
Summer Diet of Horses (Equus ferus caballus Linn.), Guanacos (Lama guanicoe Müller), and European Brown Hares (Lepus europaeus Pallas) in the High Andean Range of the Coquimbo Region, Chile

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
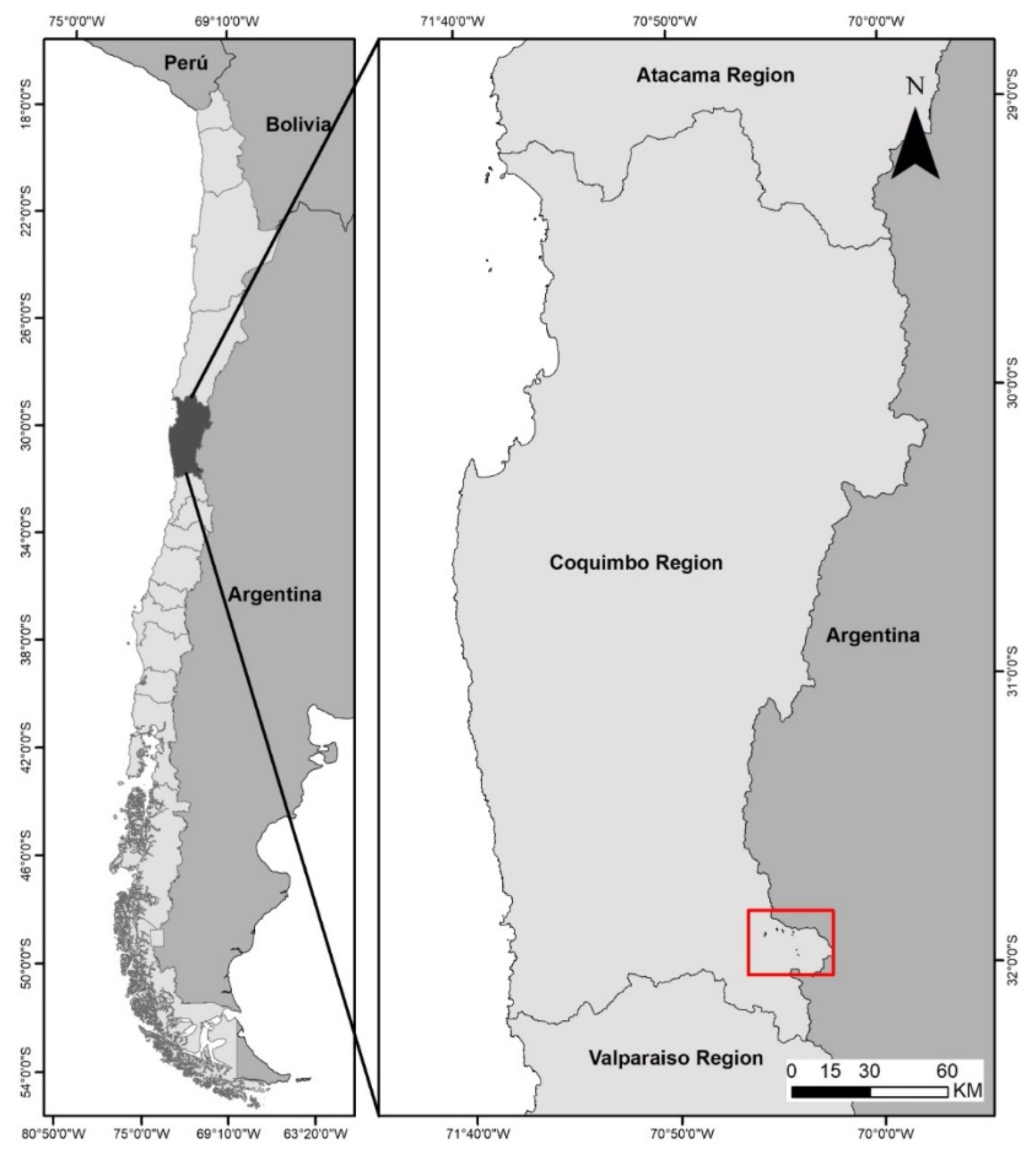
2.1. Characteristics of Study Area
2.2. Methods
2.3. Data Analysis
3. Results
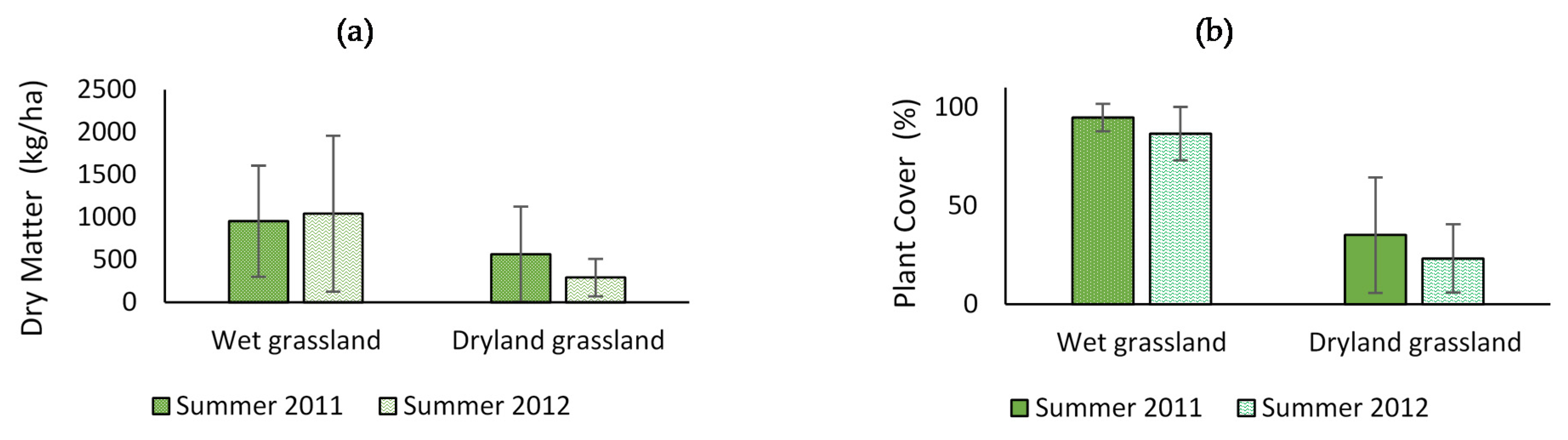
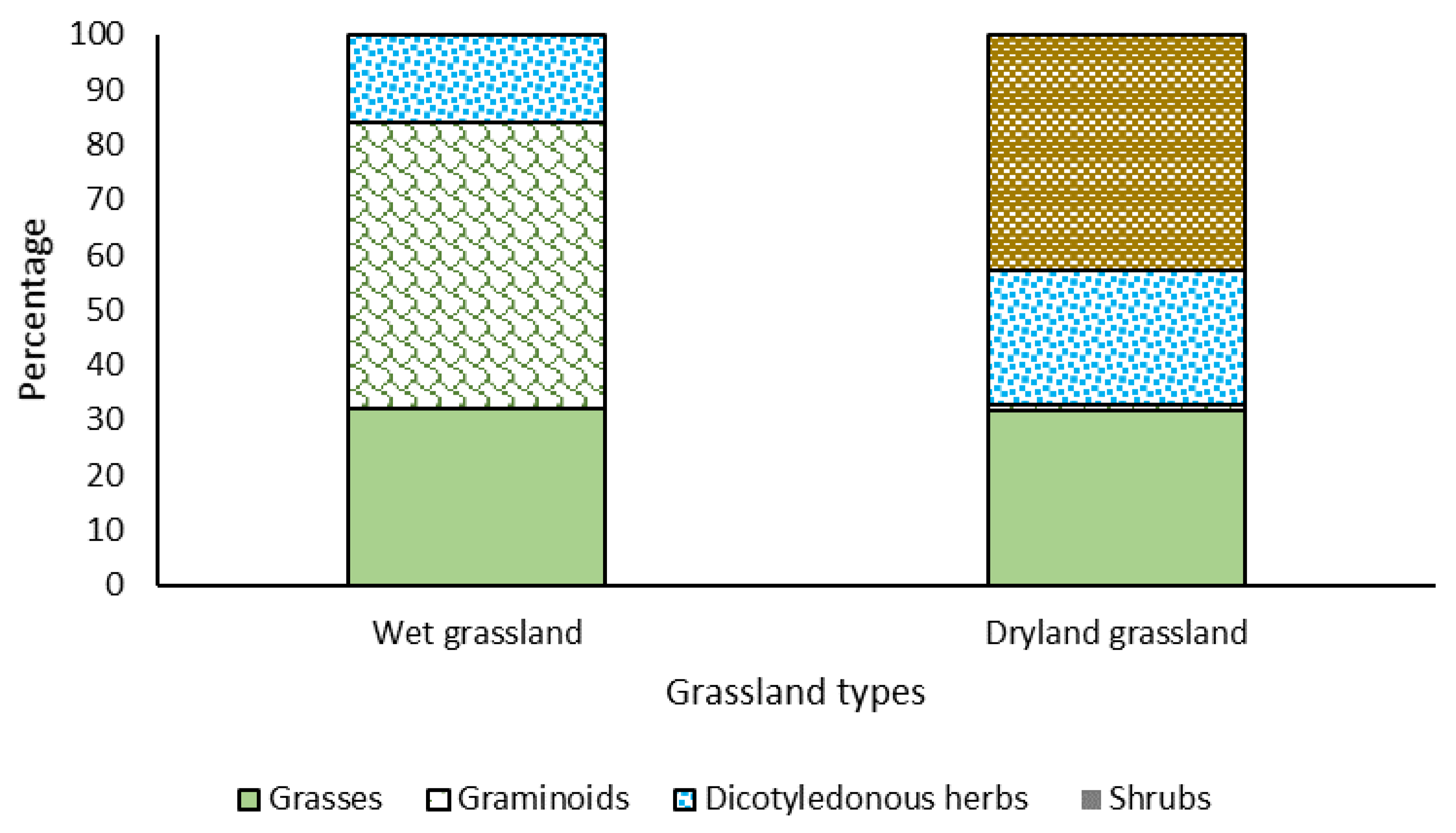
3.1. Dry Matter Production, Vegetation Cover and Grassland Botanical Composition
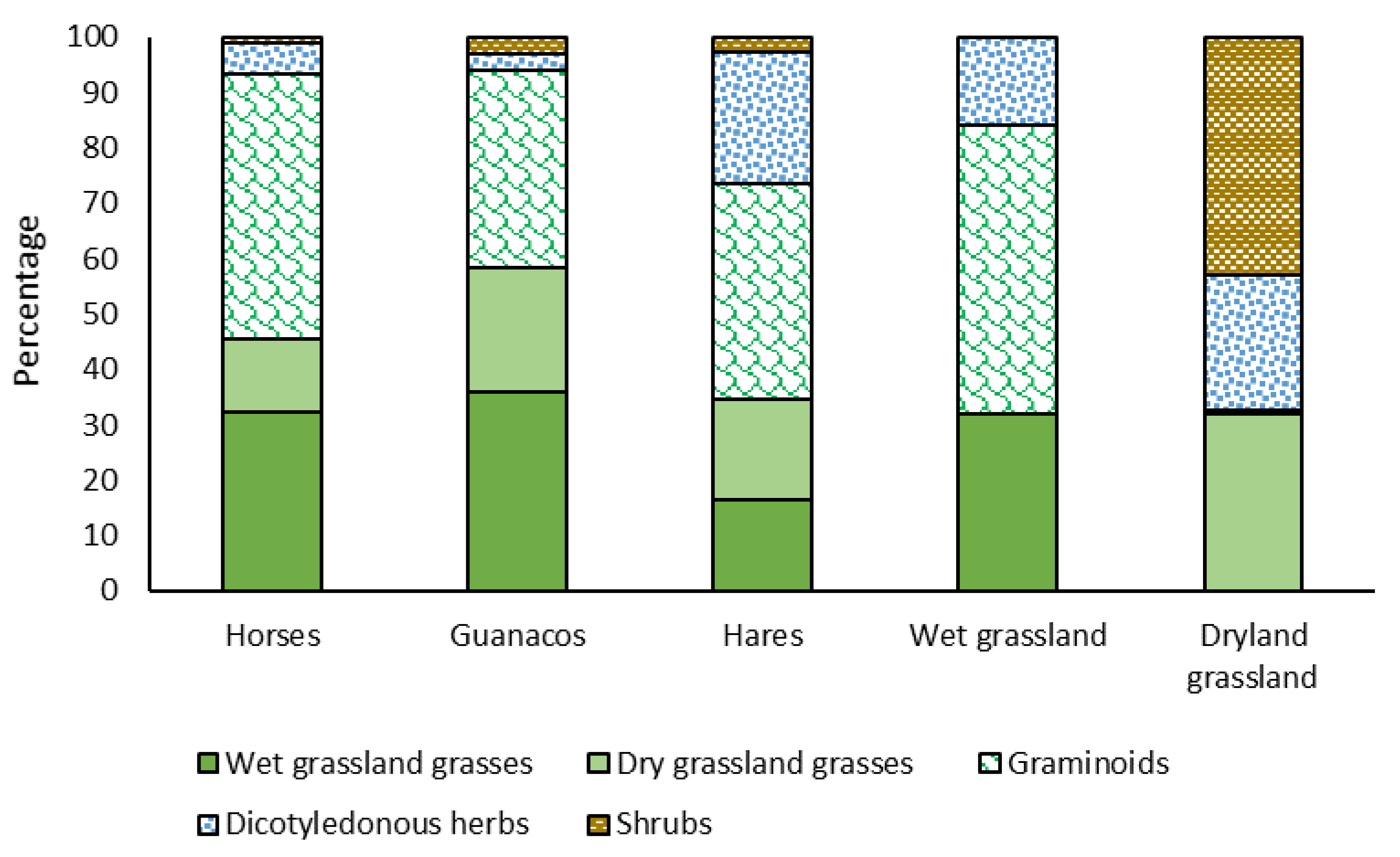
3.2. Diet’s Botanical Composition
3.3. Diet’s Relative Diversity Index
3.4. Diet Overlap
3.5. Selectivity Index of the Main Consumed Plant Species of Wet Grassland
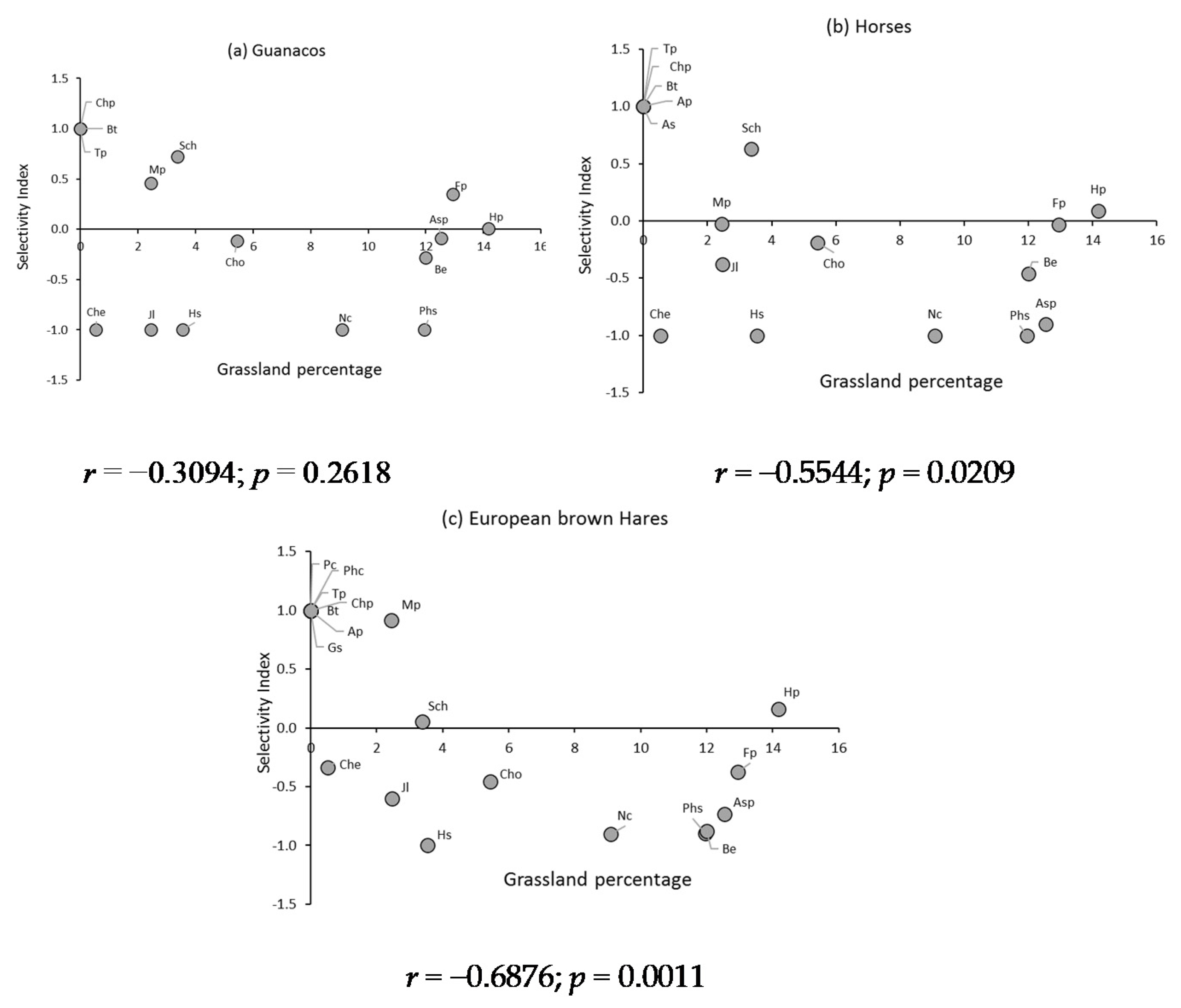
3.6. Selectivity Index of the Primarily Consumed Plant Species of Dryland Grassland
4. Discussion
4.1. Dry Matter Production, Plant Cover and Botanical of the Grasslands
4.2. Diet’s Botanical Composition
4.3. Diet Diversity
4.4. Dietary Overlap
4.5. Selectivity of the Main Plant Species Consumed
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Instituto Nacional de Estadísticas (INE). Estudio 2006; Ganadería Caprina: Limarí y Choapa, Chile, 2006; p. 42. [Google Scholar]
- Castellaro, G.G.C.; Orellana, M.J.P.; Escanilla, C.L.; Freire, M. Caracterización Productiva de Praderas Hidromórficas en la Alta Cordillera de Cuncumén, IV Región de Coquimbo, Chile. II. Valor Pastoral y Nutricional. In Proceedings of the XXIV Congreso de la Asociación Latinoamericana de Producción Animal, XL Congreso Anual Sociedad Chilena de la Producción Animal (SOCHIPA), Puerto Varas, Chile, 9–13 November 2015. [Google Scholar]
- Castellaro, G.G.C.; Orellana, M.J.P.; Escanilla, C. Caracterización productiva de praderas hidromórficas en la alta cordillera de Cuncumén, IV Región de Coquimbo, Chile. I. Producción de materia seca y capacidad de carga. In Proceedings of the XXXVIII Congreso Anual Sociedad Chilena de la Producción Animal (SOCHIPA), Frutillar, Chile, 23–25 October 2013. [Google Scholar]
- Holechek, J.L.; Pieper, R.D.; Herbel, C.H. Range Management, Principles and Practices, 6th ed.; Prentice Hall: Upper Saddle River, NJ, USA, 2011; p. 444. [Google Scholar]
