
Microsatellite-Based Genetic Characterization of the Indigenous Katjang Goat in Peninsular Malaysia

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
3. Results
3.1. Microsatellite Marker Assessments
3.2. Genetic Diversity across the Studied Microsatellite Loci
3.3. Genetic Diversity within Katjang Populations
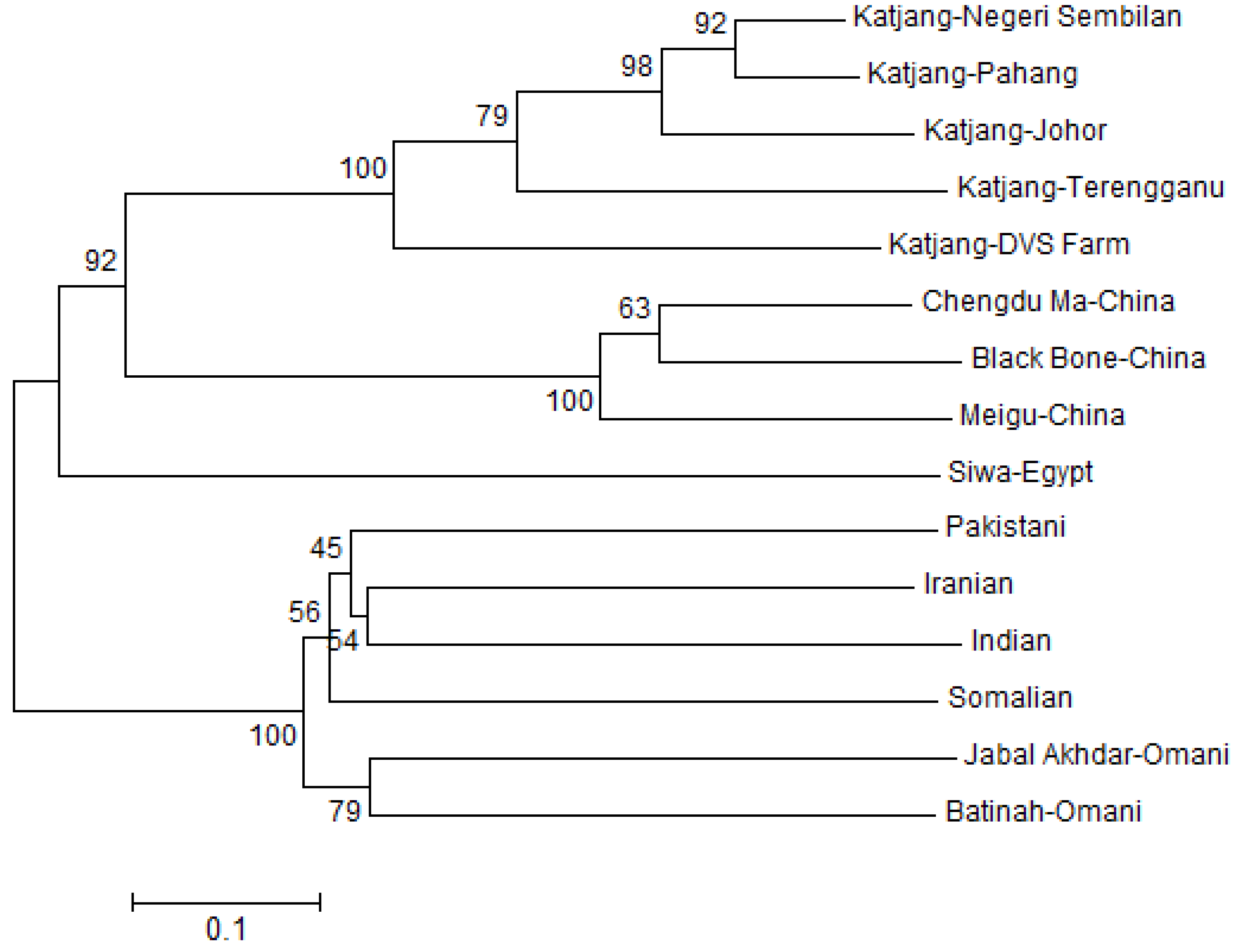
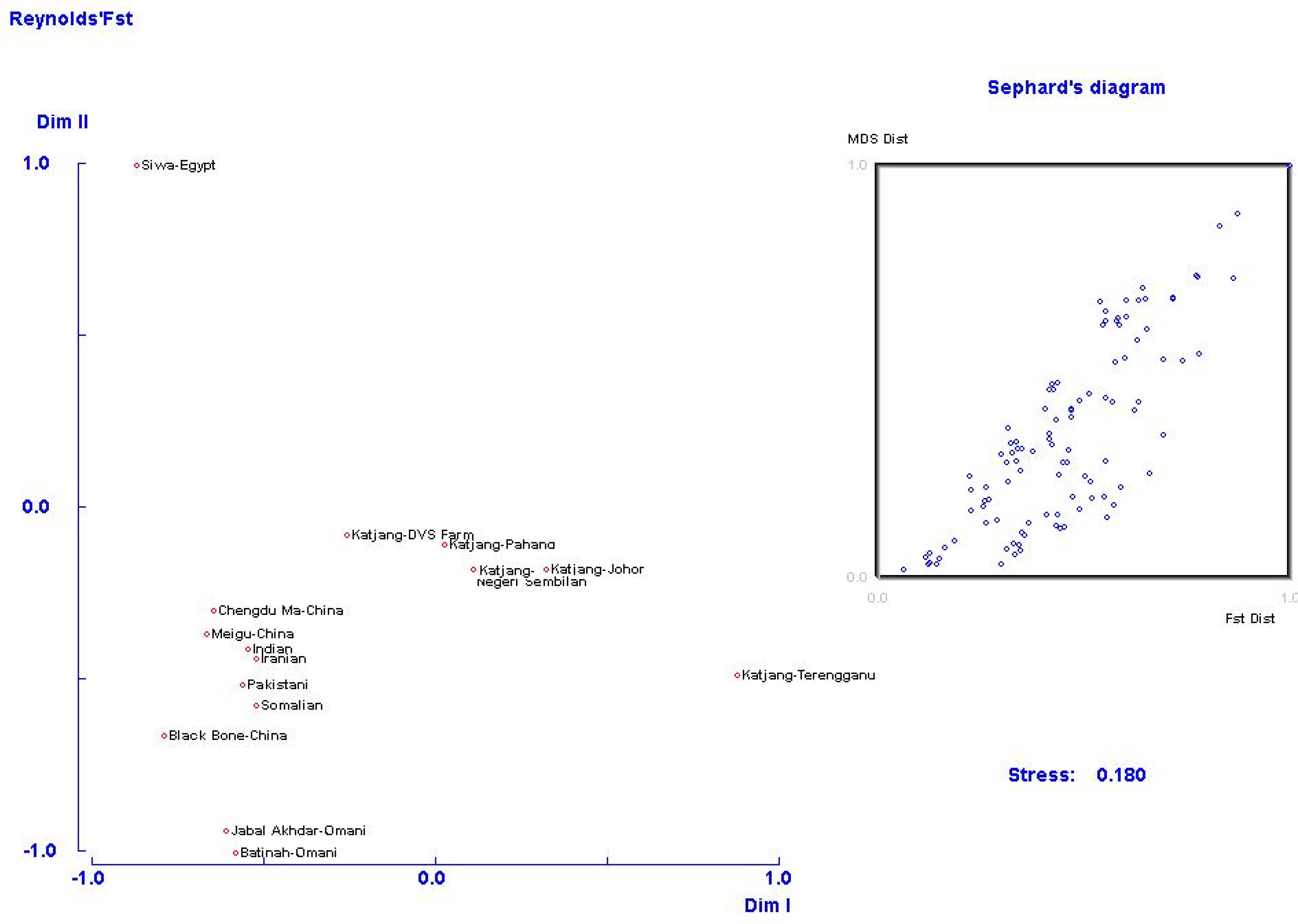
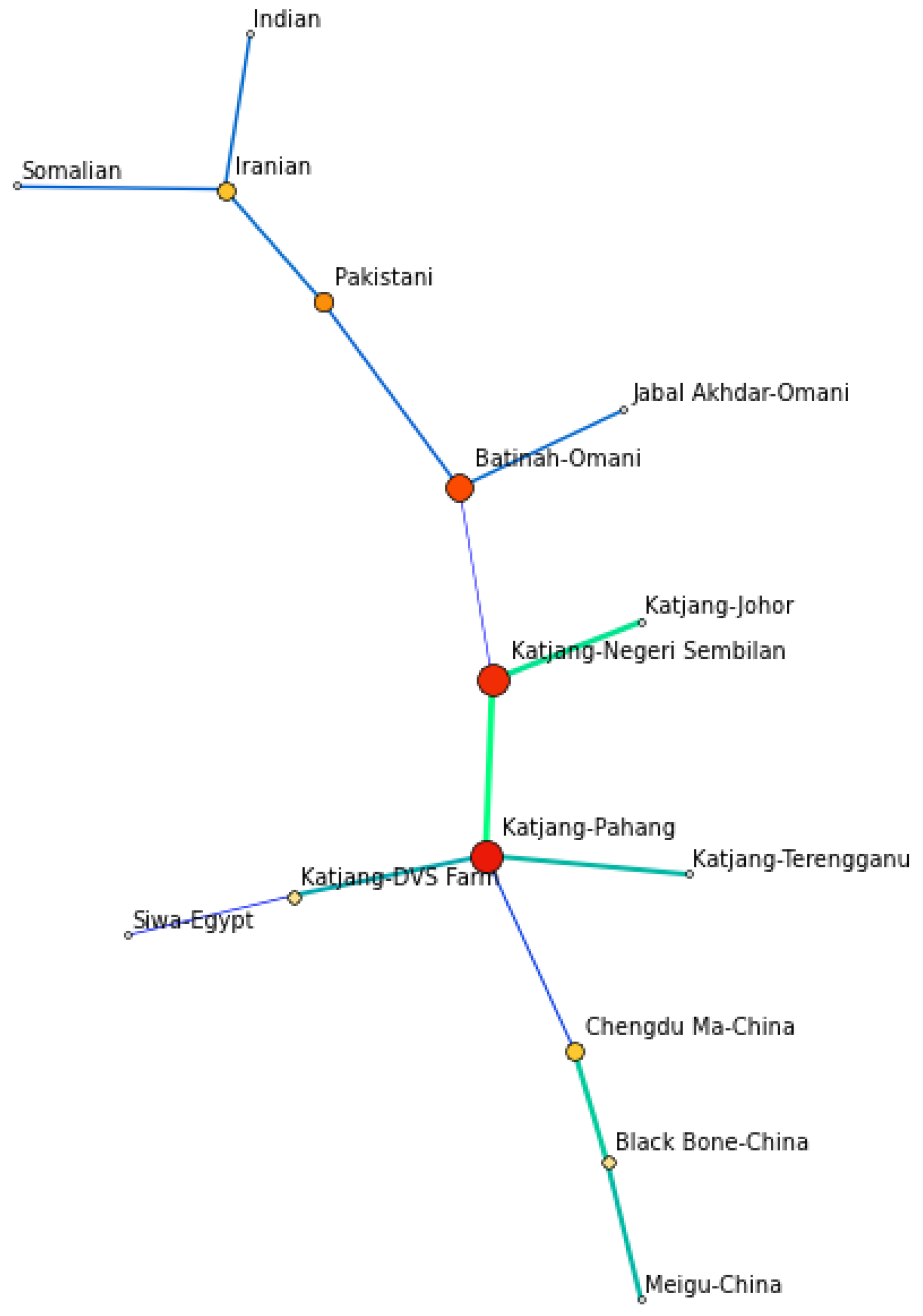
3.4. Population Genetic Relationship and Structure
4. Discussion
4.1. Microsatellite Markers Suitability
4.2. Genetic Diversity of the Katjang Goat
4.3. Population Genetic Relationship and Structure
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Department of Veterinary Services Malaysia. Malaysia: Perangkaan Ternakan. Available online: http://www.dvs.gov.my/dvs/resources/user_1/2020/BP/Perangkaan/3.Muka_Surat_1-12_OK_.pdf (accessed on 28 February 2021).
- Department of Veterinary Services Malaysia. Malaysia: Perangkaan Ternakan Import. Available online: http://www.dvs.gov.my/dvs/resources/user_1/2020/BP/Perangkaan/2._Malaysia_Import_.pdf (accessed on 28 February 2021).
- Devendra, C. Goat Breeds of Malaysia. Malays. Agric. J. 1966, 45, 268–274. [Google Scholar]
- Devendra, C.; Nozawa, K. Goats in South East Asia-Their Status and Production. Z. Tierz. Züchtungsbiol. 1976, 93, 101–120. [Google Scholar] [CrossRef]
- Devendra, C. Goats: Biology, Production, and Development in Asia; Academy of Sciences Malaysia: Kuala Lumpur, Malaysia, 2007. [Google Scholar]
