
Abstract
In the present study, we investigated the binding of location and response in a detection task of the target-target paradigm of inhibition of return (IOR). Results showed a cost of responding to a target at the repeated location (IOR) when the response was not repeated and an effect of facilitation of return (FOR) when the response was repeated. These findings suggest that when responding to a target, its location and the response to it are integrated together. In addition, an analysis of the Vincentized cumulative response time (RT) distribution further showed that memory retrieval of event representations requires time to operate. These findings were discussed according to the theoretical framework of event files.
Similar content being viewed by others


Introduction
Inhibition of return (IOR) refers to delayed response to a stimulus presented at a previously cued location (Posner & Cohen, 1984; Posner et al., 1985; see Klein, 2000, and Lupiáñez et al., 2006, for reviews). Regarding the source of IOR, Posner et al. (1985) proposed that IOR reflects the aftereffect of attentional inhibition of previously attended spatial locations. After this proposal, different accounts of IOR have been developed. Milliken et al. (2000) proposed the constructive retrieval account, suggesting that IOR occurs when the retrieval of previous memory episodes and the integration of the retrieved representation and the current stimulus is less efficient than attentional capture by novel stimuli. Dukewich (2009) proposed the habituation account, suggesting that IOR reflects the effects of habituation. In addition, Lupiáñez and colleagues (Lupiáñez, 2010; Lupiáñez et al., 2007, Lupiáñez et al., 2013; Martín-Arévalo et al., 2013) proposed the detection cost account, suggesting that IOR reflects the difficulty of detecting a target that is similar to the previous cue. In these accounts after Posner et al. (1985), mechanisms other than attentional inhibition are assumed to play an important role.
Recently, the role of response in IOR has become an intriguing issue. Ivanoff and Klein (2001) found that a nonresponding effector on an alternative response key increased IOR, suggesting the involvement of a motoric component. However, IOR may not simply reflect motor inhibition. Using event-related brain potential (ERP) components as indexes, Prime and Ward (2004) suggested that IOR involves perceptual processes rather than inhibition of motor processes. In a later study of long-term and short-term (standard) IOR, Wilson et al. (2006) found delayed response to a target at a location that was previously associated with a no-go stimulus, regardless of whether the temporal interval between the no-go event and the current target was short (1,500 ms) or long (10,500 or 13,500 ms). To account for their findings, rather than suggesting the encoding of the attentional inhibitory effect into the representation of the cue event (e.g., Grison et al., 2005; Kessler & Tipper, 2004; Tipper et al., 2003), Wilson et al. proposed the response retrieval account that the response associated with the cue event is encoded into the memory representations and can produce IOR when these memory representations are retrieved and incompatible with current task demand (e.g., retrieval of a no-go response while the current task demand is a go response). Taken together, IOR may be related to memory retrieval of the representations that involves the integration of response and location but not attentional and response inhibition.
The idea that IOR involves the integration of response and location can fit into a broader theoretical framework: the event files (Hommel, 1998, 2004). Event files can be viewed as an extension of object files (Kahneman et al., 1992), proposing that in addition to feature binding, actions and stimulus features are bound together. In Hommel et al.’s (2001) theory of event coding (TEC), perception and action share common coding and representations. That is, the relationship between perception and action is similar to two sides of one coin. Frings et al. (2020) further proposed the framework of binding and retrieval in action control (BRAC), suggesting that there are two independent processes that contribute to the aftereffect of binding: binding and retrieval. They further suggest that these two processes are modulated by top-down and bottom-up processes independently. According to the theoretical framework based on event files (Frings et al., 2020), it may account for phenomena that involve the influence of a previous event on the current event, such as negative priming (Tipper, 1985; Tipper & Cranston, 1985; see D’Angelo et al., 2016; Fox, 1995; Frings et al., 2015; May et al., 1995; Neill et al., 1995 for reviews) and IOR.
Back to the phenomenon of IOR, according to the framework of event files (Frings et al., 2020; Hommel, 1998, 2004; Hommel et al., 2001), the response, location, and other stimulus features of the cue would be bound together into a representation of the cue event. When there is a retrieval cue for retrieving this representation, such as when the target is presented at the repeated location, this representation will be retrieved (e.g., Frings et al., 2020). Match between the cue and target events will lead to faster performance, and mismatch will lead to slower responses (e.g., Rajsic et al., 2014).
Consistent with this prediction, greater IOR has been observed when the features of the cue were repeated, especially in the cue-target paradigm of IOR (e.g., Hu et al., 2011; Klein et al., 2015; see Huffman et al., 2018, for a review). Hu et al. (2011) and Klein et al. (2015) observed greater IOR when the color was repeated. In addition, Klein et al. demonstrated that repetition of a nonspatial feature (i.e., color in their study) increases IOR in both detection and discrimination tasks but only when the nonspatial feature is task irrelevant.
However, there are also circumstances in which stimulus repetition did not affect IOR, especially in the target-target paradigm of IOR. For instance, it has been suggested that feature integration occurred when the task was discrimination but did not occur when the task was detection or localization in the target-target paradigm of IOR (e.g., Huffman et al., 2018; Taylor & Donnelly, 2002; but see Chao et al., 2020). When the task was detection or localization, repetition of task-irrelevant stimulus features such as form and color did not affect the effect of IOR, especially in the target-target paradigm of IOR (Huffman et al., 2018). That is, IOR remained similar in size regardless of whether the form or color of the cue event was repeated in the target event or not. Similarly, using a prime-probe paradigm, Schöpper et al. (2020) found evidence for feature-response binding in a discrimination task but not in a detection task.