- Bonino, N. Interacción trófica entre el conejo silvestre europeo y el ganado doméstico en el noroeste de la Patagonia Argentina. Ecol. Austral 2006, 16, 135–142. [Google Scholar]
- Bonino, N.; Borrelli, L. Variación estacional en la dieta del conejo silvestre europeo (Oryctolagus cuniculus) en la región andina de Neuquén, Argentina. Ecol. Austral 2006, 16, 7–13. [Google Scholar]
- Borgnia, M.; Vilá, B.L.; Cassini, M.H. Interaction between wild camelids and livestock in an Andean semi-desert. J. Arid Environ. 2008, 72, 2150–2158. [Google Scholar] [CrossRef]
- Vila, A.R.; Borrelli, L.; Martínez, L. Dietary Overlap between Huemul and Livestock in Los Alerces National Park, Argentina. J. Wildl. Manag. 2009, 73, 368–373. [Google Scholar] [CrossRef]
- Johnson, M.K. Foods of primary consumers on cold dessert shrubb-steppe of South Central Idaho. J. Range Manag. 1979, 32, 365–368. [Google Scholar] [CrossRef]
- Vallentine, J.F. Grazing Management, 2th ed.; Academic Press: San Diego, CA, USA, 2001; p. 659. [Google Scholar]
- Gastó, J.; Cosio, F.; Panario, D. Clasificación de Ecorregiones y Determinación de Sitio y Condición. Manual de Aplicación a Municipios y Predios Rurales; Red de Pastizales Andinos: Santiago, Chile, 1993; p. 254. [Google Scholar]
- Köeppen, W. Climatología; Fondo de Cultura Económica: Carretera Picacho-Ajusco, Mexico, 1948; p. 478. [Google Scholar]
- Luebert, F.; Pliscoff, P. Sinopsis Bioclimática y Vegetacional de Chile; Editorial Universitaria: Santiago de, Chile, 2018; p. 384. [Google Scholar]
- Squeo, F.A.; Barry, G.; Warner, B.G.; Aravena, R.; Espinoza, D. Bofedales: High altitude peatlands of the central Andes. Rev. Chil. Hist. Nat. 2006, 79, 245–255. [Google Scholar] [CrossRef] [Green Version]
- Etienne, M.; Prado, C. Descripción de la Vegetación Mediante Cartografía de Ocupación de Tierras. Conceptos y Manual de Uso Práctico. Ciencias Agrícolas Nº10; Universidad de Chile, Facultad de Ciencias Agrarias: Veterinarias y Forestales, Chile, 1982; p. 120. [Google Scholar]
- Daget, P.H.; Poissonet, J. Une méthode d’analyse phytologique des prairies, critères d’application. Ann. Agron. 1971, 22, 5–41. [Google Scholar]