- Barker, J.S.F.; Tan, S.G.; Moore, S.S.; Mukherjee, T.K.; Matheson, J.L.; Selvaraj, O.S. Genetic Variation within and Relationships Among Populations of Asian Goats (Capra Hircus). J. Anim. Breed. Genet. 2001, 118, 213–233. [Google Scholar] [CrossRef]
- Solaiman, S.G. Goat Science and Production; Wiley-Blackwell: Ames, IA, USA, 2010. [Google Scholar]
- Alimon, A.R. Penternakan Kambing; Dewan Bahasa dan Pustaka: Kuala Lumpur, Malaysia, 1988. [Google Scholar]
- Department of Veterinary Services Malaysia. First Report on the State of the World’s Animal Genetic Resources: Animal Genetic Resources in Malaysia; DVS, MOA: Putrajaya, Malaysia, 2003.
- Devendra, C. The Importance of Goats in Malaya. J. Anim. Breed. Genet. 1966, 83, 72–79. [Google Scholar] [CrossRef]
- Devendra, C.; McLeroy, G.B. Goat and Sheep Production in the Tropics; Longman Scientific: London, UK, 1982. [Google Scholar]
- Devendra, C. Studies in the Nutrition of the Indigenous Goat of Malaya. 1. The Body Measurements, Composition of Sample Joints and Their Relationship to Carcass Composition. Malays. Agric. J. 1966, 45, 345–369. [Google Scholar]
- Devendra, C. Pemeliharaan dan Pengeluaran Kambing; Dewan Bhs. Dan Pustaka: Kuala Lumpur, Malaysia, 1986. [Google Scholar]
- Devendra, C.; Burns, M. Goat production in the tropics. In Technical Communications. Commonwealth Bureau of Animal Breeding and Genetics; Commonwealth Agricultural Bureaux: Farnham Royal, UK, 1970. [Google Scholar]
