
Abstract
The contingent capture account of involuntary attention claims that it is guided by top-down factors, such as volitional goals or task instructions. The contrasting rapid disengagement account holds that the contingent capture account relies on the spatial precueing paradigm, which is vulnerable to the elimination of the cue-validity effect through rapid attentional disengagement. In the present study, five experiments were conducted to examine whether a spatial cue presented in a target-defining or distractor-defining color that predicted the location of a subsequently presented target at the chance level involuntarily captures attention by measuring the cue-validity effect. Additionally, to examine the influence of cue–target compatibility as an alternative indicator of attentional capture, an object identical to or different from the target object was presented at the cued location in the cue display in all experiments. The results showed that the cue-validity effect and the cue–target compatibility effect were present only when the target-color cue was presented. The object of the target display presented at the location cued by the target color was recognized even on invalid trials. By contrast, the distractor color cue did not show any indication of attentional capture or postattentive inhibition. These results imply that preattentive selection and postattentive inhibition depend on top-down attentional control setting. Furthermore, the absence of a cue-validity effect with a distractor feature is not due to the inhibition of the cued location after attentional disengagement.
Similar content being viewed by others

Despite their effort to pay attention to stimuli relevant for achieving goals at hand, humans often are distracted by goal-irrelevant stimuli. This involuntary attentional shift to an external event is known as attentional capture or exogenous attentional orienting (Posner, 1980; Posner & Cohen, 1984). Many researchers have investigated how some objects capture attention involuntarily for priority processing while others are left unattended (e.g., Folk et al., 1992; Jonides & Yantis, 1988; Theeuwes, 1991). The salience-driven account of attentional capture argues that attentional capture occurs based on the feature discontinuity (salience) of objects calculated during a preattentive processing stage, in which the basic properties of objects are rapidly processed in a parallel manner (e.g., Theeuwes, 1991, 1992, 1994; Yantis & Jonides, 1990). Consistent with this view, some researchers have suggested that the allocation of spatial attention is simply the process of serially checking locations that were previously labeled as salient (Sagi & Julesz, 1985; Ullman, 1984). Proponents of the salience-driven attentional capture account utilize an experimental procedure called the additional-singleton paradigm, in which researchers instruct participants to locate a target that is defined by color or shape and to respond to the object inside it. For example, Theeuwes (1992) had participants search for a green circle (color singleton) surrounded by red circles. Occasionally, one of the red circles was replaced with a red diamond distractor (shape singleton). The presentation of the diamond distractor interfered with searching for the target object despite the shape’s irrelevance to the task, indicating attentional capture.
In contrast with the salience-driven account, the contingent capture account suggests that the allocation of exogenous attention is guided by top-down attentional control settings, which are established with a volitional goal or a task instruction (Folk et al., 1992). In this account, an attentional control setting, which contains mental descriptions of features of interest, has more control over exogenous attention than does any feature discontinuity or salience (e.g., Folk et al., 1994; Folk et al., 2002; Folk & Remington, 1998, 2006; Pratt & Hommel, 2003; Remington et al., 2001). In Folk and Remington’s (1998) experiments with the spatial cueing paradigm, participants responded to the identity of a target object defined by color, presented in one of four possible locations. In each of the four locations, a uniquely colored object appeared (e.g., one green object, one red object, and two white filler objects). A noninformative red or green cue presented in one of the four locations preceded the target display. If the color of the cue matched the color of the target object, a significant reaction time (RT) cost occurred when the target object appeared at an uncued (invalid) location relative to when the target appeared at the cued (valid) location, resulting in what is known as the cue-validity effect. In contrast, when the cue color differed from the target color, no cue-validity effect occurred, which is consistent with the hypothesis that the allocation of exogenous attention is contingent upon attentional control setting.
Although significant cue-validity effects consistently result from using target-feature cues and not from nontarget-feature cues, it is possible that the temporal gap between the onsets of the cue and the target obscures the evidence of attentional capture by a nontarget-feature cue because, in most spatial cueing experiments, the target appears after the cue (Belopolsky et al., 2010). According to Belopolsky et al.’s (2010) rapid disengagement account, initially proposed by Theeuwes et al. (2000), a singleton color cue can involuntarily capture attention, regardless of its task relevance. However, when a nontarget-feature cue is presented, attention rapidly disengages from the cued location after attentional capture because it does not match the features specified in the attentional control setting, resulting in no cue-validity effect. Moreover, Belopolsky et al. showed that an initially attended location is inhibited after attentional disengagement from the cued location when the attentional control is strong enough. They employed a spatial cueing task in conjunction with a “go/no-go” task, in which they informed participants of a specific target domain (onset or color singleton) at the beginning of each trial. They instructed participants to withhold their response when the presented target domain did not belong to the domain specified in advance. The researchers found that the mean RT was greater for valid trials than for invalid ones when they presented a no-go feature cue (Experiments 3 & 4). They contend that the location cued by a no-go feature was inhibited when participants disengaged their attention from the cued location. Therefore, it took more time to respond to a target presented at the inhibited location than to one presented at a different location, which resulted in a reversed cue-validity effect.
The rapid disengagement account attributes this reversed cue-validity effect to the inhibition of the cued location after attentional disengagement (Belopolsky et al., 2010), but the capture-independent inhibition account, which is based on the contingent capture account, suggests that the effect reflects a successful top-down inhibition rooted in the response selection stage, which is independent of any attentional capture mechanism (Anderson & Folk, 2012). Anderson and Folk (2012) disagree with the rapid disengagement account’s claim that attention rapidly disengages from the location cued by a nontarget salient feature even when the cue successfully predicts the target location. They demonstrated that inhibition could occur at a location cued by nontarget features without attentional disengagement. In their study, participants performed a go/no-go task incorporated into a spatial cueing paradigm, responding to a “go” color singleton target and withholding their response to a “no-go” color singleton target (Experiments 4, 5, & 6). Both the contingent capture account and the salience-driven capture account posit that a singleton cue can capture attention regardless of the attentional control setting when the target is defined by a singleton (Anderson & Folk, 2012; Folk & Anderson, 2010). Therefore, a go/no-go task with a singleton target made it possible to examine whether a task-relevant feature associated with a no-go response would capture attention. The results showed that inhibition occurred at the location occupied by the no-go color cue, even when attentional disengagement from the cued location was not necessary because the target was subsequently presented at the same location. This finding is inconsistent with the main argument of the rapid disengagement account.
Present study
In this study, we performed a finer-grained search for evidence of attentional capture by adopting a spatial cueing paradigm to unveil traces of attentional capture, which may be clouded by rapid disengagement from spatial cues before the onset of subsequent targets. To this end, we instructed participants to identify a target defined by a specific color (e.g., red) and to ignore a distractor of another color (e.g., green), presented along with two white filler objects. At the onset of the target display, a spatial color cue primed one of four possible target locations (target-defining color or distractor color). Moreover, to determine whether the absence of the cue-validity effect necessarily indicates that the cued location was unattended or inhibited, we manipulated the compatibility between the cued object in the cue display and the target in the target display, as performed by Folk and Remington (1998, 2006). Specifically, one of two different objects that either matched (compatible) or did not match (incompatible) the target object appeared inside each placeholder of the cue display. This manipulation assumes that attentional allocation to the cued object affects responses to the target object (Remington et al., 2001; Shih & Sperling, 1996). In Folk and Remington’s (2006) spatial cueing experiments, letter characters (“x” or “=”) appeared inside all placeholders in the cue display and the target display. A significant cue–target compatibility effect occurred only with the presentation of a target color cue, indicating that the letter cued by the distractor color was unattended. This study adopted the same approach but without the visual noise masks presented between the cue and target displays to obtain more robust cue–target compatibility effects.
By design, this study enabled the comparison of predictions drawn from the rapid disengagement and contingent capture accounts. The former predicts that the distractor color cue will elicit either no cue-validity effect or a reversed one, depending on the speed of attentional disengagement from the cued location. Nevertheless, upon the allocation of attention to the cued location, participants process and recognize the object at the cued location, regardless of any attentional disengagement or inhibition of the cued location. Thus, we expected a significant cue–target compatibility effect if the distractor color cue captured attention, even though attentional disengagement possibly clouded or reversed the cue-validity effect elicited by the distractor color cue. In contrast, the latter account predicts neither a cue–target compatibility effect nor a cue-validity effect with the distractor color cue because top-down attentional control settings govern the involuntary allocation of attention.