- Passera, C.B.; Dalmasso, A.D.; Borsetto, O. Metodo de “Point Quadrat Modificado”. In Taller de Arbustos Forrajeros para Zonas Áridas y Semiáridas, 2nd ed.; Subcomité Asesor del Árido Subtropical Argentino. Amawald. S. A.: Buenos Aires, Argentina, 1983; p. 107. [Google Scholar]
- Sparks, D.R.; Malechek, J.C. Estimating percentage dry weight in diets using a microscope technique. J. Range Manag. 1968, 21, 264–265. [Google Scholar] [CrossRef] [Green Version]
- Garnick, S.; Barboza, P.S.; Walker, J.W. Assessment of Animal-Based Methods Used for Estimating and Monitoring Rangeland Herbivore Diet Composition. Rangel. Ecol. Manag. 2018, 71, 449–457. [Google Scholar] [CrossRef]
- Catán, A.; Degano, C.A.; Werenitzky, D. Evaluación de criterios de lectura microhistológica para la cuantificación de Sphaeralcea bonariensis (Cav.), Pl Lorentz en mezclas manuales. Tec. Pecu. Méx. 2007, 45, 77–83. [Google Scholar]
- Castellaro, G.; Squella, F.; Ullrich, T.; León, F.; Raggi, A. Algunas técnicas microhistológicas utilizadas en la determinación de la composición botánica de dietas de herbívoros. Agric. Téc. Chile. 2007, 67, 86–93. [Google Scholar] [CrossRef]
- Ortega, I.M.; Berger, M.I.; Flores, M. Manual de Técnica Microhistológica. In IBTA 113/Textos y Manuales 04/Rumiantes Menores (SR-CRSP) 05/; IBTA: La Paz, Bolivia, 1993; p. 48. [Google Scholar]
- Smith, R.L.; Smith, T.M. Ecology, 8th ed.; Addison Wesley Longman: Boston, MA, USA, 2012; p. 683. [Google Scholar]
- Olsen, F.W.; Hansen, R.M. Food relations of wild free-ranging horses to livestock and big game, Red Desert, Wyoming. J. Range Manag. 1977, 30, 17–20. [Google Scholar] [CrossRef] [Green Version]
- Stuth, J.W. Foraging Behavior. Cap; Grazing Management. In An Ecological Perspective; Heitschmidt, R.K., Stuth, J.W., Eds.; Timber Press: Portland, OR, USA, 1991; p. 259. [Google Scholar]
- Krebs, C.J. Ecological Methodology; Harper Collins Publisher: New York, NY, USA, 1989; p. 654. [Google Scholar]
- Kaps, M.; Lamberson, W. Estimation of Parameters; Biostatistics for Animal Science Chap 5; CABI Publishing: Cambridge, MA, USA, 2004; p. 439. [Google Scholar]
