Dendranthema zawadskii var. lucidum (Nakai) J.H. Park Extract Inhibits Cellular Senescence in Human Dermal Fibroblasts and Aging-Related Inflammation in Rats

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.1.1. Plant Resource and Extract Preparation
2.1.2. Chemicals
2.2. Cell Line and Cell Culture
2.3. Cell Viability Assay
2.4. Induction of Cellular Senescence by Doxorubicin
2.5. Senescence-Associated β-Galactosidase (SA-β-gal) Activity Assay
2.6. Experimental Animals and Treatment
2.7. Tissue Preparation and Measurement of Oxidative Stress
2.8. Western Blot Analysis of Aorta Tissue
2.9. Phenolic Compound Analysis Using a HPLC–UVD
2.10. Statistical Analysis
3. Results
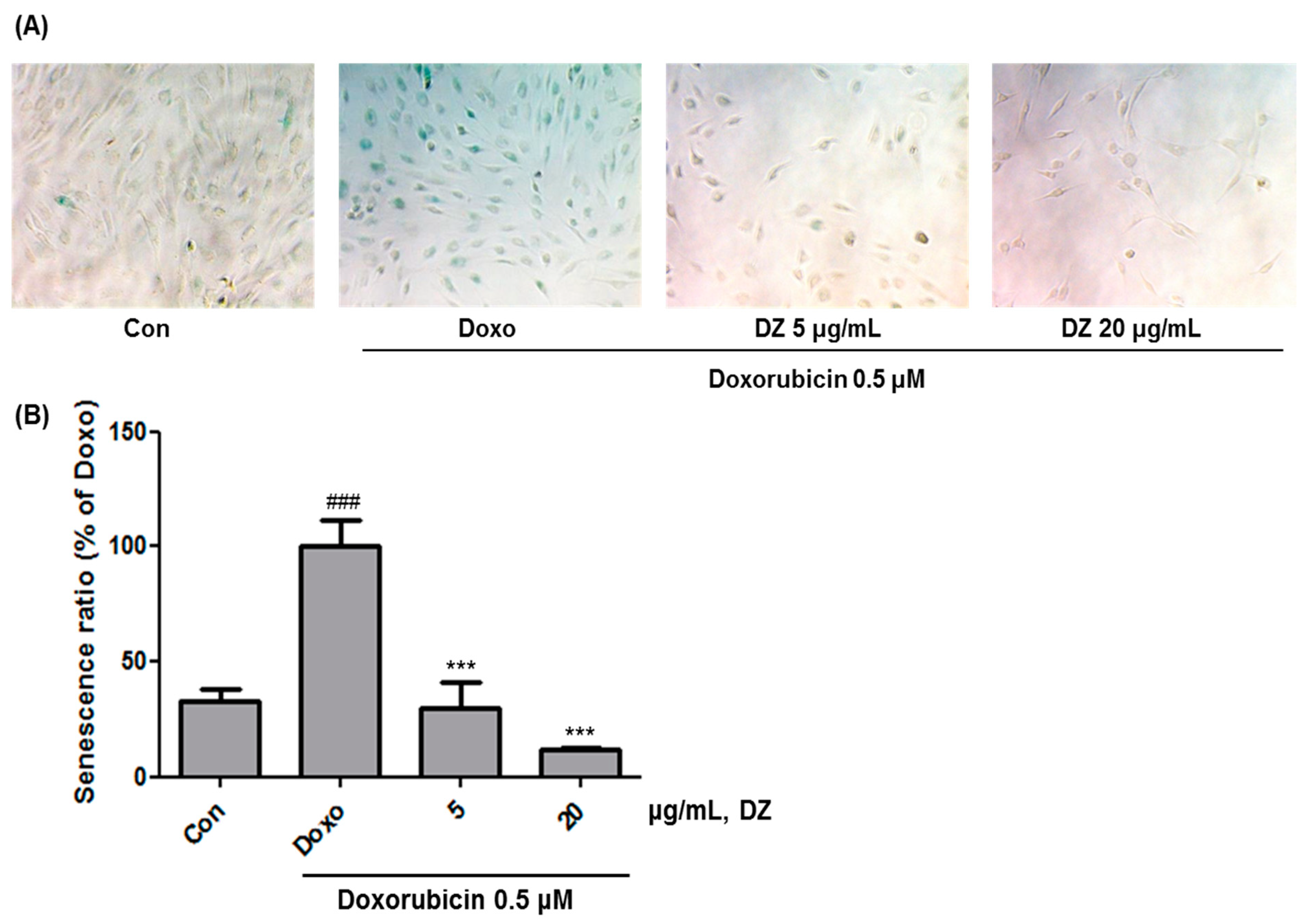
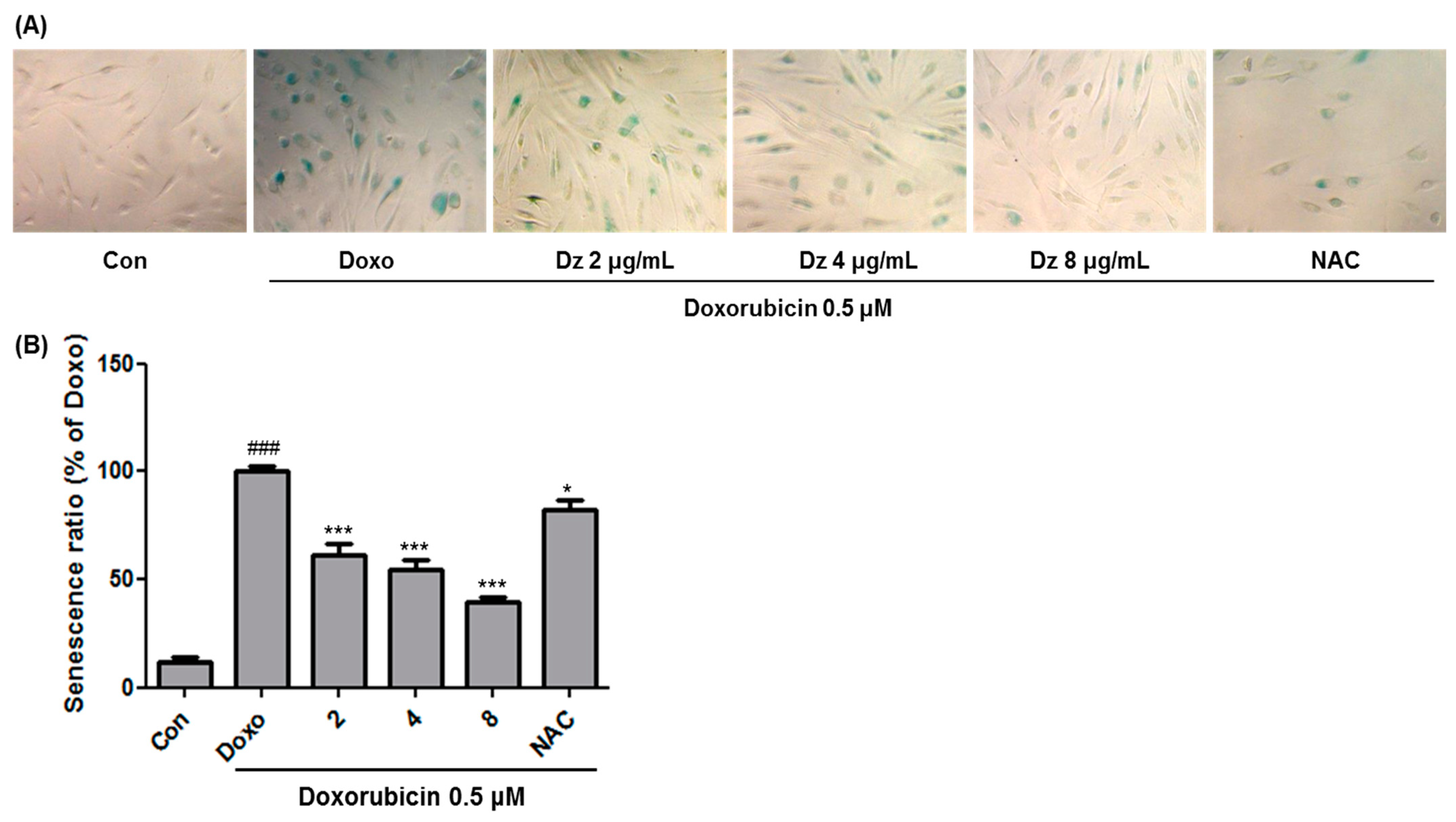
3.1. Inhibition of Cellular Senescence in HDFs by DZ Extract
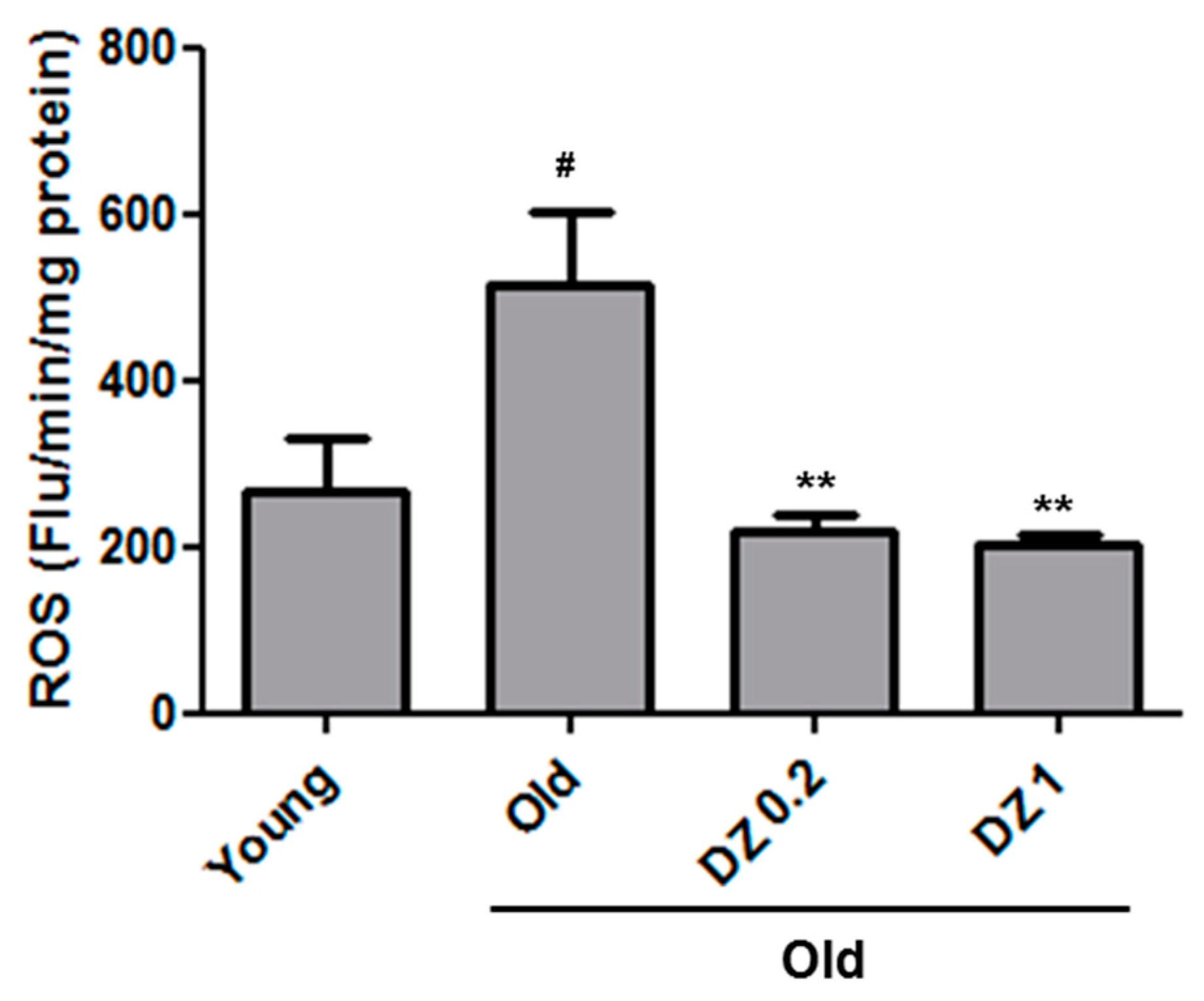
3.2. Effects of DZ Extract on Redox Status in the Descending Aorta of Aged Rats
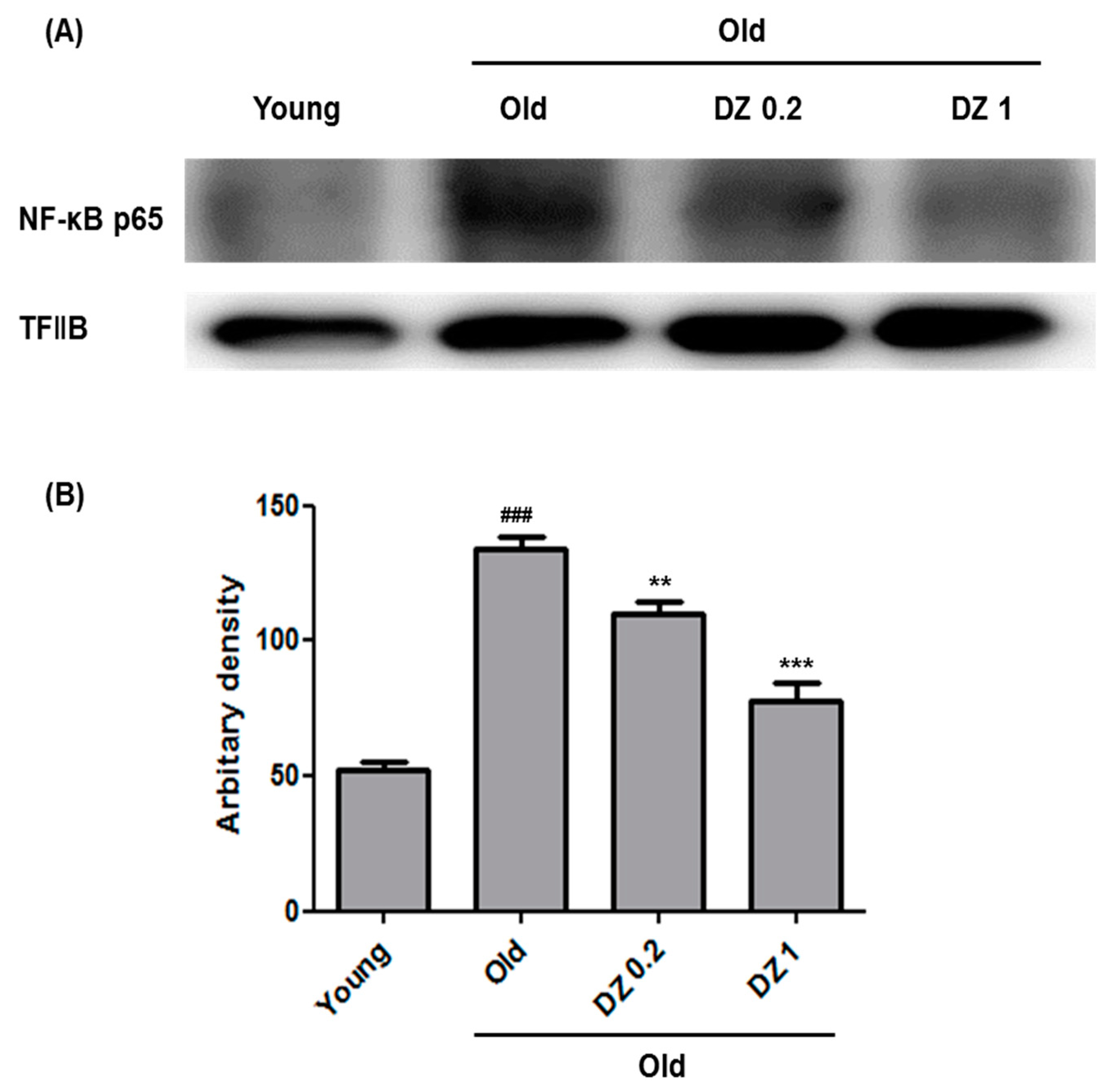
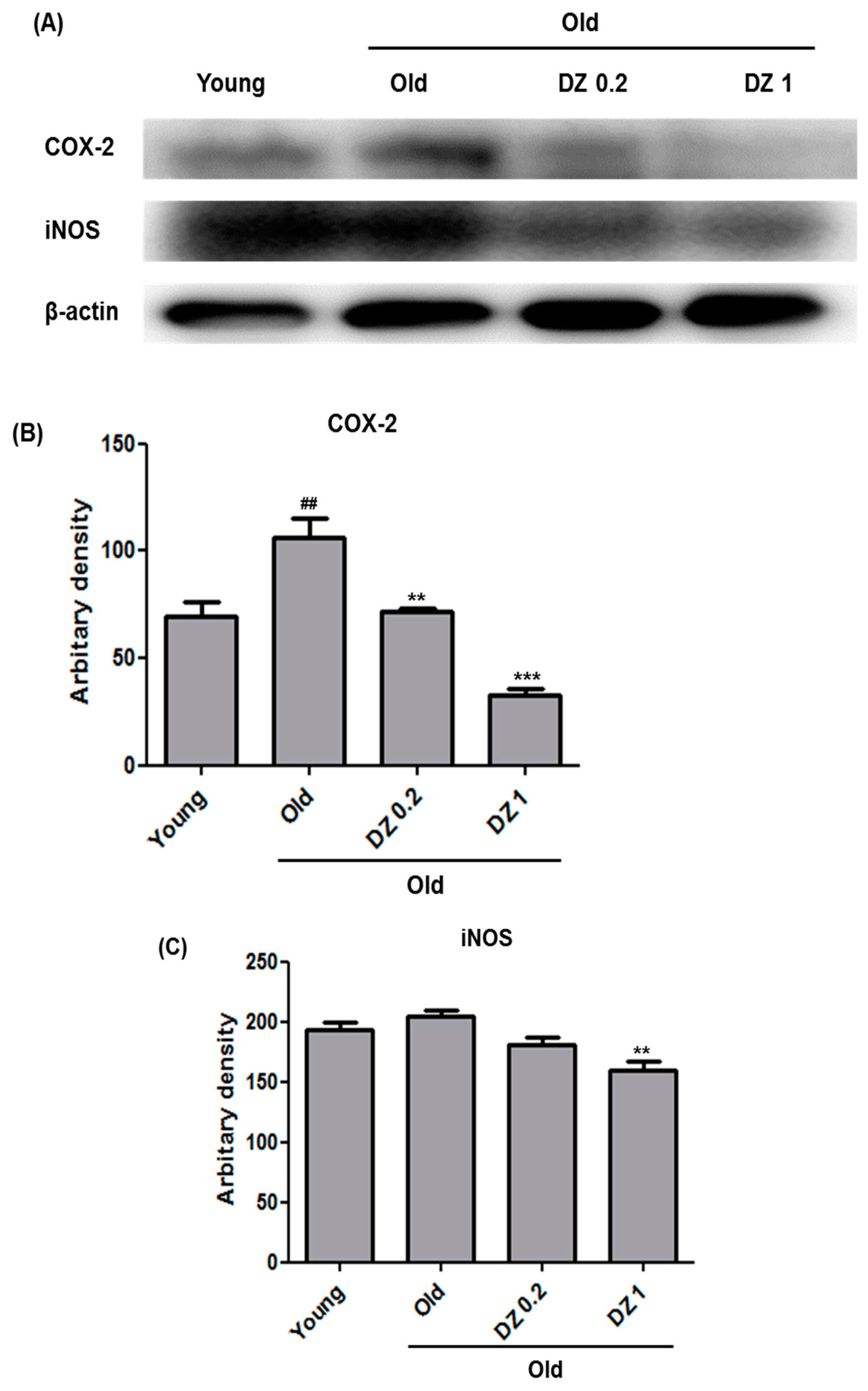
3.3. Suppressive Effect of DZ Extract on Age-Induced Nuclear Factor Kappa-Light-Chain-Enhancer of Activated B Cells (NF-κB) Activation and Inflammation in Rat Aorta
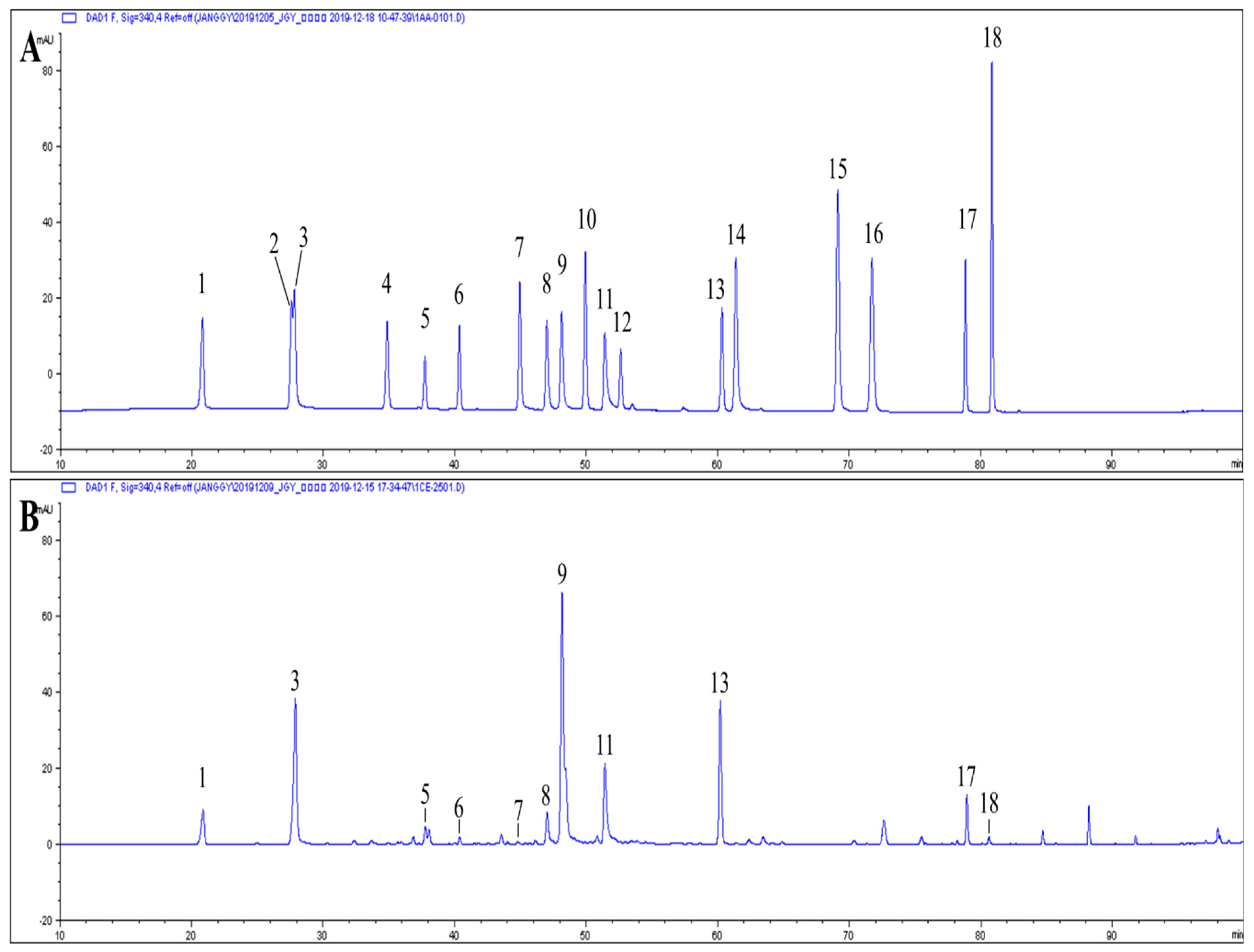