Experiment 1
Experiment 1 examined whether a distractor color cue captures attention by using the spatial cueing paradigm. To this end, vertically or horizontally oriented bars appeared within all four placeholders in both the cue and target displays. Accordingly, the orientation of the target object matched that of the cued object for half of the trials, but not for the other half. Participants responded to the target color bar in the target display, which contained a bar of the target-defining color, a bar of the distractor color, and two other white bars. Before the target display, the target-defining color or the distractor color cued one of the four placeholders. If a color cue were to involuntarily capture attention based on its status as a singleton, a significant cue-validity effect would occur regardless of task-relevance, as the salience-driven attentional capture account suggests. Moreover, if attention is allocated to a location cued by the distractor color, the object in the cued location should be recognized, resulting in cue–target compatibility effect. However, if the color cue captures attention only when it has a target-defining feature, as the contingent capture account suggests, a cue-validity effect would occur with the target color cue but not with the distractor color cue. Also, the cue–target compatibility effect would be evident only with presentation of the target color cue.
Method
Participants
To determine the optimal number of participants, we conducted a power analysis using the G*Power software (Faul et al., 2009), with power (1 − β) set to .95 and the alpha level set to 5%. The analysis indicated that a minimum sample size of 12 was required to detect an effect size of \( {\eta}_p^2 \) = .25, which was the effect size estimated based on the F value of the interaction of cue congruency and cue validity in Folk and Remington’s (2006) Experiment 1. For the experiment, we recruited 20 undergraduate students from Korea University (ages 21 to 27) to ensure sufficient power to detect attentional capture by distractor color cues. All participants reported having normal or corrected-to-normal visual acuity and color vision. We provided a reward of 6,000 KRW (approximately 5 USD) for participation in the study. This study was approved by the institutional review board at Korea University (KU-IRB-15-25-P-1).
Apparatus
A 17-inch CRT monitor displayed all stimuli at a viewing distance of approximately 60 cm. We used E-Prime software (Version 1.2, Psychology Software Tools, Pittsburgh, PA) to present stimuli and record participants’ responses with the “f” and “j” keys on a standard computer keyboard.
Stimuli
Each trial consisted of a placeholder display, a cue display, and a target display. The placeholder display contained a fixation cross (0.78° of visual angle) at the center of the display and four peripheral square placeholders (2.08° × 2.08°) located at the upper, lower, left, and right sides of the fixation cross. Each placeholder was located approximately 2.10° from the fixation cross. The fixation cross and the placeholders were white (RGB: 255, 255, 255; CIE: x = .31, y = .33). The cue display also contained a fixation cross and four placeholders in the same locations and of the same color. However, in the cue display, a set of four small dots (0.39° in diameter) surrounded each of the four placeholders. One set of dots was either red (RGB: 255, 0, 0; CIE: x = .64, y = .33) or green (RGB: 13, 147, 13; CIE: x = .30, y = .55), and the other sets were white. There was either a horizontally or vertically oriented white bar (1.3° wide, 0.12° thick) within the placeholder. A horizontally or vertically oriented bar appeared inside all placeholders in the target display and in the cue display, as well. In the target display, one of the four bars was red, another was green, and the other two were white. All placeholders were white.
Design and procedure
The experiment began with a practice block of 20 trials followed by four experimental blocks, each of which comprised 128 trials. The cue predicted the upcoming target location with 25% probability (i.e., the level of chance). The two types of cues appeared equally often. Moreover, each bar that appeared inside the placeholders of the cue and target displays was randomly chosen, and all appeared equally often.
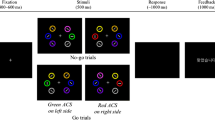
Figure 1 displays the trial sequence of the experiment. Each trial began with the placeholder display for 1,000 ms, followed by a 50-ms cue display. After the offset of the cue display, the placeholder display appeared again for 100 ms. The target display was presented for 50 ms. We instructed participants to focus on the fixation cross and to respond to the orientation of the target color bar as rapidly and accurately as possible when the target was on the screen. They responded by pressing the “f” key with their left index finger for a target-colored, vertically oriented bar and the “j” key with their right index finger for a target-colored, horizontally oriented bar. We fully informed participants of the relationship between the cue and the target and instructed them to ignore the cue and the distractor. A 150-ms tone followed all incorrect responses or responses occurring after more than 1,500 ms.

Example of a trial sequence in Experiment 1
Results
Response times (RTs) shorter than 130 ms and longer than 1,300 ms were excluded as outliers (0.18%). The mean correct RT and percentage of error (PE) were calculated for each participant as a function of cue congruency (target color or distractor color), cue validity (valid or invalid), and cue–target compatibility (compatible or incompatible). Repeated-measures analyses of variance (ANOVAs) were conducted on the mean correct RT and PE data with the above variables as within-subject variables (see Fig. 2).

Mean RT and PE as a function of congruency, validity, and compatibility in Experiment 1. The error bar indicates within-subject standard error
RT
The main effect of cue congruency was significant, F(1, 19) = 15.7, p < .001, MSe = 174, ηp2 = .45. The mean RT was greater when the target color cue was presented (M = 541 ms) than when the distractor color cue was presented (M = 533 ms). The main effect of cue validity was also significant, F(1, 19) = 26.4, p < .0001, MSe = 390, ηp2 = .58. The mean RT was shorter for valid trials (M = 529 ms) than for invalid ones (M = 545 ms), indicating a 16-ms cue-validity effect. The main effect of cue–target compatibility was significant, F(1, 19) = 4.66, p < .05, MSe = 359, ηp2 = .20. The mean RT was 6 ms shorter for cue–target compatible trials (M = 534 ms) than for incompatible ones (M = 540 ms).
Cue validity interacted with cue congruency, F(1, 19) = 85.2, p < .0001, MSe = 148, ηp2 = .82. Further analyses demonstrated that a cue-validity effect occurred when the target color cue was presented (34 ms), F(1, 19) = 63, p < .0001, MSe = 182, ηp2 = .77, but not when the distractor color cue was presented (−2 ms), F(1, 19) < 1, p = .56. Cue–target compatibility interacted with cue validity, F(1, 19) = 9.63, p < .01, MSe = 214, ηp2 = .34. Further analyses at each cue validity level showed that a significant cue–target compatibility effect occurred when the target was presented at the cued location, F(1, 19) = 9.83, p < .01, MSe = 189, ηp2 = .34, but not when it was presented at an uncued location, F(1, 19) < 1, p = .82. Cue–target compatibility interacted with cue congruency, F(1, 19) = 11.5, p < .01, MSe = 234, ηp2 = .38. Further analyses at each cue congruency level revealed a significant, 15-ms cue–target compatibility effect for congruent trials, F(1, 19) = 10.7, p < .01, MSe = 202, ηp2 = .36, but not for incongruent ones, F(1, 19) < 1, p = .58.
The three-way interaction of cue congruency, cue validity, and cue–target compatibility was also significant, F(1, 19) = 15.4, p < .001, MSe = 142, ηp2 = .45. A significant cue–target compatibility effect occurred only when the target appeared at the location cued by the target color, F(1, 19) = 23, p < .0001, MSe = 372, ηp2 = .55. Cue–target compatibility was not significant when the target appeared at a location other than that cued by the target color, F(1, 19) < 1, p = .98; at the location cued by the distractor color, F(1, 19) < 1, p = .74; or at a location other than that cued by the distractor color, F(1, 19) < 1, p = .6.
PE
No main effects or interactions reached significance.
Discussion
When the target color cue was presented, a significant 34-ms cue-validity effect occurred. Moreover, a significant cue–target compatibility effect occurred for valid trials, indicating attentional capture by the target color cue. Importantly, no cue–target compatibility effect occurred for invalid trials when the target cue was presented. This lack of a compatibility effect on invalid trials was possibly due to forward visual masking, or the contribution of the compatibility between a target display object at the cued location and the target, as the cue and the target appeared in different locations on invalid trials.
In contrast, when the distractor color cue was presented, no sign of attentional capture emerged, which is inconsistent with Belopolsky et al.’s (2010) rapid disengagement account. Specifically, the cue-validity effect was not significant, and no cue–target compatibility effect occurred, regardless of whether the target was presented at the cued location or an uncued location. If attention rapidly disengaged from the location cued by the distractor color, as Belopolsky et al. have suggested, a reversed cue-validity effect should have resulted.