- Castellaro, G.; Araya, R. Manejo de Praderas Altiplánicas. Antecedentes, Descripción, Evaluación y Manejo; Fundación para la Innovación Agraria: Santiago, Chile, 2012; p. 65. [Google Scholar]
- Le Houérou, H.N. Rain use efficiency a unifying concept in arid land ecology. J. Arid Environ. 1984, 7, 213–247. [Google Scholar]
- Castellaro, G.; Araya, R.; Escanilla, J.P. Buenas Prácticas para la Actividad Ganadera. Capítulo 2.2. In Manual de Buenas Prácticas para Uso Sustentable de Ecosistemas de Montaña; Fredericksen, N., Pérez, J., Contreras, X., Eds.; Universidad de Chile–Servicio Agrícola y Ganadero: Santiago, Chile, 2011; p. 117. [Google Scholar]
- Ormaechea, S.G.; Utrilla, V.R.; Suarez, D.D.; Peri, P.L. Evaluación objetiva de la condición de mallines de Santa Cruz. Prod. Anim. 2010, 12, 47–52. [Google Scholar]
- Gamound, M. Rain use efficiency, primary production and rainfall relationships in desert rangelands of Tunisia. Land Degrad. Develop. 2016, 27, 738–747. [Google Scholar] [CrossRef]
- Baldi, R.; Pelliza-Sbriller, A.; Elston, D.; Albon, S. High potential for competition between guanacos and sheep in Patagonia. J. Wildl. Manag. 2004, 68, 924–938. [Google Scholar] [CrossRef]
- Arias, N.; Feijóo, S.; Quinteros, P.; Bava, J. Composición botánica de la dieta del guanaco (Lama guanicoe) en la Reserva Corazón de la Isla, Tierra del Fuego (Argentina): Utilización estacional de Nothofagus spp. Bosque 2015, 36, 71–79. [Google Scholar] [CrossRef] [Green Version]
- Puig, S.; Videla, F.; Cona, M.; Monge, S. Use of food availability by guanacos (Lama guanicoe) and livestock in Northern Patagonia (Mendoza, Argentina). J. Arid Environ. 2001, 47, 291–308. [Google Scholar] [CrossRef]
- National Reseach Council (NRC). Nutrient Requirements of Small Ruminants. Sheep, Goats, Cervids, and New World Camelids, 6th ed.; National Academic Press: Washington, DC, USA, 2007; p. 362. [Google Scholar]
- Muñoz, A.E.; Simonetti, J.A. Diet of guanaco in sheep-free rangeland in Tierra del Fuego, Chile. Cienc. Inv. Agr. 2013, 40, 185–191. [Google Scholar] [CrossRef] [Green Version]
- Bonino, N.; Pelliza-Sbriller, A. Composición botánica de la dieta del guanaco (Lama guanicoe) en dos ambientes contrastantes de Tierra del Fuego, Argentina. Ecol. Austral 1991, 1, 97–102. [Google Scholar]