3.4. Major Phenolic Compounds of Dendranthema zawadskii (DZ) Extract
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Dimri, G.P.; Lee, X.; Basile, G.; Acosta, M.; Scott, G.; Roskelley, C.; Medrano, E.E.; Linskens, M.; Rubelj, I.; Pereira-Smith, O. A biomarker that identifies senescent human cells in culture and in aging skin in vivo. Proc. Natl. Acad. Sci. USA 1995, 92, 9363–9367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harman, D. Aging: A theory based on free radical and radiation chemistry. J. Gerontol. 1955, 11, 298–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, H.Y.; Cesari, M.; Anton, S.; Marzetti, E.; Giovannini, S.; Seo, A.Y.; Carter, C.; Yu, B.P.; Leeuwenburgh, C. Molecular inflammation: Underpinnings of aging and age-related diseases. Ageing Res. Rev. 2009, 8, 18–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, H.T.; Kim, Y.S. The Establishment of conservation area and conservation strategy in Ulleung Island(1) flora. Korean J. Environ. Ecol. 2002, 16, 195–216. [Google Scholar]
- Lee, C.B. Illustrated Flora of Korea; Hyangmoonsa: Seoul, Korea, 2003. [Google Scholar]
- Choi, Y.J. Traditional Plant in Korea; Academic Books: Seoul, Korea, 1992. [Google Scholar]
- Shin, G.C.; Shin, Y.C. New Our Talk Large a Dictionary.; Samsung Publishing Company: Seoul, Korea, 1992. [Google Scholar]
