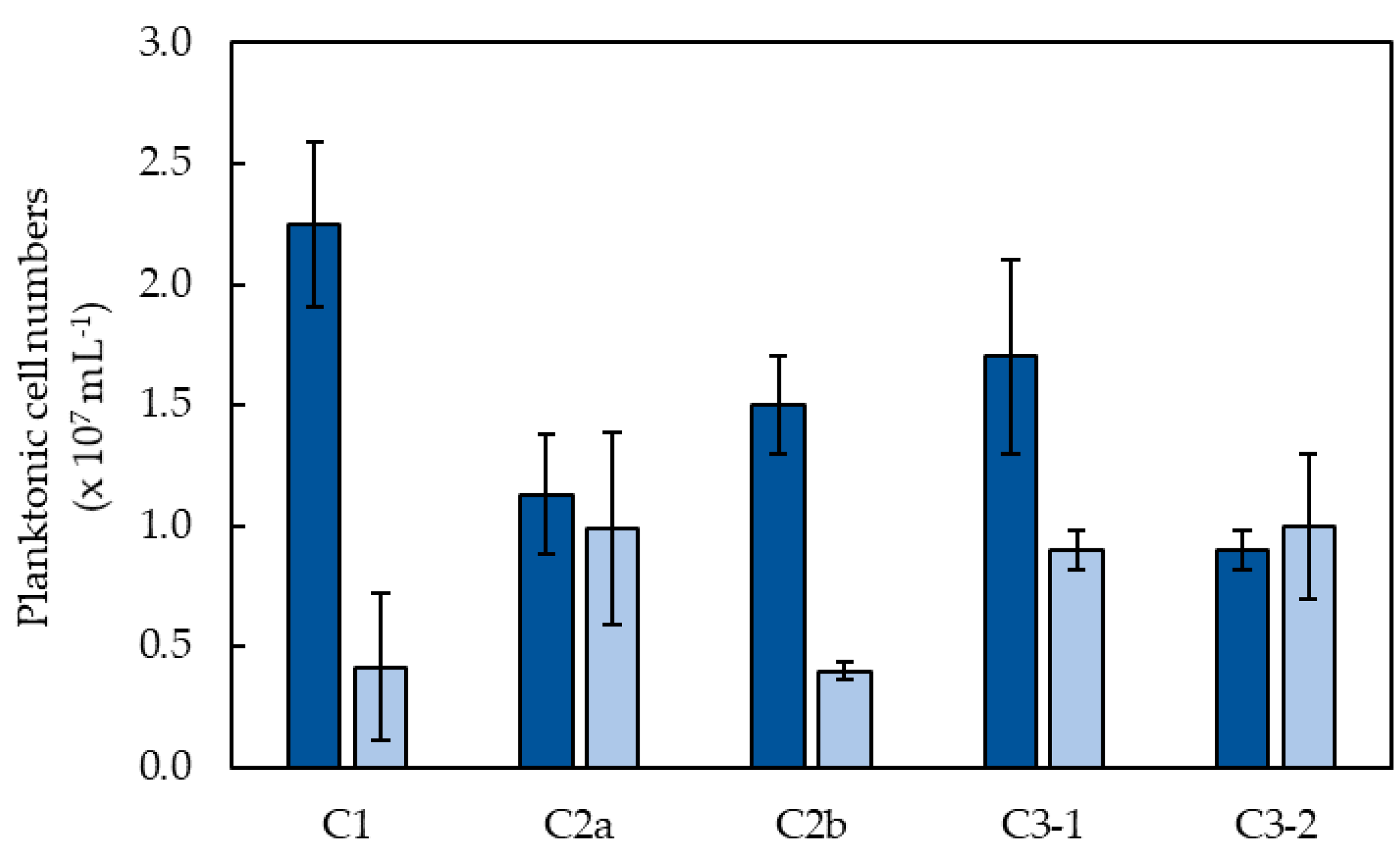
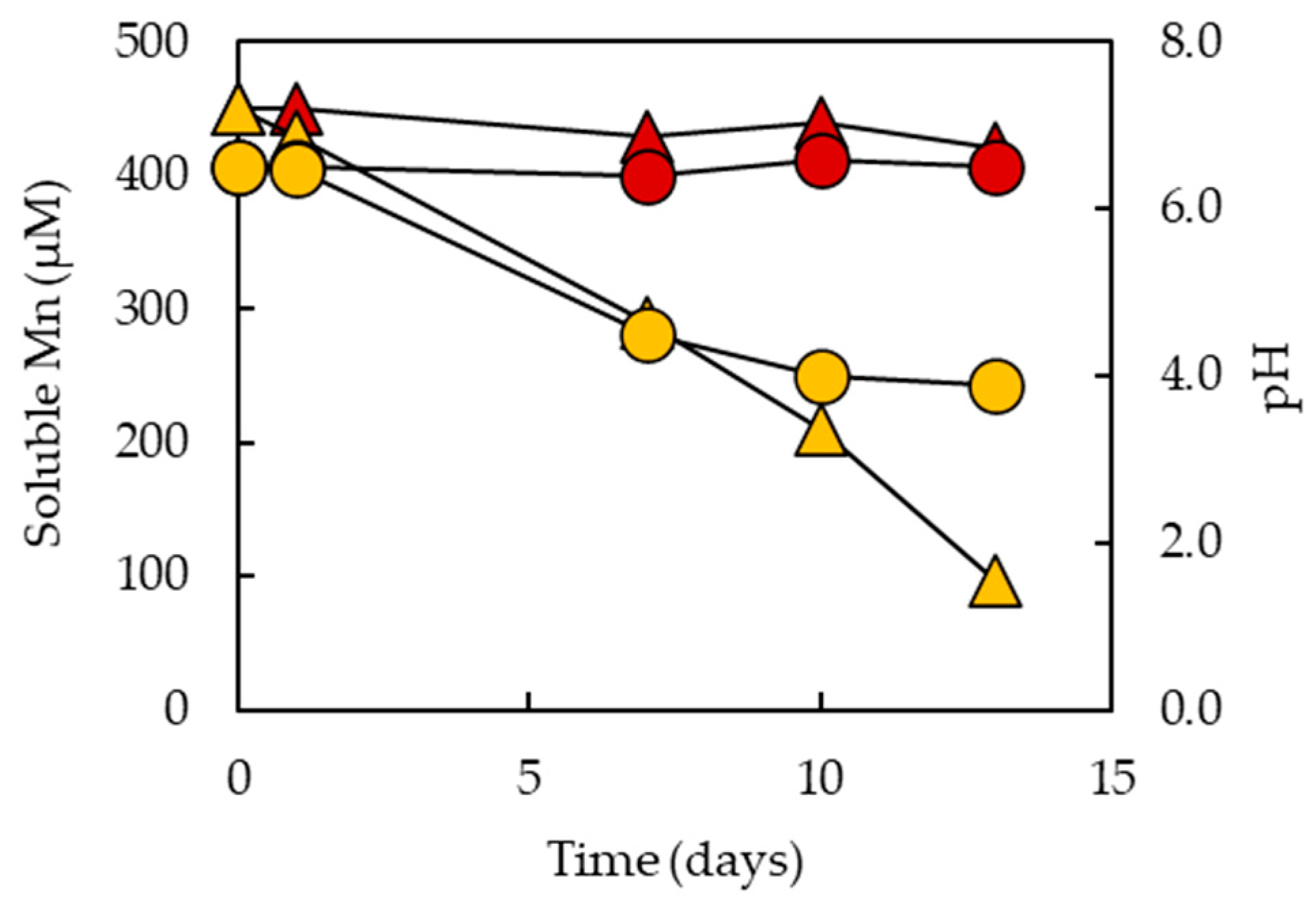
3.1. Removal of Soluble Mn from Partly Processed Mine Water Using the MnOBs
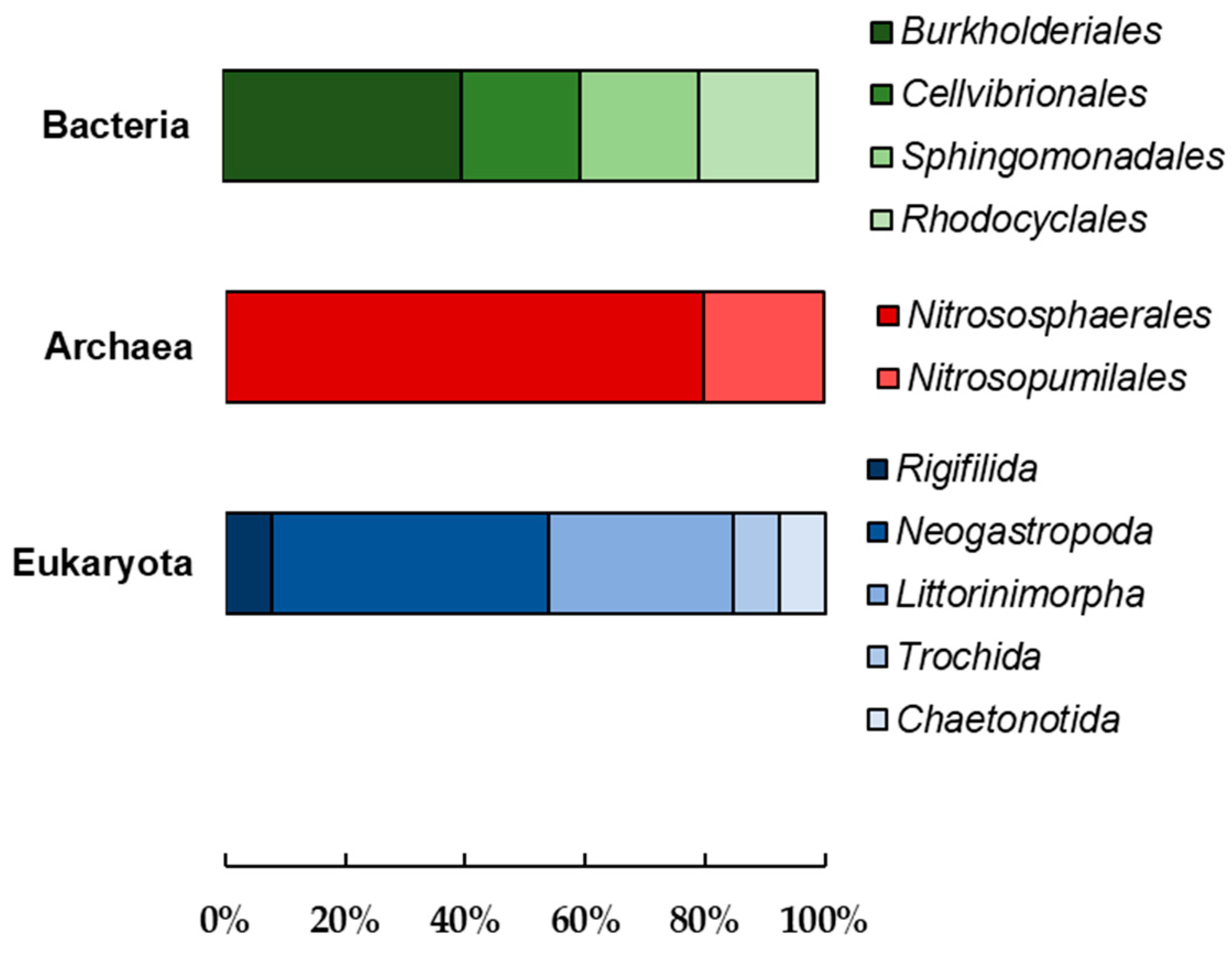
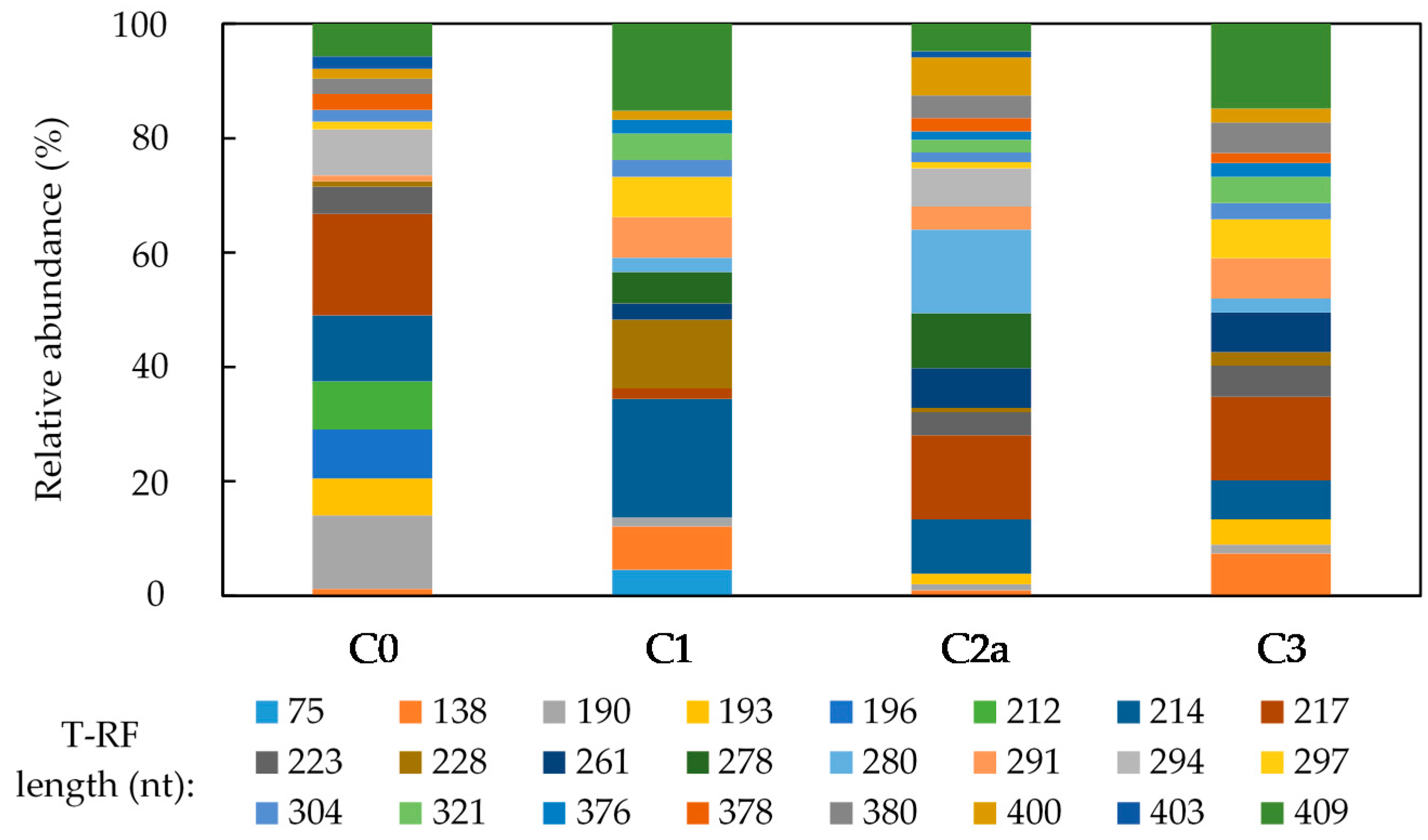
3.2. Microbial Community Analysis of the Mn (IV) Biofilm-Colonized Pebbles
3.3. Characterization of a Mn (II)-Oxidizing Fungal Isolate
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Ehrlich, H.L.; Newman, D.K. Geomicrobiology of Manganese. In Geomicrobiology, 5th ed.; Ehrlich, H.L., Newman, D.K., Eds.; CRC Press: Boca Raton, FL, USA, 2008; pp. 347–420. [Google Scholar]
- Vanẏsek, P. Electrochemical Series. In Handbook of Chemistry and Physics, 93th ed.; Haynes, W.M., Ed.; CRC Press: Boca Raton, FL, USA, 2012; pp. 5–80. [Google Scholar]
- Tebo, B.M.; Bargar, J.R.; Clement, B.G.; Dick, G.J.; Murray, K.J.; Parker, D.; Verity, R.; Webb, S.M. Biogenic Manganese Oxides: Properties and Mechanisms of Formation. Annu. Rev. Earth Planet. Sci. 2004, 32, 287–328. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.; Leadbetter, J.R. Bacterial chemolithoautotrophy via manganese oxidation. Nat. Res. 2020, 583, 453–465. [Google Scholar] [CrossRef] [PubMed]