- Barri, F.R.; Falczuk, V.; Cingolani, A.M.; Díaz, S. Dieta de la población de guanacos (Lama guanicoe) reintroducida en el Parque Nacional Quebrada del Condorito, Argentina. Ecol. Austral 2014, 24, 203–211. [Google Scholar] [CrossRef]
- Candia, R.; Dalmasso, A.D. Dieta del guanaco (Lama guanicoe) y productividad del pastizal en la reserva La Payunia, Mendoza (Argentina). Multequina 1995, 4, 5–15. [Google Scholar]
- Castellaro, G.; Ullrich, T.; Wackwitz, B.; Raggi, A. Composición botánica de la dieta de alpacas (Lama pacos L.) y llamas (Lama glama L.) en dos estaciones del año, en praderas altiplánicas de un sector de la Provincia de Parinacota. Agric. Téc. 2004, 64, 353–364. [Google Scholar] [CrossRef]
- Soler, R.M.; Martínez-Pastur, G.; Lencinas, M.V.; Borrelli, L. Seasonal diet of Lama guanicoe (Camelidae: Artiodactyla) in a heterogeneous landscape of South Patagonia. Bosque 2013, 34, 129–141. [Google Scholar] [CrossRef] [Green Version]
- Karmiris, I.; Panagiotis, D.P.; Kazantzidis, S.; Papachristou, T.G. Diet Selection by Domestic and Wild Herbivore Species in a Coastal Mediterranean Wetland. Ann. Zool. Fenn. 2011, 48, 233–242. [Google Scholar] [CrossRef]
- Osoro, K.; Ferreira, M.; García, U.; Martínez, A.; Celaya, R. Forage intake, digestibility and performance of cattle, horses, sheep and goats grazing together on an improved heathland. Anim. Prod. Sci. 2015, 57, 102–109. [Google Scholar] [CrossRef]
- Scasta, J.D.; Beck, J.L.; Angwin, C.J. Meta-Analysis of Diet Composition and Potential Conflict of Wild Horses with Livestock and Wild Ungulates on Western Rangelands of North America. Rangel. Ecol. Manag. 2016, 69, 10–318. [Google Scholar] [CrossRef]
- Fleurance, G.; Duncan, P.; Mallevaud, B. Daily intake and the selection of feeding sites by horses in heterogeneous wet grasslands. Anim. Res. 2001, 50, 149–156. [Google Scholar] [CrossRef] [Green Version]
- Cheeke, P.R.; Dierenfeld, E.S. Comparative Animal Nutrition and Metabolism; CABI Org.: Oxford, UK, 2010; p. 339. [Google Scholar]
- Dryden, G.M. Animal Nutrition Science; CABI: Oxfordshire, UK, 2008; p. 415. [Google Scholar]
- Smith, C.; Valdez, R.; Holechek, J.L.; Zwank, P.J.; Cardenas, M. Diets of native and non-native ungulates in southcentral New Mexico. Southwest. Nat. 1998, 43, 163–169. [Google Scholar]