- Woo, K.S.; Yu, J.S.; Hwang, I.G.; Lee, Y.R.; Lee, C.H.; Yoon, H.S.; Lee, J.S.; Jeong, H.S. Antioxidative activity of volatile compounds in flower of Chrysanthemum indicum, C. morifolium, and C. zawadskii. J. Korean Soc. Food Sci. Nutr. 2008, 37, 805–809. [Google Scholar] [CrossRef]
- Seo, J.Y.; Lim, S.S.; Park, J.; Lim, J.-S.; Kim, H.J.; Kang, H.J.; Park, J.H.Y.; Kim, J.-S. Protection by Chrysanthemum zawadskii extract from liver damage of mice caused by carbon tetrachloride is maybe mediated by modulation of QR activity. Nutr. Res. Pract. 2010, 4, 93. [Google Scholar] [CrossRef]
- Gu, D.R.; Hwang, J.-K.; Erkhembaatar, M.; Kwon, K.-B.; Kim, M.S.; Lee, Y.-R.; Lee, S.H. Inhibitory effect of Chrysanthemum zawadskii Herbich var. latilobum Kitamura extract on RANKL-induced osteoclast differentiation. Evid.-Based Compl. Alt. Med. 2013, 2013. [Google Scholar] [CrossRef]
- Byun, J.-H.; Choi, C.-W.; Jang, M.-J.; Lim, S.H.; Han, H.J.; Choung, S.-Y. Anti-Osteoarthritic Mechanisms of Chrysanthemum zawadskii var. latilobum in MIA-Induced Osteoarthritic Rats and Interleukin-1β-Induced SW1353 Human Chondrocytes. Medicina 2020, 56, 685. [Google Scholar] [CrossRef]