- Akob, D.M.; Bohu, T.; Beyer, A.; Schäffner, F.; Händel, M.; Johnson, C.A. Identification of Mn(II)-oxidizing bacteria from a low-pH contaminated former uranium mine. Appl. Environ. Microbiol. 2014, 80, 5086–5097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bohu, T.; Akob, D.M.; Abratis, M.; Lazar, C.S.; Kusel, K. Biological low-pH Mn(II) oxidation in a manganese deposit influenced by metal-rich groundwater. Appl. Environ. Microbiol. 2016, 82, 3009–3021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hallberg, K.B.; Johnson, D.B. Biological manganese removal from acid mine drainage in constructed wetlands and prototype bioreactors. Sci. Total Environ. 2005, 338, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Monhemius, A.J. Precipitation diagrams for metal hydroxides, sulfides, arsenates and phosphates. Trans. Inst. Min. Metall. 1977, 86, C202–C206. [Google Scholar]
- Jacob, J.; Raignault, I.; Battaglia-Brunet, F.; Mailhan-Muxi, C.; Engevin, J.; Djemil, M. Biological Manganese Removal from Mine Drainage in a Fixed Bed Bioreactor at Pilot Scale. In Proceedings of the 13th International Conference on Mine Closure, Perth, Australia, 3–5 September 2019; pp. 911–920. [Google Scholar]