- Peters, K.; Deichert, G.; Drewes, E.; Fichtner, G.; Moll, S.; Chavarria, F.; Diakite, B. Goat Production in Low Income Economic Units of Selected Areas in West-Malaysia. In Proceedings of the Schriften des Seminars fuer Landwirtschaftliche Entwicklung; Technical University of Berlin: Berlin, Germany, 1979. [Google Scholar]
- Gosling, L.A.P. Patterns and Problems of Livestock Production in Malaya. Ph.D. Thesis, University of Michigan, Ann Arbor, MI, USA, 1958. [Google Scholar]
- Hirooka, H.; Mukherjee, T.K.; Panandam, J.M.; Horst, P. Genetic Parameters for Growth Performance of the Malaysian Local Goats and Their Crossbreds with the German (Improved) Fawn Goats. J. Anim. Breed. Genet. 1997, 114, 191–199. [Google Scholar] [CrossRef]
- Porter, V.; Alderson, L.; Hall, S.J.G.; Sponenberg, D.P. Mason’s World Encyclopedia of Livestock Breeds and Breeding; CABI: Wallingford, UK, 2016. [Google Scholar]
- Ariff, O.M.; Hifzan, R.M.; Zuki, A.B.M.; Jiken, A.J.; Lehan, S.M. Maturing Pattern for Body Weight, Body Length and Height at Withers of Jamnapari and Boer Goats. Pertanika J. Trop. Agric. Sci. 2010, 33, 269–276. [Google Scholar]
- Hifzan, M.R.; Nor Amna, A.M.N.; Izuan Bahtiar, A.J.B.; Amie Marini, A.B.; Mohd Hafiz, A.W. Manipulating of Katjang Goat Genetic Material for Sustainable Goat Industry in Malays. Available online: http://ap.fftc.agnet.org/ap_db.php?id=950&print=1 (accessed on 26 October 2019).