- Reichlin, T.; Klansek, E.; Hackländer, K. Diet selection by hares (Lepus europaeus) in arable land and its implication for habitat management. Eur. J. Wildl. Res. 2006, 52, 109–118. [Google Scholar] [CrossRef]
- López-Cortés, F.; Cortés, A.; Miranda, E.; Rau, J.R. Dietas de Abrothrix andinus, Phyllotis xanthopygus (Rodentia) y Lepus europaeus (Lagomorpha) en un ambiente altoandino de Chile. Rev. Chil. Hist. Nat. 2007, 80, 3–12. [Google Scholar] [CrossRef]
- Katona, K.; Bíró, Z.; Hahn, I.; Kertész, M.; Altbäcker, V. Competition between European hare and European rabbit in a lowland área, Hungary: A long-term ecological study in the period of rabbit extinction. Folia Zool. 2004, 53, 255–268. [Google Scholar]
- Puig, S.; Videla, F.; Cona, M.I.; Monge, S.A. Diet of the brown hare (Lepus europaeus) and food availability in northern Patagonia (Mendoza, Argentina). Mamm. Biol. 2006. [Google Scholar] [CrossRef]
- Wolff, J.O. Food Habits of Snowshoe Hares in Interior Alaska. J. Wildl. Manag. 1978, 42, 148–153. [Google Scholar] [CrossRef]
- McInnis, M.L.; Vavra, M. Dietary Relationships among Feral Horses, Cattle, and Pronghorn in Southeastern Oregon. J. Range Manag. 1987, 40, 60–66. [Google Scholar] [CrossRef]
- Krysl, L.J.; Sowell, B.F.; Hubbert, M.E.; Plumb, G.E.; Jewett, T.K.; Smith, M.A.; Waggoner, J.W. Horses and Cattle Grazing in the Wyoming Red Desert, II. Dietary Quality. J. Range Manag. 1984, 37, 252–256. [Google Scholar] [CrossRef]
- Hosten, P.E.; Whitridge, H.; Broyles, M. VDiet Overlap and Social Interactions among Cattle, Horses, Deer and Elk in the Cascade-Siskiyou National Monument, Southwest. Oregon. Department of the Interior, Bureau of Land Management, US Medford. 2007. Available online: https://www.blm.gov/or/districts/medford/files/dietoverlap.pdf (accessed on 5 October 2020).
- Uresk, D.W. Diets of the Black-tailed Hare in Steppe Vegetation. J. Range Manag. 1978, 31, 439–442. [Google Scholar] [CrossRef] [Green Version]
- Castellaro, G.; Urra, H.; Hidalgo, J.; Orellana, C.; Escanilla, J.P. Sheep and goat grazing diets on an annual Mediterranean grassland containing tall wheatgrass (Thinopyrum ponticum (PODP.)). Cien. Inv. Agr. 2018, 45, 240–250. [Google Scholar] [CrossRef]