- Santos, A.L.; Johnson, D.B. The effects of temperature and pH on the kinetics of an acidophilic sulfidogenic bioreactor and indigenous microbial communities. Hydrometallurgy 2017, 168, 116–120. [Google Scholar] [CrossRef]
- Mariner, R.; Johnson, D.B.; Hallberg, K.B. Characterisation of an attenuation system for the remediation of Mn (II) contaminated waters. Hydrometallurgy 2008, 94, 100–104. [Google Scholar] [CrossRef]
- Lane, D.J. 16S/23S rRNA Sequencing. In Nucleic Acid Techniques in Bacterial Systematics; Stackebrandt, E., Goodfellow, M., Eds.; John Wiley & Sons: Chichester, UK, 1991; pp. 115–175. [Google Scholar]
- Kay, C.M.; Rowe, O.F.; Rocchetti, L.; Coupland, K.; Hallberg, K.B.; Johnson, D.B. Evolution of microbial “streamer” growths in an acidic metal-contaminated stream draining an abandoned underground copper mine. Life 2013, 3, 189–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falagán, C.; Sánchez-Espana, J.; Johnson, D.B. New insights into the biogeochemistry of extremely acidic environments revealed by a combined cultivation-based and culture-independent study of two stratified pit lakes. FEMS Microbiol. Ecol. 2014, 87, 231–243. [Google Scholar] [CrossRef] [PubMed]