- Tsukahara, Y.; Chomei, Y.; Oishi, K.; Kahi, A.K.; Panandam, J.M.; Mukherjee, T.K.; Hirooka, H. Analysis of Growth Patterns in Purebred Kambing Katjang Goat and its Crosses with the German Fawn. Small Rumin. Res. 2008, 80, 8–15. [Google Scholar] [CrossRef]
- Department of Veterinary Services Malaysia; Livestock Breeding Policy. Malaysian Livestock Breeding Policy 2013. Available online: http://www.dvs.gov.my/dvs/resources/user_1/DVS pdf/Livestock_Breeding_Policy.pdf (accessed on 26 October 2019).
- DAD-IS Domestic Animal Diversity Information System (DAD-IS). Available online: http://www.fao.org/dad-is/browse-by-country-and-species/en/ (accessed on 27 October 2019).
- FAO. In Vivo Conservation of Animal Genetic Resources; FAO: Rome, Italy, 2013. [Google Scholar]
- Marwal, A.; Sahu, A.K.; Gaur, R.K. Molecular Markers: Tool for Genetic Analysis. In Animal Biotechnology Models in Discovery and Translation; Verma, A., Singh, A., Eds.; Academic Press: Waltham, MA, USA, 2014; pp. 289–305. [Google Scholar]
- FAO. Molecular Genetic Characterization of Animal Genetic Resources; FAO: Rome, Italy, 2011. [Google Scholar]
- Li, Y.C.; Korol, A.B.; Fahima, T.; Beiles, A.; Nevo, E. Microsatellites: Genomic Distribution, Putative Functions and Mutational Mechanisms: A Review. Mol. Ecol. 2002, 11, 2453–2465. [Google Scholar] [CrossRef]
- Marini, A.; Hifzan, M. Genetic Variation of Four Goat Breeds in Malaysia Using Microsatellite Polymorphism Markers. Malays. J. Anim. Sci. 2013, 16, 1–8. [Google Scholar]
- Li, M.H.; Zhao, S.H.; Bian, C.; Wang, H.S.; Wei, H.; Liu, B.; Yu, M.; Fan, B.; Chen, S.L.; Zhu, M.J.; et al. Genetic Relationships Among Twelve Chinese Indigenous Goat Populations Based on Microsatellite Analysis. Genet. Sel. Evol. 2002, 34, 729–744. [Google Scholar] [CrossRef] [Green Version]
- Kemp, S.J.; Brezinsky, L.; Teale, A.J. A Panel of Bovine, Ovine and Caprine Polymorphic Microsatellites. Anim. Genet. 1993, 24, 363–365. [Google Scholar] [CrossRef] [PubMed]
- Balasingham, T.G.; Robinson, N.A.; McGregor, B.A. Implications for the Conservation of Genetic Diversity in Mohair Goats from a Comparison of a Relic Island Population with Breeds Farmed in Australia. Aust. J. Exp. Agric. 1999, 39, 411–418. [Google Scholar] [CrossRef]
- Lazar, I.; Lazar, I. Gel Analyzer 2010: Freeware 1D gel Electrophoresis Image Analysis Software. Available online: http://www.gelanalyzer.com (accessed on 1 June 2018).