Experiment 2
Experiment 2 investigated attentional capture by a distractor color cue in a task requiring enhanced attentional control. Belopolsky et al. (2010) argue that the occurrence of postattentive suppression depends on the robustness of top-down attentional control. That is, when the attentional control setting is not sufficiently strong, attention captured by a nontarget feature is unable to disengage rapidly from the initially attended location, resulting in a cue-validity effect. In contrast, when enhanced attentional control is required to perform a task, attention can disengage rapidly from a location occupied by a nontarget feature, and top-down modulated inhibition is induced after attentional disengagement from the cued location.
In Experiment 1, participants searched for a target bar defined by color and discerned its orientation, which required less attentional resources because it was easy to distinguish vertically oriented bars from horizontally oriented ones. Hence, the attentional control setting might have been too weak to induce postattentive suppression where the distractor color cue was attended. Therefore, in Experiment 2, we utilized the upright capital English letters “T” and “L” instead of bars to make it more difficult to distinguish the target shapes. This design was intended to encourage participants to adopt a stronger attentional-control setting. If more robust attentional control elicits postattentive suppression, as Belopolsky et al. (2010) have suggested, a reversed cue-validity effect should occur for the distractor color cue. In contrast, if the location of the distractor color cue is unattended, as the contingent capture account suggests, neither a cue-validity effect nor a cue–target compatibility effect would result when the distractor color cue was presented.
Method
Participants
We recruited a new group of 20 undergraduate students (ages 18 to 29) from the same participant pool used in Experiment 1. As before, participants received 6,000 KRW in compensation, and all reported normal or corrected-to-normal visual acuity and color vision.
Apparatus and stimuli
The apparatus and stimuli were the same as those used in Experiment 1, with one exception. We replaced the bars in the cue and target displays with the white upright letters “T” (0.48° × 0.66°) and “L” (0.42° × 0.66°). In the target display, one target-colored, one distractor-colored, and two white Ts and Ls were randomly assigned to each of the four placeholders.
Design and procedure
The design and procedure of Experiment 2 were identical to those of Experiment 1. The participants responded only to the target-colored letter, ignoring the distractor-colored letters. They pressed the “f” key with their left index finger in response to the target letter “T” and the “j” key with their right index finger in response to the target letter “L.” We emphasized the importance of accuracy and speed, as well.
Results
Adopting the same criteria used in Experiment 1, we excluded 0.56% of all trials from the analyses as outliers. The mean correct RT and PE were calculated for each participant as a function of cue congruency (target color or distractor color), cue validity (valid or invalid), and cue–target compatibility (compatible or incompatible). ANOVAs were conducted on the mean correct RTs and PEs, with the above factors as within-subject variables (see Fig. 3).

Mean RT and PE as a function of congruency, validity, and compatibility in Experiment 2
RT
As in Experiment 1, the main effect of cue congruency was significant, F(1, 19) = 23, p < .0001, MSe = 367, ηp2 = .55. The mean RT was greater for congruent trials (M = 546 ms) than for incongruent trials (M = 532 ms). The main effect of cue validity was also significant, F(1, 19) = 5.64, p < .05, MSe = 614, ηp2 = .23. The mean RT was greater for invalid trials (M = 544 ms) than for valid trials (M = 534 ms), indicating a 10-ms cue-validity effect. However, the main effect of cue–target compatibility was not significant, F(1, 19) = 3.59, p = .074.
Cue validity interacted with cue congruency, F(1, 19) = 15.7, p < .001, MSe = 269, ηp2 = .45. Further analyses of the cue-validity effect at each level of cue congruency revealed a significant 20-ms cue-validity effect when the target color cue was presented, F(1, 19) = 12.6, p < .01, MSe = 305, ηp2 = 40, but not when the distractor color cue was presented, F(1, 19) < 1, p = .79. As in Experiment 1, cue validity interacted with cue–target compatibility, F(1, 19) = 7.43, p < .05, MSe = 339, ηp2 = .28. Further analyses revealed a significant cue–target compatibility effect of 14 ms for valid trials, F(1, 19) = 6.92, p < .05, MSe = 268, ηp2 = .27, but none for invalid trials, F(1, 19) < 1, p = .44. The interaction between cue congruency and cue–target compatibility was not significant, F(1, 19) < 1, p = .36.
The three-way interaction of cue congruency, cue validity, and cue–target compatibility was not significant, F(1, 19) = 1.24, p = .28; however, a cue–target compatibility effect occurred when the target appeared at the location cued by the target color (20 ms), F(1, 19) = 7.08, p < .05, MSe = 543, ηp2 = .27, but not when the target appeared at a location other than that cued by the target color (−2 ms), F(1, 19) < 1, p = .64; at the location cued by the distractor color (8 ms), F(1, 19) = 1.01, p = .33; or at a location other than that cued by the distractor color (−3 ms), F(1, 19) < 1, p = .47.
PE
The main effect of cue validity was significant, F(1, 19) = 6.22, p < .05, MSe = .001, ηp2 = .25, indicating that PE was higher on invalid trials (4.53%) than on valid ones (3.20%). No other main effects or interactions reached significance.
Discussion
As in Experiment 1, no evidence for attentional capture or postattentive inhibition emerged when the distractor color cue was presented, which is inconsistent with the rapid disengagement account. Specifically, neither a cue-validity effect nor a cue–target compatibility effect occurred with the distractor color cue. However, a cue-validity effect emerged when the target color cue was presented, even in the context of more complex stimuli. When the target color cue was presented, there was a significant effect of the compatibility between the cued and target objects on valid trials, but not on invalid trials. As suggested earlier, on invalid trials, it is possible that the target display’s nontarget object was recognized at the cued location, resulting in the dilution of the cued object’s influence. Alternatively, on valid trials, forward masking could have occurred when the cued object in the cue display and the target were perceptually different, resulting in a delayed perception of the target relative to when the cued object and the target were perceptually identical. On the other hand, forward masking might have been absent on invalid trials, which could explain the observation that the cue–target compatibility effect was present only on valid trials cued by the target color.
Experiment 3
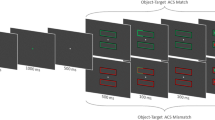
Experiment 3 examined whether the effect of the compatibility between cued and target objects occurs due to perceptual forward masking elicited when the objects in the cue and target displays are perceptually different. In Experiment 2, capital Ts and Ls appeared in both the cue and target displays. The letters were identical in their physical shapes and names on cue–target compatible trials and different on incompatible trials. Accordingly, forward masking, if it existed, would have been greater for cue–target incompatible trials than for compatible ones. This experiment utilized lowercase letters in the cue display and capital letters in the target display so that the shapes of the cued and target objects were different on all trials, thereby minimizing the asymmetrical perceptual forward masking between cue–target compatible and incompatible trials (see Fig. 4). If the cue–target compatibility effect obtained on valid trials cued by the target color reflected the forward masking effect, it could have been reduced or eliminated when the cued and target objects were presented in different physical shapes on each trial. However, if the recognition of the cued object elicited a significant cue–target compatibility effect, the cue–target compatibility effect would emerge for valid trials cued by the target color, as in the previous experiments.

Example of a trial sequence in Experiment 3
Method
Participants
A new group of 20 undergraduate students (ages 19 to 33), drawn from the same pool as in the previous experiments, participated in this experiment. As before, the participants received 6,000 KRW in compensation, and all reported normal or corrected-to-normal visual acuity and color vision.
Apparatus and stimuli
The apparatus and stimuli were the same as those used in Experiment 2, with one modification. We replaced the letters “T” and “L” within the placeholders in the cue display with white lowercase letters “a” (0.54° × 0.57°) and “h” (0.51° × 0.69°). In the target display, one target-colored, one distractor-colored, and two white capital As (0.66° × 0.66°) and Hs (0.66° × 0.60°) were randomly assigned to each of the four placeholders.
Design and procedure
The participants responded to the target-colored letter while ignoring nontarget-colored letters. They pressed the “f” key with their left index finger in response to the target letter “A” and the “j” key with their right index finger in response to the target letter “H.” Again, we emphasized the importance of accuracy and speed in performance. Finally, the target and distractor colors were counterbalanced across participants.
Results
Using the same criteria as in the previous experiments, we excluded 0.12% of all trials from the analyses as outliers. We calculated the mean correct RT and PE for each participant as a function of cue congruency (target color or distractor color), cue validity (valid or invalid), and cue–target compatibility (compatible or incompatible). We conducted ANOVAs on the mean correct RT and PE data with the abovementioned variables as within-subject variables (see Fig. 5).