- Ñancucheo, I.; Rowe, O.F.; Hedrich, S.; Johnson, D.B. Solid and liquid media for isolating and cultivating acidophilic and acid-tolerant sulfate-reducing bacteria. FEMS Microbiol. Lett. 2016, 363, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mariner, R.; Johnson, D.B.; Hallberg, K.B. Development of a novel biological system for removing manganese from contaminated waters. Adv. Mater. Res. 2007, 20–21, 267–270. [Google Scholar] [CrossRef]
- Duckworth, O.W.; Holmström, S.J.M.; Peña, J.; Sposito, G. Biogeochemistry of iron oxidation in a circum-neutral freshwater habitat. Chem. Geol. 2009, 260, 149–158. [Google Scholar] [CrossRef]
- Stumm, W.; Morgan, J.J. Aquatic Chemistry: An Introduction Emphasizing Chemical Equilibria in Natural Waters; Wiley Interscience: New York, NY, USA, 1981; p. 583. [Google Scholar]
- Ehrlich, H.L. Geomicrobiology, 4th ed.; Marcel Dekker: New York, NY, USA, 2002; p. 800. [Google Scholar]
- Rusconi, R.; Guasto, J.S.; Stocker, R. Bacterial transport suppressed by fluid shear. Nat. Phys. 2014, 10, 212–217. [Google Scholar] [CrossRef] [Green Version]
- Adams, L.F.; Ghiorse, W.C. Physiology and ultrastructure of Leptothrix discophora SS-I*. Arch. Microbiol. 1986, 145, 126–135. [Google Scholar] [CrossRef]