- Jung, S.-Y.; Park, S.-H.; Nam, C.-H.; Lee, H.-J.; Lee, Y.-M.; Chang, K.-S. The distribution of vascular plants in Ulleungdo and nearby island regions (Gwaneumdo, Jukdo), Korea. J. Asia-Pacific Biodiv. 2013, 6, 123–156. [Google Scholar] [CrossRef] [Green Version]
- Woo, J.-H.; Shin, S.-L.; Lee, C.-H. Antioxidant effect of 80% ethanol extracts obtained from three Dendranthema species. Korean J. Plant. Res. 2010, 23, 47–53. [Google Scholar]
- Yoon, I.K.; Kim, H.K.; Kim, Y.K.; Song, I.H.; Kim, W.; Kim, S.; Baek, S.H.; Kim, J.H.; Kim, J.R. Exploration of replicative senescence-associated genes in human dermal fibroblasts by cDNA microarray technology. Exp. Gerontol. 2004, 39, 1369–1378. [Google Scholar] [CrossRef]
- Kim, K.S.; Seu, Y.B.; Baek, S.H.; Kim, M.J.; Kim, K.J.; Kim, J.H.; Kim, J.R. Induction of cellular senescence by insulin-like growth factor binding protein-5 through a p53-dependent mechanism. Mol. Biol. Cell 2007, 18, 4543–4552. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.H.; Hwangbo, K.; Zheng, M.S.; Cho, J.H.; Son, J.K.; Kim, H.Y.; Baek, S.H.; Choi, H.C.; Park, S.Y.; Kim, J.R. Quercetin-3-O-β-d-glucuronide isolated from Polygonum aviculare inhibits cellular senescence in human primary cells. Arch. Pharmacal Res. 2014, 37, 1219–1233. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.H.; Hwangbo, K.; Zheng, M.S.; Son, J.K.; Kim, H.Y.; Baek, S.H.; Choi, H.C.; Park, S.Y.; Kim, J.R. Inhibitory effects of juglanin on cellular senescence in human dermal fibroblasts. J. Nat. Med. 2014, 68, 473–480. [Google Scholar] [CrossRef]
- Lee, E.K.; Jang, E.J.; Jung, K.J.; Kim, D.H.; Yu, B.P.; Chung, H.Y. Betaine attenuates lysophosphatidylcholine-mediated adhesion molecules in aged rat aorta: Modulation of the nuclear factor-κB pathway. Exp. Gerontol. 2013, 48, 517–524. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.J.; Kim, H.S.; Lee, J.; Chung, J.; Lee, J.S.; Choi, J.S.; Yoon, T.R.; Kim, H.K.; Chung, H.Y. Down-regulation of oxidative stress and COX-2 and iNOS expressions by dimethyl lithospermate in aged rat kidney. Arch. Pharm. Res. 2014, 37, 1032–1038. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.H.; Hwangbo, K.; Zheng, M.S.; Cho, J.H.; Son, J.-K.; Kim, H.Y.; Baek, S.H.; Choi, H.C.; Park, S.Y.; Kim, J.-R. Inhibitory effects of (−)-loliolide on cellular senescence in human dermal fibroblasts. Arch. Pharmacal Res. 2014, 38, 876–884. [Google Scholar] [CrossRef] [PubMed]
- Yu, B.P. Aging and oxidative stress: Modulation by dietary restriction. Free Radic. Biol. Med. 1996, 21, 651–668. [Google Scholar] [CrossRef]