Mean RT and PE as a function of congruency, validity, and compatibility in Experiment 3
RT
The main effects of all three independent variables were significant. The mean RT was greater for congruent trials (M = 519 ms) than for incongruent ones (M = 511 ms), F(1, 19) = 15.7, p < .001, MSe = 149, ηp2 = .45. Moreover, the mean RT was greater on invalid trials (M = 521 ms) than on valid ones (M = 509 ms), F(1, 19) = 21.9, p < .001, MSe = 282, ηp2 = .54, indicating a 12-ms cue-validity effect. The mean RT was greater for cue–target incompatible trials (M = 520 ms) than for compatible ones (M = 510 ms), F(1, 19) = 14.3, p < .01, MSe = 255, ηp2 = .43.
Cue validity interacted with cue congruency, F(1, 19) = 16.8, p < .001, MSe = 355, ηp2 = .47. Further analyses revealed a significant cue-validity effect on congruent trials (25 ms), F(1, 19) = 38.9, p < .0001, MSe = 156, ηp2 = .67, but not on incongruent ones (0 ms), F(1, 19) < 1, p = .95. Cue–target compatibility interacted with cue validity, F(1, 19) = 10.7, p < .01, MSe = 150, ηp2 = .36. A significant 16-ms cue–target compatibility effect occurred when the target was presented at the cued location, F(1, 19) = 16.42, p < .001, MSe =153, ηp2 = .46, but not when it was presented at an uncued location, F(1, 19) = 2.08, p =.17. Cue congruency also interacted with cue–target compatibility, F(1, 19) = 10, p < .01, MSe = 169, ηp2 = .35. Further analyses at each cue congruency level showed a significant 16-ms cue–target compatibility effect for congruent trials, F(1, 19) = 16, p < .001, MSe = 161, ηp2 = .46, but no effect for incongruent ones, F(1, 19) = 1.79, p = .20.
The three-way interaction of cue congruency, cue validity, and cue–target compatibility was marginally significant, F(1, 19) = 3.01, p = .098, MSe =117, ηp2 = .14. When the cue–target compatibility effect was examined at each cue congruency and cue-validity level, significant cue–target compatibility effects emerged both when the target appeared at the location cued by the target color, F(1, 19) = 17.4, p < .001, MSe = 369, ηp2 = .48, and when the target appeared at an uncued location with the target color cue, F(1, 19) = 5.42, p < .05, MSe = 83.3, ηp2 = .22. When the distractor color cues were presented, no cue–target compatibility effect occurred for valid, F(1, 19) = 2.37, p = .14, or invalid trials, F(1, 19) < 1, p = .90. Furthermore, separate two-way ANOVAs conducted for the congruent trials indicated that the cue–target compatibility effect was larger for valid trials than for invalid ones, F(1, 19) = 13.3, p < .01, MSe = 131, ηp2 = .41, and this interaction was not significant for incongruent trials, F(1, 19) = 1.66, p =.21.
PE
The main effect of cue validity was significant, F(1, 19) = 5.62, p < .05, MSe = .0009, ηp2 = .23. PE was higher for invalid trials (3.3%) than for valid ones (2.5%). No other main effects or interactions reached significance.
Discussion
As in the previous experiments, a significant cue-validity effect emerged only when the target color cue was presented, indicating the occurrence of attentional capture. Importantly, even after minimizing asymmetric forward masking effects, a 26-ms, significant cue–target compatibility still emerged for valid trials cued by the target color, indicating that the cued letter was recognized. Although a significant 7-ms cue–target compatibility effect also occurred for the invalid trials, the cue–target compatibility effect was significantly smaller for invalid trials than for valid ones, possibly reflecting the modulation of the cue–target compatibility effect due to the object of the target display that appeared at the cued location. That is, the object presented at the cued location in the target display may have been recognized even on invalid trials because attention was allocated to the cued location when the target color cue appeared and remained there for a short time after the onset of the target display (Becker et al., 2010).
To examine this possibility, we conducted additional ANOVAs for the mean correct RTs on invalid trials only using cue congruency, cue–target compatibility, and compatibility between the target object and the nontarget object in the target display that appeared at the cued location—that is, target display compatibility (TDC), determined as compatible or incompatible—as within-subject variables (see Fig. 6). The results indicated that the interaction of congruency and TDC was significant, F(1, 19) = 12, p < .01, MSe = 71.8, ηp2 = .39. When the target color cue was presented, a significant TDC effect emerged, F(1, 19) = 73.8, p < .0001, MSe = 20.9, ηp2 = .80, indicating that the object was recognized at the cued location in the target display. However, no such compatibility effect occurred when the distractor color cue was presented, F(1, 19) = 1.76, p = .20, MSe = 55.3, ηp2 = .08, which provides additional evidence that attention was not captured by the distractor color cue. These results indicate that, as posited by the contingent capture account (Folk et al., 1992), the color cue captures attention only when it is presented in a target-defining color.

Mean RT and as a function of congruency, compatibility, and target display objects compatibility (TDC) on invalid trials of Experiment 3. The error bar indicates within-subject standard error
Experiment 4
In Experiments 1–3, neither a cue-validity nor a reversed cue-validity effect occurred with the presentation of the distractor color cue. This lack of effect was possibly because the distractor cue was not sufficiently strong or salient to capture attention. Thus, to increase the magnitude of attentional capture so that it could be detected more easily using the cue-validity and the cue–target compatibility effects, we made two additional changes in Experiment 4. First, we increased the duration of the cue display from 50 to 150 ms to enhance the conscious perception of the cue. Second, we reduced cue-offset/target-onset asynchrony (CTOA) to 0 ms (150-ms stimulus onset asynchrony [SOA]) to prevent attentional disengagement from the distractor color cue, assuming that the cue captured attention. In Theeuwes and Godijn’s (2002) experiments, which used a similar approach, the researchers shortened the SOA between the cue display and the target display from 1,300 ms (Experiment 1) to 133 ms (Experiment 2). They found a reversed cue-validity effect for a no-go feature cue when the SOA was 1,300 ms, and this became a positive cue-validity effect when the SOA was shortened to 133 ms. Theeuwes and Godijn argue that the 130-ms SOA window allowed no extra time for attention to reorient to another location, thus revealing a facilitation effect that might have been obscured by the postattentive suppression.
If these changes successfully prevent attentional disengagement from the cued location, which possibly clouded any indication of attentional capture by distractor color cues in the previous experiments, significant cue validity and cue–target compatibility effects would be evident with the distractor color cue. Nevertheless, if the location cued by the distractor color is unattended, as the contingent capture account suggests, neither the cue-validity effect nor the cue–target compatibility effect would be evident on trials with the distractor color cue.
Method
Participants
Drawn from the same participant pool as in Experiment 1, 16 new undergraduate students participated in the experiment (ages 20 to 29). Participants received 7,000 KRW (approximately 6 USD) as compensation. All reported having normal or corrected-to-normal visual acuity and color vision.
Apparatus and stimuli
The apparatus and stimuli were the same as those used in Experiment 2. White Ts and Ls were randomly placed in the cue display, whereas one target-colored, one distractor-colored, and two white Ts and Ls were randomly assigned in the target display.
Design and procedure
The design and procedure of Experiment 4 were similar to those of Experiment 2. We made two modifications in this experiment, one of which was to increase the duration of the cue display from 50 to 150 ms. More importantly, we changed the total time between the offset of the cue display and the onset of the target display from 150 to 0 ms. Therefore, the target display appeared immediately after the cue display disappeared. The participants waited for the target display, then responded to the target-colored letter.
Results
With the same criteria as those in the previous experiments, we excluded 35 trials from the analyses, accounting for 0.36% of the total data. The mean correct RT and PE were calculated for each participant as a function of cue congruency (target color or distractor color), cue validity (valid or invalid), and cue–target compatibility (compatible or incompatible). ANOVAs were conducted on the mean correct RT and PE data with the above factors as within-subject variables (see Fig. 7).

Mean RT and PE as a function of congruency, validity, and compatibility in Experiment 4
RT
The main effect of cue congruency was significant, F(1, 15) = 14.9, p < .01, MSe = 912, ηp2 = .50. The mean RT was greater when the target color cue appeared (M = 564 ms) than when the distractor color cue appeared (M = 543 ms). The main effect of cue-validity was significant, F(1, 15) = 11.2, p < .01, MSe = 476, ηp2 = .43. The mean RT was longer for invalid trials (M = 560 ms) than for valid ones (M = 547 ms), indicating a 13-ms cue-validity effect. The main effect of cue–target compatibility was significant, F(1, 15) = 17.9, p < .001, MSe = 570, ηp2 = .55. Participants responded to the target approximately 17 ms more slowly on incompatible trials (M = 562 ms) than on compatible ones (M = 545 ms).