The follow-up question is: does stimulus-response binding exist in a detection task in the target-target paradigm of IOR, and is there any method to reveal (and/or induce) the stimulus-response binding? On the one hand, it is possible that because responses are usually very fast in a detection task (Schöpper et al., 2020), binding or retrieval may occur too late to affect the detection performance. On the other hand, it is also possible that top-down factors such as task demand may affect the integration process in IOR (Frings et al., 2020; Hommel, 2004). When a particular response is varied trial-by-trial in a detection task, responses must be processed because it is highly task relevant. Hence, a stimulus-response binding is more likely to occur.
Therefore, the first issue to be investigated in the present study was whether there is an effect of location-response binding in a detection task of the target-target paradigm of IOR. To achieve this purpose, we combined Hommel’s (1998) procedure with the target-target paradigm of IOR. At the beginning of each trial, the required response when detecting a target was cued and manipulated. Hence, although a detection task was used in the present study, the response to each target could be manipulated independent of the features. Via this design, we can examine the interaction between location and response repetitions in a detection task of the target-target paradigm of IOR. On the one hand, if binding would not occur in a detection task, the interaction between location and response repetitions was not expected. On the other hand, if binding of location and response was possible in a detection task when the response was highly relevant to the task demand, the interaction between location and response repetitions should be observed. In addition, considering Wilson et al.’s (2006) proposal that the IOR effect occurs when retrieved response code is incompatible with the current task demand, and considering the framework of transfer-inappropriate processing and transfer-appropriate processing (Neill, 2007; Neill & Mathis, 1998) that the retrieved memory episodes can lead to a response cost when they are incompatible with the current task setting and a response benefit when they are compatible (Lupiáñez et al., 2013), we expected a repetition benefit of location when the response was repeated and a repetition cost of location when the response was not repeated.
The second issue is the temporal requirement of memory retrieval of the cue representations. In order to explain the absence of the binding effect in a detection task, Schöpper et al. (2020) suggested that detection processes may occur too fast for memory retrieval to manifest. In this case, it is possible that the aftereffect of binding may take place when there is more time for memory retrieval to occur and affect the performance. Therefore, we analyzed the Vincentized cumulative RT distribution (Ratcliff, 1979) to investigate the temporal requirement of memory retrieval of the cue representations. To perform this analysis, the RTs in each condition were sorted into different RT bins, from the shorted to the longest. If memory retrieval of the cue representations was time invariant, no interaction between RT bins and the cuing effect was expected. On the other hand, if memory retrieval was time dependent (e.g., Schöpper et al., 2020), an interaction between RT bins and the cuing effect was expected, and the cuing effect should be larger in the longer RT bins.
The final issue is the effect of repeating a nonspatial and task-irrelevant feature on IOR. On the one hand, considering previous findings of Huffman et al. (2018) and Schöpper et al. (2020), the repetition of a nonspatial and task-irrelevant feature should not affect IOR when the task is detection in a target-target paradigm. This is consistent with Memelink and Hommel’s (2013) proposal of the intentional weighting mechanism – task-relevant features should receive larger weight than task-irrelevant features. In addition, Hommel et al. (2014) further showed that task demand affected which information was retrieved from the event files. Hence, it is expected that a nonspatial and task-irrelevant feature should be less likely to be processed and encoded into/retrieved from the cue representation and therefore less likely to affect IOR. On the other hand, if the findings in the cue-target paradigm (e.g., Hu et al., 2011; Klein et al., 2015) could be applied to the target-target paradigm, the repetition of a task-irrelevant feature might modulate IOR. For instance, Chao et al. (2020) used a detection task in a target-target paradigm and observed larger spatial IOR when the color was repeated. In the present study, therefore, we also manipulated nonspatial feature (form) repetition to examine the effect of repeating a nonspatial task-irrelevant feature on IOR. If all the features of the cue were integrated into the cue representations, the effect of cue repetition should interact with location repetition and response repetition. On the other hand, if the task demand modulated encoding and binding, the repetition of the form should not affect the performance.
Taken together, in order to investigate these three issues, we used a target-target paradigm of IOR with a location repetition × response repetition × form repetition design. At the beginning of each trial, the required response upon detecting the target was cued and manipulated. Hence, we manipulated the repetition of location, required response, and form of the target orthogonally in the present study.
Method
Participants
A total of 28 undergraduate students (21 females and seven males; average age = 22.2 years) at Chung Yuan Christian University participated in this experiment and received NT$300 as compensation. They had normal or corrected-to-normal vision. Each participant read and signed an informed consent form before the start of the experiment.
Stimuli
Three boxes were presented in white as placeholders. At a viewing distance of approximately 57 cm, each box subtended a width of 3.0° and a height of 2.0°, with a border thickness of 0.1°. The three boxes were presented at the center of the screen, aligned on the horizontal meridian. The center-to-center distance between adjacent boxes was 4.8°. The central box was cued at the beginning of each trial by increasing the width of that box outline to 0.3°; at the same time, a Chinese word (left or right) was presented in the center of the central box as a response cue, referring either to the left button or the right button. The height and width of the Chinese word was 0.8°. The target was either a rectangle or an oval presented in the center of either the left or the right box. The length and width of the rectangle and oval were 1.1° and 0.7° and 1.1° and 0.7°, respectively. The color of the target was always yellow, and the orientation of the target was always vertical.