- Chung, H.Y.; Kim, H.J.; Kim, J.W.; Yu, B.P. The inflammation hypothesis of aging. Ann. N. Y. Acad. Sci. 2001, 928, 327–335. [Google Scholar] [CrossRef]
- Chung, H.Y.; Kim, H.J.; Kim, K.W.; Choi, J.S.; Yu, B.P. Molecular inflammation hypothesis of aging based on the anti-aging mechanism of calorie restriction. Microsc. Res. Techniq. 2002, 59, 264–272. [Google Scholar] [CrossRef]
- Zou, Y.; Kim, D.H.; Jung, K.J.; Heo, H.S.; Kim, C.H.; Baik, H.S.; Yu, B.P.; Yokozawa, T.; Chung, H.Y. Lysophosphatidylcholine enhances oxidative stress via the 5-lipoxygenase pathway in rat aorta during aging. Rejuv. Res. 2009, 12, 15–24. [Google Scholar] [CrossRef]
- Hao, B.-J.; Wu, Y.-H.; Wang, J.-G.; Hu, S.-Q.; Keil, D.J.; Hu, H.-J.; Lou, J.-D.; Zhao, Y. Hepatoprotective and antiviral properties of isochlorogenic acid A from Laggera alata against hepatitis B virus infection. J. Ethnopharmacol. 2012, 144, 190–194. [Google Scholar] [CrossRef]
- Ganzon, J.G.; Chen, L.-G.; Wang, C.-C. 4-O-Caffeoylquinic acid as an antioxidant marker for mulberry leaves rich in phenolic compounds. J. Food Drug Anal. 2018, 26, 985–993. [Google Scholar] [CrossRef]