- Miyata, N.; Maruo, K.; Tani, Y.; Tsuno, H.; Seyama, H.; Soma, M.; Iwahori, K. Production of biogenic manganese oxides by anamorphic Ascomycete fungi isolated from streambed pebbles. J. Geomicrobiol. 2006, 23, 63–73. [Google Scholar] [CrossRef]
- Santelli, C.M.; Pfister, D.H.; Lazarus, D.; Sun, L.; Burgos, W.D.; Hansel, C.M. Promotion of Mn(II) oxidation and remediation of coal mine drainage in passive treatment systems by diverse fungal and bacterial communities. Appl. Environ. Microbiol. 2010, 76, 4871–4875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasaki, K.; Konno, H.; Endo, M.; Takano, K. Removal of Mn(II) ions from aqueous neutral media by manganese-oxidizing fungus in the presence of carbon fiber. Biotechnol. Bioeng. 2004, 85, 489–496. [Google Scholar] [CrossRef] [PubMed]
- Industrial Effluent Discharge Act 2011 (No. 430/2011) Brasília: Conselho Nacional Do Meio Ambiente. Available online: http://www.mma.gov.br/port/conama/legiabre.cfm?codlegi=646 (accessed on 21 January 2015).
- Meybeck, M. Carbon, nitrogen, and phosphorus transport by world rivers. Am. J. Sci. 1982, 282, 401–450. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phases | Feed Liquor pH | Flow Rate (mL h−1) | HRT (h) | Duration (Days) |
|---|---|---|---|---|