Cue-validity interacted with cue congruency, F(1, 15) = 19.9, p < .001, MSe = 543, ηp2 = .57. Additional analyses of the cue-validity effect at each congruency level confirmed a significant 31-ms cue-validity effect when the target color cue was presented, F(1, 15) = 17.6, p < .001, MSe = 444, ηp2 = .54. In contrast, a marginally significant reversed 5-ms cue-validity effect occurred when the distractor color cue was presented, F(1, 15) = 3.64, p = .076, MSe = 66.0, ηp2 = .20. Cue–target compatibility interacted with cue-validity, F(1, 15) = 18.6, p < .001, MSe = 545, ηp2 = .55. A significant 35-ms compatibility effect occurred for valid trials, F(1, 15) = 19.7, p < .001, MSe = 517, ηp2 = .57, but not for invalid ones, F(1, 15) < 1, p = .97. The interaction of cue–target compatibility with cue congruency was significant, F(1, 15) = 32.2, p < .0001, MSe = 266, ηp2 = .68. Further analyses at each cue congruency level indicated that the cue–target compatibility effect was significant for congruent trials (34 ms), F(1, 15) = 36.3, p < .0001, MSe = 258, ηp2 = .71, but not for incongruent ones (2 ms), F(1, 15) < 1, p = .73.
The three-way interaction of cue congruency, cue validity, and cue–target compatibility was significant, F(1, 15) = 21, p < .001, MSe = 214, ηp2 = .58. Further analyses revealed a significant cue–target compatibility effect when the target was presented at the location cued by the target color (64 ms), F(1, 15) = 54.4, p < .0001, MSe = 601, ηp2 = .78. However, a nonsignificant compatibility effect occurred when the target was presented at a location other than that cued by the target color (5 ms), F(1, 15) = 1.33, p = .27. When the distractor color cue was presented, no compatibility effect occurred on valid trials (8 ms), F(1, 15) < 1, p = .45, or on invalid trials (-4 ms), F(1, 15) = 1.53, p = .24.
As in Experiment 3, we conducted an additional ANOVA on the data from invalid trials using cue congruency, cue–target compatibility, and target display compatibility (TDC) as within-subject variables (see Fig. 8). The interaction of cue–target compatibility and TDC was significant, F(1, 15) = 12.2, p < .01, MSe = 111, ηp2 = .45. The mean RT was shorter for cue–target compatible trials (M = 553 ms) than for incompatible ones (M = 561 ms) when the object at the cued location in the target display matched the target, F(1, 15) = 4.86, p < .05, MSe = 125, ηp2 = .24. The mean RT was marginally longer in the cue–target compatible trials (M = 563 ms) than in the incompatible ones (M = 559 ms) when the cued nontarget object in the target display differed from the target, F(1, 15) = 4.11, p = .061, MSe = 35, ηp2 = .22. Importantly, the interaction of cue congruency and TDC was marginally significant, F(1, 15) = 3.87, p = .068, MSe = 136, ηp2 = .2. TDC was significant when the target color cue was presented, F(1, 15) = 5.38, p < .05, MSe = 86.7, ηp2 = .26, indicating that the object presented at the cued location in the target display was recognized. However, the effect of TDC was not significant when the distractor color cue was presented, F(1, 15) < 1, p = .86, indicating that the object presented at the cued location in the target display was not recognized.

Mean RT and as a function of congruency, compatibility, and target display objects compatibility (TDC) on invalid trials of Experiment 4
PE
The main effects of cue congruency and cue validity were significant. PE was higher for congruent trials (6.6%) than for incongruent ones (4.1%), F(1, 15) = 15.9, p < .01, MSe = .0013, ηp2 = .52. PE was also higher for invalid trials (6.0%) than for valid ones (4.7%), F(1, 15) = 6.45, p < .05, MSe = .0009, ηp2 = .30. No other interactions reached significance.
Discussion
In Experiment 4, the magnitude of the cue-validity effect was 39 ms, the largest among the cue-validity effects observed on congruent trials in the previous experiments. Therefore, the changes implemented in Experiment 4 successfully induced stronger attentional capture using salient cues without directly manipulating attentional control. Nevertheless, evidence of attentional capture was apparent only when the target color cue was presented, as suggested by the contingent capture account (Folk et al., 1992). A significant cue-validity effect occurred when the target color cue appeared, and the cue–target compatibility effect was significant only when the target appeared at the location cued by the target color. Moreover, the target-display object at the cued location was recognized for invalid trials but only when it was cued by the target color. In contrast, although a marginally significant reversed cue-validity effect occurred when the distractor color cue appeared, which may be taken to indicate attentional inhibition, a reversed cue-validity effect likely did not occur after attentional disengagement because there was no evidence indicating that the cued object or the object at the cued location of the target display was recognized.
Experiment 5
In Experiments 1–3, no significant reversed cue-validity effect occurred when the distractor color cue appeared, possibly suggesting that the attentional control setting recruited by the task was not strong enough to induce rapid attentional disengagement and postattentive inhibition. The contingent capture account posits the nonsignificant cue-validity effect as an indication of no attentional capture by nontarget features, whereas the salience-driven capture account posits it as an indication of rapid attentional disengagement after attentional capture. We conducted Experiment 5 to resolve this discrepancy between the two accounts by examining the cue–target compatibility effect in a situation showing a reversed cue-validity effect.
Therefore, we adopted Belopolsky et al.’s (2010) approach to enhance the attentional control setting and increase the speed of attentional disengagement. For example, Belopolsky et al. found a reversed cue-validity effect in experiments using a go/no-go task, but there was no reversed cue-validity effect in experiments using a target identification task. They claim that participants exhibited strong, sustained attentional control throughout the whole block during the go/no-go task because they were constantly relying on the attentional control setting to identify whether a target was a go target or a no-go target.
However, in Anderson and Folk’s (2012) Experiments 4 and 5, where the go target color was unpredictably either red or green, and the no-go target color was always blue, significant cue-validity effects occurred for the go color cue but not for the no-go color cue. Interestingly, the mean RT for invalid trials in that experiment did not differ significantly with the color of the cue. They suggest that both color cues captured attention. However, due to capture-independent inhibition, which is unrelated to attentional disengagement, the location cued by the no-go color was suppressed, resulting in delayed responses for valid trials. Nevertheless, there was no significant difference between the invalid trials with the go color cue and those with the no-go color cue because there was no capture-independent inhibition on invalid trials.
The paradigm and stimuli used in Experiment 5 were the same as those in Experiment 2, except that we introduced a go/no-go task. To this end, we presented a color singleton target in the target display, and its color (three possibilities) was randomly selected. Participants responded to the identity of the singleton target when the target was red or green and withheld their response when the target was blue. We examined the cue-validity effect and the cue–target compatibility effect to determine whether a no-go color cue could capture attention when engaging a strong attentional control setting.
Method
Participants
Drawn from the same pool as in the previous experiments, 16 new undergraduate students (ages 20 to 30) participated in this experiment. All received KRW 8,000 (approximately 7 USD) for their participation. All reported having normal or corrected-to-normal visual acuity and color vision.
Apparatus and stimuli
The apparatus and stimuli were the same as those used in Experiment 2, with the exception that we changed the task to a go/no-go task. We removed a colored distractor from the target display so that the target object was the only color singleton item in the target display. We presented the spatial cues and the target letter randomly in red (RGB: 255, 0, 0; CIE: x = .64, y = .33), green (RGB: 13, 147, 13; CIE: x = .30, y = .55), or blue (RGB: 0, 0, 255; CIE: x = .22, y = .32). The other filler cues and letters were white. We designated red and green as the go colors and blue as the no-go color.
Design and procedure
We made two modifications in Experiment 5. First, we changed the nature of the task from target identification to go/no-go (see Fig. 9). Participants responded to red or green letters appearing in the target display and withheld their response when blue letters appeared in the display. One of the three colors for the spatial cue was randomly selected on each trial (256 trials for each color). The ratio of go color-cue trials to no-go color-cue trials was 2 to 1 because there were two different go-colors and one no-go color. As in previous experiments, a trial was labeled with respect to the color of the spatial cue, rather than the color of the target object, such that a trial cued by red or green dots was labeled as a go cue trial (congruent), while a trial cued by blue dots was labeled as a no-go cue trial (incongruent). A trial with a red “target” preceded by a green “cue,” for example, was labeled as congruent because the cue color matched one of the target-defining features in the attentional control setting, although the cue and target colors did not match. Second, we increased the number of trials to compensate for the loss of reaction time data from the no-go trials. All participants first completed a practice block of 20 trials and then performed four main blocks of 192 trials.