- Glaubitz, J.C. CONVERT: A User-Friendly Program to Reformat Diploid Genotypic Data for Commonly used Population Genetic Software Packages. Mol. Ecol. Notes 2004, 4, 309–310. [Google Scholar] [CrossRef]
- Yeh, F.; Boyle, T. Population Genetic Analysis of Co-Dominant and Dominant Markers and Quantitative Traits. Belg. J. Bot. 1997, 129, 157–163. [Google Scholar]
- Kimura, M.; Crow, J.F. The Number of Alleles that can be Maintained in a Finite Population. Genetics 1964, 49, 725–738. [Google Scholar] [CrossRef] [PubMed]
- Levene, H. On a Matching Problem Arising in Genetics. Ann. Math. Stat. 1949, 20, 91–94. [Google Scholar] [CrossRef]
- Stern, C. The Hardy-Weinberg Law. Science 1943, 97, 137–138. [Google Scholar] [CrossRef] [Green Version]
- Nei, M. Molecular Evolutionary Genetics; Columbia University Press: New York, NY, USA, 1987. [Google Scholar]
- Piry, S.; Luikart, G.; Cornuet, J.M. Bottleneck: A Computer Program for Detecting Recent Reductions in the Effective Population Size Using Allele Frequency Data. J. Hered. 1999, 90, 502–503. [Google Scholar] [CrossRef]
- Di Rienzo, A.; Peterson, A.C.; Garza, J.C.; Valdes, A.M.; Slatkin, M.; Freimer, N.B. Mutational Processes of Simple-Sequence Repeat Loci in Human Populations. Proc. Natl. Acad. Sci. USA 1994, 91, 3166–3170. [Google Scholar] [CrossRef] [Green Version]
- Wilcoxon, F. Individual Comparisons by Ranking Methods. Biom. Bull. 1945, 1, 80–83. [Google Scholar] [CrossRef]
- Luikart, G.; Allendorf, F.W.; Cornuet, J.M.; Sherwin, W.B. Distortion of Allele Frequency Distributions Provides a Test for Recent Population Bottlenecks. J. Hered. 1998, 89, 238–247. [Google Scholar] [CrossRef] [PubMed]
- Guang-Xin, E.; Zhao, Y.J.; Chen, L.P.; Ma, Y.H.; Chu, M.X.; Li, X.L.; Hong, Q.H.; Li, L.H.; Guo, J.J.; Zhu, L.; et al. Genetic Diversity of the Chinese Goat in the Littoral Zone of the Yangtze River as Assessed by Microsatellite and mtDNA. Ecol. Evol. 2018, 8, 5111–5123. [Google Scholar]
- Al-Araimi, N.A.; Gaafar, O.M.; Costa, V.; Neira, A.L.; Al-Atiyat, R.M.; Beja-Pereira, A. Genetic Origin of Goat Populations in Oman Revealed by Mitochondrial DNA Analysis. PLoS ONE 2017, 12, 1–16. [Google Scholar] [CrossRef] [Green Version]
- El-Sayed, M.; Al-Soudy, A.; El Badawy, A. Microsatellite Markers Polymorphism between Two Egyptian Goat Populations (Capra Hircus). Egypt. J. Genet. Cytol. 2016, 45, 89–103. [Google Scholar] [CrossRef]
- Nei, M.; Tajima, F.; Tateno, Y. Accuracy of Estimated Phylogenetic Trees from Molecular Data. J. Mol. Evol. Evol. 1983, 19, 153–170. [Google Scholar] [CrossRef]
- Takezaki, N.; Nei, M.; Tamura, K. POPTREE2: Software for Constructing Population Trees from Allele Frequency Data and Computing other Population Statistics with Windows Interface. Mol. Biol. Evol. 2010, 27, 747–752. [Google Scholar] [CrossRef] [Green Version]
- Pena, J.; Gómez-Pérez, L.; Alfonso-Sánchez, M. GenoCline: On the Trail of Spatial Patterns of Genetic Variation. Authorea Prepr. 2020. [Google Scholar] [CrossRef]
- Kruskal, J.B. Nonmetric Multidimensional Scaling: A Numerical Method. Psychometrika 1964, 29, 115–129. [Google Scholar] [CrossRef]
- Reynolds, J.; Weir, B.S.; Cockerham, C.C. Estimation of the Coancestry Coefficient: Basis for a Short-Term Genetic Distance. Genetics 1983, 105, 767–779. [Google Scholar] [CrossRef] [PubMed]
- Kivelä, M.; Arnaud-Haond, S.; Saramäki, J. EDENetworks: A User-Friendly Software to Build and Analyse Networks in Biogeography, Ecology and Population Genetics. Mol. Ecol. Resour. 2015, 15, 117–122. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, D.B.; Ruiz Linares, A.; Cavalli-Sforza, L.L.; Feldman, M.W. Genetic Absolute Dating Based on Microsatellites and the Origin of Modern Humans. Proc. Natl. Acad. Sci. USA 1995, 92, 6723–6727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stauffer, D.; Aharony, A. Introduction to Percolation Theory; Taylor & Francis: Bristol, PA, USA, 1994. [Google Scholar]