Procedure
The experiment was conducted using DMDX software (Forster & Forster, 2003). As a target-target paradigm of IOR, couplets of two successive trials were used. In test couplets, a target was presented in both the first and the second trials. In catch couplets, the target was absent in the first trial, in the second trial, or in both trials. The target in the first trial served as the cue, and performance in the second trial could reveal an IOR effect. The location (left or right), required response (a left or a right button), and form (oval or rectangle) of each target were manipulated, leading to a location repetition (location repeated/unrepeated) × form repetition (form repeated/unrepeated) × response repetition (response repeated/unrepeated) design.
There was a practice block of 12 test couplets and three catch couplets at the beginning of the experiment, followed by 384 test couplets and 96 catch couplets. In the location repeated condition, the target of the second trial was presented at the location of the target of the first trial; in the location unrepeated condition, the target of the second trial was presented at the other location. In the form repeated condition, the form of the target in the second trial was identical to that in the first trial; and the two targets in one couplet were different in the form unrepeated. Finally, the same responses were required for the two targets of a couplet in the same response condition, while different responses were required in the different response condition. Location, form, and response repetitions were manipulated orthogonally. There were equal couplets (48 couplets) in each combination of these three variables.
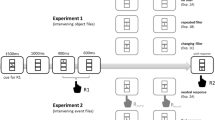
There were two trials in a couplet (see Fig. 1 for an example). Each trial began with the display of three placeholders for 200 ms. Then a 1,200-ms fixation cue was presented in the central box. At the same time, a response cue indicating which response to make upon detecting the presence of a target was presented in this central box. After the offset of the central and response cues, the placeholders were presented on the screen for another 200 ms. Then a target was presented in either the left or the right box or no target in catch trials. This target was presented for 1,000 ms or until a response was made. Participants were instructed to press the button of the mouse specified by the response cue when they saw a target, regardless of its form and location. They pressed the left button of the mouse with their index finger and the right button of the mouse with their middle finger. There were rest breaks after each block of 30 couplets.

An example of a couplet (not to scale). Note. There were two trials in each couplet. Each trial began with the display of three placeholders for 200 ms. A 1,200-ms fixation cue was then presented in the central box by increasing the width of that box outline. At the same time, a response cue indicating which response to make when detecting a target was presented in the central box. The response cue was presented in Chinese in the actual experiment. After the offset of the fixation cue, the placeholders were presented on the screen for 200 ms. A target was then presented in either the left or right box, or no target was presented
Results
The response to the target in the second trial of each couplet was included and analyzed only if the response to the target in the first trial of the same couplet was correct. In sum, 1.0% of the data were excluded because of errors of responding to the target in the first trial.
Analysis of the overall data
Table 1 shows the averaged median correct RTs and error rates in each condition. Both the RT and error rate data were analyzed by a 2 (location repetition: repeated/unrepeated) × 2 (form repetition: repeated/unrepeated) × 2 (response repetition: repeated/unrepeated) repeated-measures analysis of variance (ANOVA).
Analysis of the RT data showed a significant main effect of response repetition [F (1, 27) = 10.85, p < .01, \( {\eta}_{\mathrm{p}}^2 \) = .29]. More importantly, the interaction between location and response repetitions was significant [F (1, 27) = 27.56, p < .001, \( {\eta}_{\mathrm{p}}^2 \) = .51], indicating that location repetition produced a significant repetition benefit when the response was repeated [10 ms; F (1, 54) = 23.26, p < .001, \( {\eta}_{\mathrm{p}}^2 \) = .30] and a significant repetition cost when the response was not repeated (-22 ms; F [1, 54] = 16.00, p < .001, \( {\eta}_{\mathrm{p}}^2 \) = .23). The effects involving form repetition were not significant, including the main effect of form [F (1, 27) = 0.02, p = .598, \( {\eta}_{\mathrm{p}}^2 \) = .00], interaction between location and form repetitions [F (1, 27) = 0.42, p = .528, \( {\eta}_{\mathrm{p}}^2 \) = .02], interaction between form and response repetitions [F (1, 27) = 1.56, p = .221, \( {\eta}_{\mathrm{p}}^2 \) = .05], and three-way interaction of all three variables [F (1, 27) = 0.46, p = .509, \( {\eta}_{\mathrm{p}}^2 \) = .02].
Analysis of error rates revealed a significant interaction between location and response repetitions [F (1, 27) = 14.85, p < .001, \( {\eta}_{\mathrm{p}}^2 \) = .35]. This indicated that location repetition produced a significant repetition benefit when the response was repeated [1.0%; F (1, 54) = 6.68, p < .05, \( {\eta}_{\mathrm{p}}^2 \) = .11] and a significant repetition cost when the response was not repeated [-0.9%; F (1, 54) = 11.30, p < .01, \( {\eta}_{\mathrm{p}}^2 \) = .17].