Example of a trial sequence in Experiment 5
Results
Using the same criteria as those in the previous experiments, we excluded 1.7% of the total trials from the analyses as outliers. The mean correct RT and PE were calculated for each participant as a function of cue congruency (go color cue or no-go color cue), cue validity (valid or invalid), and cue–target compatibility (compatible or incompatible). ANOVAs were conducted on the mean RT and PE data with the above factors as within-subject variables (see Fig. 10).

Mean RT and PE as a function of congruency, validity, and compatibility in Experiment 5
RT
The main effect of cue congruency was significant, F(1, 15) = 14.7, p < .01, MSe = 251, ηp2 = .49. The mean RT was greater when the no-go color cue was presented (M = 611 ms) than when the go color cue was presented (M = 600 ms). Moreover, the main effect of cue validity was significant, F(1, 15) = 49.6, p < .0001, MSe = 208, ηp2 = .77. The mean RT was greater for invalid trials (M = 615 ms) than for valid ones (M = 597 ms). The main effect of cue–target compatibility was also significant, F(1, 15) = 7.56, p < .05, MSe = 409, ηp2 = .34. The mean RT was greater for incompatible trials (M = 610 ms) than for compatible ones (M = 601 ms).
Cue validity interacted with cue congruency, F(1, 15) = 6.55, p < .05, MSe = 445, ηp2 = .30. A significant cue-validity effect occurred for go cue trials (27 ms), F(1, 15) = 96.1, p < .0001, MSe = 62.8, ηp2 = .87, but not for no-go cue trials (8 ms), F(1, 15) = 2.14, p = .16. The two-way interaction of cue–target compatibility and cue validity was significant, F(1, 15) = 13.6, p < .01, MSe = 278, ηp2 = .48. A significant 21-ms cue–target compatibility effect occurred when the target was presented at the cued location, F(1, 15) = 12.1, p < .01, MSe = 285, ηp2 = .45, but not when it was presented at an uncued location, F(1, 15) = .15, p = .70. The three-way interaction of cue congruency, cue validity, and cue–target compatibility was not significant, F(1, 15) = .01, p = .92.
As in Experiments 3 and 4, additional ANOVAs were conducted on the mean correct RT from invalid trials with cue congruency, cue–target compatibility, and TDC as within-subject variables (see Fig. 11). The main effect of TDC, F(1, 15) = 5.89, p < .05, MSe = 288, ηp2 = .28, and the interaction of cue–target compatibility and TDC, F(1, 15) = 28.5, p < .001, MSe = 1,724, ηp2 = .66, were significant. The three-way interaction of cue–target compatibility, cue congruency, and TDC was also significant, F(1, 15) = 5.22, p < .05, MSe = 412, ηp2 = .26. When the go color cue was presented, the interaction of cue–target compatibility and TDC was significant, F(1, 15) = 37.7, p < .0001, MSe = 954, ηp2 = .72. The cue–target compatibility effect occurred when the object of the target display presented at the cued location matched the target (TDC compatible), F(1, 15) = 37, p < .0001, MSe = 829, ηp2 = .71, while a significant reversed cue–target compatibility effect occurred when the object of the target display at the cued location differed from the target (TDC incompatible), F(1, 15) = 24, p < .001, MSe = 360, ηp2 = .62. When the no-go color cue was presented, unlike in the previous experiments, TDC modulated the cue–target compatibility effect, F(1, 15) = 13, p < .01, MSe = 1,182, ηp2 = .46. The cue–target compatibility effect was significant when the object of the target display presented at the cued location matched the target, F(1, 15) = 10.3, p < .01, MSe = 1,242, ηp2 = .41, while a reversed cue–target compatibility effect occurred when the object differed from the target, F(1, 15) = 7.49, p < .05, MS = 515, ηp2 = .33.

Mean RT and as a function of congruency, compatibility, and target display objects compatibility (TDC) on invalid trials of Experiment 5. Dashed lines on the lower-left corner indicate the hypothesized path of attention
PE
The main effect of cue validity was significant, F(1, 15) = 5.24, p < .05, MSe = .0008, ηp2 = .26. PE was higher for invalid trials (3.0%) than for valid ones (1.9%). No other main effect or interaction was significant.
Discussion
In Experiment 5, the go color cues captured attention, as indicated by the significant cue-validity effect and cue–target compatibility effect. As in the previous experiments, no cue–target compatibility effect occurred for invalid trials when the go color cues appeared. Importantly, when the no-go color cue was presented, a significant cue–target compatibility effect occurred for valid trials but not for invalid ones, although no cue-validity effect emerged. Similar to the results of Belopolsky et al.’s (2010) experiments, there was a 20-ms increase in RT for valid trials when the no-go color cue was presented compared to when the go color cues were presented. This RT cost for valid trials eventually clouded the cue-validity effect for the no-go color cue, which aligns with the rapid disengagement account.
However, it seems unlikely that the RT cost occurring on valid trials with the no-go color cue would have been the result of postattentive inhibition induced by attentional disengagement. As in Anderson and Folk’s (2012) experiments, we found no evidence for rapid attentional disengagement on invalid trials. The mean RT of invalid trials was 615 ms when the no-go color cue was presented and 614 ms when the go color cues were presented, and this difference was not significant. Moreover, unlike in the previous experiments, the cue–target compatibility effect was significant on valid trials but not on invalid ones, regardless of whether the go color or no-go color cue was presented. Lastly, the main effect of TDC was significant, but it did not interact with cue congruency, which suggests that the object at the cued location in the target display was recognized regardless of whether it was cued by a go color cue or a no-go color cue. These results indicate that both the no-go color cue and the go color cues captured attention.
Unlike in the previous experiments, the reversed cue–target compatibility effect occurred on TDC incompatible trials regardless of cue congruency, even though this pattern was significantly more evident when the go color cue was presented than when the no-go color cue was presented (see Fig. 11). This finding indicates that the compatibility between the cued object in the cue display and the nontarget object in the target display that appeared at the cued location exerted a greater influence on task performance than did cue–target compatibility. This influence might have been amplified, possibly because of the increased task difficulty in Experiment 5.
General discussion
In this study, we manipulated the compatibility of cued and target objects to more precisely investigate attentional capture by a task-irrelevant feature. Additionally, we investigated the presence of attentional capture and postattentive inhibition in cases of enhanced attentional control setting (Experiment 2) or with induction of more robust attentional capture (Experiment 4) to ensure detection of attentional capture by the distractor color cue. Nevertheless, we found no such direct indication of attentional capture by the distractor color cue. In all experiments, significant cue-validity effects occurred when the target color cue was presented, but not when the distractor color cue was presented. Importantly, when the target color cue was presented, a compatibility effect between the cued and target objects occurred for valid trials but not for invalid trials in Experiments 1–4. Moreover, this finding was still evident after minimizing the asymmetric influences of perceptual forward masking for cue–target compatible and incompatible trials. These findings suggest that the object in the location cued by the target color was recognized, resulting in a significant cue–target compatibility effect for valid trials. Furthermore, the object at the cued location in the target display diluted the cue–target compatibility effect for invalid trials cued by the target color.
However, when the distractor color cue was presented, as suggested by the contingent capture account (e.g., Folk et al., 1992), we found no significant (reversed) cue-validity effect in Experiments 1–3. Moreover, no cue–target compatibility effect occurred, regardless of cue validity. This pattern of results resurfaced after we shortened the cue–target interval and presented the spatial cue for a longer time in Experiment 4. A marginal reversed cue-validity effect occurred without a cue–target compatibility effect in Experiment 4. Nevertheless, it is highly likely that the reversed cue-validity effect is independent of inhibition after disengagement because there was less time for attention to reorient to another location, relative to the other experiments where no reversed cue-validity occurred. Altogether, these findings imply that salient cues containing to-be-ignored features do not capture attention when the attentional control setting is tuned to a specific feature during visual search.