- Al-Araimi, N.A.; Al-Atiyat, R.M.; Luzuriaga-Neira, A.; Mahgoub Gaafar, O.; Kadim, I.T.; Al-Marzooqi, W.; Babiker, H.A.; Al-Kindi, M.N.; Al-Ansari, A.S.; Al-Lawati, A.H.; et al. Genetic Structure of Omani Goats Reveals Admixture Among Populations from Geographically Proximal Sites. Small Rumin. Res. 2019, 178, 1–6. [Google Scholar] [CrossRef]
- Barker, J.S.F. A Global Protocol for Determining Genetic Distances Among Domestic Livestock Breeds. In Proceedings of the 5th World Congress on Genetics Applied to Livestock Production, Guelph, ON, Canada, 7–12 August 1994; Volume 21, pp. 501–508. [Google Scholar]
- FAO. The State of The World’s Animal Genetic Resources for Food and Agriculture; FAO: Rome, Italy, 2007. [Google Scholar]
- Chapuis, M.P.; Estoup, A. Microsatellite Null Alleles and Estimation of Population Differentiation. Mol. Biol. Evol. 2007, 24, 621–631. [Google Scholar] [CrossRef] [Green Version]
- Callen, D.F.; Thompson, A.D.; Shen, Y.; Phillips, H.A.; Richards, R.I.; Mulley, J.C.; Sutherland, G.R. Incidence and Origin of “Null” Alleles in the (AC)n Microsatellite Markers. Am. J. Hum. Genet. 1993, 52, 922–927. [Google Scholar]
- Singh, G.; Thakur, Y.; Kour, A.; Sankhyan, V.; Katoch, S. Genetic Characterization of Gaddi Goat Breed of Western Himalayas Using Microsatellite Markers. Vet. World 2015, 8, 527–531. [Google Scholar] [CrossRef] [Green Version]
- Hirbo, J.; Kemp, S.J.; Hanotte, O.; Kifaro, G.C.; Watts, P.C.; Gwakisa, P.S.; Petersen, P.H.; Chenyambuga, S.W.; Rege, J.E.O. Genetic Characterization of Indigenous Goats of Sub-Saharan Africa Using Microsatellite DNA Markers. Asian-Australas. J. Anim. Sci. 2014, 17, 445–452. [Google Scholar]
- Kharkar, K.; Kuralkar, S.V.; Kuralkar, P. Molecular Genetic Characterization of Berari Breed of Goat Using Microsatellite Markers. Indian J. Anim. Res. 2015, 49, 423–428. [Google Scholar] [CrossRef]
- Jarne, P.; Lagoda, P.J.L. Microsatellites, from Molecules to Populations and Back. Trends Ecol. Evol. 1996, 11, 424–429. [Google Scholar] [CrossRef]
- ISAG/FAO. Secondary Guidelines for Development of National Farm Animal Genetic Resources Management Plans. Measurement of Domestic Animal Diversity (MoDAD): Recommended Microsatellite Markers; FAO: Rome, Italy, 2004. [Google Scholar]
- Hanotte, O.; Jianlin, H. Genetic Characterization of Livestock Populations and its Use in Conservation Decision Making. Role Biotechnol. Explor. Prot. Genet. Resour. 2005, 131–136. [Google Scholar]
- Martínez, A.M.; Carrera, M.P.; Acosta, J.M.; Rodríguez-Gallardo, P.P.; Cabello, A.; Camacho, E.; Delgado, J.V. Genetic Characterisation of the Blanca Andaluza Goat Based on Microsatellite Markers. S. Afr. J. Anim. Sci. 2004, 34, 17–19. [Google Scholar]
- Álvarez, I.; Traoré, A.; Kaboré, A.; Zaré, Y.; Fernández, I.; Tamboura, H.H.; Goyache, F. Microsatellite Analysis of the Rousse de Maradi (Red Sokoto) goat of Burkina Faso. Small Rumin. Res. 2012, 105, 83–88. [Google Scholar] [CrossRef] [Green Version]
- Li, X.L.; Valentini, A. Genetic Diversity of Chinese Indigenous Goat Breeds Based on Microsatellite Markers. J. Anim. Breed. Genet. 2004, 121, 350–355. [Google Scholar]
- Tefiel, H.; Ata, N.; Chahbar, M.; Benyarou, M.; Fantazi, K.; Yilmaz, O.; Cemal, I.; Karaca, O.; Boudouma, D.; Gaouar, S.B.S. Genetic Characterization of four Algerian Goat Breeds Assessed by Microsatellite Markers. Small Rumin. Res. 2018, 160, 65–71. [Google Scholar] [CrossRef]
- Korkmaz Aĝaoĝlu, Ö.; Ertuĝrul, O. Assessment of Genetic Diversity, Genetic Relationship and Bottleneck Using Microsatellites in Some Native Turkish Goat Breeds. Small Rumin. Res. 2012, 105, 53–60. [Google Scholar] [CrossRef]