Analysis of data as a function of RT bins
To analyze the RT data as a function of RT bins, the RT data were further analyzed by a 2 (location repetition: repeated/unrepeated) × 2 (form repetition: repeated/unrepeated) × 2 (response repetition: repeated/unrepeated) × 8 (RT bin: 12.5%/25%/37.5/50%/62.5%/75%/87.5%/100%) repeated-measures ANOVA. Table 2 shows the data in all conditions. To simplify data illustration, Fig. 2 shows the effect of location repetition (unrepeated-location RT – repeated-location RT) as a function of form repetition, response repetition, and RT bins. Because there were too few errors in each condition, error rates were not analyzed.

The effect of location repetition as a function of form repetition, response repetition, and RT bin. Note. The effect of location repetition = unrepeated-location RT – repeated-location RT. Error bars represent the 95% confidence interval based on Jarmasz and Hollands’s (2009) method. The MSE was based on an ANOVA of the effect of location repetition
Analysis of RT data as a function of RT bins also showed a significant main effect of response repetition [F (1, 27) = 18.59, p < .001, \( {\eta}_{\mathrm{p}}^2 \) = .41] and a significant interaction between location and response repetitions [F (1, 27) = 47.04, p < .001, \( {\eta}_{\mathrm{p}}^2 \) = .64]. In addition, the main effect of RT bin was significant [F (7, 189) = 214.50, p < .001, \( {\eta}_{\mathrm{p}}^2 \) = .89]. The interaction between response repetition and RT bin was also significant [F (7, 189) = 5.46, p < .001, \( {\eta}_{\mathrm{p}}^2 \) = .17]. More importantly, the interaction of location repetition, response repetition, and RT bin was significant [F (7, 189) = 17.13, p < .001, \( {\eta}_{\mathrm{p}}^2 \) = .39]. This three-way interaction suggested that the benefit of location repetition when the response was repeated was significant in the 37.5% [13 ms; F [(1, 432) = 7.37, p < .01, \( {\eta}_{\mathrm{p}}^2 \) = .02], 50% [17 ms; F (1, 432) = 11.31, p < .01, \( {\eta}_{\mathrm{p}}^2 \)= .03], 62.5% [19 ms; F (1, 432) = 14.22, p < .001, \( {\eta}_{\mathrm{p}}^2 \) = .03], 75% [23 ms; F (1, 432) = 21.92, p < .001, \( {\eta}_{\mathrm{p}}^2 \) = .05], 87.5% [27 ms; F (1, 432) = 30.26, p < .001, \( {\eta}_{\mathrm{p}}^2 \) = .07], and 100% [44 ms; F (1, 432) = 77.28, p < .01, \( {\eta}_{\mathrm{p}}^2 \) = .15] RT bins. The repetition benefit increased as a function of RT bins. Moreover, the cost of location repetition when the response was not repeated was significant in the 50% [-12 ms; F (1, 432) = 5.61, p < .01, \( {\eta}_{\mathrm{p}}^2 \) = .01], 62.5% [-16 ms; F (1, 432) = 10.20, p < .01, \( {\eta}_{\mathrm{p}}^2 \) = .02], 75% [-18 ms; F (1, 432) = 13.72, p < .001, \( {\eta}_{\mathrm{p}}^2 \) = .03], 87.5% [-21 ms; F (1, 432) = 18.47, p < .001, \( {\eta}_{\mathrm{p}}^2 \) = .04], and 100% [-37 ms; F (1, 432) = 55.78, p < .001, \( {\eta}_{\mathrm{p}}^2 \) = .11] RT bins. The repetition cost also increased as a function of RT bins.
Discussion
The present study investigated the effect of location-response binding in a detection task of the target-target paradigm of IOR by integrating Hommel’s (1998) response-cuing procedure with the target-target paradigm of IOR. Results provided clear evidence for location-response binding: location repetition produced IOR (i.e., cost of location repetition) when the response was not repeated and a facilitation of return (FOR; i.e., benefit of location repetition) when the response was repeated. It should be noted that the FOR observed in the present study was not simply the main facilitatory effect of response repetition (e.g., Welsh & Pratt, 2006). Because location and response repetitions were manipulated orthogonally in the present study, FOR was observed only when the response was also repeated but not when the response was not repeated. Hence, it is important to propose a mechanism that binds the location and response together.
The interaction of location and response repetitions was consistent with the frameworks of event files (Frings et al., 2020; Hommel, 1998, 2004; Hommel et al., 2001), suggesting that the spatial location of a target and the response to it are bound together into an event file, and retrieval of the previous file can modulate the response. On the one hand, when the target matches the retrieved representation (i.e., the same location and response), the response to it is facilitated. On the other hand, when the target partially matches the retrieved representation (i.e., a cue for memory retrieval but the retrieved episode is not fully compatible), the response to it is slowed (Milliken et al., 1998, 2000). The findings that memory retrieval of previous memory episodes led to IOR or FOR, depending on the match between the retrieved memory representations and the current task demand, is consistent with the framework of transfer-inappropriate processing and transfer-appropriate processing (Neill, 2007; Neill & Mathis, 1998; see also Lupiáñez et al., 2013).
Recently, Hommel and Frings (2020) showed that the event files decay over time, lasting for about 5 s. In the present study, after the offset of the target of the first trial, there was a 1,600-ms temporal interval before the onset of the target of the second trial. Hence, the inter-stimulus interval between the targets of the first and second trials was appropriate for observing the impact of the event files. However, it should be noted that Wilson et al. (2006) found an IOR effect when the temporal intervals were 1,500, 10,500, and 13,500 ms and proposed the role of response retrieval. It was possible that short-term (1,500 ms) and long-term (10,500 or 13,500 ms) IOR may involve different representations, and it was also possible that the event files may last longer than 5 s or be transformed into more durable long-term representations in certain contexts.