- Han, X.; Wu, Y.C.; Meng, M.; Sun, Q.S.; Gao, S.M.; Sun, H. Linarin prevents LPS-induced acute lung injury by suppressing oxidative stress and inflammation via inhibition of TXNIP/NLRP3 and NF-κB pathways. Int. J. Mol. Med. 2018, 42, 1460–1472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, B.; Lee, J.H.; Seo, M.-J.; Eom, S.H.; Kim, W. Linarin down-regulates phagocytosis, pro-inflammatory cytokine production, and activation marker expression in RAW264. 7 macrophages. Food Sci. Biotechnol. 2016, 25, 1437–1442. [Google Scholar] [CrossRef]
- Su, X.; Zhu, Z.-h.; Zhang, L.; Wang, Q.; Xu, M.-m.; Lu, C.; Zhu, Y.; Zeng, J.; Duan, J.-A.; Zhao, M. Anti-inflammatory property and functional substances of Lonicerae Japonicae Caulis. J. Ethnopharmacol. 2021, 267, 113502. [Google Scholar] [CrossRef] [PubMed]
- Chai, R.K.; Zhou, X.M.; Xia, M.L.; Pan, X.Y. Effects of dendranthema morifolium on learning, memory and moving abilities in aging mice induced by D-galactose. Cell Biol. Intern. 2008, 32, S8. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phenolic Compounds (1) | Content (μg/g Extract, d.b.) | ||
|---|---|---|---|
| Neochlorogenic acid | 5091.90 | ± | 105.83 |
| 4-O-caffeoylquinic acid | 16,985.07 | ± | 198.14 |
| Vicenin-1 | 1963.01 | ± | 181.48 |
| Isoschaftoside | 1005.34 | ± | 66.35 |
| Luteolin 7-O-glucoside | 590.92 | ± | 62.22 |