- Naqvi, A.N.; Bukhari, J.F.; Vahidi, S.M.F.; Utsunomiya, Y.T.; Garcia, J.F.; Babar, M.E.; Han, J.L.; Pichler, R.; Periasamy, K. Microsatellite Based Genetic Diversity and Mitochondrial DNA D-Loop Variation in Economically Important Goat Breeds of Pakistan. Small Rumin. Res. 2017, 148, 62–71. [Google Scholar] [CrossRef] [Green Version]
- Al-Atiyat, R.M.; Alobre, M.M.; Aljumaah, R.S.; Alshaikh, M.A. Microsatellite Based Genetic Diversity and Population Structure of Three Saudi Goat Breeds. Small Rumin. Res. 2015, 130, 90–94. [Google Scholar] [CrossRef]
- Negrini, R.; D’Andrea, M.; Crepaldi, P.; Colli, L.; Nicoloso, L.; Guastella, A.M.; Sechi, T.; Bordonaro, S.; Ajmone-Marsan, P.; Pilla, F. Effect of Microsatellite Outliers on the Genetic Structure of Eight Italian Goat Breeds. Small Rumin. Res. 2012, 103, 99–107. [Google Scholar] [CrossRef]
- Awobajo, O.K.; Salako, A.E.; Osaiyuwu, O.H. Analysis of Genetic Structure of Nigerian West African Dwarf Goats by Microsatellite Markers. Small Rumin. Res. 2015, 133, 112–117. [Google Scholar] [CrossRef]
- Cardoso, T.F.; Amills, M.; Bertolini, F.; Rothschild, M.; Marras, G.; Boink, G.; Jordana, J.; Capote, J.; Carolan, S.; Hallsson, J.H.; et al. Patterns of Homozygosity in Insular and Continental Goat Breeds. Genet. Sel. Evol. 2018, 50, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Cornuet, J.M.; Luikart, G. Description and Power Analysis of Two Tests for Detecting Recent Population Bottlenecks from Allele Frequency Data. Genetics 1996, 144, 2001–2014. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Goat Population | Origin | Data Numbers | References |
|---|---|---|---|
| Chengdu Ma breed | Chengdu, Sichuan, China | 30 | [43] |
| Meigu breed | Meigu, Sichuan, China | 34 | [43] |
| Black-bone breed | Wuhan, Hubei, China | 24 | [43] |
| Siwa, Egypt | Siwa, Egypt | 20 | [45] |
| Jabal Akhdar breed | Oman | 31 | [54] |
| Batinah breed | Oman | 30 | [54] |
| Somalian | Hargeisa, Somalia | 28 | [54] |
| Iranian | Bandar Abbas, Iran | 21 | [54] |
| Pakistani | Gwadar, Pakistan | 26 | [54] |
| Indian | Malegaon, Nasik, India | 21 | [54] |
| Breed/ Population | Microsatellite Loci | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| SRCRSP5 | MAF065 | MAF70 | OarFCB48 | SRCRSP9 | SPS113 | OarFCB20 | CSRD247 | ILSTS029 | SRCRSP8 | OarAE54 | |
| Katjang | X | X | X | X | X | X | X | X | X | X | X |
| Chengdu Ma | X | X | X | X | X | X | X | X | X | X | X |
| Meigu | X | X | X | X | X | X | X | X | X | X | X |
| Black-bone | X | X | X | X | X | X | X | X | X | X | X |
| Siwa | X | X | X | X | X | X | X | ||||
| Jabal Akhdar | X | X | X | X | X | ||||||
| Batinah | X | X | X | X | X | ||||||
| Somalian | X | X | X | X | X | ||||||
| Iranian | X | X | X | X | X | ||||||
| Pakistani | X | X | X | X | X | ||||||
| Indian | X | X | X | X | X | ||||||
| No | Microsatellite Loci | Loci Reference | Allele Size Range (Base Pair) | Allele Polymorphism |
|---|---|---|---|---|
| 1. | SRCRSP5 | [26] | 162–185 | Polymorphic |
| 2. | MAF065 | [26] | 119–144 | Polymorphic |
| 3. | MAF70 | [26] | 137–172 | Polymorphic |
| 4. | SRCRSP23 | [26] | 86–107 | Less than 4 alleles |
| 5. | OarFCB48 | [26] | 148–181 | Polymorphic |
| 6. | INRA023 | [26] | 210–219 | Less than 4 allele |
| 7. | SRCRSP9 | [26] | 112–140 | Polymorphic |
| 8. | SPS113 | [26] | 134–157 | Polymorphic |
| 9. | INRABERN172 | [26] | 247 | Monomorphic |
| 10. | OarFCB20 | [26] | 90–112 | Polymorphic |
| 11. | CSRD247 | [26] | 210–273 | Polymorphic |
| 12. | MCM527 | [26] | 154–165 | Less than 4 alleles |
| 13. | ILSTS087 | [26] | 144–165 | Polymorphic |
| 14. | INRA063 | [26] | 174–184 | Less than 4 alleles |
| 15. | ILSTS011 | [26] | 241–297 | Polymorphic |
| 16. | ILSTS005 | [26] | 180 | Monomorphic |