In Experiment 5, in which participants performed a go/no-go task, the cue-validity effect occurred only when the go target color cue was presented but not for the no-go cue, as in the other experiments. Furthermore, as the rapid disengagement account predicts, the mean RT for the target presented at the cued location was longer when the no-go color cue was presented than when the go target color cue was presented. Importantly, unlike in the other experiments, a 21-ms compatibility effect occurred at the cued location primed by a go target color, and a 20-ms compatibility effect occurred at the cued location primed by the no-go target color, indicating attentional capture by both types of cues. However, this significant cue–target compatibility effect does not necessarily indicate that the lack of the cue-validity effect with the no-go color cue was due to location-specific postattentive inhibition, as Belopolsky et al. (2010) suggest. If attention was rapidly disengaged from the cued location, the mean RT for invalid trials should have been shorter with the no-go target color cues than with the go target color cues.
Rather, it seems more plausible that the no-go target cue simply captured attention, but attention was not disengaged for valid trials. Indeed, the contingent capture account suggests that a no-go target feature can capture attention when the target is defined as a singleton object (Bacon & Egeth, 1994). Furthermore, the target was always a singleton object, and the no-go target color was equally important as the go target color in performing the go/no-go task. Accordingly, simply inhibiting the no-go target color could have impaired task performance in Experiment 5, unlike in the previous experiments where inhibiting the distractor color would have resulted in improved task performance. An alternative possibility for the RT cost observed for valid trials is that the go/no-go task requirements induced it, which would be independent of attentional disengagement or postattentive inhibition, as Anderson and Folk (2012) note. In a similar vein, some studies have reported reversed cue-validity effects, and attributed the reversed effects to sources other than postattentive inhibition (Carmel & Lamy, 2014, 2015; Lamy et al., 2004).
The cue–target compatibility effect
Although attentional capture by the target color cue occurred consistently in Experiments 1–4, the cue–target compatibility effect occurred for valid but not invalid trials. Folk and Remington (2006) found that the cue–target compatibility effect decreased considerably for invalid trials relative to valid trials when the target color cue was presented, although this difference was not statistically significant. The decreased cue–target compatibility effects or the absence of compatibility effects were possibly due to postattentive inhibition. It is widely accepted that attentional disengagement leads to the suppression of a previously attended location (e.g., Belopolsky et al., 2010; Klein, 2000). Thus, responses to the target presented at the cued location are typically delayed when CTOA is longer (>300 ms; e.g., Tipper et al., 1994), a phenomenon called the inhibition of return (Posner & Cohen, 1984).
Fuentes et al. (1999) argue that as attention is disengaged from a location, temporary inhibition applies not only to previously attended locations, but also to the stimuli that had been presented at that location. They found a significant flanker-compatibility effect when a distractor was presented at an uncued location but a reversed flanker compatibility effect when it was presented at a previously cued location. Similarly, Wyatt and Machado (2013) demonstrated that the flanker-compatibility effect decreases as a function of the temporal delay between the onsets of a peripheral flanker and a central target. In their study, positive flanker-compatibility effects occurred at shorter delays, while reversed flanker-compatibility effects occurred at longer delays. These results imply that attentional disengagement from a location causes suppression of the location along with the stimuli presented at that location, which might be why we did not observe the cue–target compatibility effect in this study, except in valid trials cued by the target color.
However, as mentioned in the Discussion section for Experiment 3, the reason for the absence of the cue–target compatibility effect for invalid trials cued by the target color was that the object at the cued location in the target display was recognized instead of postattentive inhibition being applied to the cued object. Additional analyses of the data from invalid trials showed that, when the target color cue was presented, the compatibility between the TDC was significant in Experiments 1–4, indicating that the cued nontarget object presented in the target display was recognized. More specifically, because the focus of attention shifted to the location cued by the target color, the object subsequently presented in the target display at this attended location was recognized, although this cued object was not a target. In contrast, when the distractor color cue was presented, no TDC effect occurred in Experiments 1–4. These results imply that the null effects for cue-validity and cue–target compatibility obtained using the distractor color cue were not due to postattentive inhibition.
Preattentive suppression of distractor features
Previous event-related potentials (ERPs) research has demonstrated that a color singleton distractor elicits greater positivity at contralateral scalp sites than ipsilateral scalp sites (e.g., Gaspar et al., 2016; Hickey et al., 2009; Sawaki & Luck, 2010, 2013). Sawaki and Luck (2010) suggest that this distractor positivity (Pd) reflects the suppression of the distractor. According to their signal suppression hypothesis, although a color singleton generates a salience signal, a top-down inhibitory mechanism preattentively suppresses the signal. Additionally, Gaspelin et al. (2015) found that letter probes presented inside singleton distractor objects were less likely to be recalled than probes presented inside either nonsingleton nontarget objects or target objects. They suggested that because the salience signal generated by a distractor color singleton is suppressed without the involvement of attentional capture or attentional disengagement, the probe letter inside the color singleton distractor is less likely to be attended, resulting in poor recall performance. Moreover, Chang and Egeth (2019) suggest that the selection of a target-defining feature and the suppression of a distractor-defining one operate independently to guide attention. In their subsequent study, Chang and Egeth (2021) demonstrated that this preattentive suppression mechanism alone has the potential to override attentional capture by a distractor feature.
In this study, it is unclear whether the location cued by the distractor color was preattentively suppressed. Nevertheless, the findings also provided no evidence indicating that the distractor color cue, unlike the target color cue, captures attention or that the object cued by the distractor color or the target display object presented at the cued location is recognized. However, it is possible that the distractor color cue was suppressed preattentively based on its feature value (Gaspelin & Luck, 2018) because participants adopted a feature search mode and the target and distractor colors remained fixed throughout each experiment. Moreover, the reversed cue-validity effect obtained with the distractor color cue in Experiment 4, although it was marginally significant, implies that the location occupied by a distractor feature might have been preattentively suppressed in the earlier stages of visual information processing (i.e., preattentive selection) because it also failed to provide evidence that the cued object or the target display object at the cued location was recognized.
On the other hand, we obtained no significant reversed cue-validity effect or cue–target compatibility effect with distractor color cues. Such reversed effects would have strengthened the conclusion that the cues were preattentively suppressed, which accords with previous research revealing poorer recall performance for objects cued by to-be-ignored features compared to those cued by neutral features (e.g., Chang & Egeth, 2019; Gaspelin et al., 2015). This raises another possibility that the distractor cues were not preattentively suppressed but were simply not selected because they were not congruent with the attentional control setting. Similarly, Livingstone et al. (2017) showed that the Pd is not time-locked to the onset of cue displays but to the onset of following target displays. They suggest that the Pd component reflects the facilitation of early perceptual processing of a nontarget object in the target display that appears at the cued location rather than the preattentive suppression of the distractor feature.
Conclusion
The results of this study support the view that top-down attentional control setting plays a critical role in guiding involuntary attentional capture, demonstrating that objects primed by the target-defining feature are recognized and those primed by the distractor-defining feature are not, leaving aside attentional capture by the target-defining feature. These findings confirm that preattentive selection and postattentive inhibition depend on top-down attentional control setting. Even though future study is needed to validate the compatibility manipulations adopted in the present study, the findings of this study elucidate how our attentional focus involuntarily shifts toward external events.