| Isochlorogenic acid B | 4365.56 | ± | 293.59 |
| Isochlorogenic acid A | 26,477.89 | ± | 830.66 |
| Isochlorogenic acid C | 10,598.95 | ± | 454.09 |
| Linarin | 16,480.55 | ± | 399.13 |
| Eupatorin | 4228.48 | ± | 131.40 |
| Acacetin | 412.72 | ± | 66.59 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, J.; Jang, G.-Y.; Lee, J.; Chung, H.-Y.; Noh, H.-J.; Kim, G.-S.; Lee, S.-E. Dendranthema zawadskii var. lucidum (Nakai) J.H. Park Extract Inhibits Cellular Senescence in Human Dermal Fibroblasts and Aging-Related Inflammation in Rats. Processes 2021, 9, 801. https://doi.org/10.3390/pr9050801
Choi J, Jang G-Y, Lee J, Chung H-Y, Noh H-J, Kim G-S, Lee S-E. Dendranthema zawadskii var. lucidum (Nakai) J.H. Park Extract Inhibits Cellular Senescence in Human Dermal Fibroblasts and Aging-Related Inflammation in Rats. Processes. 2021; 9(5):801. https://doi.org/10.3390/pr9050801
Chicago/Turabian StyleChoi, Jehun, Gwi-Yeong Jang, Jeonghoon Lee, Hae-Young Chung, Hyung-Jun Noh, Geum-Soog Kim, and Seung-Eun Lee. 2021. "Dendranthema zawadskii var. lucidum (Nakai) J.H. Park Extract Inhibits Cellular Senescence in Human Dermal Fibroblasts and Aging-Related Inflammation in Rats" Processes 9, no. 5: 801. https://doi.org/10.3390/pr9050801