- Schai-Braun, S.C.; Reichlin, T.S.; Ruf, T.; Klansek, E.; Tataruch, F.; Arnold, W. The European Hare (Lepus europaeus): A Picky Herbivore Searching for Plant Parts Rich in Fat. PLoS ONE 2015, 10, e0134278. [Google Scholar] [CrossRef] [Green Version]
- Reus, M.L.; Peco, B.; De los Ríos, C.; Giannoni, S.M.; Campos, C.M. Trophic interactions between two medium-sized mammals: The case of the native Dolichotis patagonum and the exotic Lepus europaeus in a hyper-arid ecosystem. Acta Theriol. 2013, 58, 205–214. [Google Scholar] [CrossRef]
- Reus, M.L.; Cappa, F.M.; Andinoa, N.; Campos, V.E.; De los Ríos, C.; Campos, C.M. Trophic interactions between the native guanaco (Lama guanicoe) and the exotic donkey (Equus asinus) in the hyper-arid Monte desert (Ischigualasto Park, Argentina). Stud. oNeotrop. Fauna Environ. 2014, 49, 159–168. [Google Scholar] [CrossRef]
- Linares, L.; Linares, V.; Mendoza, G.; Peláez, F.; Rodríguez, E.; Phum, C. Preferencias alimenticias del guanaco (Lama guanicoe cacsilensis) y su competencia con el ganado doméstico en la Reserva Nacional de Calipuy, Perú. Sci. Agropecu. 2010, 1, 225–234. [Google Scholar] [CrossRef]
- Kuzyk, G.W.; Hudson, R.J. Animal-unit equivalence of bison, wapiti, and mule deer in the aspen parkland of Alberta. Can. J. Zool. 2007, 85, 767–773. [Google Scholar] [CrossRef]
- Kleiber, M. Body size and metabolic rate. Physiol. Rev. 1947, 27, 511–541. [Google Scholar] [CrossRef]
- Hobbs, N.T.; Carpenter, L.H. Viewpoint: Animal-Unit Equivalents Should Be Weighted by Dietary Differences. J. Range Manag. 1986, 39, 470. [Google Scholar] [CrossRef]
- Bull, J.; Bas, F.; Silva-Guzmán, M.; Wentzel, H.H.; Keim, J.P.; Gandarillas, M. Characterization of Feeding, Sport Management, and Routine Care of the Chilean Corralero Horse during Rodeo Season. Animals 2019, 9, 697. [Google Scholar] [CrossRef] [Green Version]
- De Lamo, D.A. South American Camelids in Argentina. History, Use, and Animal Health. Servicio Nacional de Sanidad y Calidad Agroalimentaria (SENASA); Ministerio de Agricultura, Ganadería y Pesca: Buenos Aires, Argentina, 2011; p. 37.