| 17. | SRCRSP15 | [26] | 180–208 | Polymorphic |
| 18. | SRCRSP3 | [26] | 107–132 | Polymorphic |
| 19. | ILSTS029 | [26] | 156–192 | Polymorphic |
| 20. | TGLA53 | [26] | 127–160 | Polymorphic |
| 21. | ETH10 | [26] | 202–212 | Less than 4 alleles |
| 22. | MAF209 | [26] | 109 | Monomorphic |
| 23. | INRABERN185 | [26] | 247–291 | Polymorphic |
| 24. | P19(DYA) | [26] | 160–195 | Polymorphic |
| 25. | TCRVB6 | [26] | 231–258 | Polymorphic |
| 26. | SRCRSP7 | [26] | 125–135 | Less than 4 alleles |
| 27. | SRCRSP8 | [26] | 209–243 | Polymorphic |
| 28. | DRBP1 | [26] | 107–146 | Polymorphic |
| 29. | OarAE54 | [26] | 114–141 | Polymorphic |
| 30. | BM6444 | [26] | - | Non-amplification |
| 31. | RM004 | [6] | 114 | Monomorphic |
| 32. | ILSTS044 | [30] | 160–172 | Polymorphic |
| 33. | TGLA245 | [6] | 125–162 | Polymorphic |
| 34. | BM1818 | [29] | 251–290 | Polymorphic |
| 35. | OarJMP29 | [31] | 123–138 | Polymorphic |
| 36. | INRA005 | [6] | 130–162 | Polymorphic |
| No | Microsatellite Loci | 1 Na | 2 Ne | 3 Ho | 4 He | 5 FIS |
|---|---|---|---|---|---|---|
| 1. | SRCRSP5 | 6 | 4.19 | 0.33 | 0.77 | 0.44 |
| 2. | MAF065 | 6 | 5.73 | 0.59 | 0.83 | 0.11 |
| 3. | MAF70 | 8 | 4.26 | 0.44 | 0.77 | 0.39 |
| 4. | OarFCB48 | 8 | 3.80 | 0.38 | 0.74 | 0.42 |
| 5. | SRCRSP9 | 8 | 5.87 | 0.41 | 0.83 | 0.34 |
| 6. | SPS113 | 6 | 3.10 | 0.22 | 0.68 | 0.65 |
| 7. | OarFCB20 | 5 | 2.26 | 0.12 | 0.56 | 0.64 |
| 8. | CSRD247 | 15 | 10.05 | 0.46 | 0.91 | 0.34 |
| 9. | ILSTS087 | 6 | 4.84 | 0.34 | 0.80 | 0.32 |
| 10. | ILSTS011 | 9 | 4.26 | 0.31 | 0.77 | 0.39 |
| 11. | SRCRSP15 | 5 | 3.51 | 0.21 | 0.72 | 0.57 |
| 12. | SRCRSP3 | 7 | 5.41 | 0.47 | 0.82 | 0.26 |
| 13. | ILSTS029 | 8 | 1.69 | 0.17 | 0.41 | 0.52 |
| 14. | TGLA53 | 7 | 2.82 | 0.38 | 0.65 | 0.33 |
| 15. | INRABERN185 | 6 | 1.37 | 0.16 | 0.27 | 0.51 |
| 16. | P19(DYA) | 6 | 2.70 | 0.48 | 0.63 | −0.06 |
| 17. | TCRVB6 | 8 | 4.95 | 0.08 | 0.80 | 0.80 |
| 18. | SRCRSP8 | 8 | 5.49 | 0.08 | 0.82 | 0.88 |
| 19. | DRBP1 | 10 | 5.54 | 0.15 | 0.83 | 0.79 |
| 20. | OarAE54 | 8 | 3.92 | 0.28 | 0.75 | 0.49 |
| 21. | ILSTS044 | 4 | 2.56 | 0.19 | 0.62 | 0.37 |
| 22. | TGLA245 | 8 | 4.54 | 0.19 | 0.79 | 0.64 |
| 23. | BM1818 | 9 | 5.78 | 0.28 | 0.83 | 0.53 |
| 24. | OarJMP29 | 4 | 2.48 | 0.28 | 0.60 | 0.31 |
| 25. | INRA005 | 6 | 3.90 | 0.31 | 0.75 | 0.37 |
| Mean | 7.24 | 4.20 | 0.29 | 0.72 | 0.46 | |
| Standard deviation | 2.24 | 1.80 | 0.14 | 0.14 |
| Population | 1 Ho | 2 He | 3 FIS |
|---|---|---|---|
| DVS Farm | 0.37 ± 0.23 | 0.52 ± 0.20 | 0.25 |
| Negeri Sembilan | 0.28 ± 0.19 | 0.63 ± 0.18 | 0.55 |
| Pahang | 0.29 ± 0.16 | 0.66 ± 0.17 | 0.56 |
| Johor | 0.30 ± 0.21 | 0.54 ± 0.19 | 0.40 |
| Terengganu | 0.25 ± 0.24 | 0.51 ± 0.26 | 0.48 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ernie Muneerah, M.A.; Md Tamrin, N.A.; Salisi, M.S.; Zulkifly, S.; Ghazali, S.S.M.; Temuli, J.J.; Rosali, M.H.; Nazari, S.; Wan Nik, W.M.K.; Mamat-Hamidi, K. Microsatellite-Based Genetic Characterization of the Indigenous Katjang Goat in Peninsular Malaysia. Animals 2021, 11, 1328. https://doi.org/10.3390/ani11051328
Ernie Muneerah MA, Md Tamrin NA, Salisi MS, Zulkifly S, Ghazali SSM, Temuli JJ, Rosali MH, Nazari S, Wan Nik WMK, Mamat-Hamidi K. Microsatellite-Based Genetic Characterization of the Indigenous Katjang Goat in Peninsular Malaysia. Animals. 2021; 11(5):1328. https://doi.org/10.3390/ani11051328
Chicago/Turabian StyleErnie Muneerah, Mohd Adhan, Nur Aida Md Tamrin, Mohd Shahrom Salisi, Shahrizim Zulkifly, Siti Shaidatul Maisarah Ghazali, Jackson Jenun Temuli, Mohd Hifzan Rosali, Shariffah Nazari, Wan Mohd Kamil Wan Nik, and Kamalludin Mamat-Hamidi. 2021. "Microsatellite-Based Genetic Characterization of the Indigenous Katjang Goat in Peninsular Malaysia" Animals 11, no. 5: 1328. https://doi.org/10.3390/ani11051328