It should be noted that, however, the frameworks of event files may not account for all IOR-related phenomena, or a cost of responding to a stimulus presented at a previously cued location. For instance, Hilchey et al. (2018) demonstrated that the binding effect manifested when the participants made manual discrimination responses to the targets, and inhibited reorientation to the target manifested when the participants localized the targets by eye movements. The present study further added that the binding effect could also manifest in a detection task. Considering that IOR may involve multiple mechanisms (e.g., Hilchey et al., 2018; Taylor & Klein, 2000; Wang et al., 2012), future study is required to reveal the effects of location and response bindings on sensory and motor components of IOR.
Regarding the reason for lack of binding effect in a detection task, Schöpper et al. (2020) suggested a possibility that the detection task operates too fast for memory retrieval of the previous episode to occur. Analysis of the Vincentized cumulative RT distribution (Ratcliff, 1979) supported Schöpper et al.’s (2020) suggestion. The IOR and FOR were observed only when the RTs were long. These findings suggested that it took time for memory retrieval of the previous episode to affect the performance.
Another consideration regarding the binding effect in a detection task in the target-target paradigm of IOR manifested in the present study but not in previous studies (e.g., Huffman et al., 2018) is the requirement of processing the response cues. That is, in order to learn what kind of response is required when detecting the target, visual discrimination of the response cues and response preparation for the target were required in the present study, trial by trial. If this task set for response cue processing was transferred to target processing, in comparison to previous studies in which a simple detection task was used, the present study may involve more discrimination processing. Although the lack of the effect of form repetition did not support the idea that the task set for response-cue processing, such as form discrimination, was transferred to target processing, this issue still deserves further investigation. In addition, considering that processing the response cues could be viewed as part of the task demand in the present study, the possibility that the response cue and target were integrated into event files also requires further investigation, such as manipulating the retrieval cues provided by either the response cues or the targets.
Moreover, regarding the effect of repeating a task-irrelevant nonspatial feature, our results showed that form repetition did not produce any significant effect. One interpretation regarding the null effect of form repetition was that the forms used in the present study were ineffective. However, because the effect of form repetition with the same set of visual forms had been observed in our laboratory (e.g., Chao, 2017), this interpretation was less likely. Instead, our findings are consistent with previous findings of Huffman et al. (2018) and Schöpper et al. (2020) and the account of the intentional weighting mechanism (Memelink & Hommel, 2013), supporting the idea that task demand affects what is encoded into the event files and/or what is retrieved from the event files (Frings et al., 2020; Hommel, 2004; Hommel et al., 2014), and task-irrelevant features are hence less likely to be encoded.
However, there are findings suggesting that task-irrelevant nonspatial features may be encoded into the representations of the target event or cue event, such as Kwak and Egeth’s (1992) Experiment 2 and Chao et al. (2020) in a target-target paradigm of IOR and Hu et al. (2011) and Klein et al. (2015) in a cue-target paradigm of IOR. A possible interpretation is that task demand (Frings et al., 2020; Hommel, 2004) and resource limitation (e.g., Lavie et al., 2004) play important roles. In the cue-target paradigm, because there is no specific response requirement for processing the cue, task-irrelevant features are more likely to be processed and encoded. In contrast, in the target-target paradigm, specific response is required for the first target; hence, attention is directed toward specific feature and response and less likely toward task-irrelevant features. Therefore, the integration of task-irrelevant nonspatial features and spatial locations is more likely to occur in the cue-target paradigm of IOR but less likely in the target-target paradigm. In addition, in comparison to Kwak and Egeth (1992) and Chao et al. (2020) in which the repetition of only two features (color and location) was manipulated, the repetition of three features (form, location, and response) was manipulated in the present study. It is possible that owing to limited attentional resource, when there are many features varied, a limited set of features can be attended and encoded. Therefore, visual form was not encoded in the present study.
Finally, in the present study, because the participants pressed either the left or the right button when detecting a target at either the left or the right location, the Simon effect (Simon, 1969; faster response to a target when it is presented at a location congruent with the direction of the response) may occur. Moreover, it has been demonstrated that the Simon effect may interact with the IOR effect (e.g., Hilchey et al., 2011; Ivanoff et al., 2002). Hence, it is plausible that the Simon effect may interact with the effect of bindings of location and response. In order to examine this possibility, we reanalyzed the data by including the congruency between the location and response associated with the target of the second trial as the fourth variable. A 2 (location repetition: repeated/unrepeated) × 2 (form repetition: repeated/unrepeated) × 2 (response repetition: repeated/unrepeated) × 2 (congruency: congruent/incongruent) repeated-measures ANOVA revealed a significant main effect of congruency, F (1, 27) = 34.96, p < .01, \( {\eta}_{\mathrm{p}}^2 \) = .56, revealing faster responses in the congruent condition (285 ms) than in the incongruent condition (303 ms). Moreover, the three-way interaction of location repetition, response repetition, and congruency was significant, F (1, 27) = 7.55, p < .05, \( {\eta}_{\mathrm{p}}^2 \) = .22. Follow-up analysis revealed that although there was a cost of location repetition when the response was not repeated regardless of congruency, this IOR effect was larger in the incongruent condition [-22 ms; F (1, 108) = 24.27, p < .01, \( {\eta}_{\mathrm{p}}^2 \) = .18] than in the congruent condition [-10 ms; F (1, 108) = 5.06, p < .05, \( {\eta}_{\mathrm{p}}^2 \) = .05]. Similarly, there was a benefit of location repetition when the response was repeated, this FOR effect was larger in the incongruent condition [25 ms; F (1, 108) = 32.36, p < .01, \( {\eta}_{\mathrm{p}}^2 \) = .23] than in the congruent condition [11 ms; F (1, 108) = 5.96, p < .05, \( {\eta}_{\mathrm{p}}^2 \) = .05]. Considering the time course of memory retrieval of the event files (Schöpper et al., 2020), it was possible that the Simon effect delayed the responses and hence allowed better memory retrieval and larger effect of the previous binding between the location and response.