| C1 | 5.0 ± 0.3 | 92.0 ± 9.7 | 11.3 ± 1.1 | 126 |
| C2a | 6.5 ± 0.3 | 92.0 ± 8.0 | 11.3 ± 1.3 | 28 |
| C2b | 6.5 ± 0.3 | 44.0 ± 2.0 | 23.6 ± 1.6 | 32 |
| C3 | 6.5 ± 0.3 | 23.0 ± 3.0 | 45.2 ± 3.5 | 78 |
| Sulfidogenic Bioreactor | MnOB—Phase C3 | |
|---|---|---|
| Influent | 234 ± 0.7 | 9.2 ± 0.5 |
| Effluent | 9.2 ± 0.5 | 2.4 ± 0.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santos, A.L.; Johnson, D.B. Design and Operation of Empirical Manganese-Removing Bioreactors and Integration into a Composite Modular System for Remediating and Recovering Metals from Acidic Mine Waters. Appl. Sci. 2021, 11, 4287. https://doi.org/10.3390/app11094287
Santos AL, Johnson DB. Design and Operation of Empirical Manganese-Removing Bioreactors and Integration into a Composite Modular System for Remediating and Recovering Metals from Acidic Mine Waters. Applied Sciences. 2021; 11(9):4287. https://doi.org/10.3390/app11094287
Chicago/Turabian StyleSantos, Ana Laura, and D. Barrie Johnson. 2021. "Design and Operation of Empirical Manganese-Removing Bioreactors and Integration into a Composite Modular System for Remediating and Recovering Metals from Acidic Mine Waters" Applied Sciences 11, no. 9: 4287. https://doi.org/10.3390/app11094287