References
Anderson, B. A., & Folk, C. L. (2012). Dissociating location-specific inhibition and attention shifts: Evidence against the disengagement account of contingent capture. Attention, Perception, & Psychophysics, 74(6), 1183–1198. https://doi.org/10.3758/s13414-012-0325-9
Bacon, W. F., & Egeth, H. E. (1994). Overriding stimulus-driven attentional capture. Perception & Psychophysics, 55(5), 485–496. https://doi.org/10.3758/BF03205306
Becker, S. L., Folk, C. L., & Remington, R. W. (2010). The role of relational information in contingent capture. Journal of Experimental Psychology: Human Perception and Performance, 36(6), 1460–1476. https://doi.org/10.1037/a0020370
Belopolsky, A. V., Schreij, D., & Theeuwes, J. (2010). What is top-down about contingent capture? Attention, Perception, & Psychophysics, 72(2), 326–341. https://doi.org/10.3758/APP.72.2.326
Carmel, T., & Lamy, D. (2014). The same-location cost is unrelated to attentional settings: An object-updating account. Journal of Experimental Psychology: Human Perception and Performance, 40(4), 1465–1478. https://doi.org/10.1037/a0036383
Carmel, T., & Lamy, D. (2015). Towards a resolution of the attentional-capture debate. Journal of Experimental Psychology: Human Perception and Performance, 41(6), 1772-1782. https://doi.org/10.1037/xhp0000118
Chang, S., & Egeth, H. E. (2019). Enhancement and suppression flexibly guide attention. Psychological Science, 30(12), 1724–1732. https://doi.org/10.1177/0956797619878813
Chang, S., & Egeth, H. E. (2021). Can salient stimuli really be suppressed?. Attention, Perception, & Psychophysics, 83, 260–269. https://doi.org/10.3758/s13414-020-02207-8
Faul, F., Erdfelder, E., Buchner, A., & Lang, A. G. (2009). Statistical power analyses using G* Power 3.1: Tests for correlation and regression analyses. Behavior Research Methods, 41(4), 1149–1160. https://doi.org/10.3758/BRM.41.4.1149
Folk, C. L., & Anderson, B. A. (2010). Target-uncertainty effects in attentional capture: Color-singleton set or multiple attentional control settings? Psychonomic Bulletin & Review, 17(3), 421–426. https://doi.org/10.3758/PBR.17.3.421
Folk, C. L., Leber, A. B., & Egeth, H. E. (2002). Made you blink! Contingent attentional capture produces a spatial blink. Perception & Psychophysics, 64(5), 741–753. https://doi.org/10.3758/BF03194741
Folk, C. L., & Remington, R. (1998). Selectivity in distraction by irrelevant featural singletons: Evidence for two forms of attentional capture. Journal of Experimental Psychology: Human Perception and Performance, 24(3), 847–858. https://doi.org/10.1037/0096-1523.24.3.847
Folk, C. L., & Remington, R. (2006). Top-down modulation of preattentive processing: Testing the recovery account of contingent capture. Visual Cognition, 14(4/8), 445–465. https://doi.org/10.1080/13506280500193545
Folk, C. L., Remington, R. W., & Johnston, J. C. (1992). Involuntary covert orienting is contingent on attentional control settings. Journal of Experimental Psychology: Human Perception and Performance, 18(4), 1030–1044. https://doi.org/10.1037/0096-1523.18.4.1030
Folk, C. L., Remington, R. W., & Wright, J. H. (1994). The structure of attentional control: Contingent attentional capture by apparent motion, abrupt onset, and color. Journal of Experimental Psychology: Human Perception & Performance, 20(2), 317–329. https://doi.org/10.1037/0096-1523.20.2.317
Fuentes, L. J., Vivas, A. B., & Humphreys, G. W. (1999). Inhibitory tagging of stimulus properties in inhibition of return, Effects on semantic priming and flanker interference. The Quarterly Journal of Experimental Psychology, 52A(1), 149–164. https://doi.org/10.1080/713755797
Gaspar, J. M., Christie, G. J., Prime, D. J., Jolicœur, P., & McDonald, J. J. (2016). Inability to suppress salient distractors predicts low visual working memory capacity. Proceedings of the National Academy of Sciences of the United States of America, 113(13), 3693–3698. https://doi.org/10.1073/pnas.1523471113
Gaspelin, N., Leonard, C. J., & Luck, S. J. (2015). Direct evidence for active suppression of salient-but-irrelevant sensory inputs. Psychological Science, 26(11), 1740–1750. https://doi.org/10.1177/0956797615597913
Gaspelin, N., & Luck, S. J. (2018). Distinguishing among potential mechanisms of singleton suppression. Journal of Experimental Psychology: Human Perception and Performance, 44(4), 626–644. https://doi.org/10.1037/xhp0000484
Hickey, C., Di Lollo, V., & McDonald, J. J. (2009). Electrophysiological indices of target and distractor processing in visual search. Journal of Cognitive Neuroscience, 21(4), 760–775. https://doi.org/10.1162/jocn.2009.21039
Jonides, J., & Yantis, S. (1988). Uniqueness of abrupt visual onset in capturing attention. Perception & Psychophysics, 43(4), 346–354. https://doi.org/10.3758/BF03208805
Klein, R. M. (2000). Inhibition of return. Trends in Cognitive Sciences, 4(4), 138–147. https://doi.org/10.1016/S1364-6613(00)01452-2
Lamy, D., Leber, A., & Egeth, H. E. (2004). Effects of task relevance and stimulus-driven salience in feature-search mode. Journal of Experimental Psychology: Human Perception and Performance, 30(6), 1019–1031. https://doi.org/10.1037/0096-1523.30.6.1019
Livingstone, A. C., Christie, G. J., Wright, R. D., & McDonald, J. J. (2017). Signal enhancement, not active suppression, follows the contingent capture of visual attention. Journal of Experimental Psychology: Human Perception and Performance, 43(2), 219–224. https://doi.org/10.1037/xhp0000339
Posner, M. I. (1980). Orienting of attention. The Quarterly Journal of Experimental Psychology, 32(1), 3–25. https://doi.org/10.1080/00335558008248231
Posner, M. I., & Cohen, Y. (1984). Components of visual orienting. Erlbaum.
Pratt, J., & Hommel, B. (2003). Symbolic control of visual attention: The role of working memory and attentional control settings. Journal of Experimental Psychology: Human Perception and Performance, 29(5), 835–845. https://doi.org/10.1037/0096-1523.29.5.835
Remington, R. W., Folk, C. L., & McLean, J. P. (2001). Contingent attentional capture or delayed allocation of attention? Perception & Psychophysics, 63(2), 298–307. https://doi.org/10.3758/BF03194470
Sagi, D., & Julesz, B. (1985). ‘Where’ and’ what’ in vision. Science, 228(4704), 1217–1219. https://doi.org/10.1126/science.4001937
Sawaki, R., & Luck, S. J. (2010). Capture versus suppression of attention by salient singletons: Electrophysiological evidence for an automatic attend-to-me signal. Attention, Perception, & Psychophysics, 72(6), 1455–1470. https://doi.org/10.3758/APP.72.6.1455
Sawaki, R., & Luck, S. J. (2013). Active suppression after involuntary capture of attention. Psychonomic Bulletin & Review, 20(2), 296–301. http://dx.doi.org/10.3758/s13423-012-0353-4
Shih, S. I., & Sperling, G. (1996). Is there feature-based attentional selection in visual search? Journal of Experimental Psychology: Human Perception and Performance, 22(3), 758–779. https://doi.org/10.1037/0096-1523.22.3.758
Theeuwes, J. (1991). Cross-dimensional perceptual selectivity. Perception & Psychophysics, 50(2), 184–193. https://doi.org/10.3758/BF03212219
Theeuwes, J. (1992). Perceptual selectivity for color and form. Perception & Psychophysics, 51(6), 599–606. https://doi.org/10.3758/BF03211656
Theeuwes, J. (1994). Stimulus-driven capture and attentional set: selective search for color and visual abrupt onsets. Journal of Experimental Psychology: Human Perception and Performance, 20(4), 799–806. https://doi.org/10.1037/0096-1523.20.4.799
Theeuwes, J., Atchley, P., & Kramer, A. F. (2000). On the time course of top-down and bottom-up control of visual attention. In S. Monsell & J. Driver (Eds.), Attention and performance XVIII: Control of cognitive processes (pp. 105–124). MIT Press.
Theeuwes, J., & Godijn, R. (2002). Irrelevant singletons capture attention: Evidence from inhibition of return. Perception & Psychophysics, 64(5), 764–770. https://doi.org/10.3758/BF03194743
Tipper, S. P., Weaver, B., Jerreat, L. M., & Burak, A. L. (1994). Object-based and environment-based inhibition of return of visual attention. Journal of Experimental Psychology: Human Perception and Performance, 20(3), 478–499. https://doi.org/10.1037/0096-1523.20.3.478
Ullman, S. (1984). Visual routines. Cognition, 18(1), 97–159. https://doi.org/10.1016/0010-0277(84)90023-4
Wyatt, N., & Machado, L. (2013). Evidence inhibition responds reactively to the salience of distracting information during focused attention. PLOS ONE, 8(4), Article e62809. https://doi.org/10.1371/journal.pone.0062809
Yantis, S., & Jonides, J. (1990). Abrupt visual onsets and selective attention: Voluntary versus automatic allocation. Journal of Experimental Psychology: Human Perception and Performance, 16(1), 121–134. https://doi.org/10.1037/0096-1523.16.1.121
Author note
This research was supported by the Korean Research Foundation Grant funded by the Korean Government (NRF-2020R1A2C2012033).
Author information
Authors and Affiliations
Corresponding author
Additional information
Open practice statements
The data and materials for all experiments are available (https://osf.io/qf4ju/?view_only=1c46bda2461741f78fd2af85b1d8b713).
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Rheem, H., Cho, Y.S. A finer-grained search reveals no evidence of the attentional capture by to-be-ignored features. Atten Percept Psychophys 83, 2441–2457 (2021). https://doi.org/10.3758/s13414-021-02305-1
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13414-021-02305-1