An alternative explanation regarding this three-way interaction was that the IOR effect modulated the processing of the task-relevant and/or -irrelevant codes (e.g., Hilchey et al., 2011; Ivanoff et al., 2002) and hence increased the Simon effect. Reanalysis of the data of the present study suggested an effect of previous cue representations or event files on the Simon effect. In brief, the Simon effect was greater when there was a partial repetition of either the location [23 ms; F (1, 108) = 25.57, p < .01, \( {\eta}_{\mathrm{p}}^2 \) = .19] or the response [28 ms; F (1, 108) = 40.67, p < .01, \( {\eta}_{\mathrm{p}}^2 \) = .27] than when there was full repetition [14 ms; F (1, 108) = 9.91, p < .01, \( {\eta}_{\mathrm{p}}^2 \) = .08] or no repetition [11 ms; F (1, 108) = 5.74, p < .01, \( {\eta}_{\mathrm{p}}^2 \) = .05] of the location and response. These results might suggest that the partial match and mismatch between the previous cue representations and the current target may delayed the target processing (e.g., Milliken et al., 1998, 2000) and hence increased the Simon effect. The impacts of the previous event files on the relevant and irrelevant information processing in the Simon task requires further investigation.
In summary, we investigated the effect of location-response binding in a detection task of the target-target paradigm of IOR. The findings of the present study are more consistent with a more general theoretical framework – the event files (Frings et al., 2020; Hommel, 1998, 2004; Hommel et al., 2001) – and suggest that IOR and FOR may involve a general mechanism.
References
Chao, H.-F. (2017). The effects of location repetition and shape repetition in a target-target paradigm: A reaction time distribution analysis. Poster presented at the 58th Annual Meeting of the Psychonomic Society, Vancouver, British Columbia, Canada.
Chao, H.-F., Kuo, C.-Y., Chen, M. S., & Hsiao, F.-S. (2020). Contextual similarity between successive targets modulates inhibition of return in the target-target paradigm. Frontiers in Psychology, 11, 2052. https://doi.org/10.3389/fpsyg.2020.02052
D’Angelo, M. C., Thomson, D. R., Tipper, S. P., & Milliken, B. (2016). Negative priming 1985 to 2015: A measure of inhibition, the emergence of alternative accounts, and the multiple process challenge. Quarterly Journal of Experimental Psychology, 69, 1890–1909. https://doi.org/10.1080/17470218.2016.1173077
Dukewich, K. (2009). Reconceptualizing inhibition of return as habituation of the orienting response. Psychonomic Bulletin & Review, 16, 238–251. https://doi.org/10.3758/PBR.16.2.238
Fox, E. (1995). Negative priming from ignored distractors in visual selection: A review. Psychonomic Bulletin & Review, 2, 145–173. https://doi.org/10.3758/BF03210958
Forster, K. I., & Forster, J. C. (2003). DMDX: A windows display program with millisecond accuracy. Behavior Research Methods, Instruments, & Computers, 35, 116–124. https://doi.org/10.3758/BF03195503
Frings, C., Hommel, B., Koch, I., Rothermund, K., Dignath, D., Giesen, C., Kiesel, A., Kunde, W., Mayr, S., Moeller, B., Möller, M., Pfister, R., & Philipp, A. (2020). Binding and retrieval in action control (BRAC). Trends in Cognitive Sciences, 24, 375–387. https://doi.org/10.1016/j.tics.2020.02.004
Frings, C., Schneider, K. K., & Fox, E. (2015). The negative priming paradigm: An update and implications for selective attention. Psychonomic Bulletin & Review, 22, 1577–1597. https://doi.org/10.3758/s13423-015-0841-4
Grison, S., Paul, M. A., Kessler, K., & Tipper, S. P. (2005). Inhibition of object identity in inhibition of return: Implications for encoding and retrieving inhibitory processes. Psychonomic Bulletin & Review, 12, 553–558. https://doi.org/10.3758/BF03193804
Hilchey, M. D., Ivanoff, J., Taylor, T. L., & Klein, R. M. (2011). Visualizing the temporal dynamics of spatial information processing responsible for the Simon effect and its amplification by inhibition of return. Acta Psychologica, 136, 235–244. https://doi.org/10.1016/j.actpsy.2010.09.003
Hilchey, M. D., Rajsic, J., Huffman, G., Klein, R. M., & Pratt, J. (2018). Dissociating orienting biases from integration effects with eye movements. Psychological Science, 29, 328–339. https://doi.org/10.1177/0956797617734021