- Frylestam, B. Growth and Body Weight of European Hares in Southern Sweden. Holarctic Ecol. 1980, 3, 81–86. [Google Scholar] [CrossRef]
- Viviano, A.; Mori, E.; Fattorini, E.; Mazza, G.; Lazzeri, L.; Panichi, A.; Strianese, L.; Mohamed, W.F. Spatiotemporal overlap between the European brown hare and its potential predators and competitors. Animals 2021, 11, 562. [Google Scholar] [CrossRef]
- Tirado, C.; Cortés, A.; Miranda–Urbina, E.; Carretero, M.A. Trophic preferences in an assemblage of mammal herbivores from Andean Puna (Northern Chile). J. Arid. Environ. 2012, 79, 8–12. [Google Scholar] [CrossRef]
- Castellaro, G.; Orellana, C.; Escanilla, J.P.; Bastías, C.; Cerpa, P.; Raggi, L. Botanical Composition and Diet Quality of the Vicuñas (Vicugna vicugna Mol.) in Highland Range of Parinacota, Chile. Animals 2020, 10, 1205. [Google Scholar] [CrossRef]
- Provenza, F.D.; Villalba, J.J.; Dziba, L.E.; Atwood, S.B.; Banner, R.E. Linking herbivore experience, varied diets and plant biochemical diversity. Small Rumin. Res. 2003, 49, 257–274. [Google Scholar] [CrossRef]
- Stamps, J.; Tanaka, S.; Krishnan, V. The relationship between selectivity and food abundance in a juvenile lizard. Ecology 1981, 64, 1079–1092. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Season | Herbivore Species | Mean | ||
|---|---|---|---|---|
| Horses | Guanacos | European Brown Hares | ||
| 2011 | 0.797 ± 0.028 | 0.824 ± 0.033 | 0.737 ± 0.037 | 0.786 ± 0.019 |
| 2012 | 0.749 ± 0.026 | 0.759 ± 0.052 | 0.722 ± 0.026 | 0.743 ± 0.021 |
| Mean | 0.773 ± 0.019 | 0.791 ± 0.031 | 0.730 ± 0.022 | |
| Herbivore Species | Herbivore Species | ||
|---|---|---|---|
| Horses | Guanacos | European Brown Hares | |
| Horses | - | 0.5571 | 0.4822 |
| Guanacos | 0.5571 | - | 0.4995 |
| European brown hares | 0.4822 | 0.4995 | - |
| Herbivore Species | Wet Grassland | Dryland Grassland |
|---|---|---|
| 0.0467 | 0.5341 | |
| Guanacos | n = 41 | n = 23 |
| p = 0.7676 | p = 0.0122 | |
| 0.2030 | 0.3650 | |
| Horses | n = 41 | n = 23 |
| p = 0.1991 | p = 0.0869 | |
| 0.0801 | 0.5734 | |
| European brown hares | n = 41 | n = 23 |
| p = 0.6126 | p = 0.0072 |
| Herbivore Species | Livestock Equivalences 1 | ||
|---|---|---|---|
| Horses | Guanaco | European Brown Hares | |
| Horses (384 kg liveweight) | - | 5.08 | 61.32 |
| Guanaco (96 kg liveweight) | 0.20 | - | 20.93 |
| European brown hares (4.2 kg liveweight) | 0.02 | 0.05 | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Castellaro, G.; Orellana, C.L.; Escanilla, J.P. Summer Diet of Horses (Equus ferus caballus Linn.), Guanacos (Lama guanicoe Müller), and European Brown Hares (Lepus europaeus Pallas) in the High Andean Range of the Coquimbo Region, Chile. Animals 2021, 11, 1313. https://doi.org/10.3390/ani11051313
Castellaro G, Orellana CL, Escanilla JP. Summer Diet of Horses (Equus ferus caballus Linn.), Guanacos (Lama guanicoe Müller), and European Brown Hares (Lepus europaeus Pallas) in the High Andean Range of the Coquimbo Region, Chile. Animals. 2021; 11(5):1313. https://doi.org/10.3390/ani11051313
Chicago/Turabian StyleCastellaro, Giorgio, Carla Loreto Orellana, and Juan Pablo Escanilla. 2021. "Summer Diet of Horses (Equus ferus caballus Linn.), Guanacos (Lama guanicoe Müller), and European Brown Hares (Lepus europaeus Pallas) in the High Andean Range of the Coquimbo Region, Chile" Animals 11, no. 5: 1313. https://doi.org/10.3390/ani11051313