Hommel, B. (1998). Event files: Evidence for automatic integration of stimulus–response episodes. Visual Cognition, 5, 183–216. https://doi.org/10.1080/713756773
Hommel, B. (2004). Event files: Feature binding in and across perception and action. Trends in Cognitive Sciences, 8, 494–500. https://doi.org/10.1016/j.tics.2004.08.007
Hommel, B., & Frings, C. (2020). The disintegration of event files over time: Decay or interference? Psychonomic Bulletin & Review, 27, 751–757. https://doi.org/10.3758/s13423-020-01738-3
Hommel, B., Memelink, J., Zmigrod, S., & Colzato, L. S. (2014). Attentional control of the creation and retrieval of stimulus–response bindings. Psychological Research, 78(4), 520-538. https://doi.org/10.1007/s00426-013-0503-y
Hommel, B., Müsseler, J., Aschersleben, G., & Prinz, W. (2001). The Theory of Event Coding (TEC): A framework for perception and action planning. Behavioral and Brain Sciences, 24, 849–878. https://doi.org/10.1017/S0140525X01000103
Hu, F. K., Samuel, A. G., & Chan, A. S. (2011). Eliminating inhibition of return by changing salient nonspatial attributes in a complex environment. Journal of Experimental Psychology: General, 140, 35–50. https://doi.org/10.1037/a0021091
Huffman, G., Hilchey, M. D., & Pratt, J. (2018). Feature integration in basic detection and localization tasks: Insights from the attentional orienting literature. Attention, Perception, & Psychophysics, 80, 1333–1341. https://doi.org/10.3758/s13414-018-1535-6
Ivanoff, J., & Klein, R. M. (2001). The presence of a nonresponding effector increases inhibition of return. Psychonomic Bulletin & Review, 8, 307–314. https://doi.org/10.3758/BF03196166
Ivanoff, J., Klein, R. M., & Lupiáñez, J. (2002). Inhibition of return interacts with the Simon effect: An omnibus analysis and its implications. Perception & Psychophysics, 64, 318–327. https://doi.org/10.3758/BF03195794
Jarmasz, J. & Hollands, J. (2009). Confidence intervals in repeated-measures designs: The number of observations principle. Canadian Journal of Experimental Psychology, 63, 124–318. https://doi.org/10.1037/a0014164
Kahneman, D., Treisman, A., & Gibbs, B. J. (1992). The reviewing of object files: Object-specific integration of information. Cognitive psychology, 24, 175–219. https://doi.org/10.1016/0010-0285(92)90007-O
Kessler, K., & Tipper, S. P. (2004). Retrieval of implicit inhibitory processes: The impact of visual field, object-identity, and memory dynamics. Visual Cognition, 11, 965–995. https://doi.org/10.1080/13506280444000012a
Klein, R. M. (2000). Inhibition of return. Trends in Cognitive Sciences, 4, 138–147. https://doi.org/10.1016/S1364-6613(00)01452-2
Klein, R. M., Wang, Y., Dukewich, K. R., He, S., & Hu, K. (2015). On the costs and benefits of repeating a nonspatial feature in an exogenous spatial cuing paradigm. Attention, Perception, & Psychophysics, 77, 2293–2304. https://doi.org/10.3758/s13414-015-0941-2
Kwak, H. W., & Egeth, H. (1992). Consequences of allocating attention to locations and to other attributes. Perception & Psychophysics, 51, 455–464. https://doi.org/10.3758/BF03211641
Lavie, N., Hirst, A., De Fockert, J. W., & Viding, E. (2004). Load theory of selective attention and cognitive control. Journal of Experimental Psychology: General, 133, 339–354. https://doi.org/10.1037/0096-3445.133.3.339
Lupiáñez, J., Martín-Arévalo, E., & Chica, A. B. (2013). Is inhibition of return due to attentional disengagement or to a detection cost? The detection cost theory of IOR. Psicológica, 34, 221–252.
Lupiáñez, J., Klein, R. M., & Bartolomeo, P. (2006). Inhibition of return: Twenty years after. Cognitive Neuropsychology, 23, 1003–1014. https://doi.org/10.1080/02643290600588095
Lupiáñez, J., Ruz, M., Funes, M. J., & Milliken, B. (2007). The manifestation of attentional capture: Facilitation or IOR depending on task demands. Psychological Research, 71, 77–91. https://doi.org/10.1007/s00426-005-0037-z
Lupiáñez, J. (2010). Inhibition of return. In A. C. Nobre & J. T. Coull (Eds.), Attention and time (pp. 17–34). Oxford, UK: Oxford University Press.
Martín-Arévalo, E., Kingstone, A., & Lupiáñez, J. (2013). Is “inhibition of return” due to the inhibition of the return of attention? Quarterly Journal of Experimental Psychology, 66, 347–359. https://doi.org/10.1080/17470218.2012.711844
May, C. P., Kane, M. J., & Hasher, L. (1995). Determinants of negative priming. Psychological Bulletin, 118, 35–54. https://doi.org/10.1037/0033-2909.118.1.35
Memelink, J., & Hommel, B. (2013). Intentional weighting: A basic principle in cognitive control. Psychological Research, 77, 249–259. https://doi.org/10.1007/s00426-012-0435-y
Milliken, B., Joordens, S., Merikle, P. M., & Seiffert, A. E. (1998). Selective attention: A reevaluation of the implications of negative priming. Psychological Review, 105, 203–229. https://doi.org/10.1037/0033-295X.105.2.203
Milliken, B., Tipper, S. P., Houghton, G., & Lupiáñez, J. (2000). Attending, ignoring, and repetition: On the relation between negative priming and inhibition of return. Perception & Psychophysics, 62, 1280–1296. https://doi.org/10.3758/BF03212130
Neill, W. T. (2007). Mechanisms of transfer-inappropriate processing. In D. S. Gorfein & C. M. MacLeod (Eds.), Inhibition in cognition (pp. 63–78). Washington, DC: American Psychological Association. https://doi.org/10.1037/11587-004
Neill, W. T., & Mathis, K. M. (1998). Transfer-inappropriate processing: Negative priming and related phenomena. In D. L. Medin (Ed.), The psychology of learning and motivation: Advances in research and theory (Vol. 38, pp. 1–44). San Diego, CA: Academic Press. https://doi.org/10.1016/S0079-7421(08)60182-6
Neill, W. T., Valdes, L. A., & Terry, K. M. (1995). Selective attention and the inhibitory control of cognition. In F. N. Dempster & C. J. Brainerd (Eds.), Interference and inhibition in cognition (pp. 207–261). New York, NY: Academic Press. https://doi.org/10.1016/B978-012208930-5/50008-8
Posner, M. I., & Cohen, Y. (1984). Components of visual orienting. In H. Bouma & D. G. Bouwhuis (Eds.), Attention and performance X: Control of language processes (pp. 531–556). Hilldale, NJ: Erlbaum.
Posner, M. I., Rafal, R. D., Choate, L. S., & Vaughan, J. (1985). Inhibition of return: Neural basis and function. Cognitive Neuropsychology, 2, 211–228. https://doi.org/10.1080/02643298508252866
Prime, D. J., & Ward, L. M. (2004). Inhibition of return from stimulus to response. Psychological Science, 15, 272–276. https://doi.org/10.1111/j.0956-7976.2004.00665.x
Rajsic, J., Bi, Y., & Wilson, D. E. (2014). Long-term facilitation of return: A response-retrieval effect. Psychonomic Bulletin & Review, 21, 418–424. https://doi.org/10.3758/s13423-013-0502-4
Ratcliff, R. (1979). Group reaction time distribution and an analysis of distribution statistics. Psychological Bulletin, 86, 446−461. https://doi.org/10.1037/0033-2909.86.3.446
Schöpper, L. M., Hilchey, M. D., Lappe, M., & Frings, C. (2020). Detection versus discrimination: The limits of binding accounts in action control. Attention, Perception, & Psychophysics, 82, 2085–2097. https://doi.org/10.3758/s13414-019-01911-4
Simon, J. R. (1969). Reactions toward the source of stimulation. Journal of Experimental Psychology, 81, 174−176. https://doi.org/10.1037/h0027448
Taylor, T. L., & Donnelly, M. P. W. (2002). Inhibition of return for target discriminations: The effect of repeating discriminated and irrelevant stimulus dimensions. Perception & Psychophysics, 64, 292–317. https://doi.org/10.3758/BF03195793
Taylor, T. L., & Klein, R. M. (2000). Visual and motor effects in inhibition of return. Journal of Experimental Psychology: Human Perception and Performance, 26, 1639−1656. https://doi.org/10.1037/0096-1523.26.5.
Tipper, S. P. (1985). The negative priming effect: Inhibitory priming by ignored objects. Quarterly Journal of Experimental Psychology, 37A, 571–590. https://doi.org/10.1080/14640748508400920
Tipper, S. P., & Cranston, M. (1985). Selective attention and priming: Inhibitory and facilitatory effects of ignored primes. Quarterly Journal of Experimental Psychology, 37A, 591–611. https://doi.org/10.1080/14640748508400921
Tipper, S. P., Grison, S., & Kessler, K. (2003). Long-term inhibition of return of attention. Psychological Science, 14, 19–25. https://doi.org/10.1111/1467-9280.01413
Wang, Z., Satel, J., & Klein, R. M. (2012). Sensory and motor mechanisms of oculomotor inhibition of return. Experimental Brain Research, 218, 441–453. https://doi.org/10.1007/s00221-012-3033-8
Welsh, T. N., & Pratt, J. (2006). Inhibition of return in cue-target and target-target tasks. Experimental Brain Research, 174, 167–175. https://doi.org/10.1007/s00221-006-0433-7
Wilson, D. E., Castel, A. D., & Pratt, J. (2006). Long-term inhibition of return for spatial locations: Evidence for a memory retrieval account. Quarterly Journal of Experimental Psychology, 59, 2135–2147. https://doi.org/10.1080/17470210500481569
Author Note
This work was supported by the Ministry of Science and Technology of Taiwan under Grant MOST 105-2410-H-033-013-MY3 to Hsuan-Fu Chao. Portions of these findings were presented at the 31st Annual Convention of the Association for Psychological Science. We have no conflicts of interest to disclose.
Author information
Authors and Affiliations
Corresponding author
Additional information
Open Practices Statements
The data and materials for this experiment are available upon request and this experiment was not preregistered.
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Chao, HF., Hsiao, FS. Location-response binding and inhibition of return in a detection task. Atten Percept Psychophys 83, 1992–2001 (2021). https://doi.org/10.3758/s13414-021-02283-4
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13414-021-02283-4