
Response of Gross Mineralization and Nitrification Rates to Banana Cultivation Sites Converted from Natural Forest in Subtropical China

Abstract
:1. Introduction
2. Material and Methods
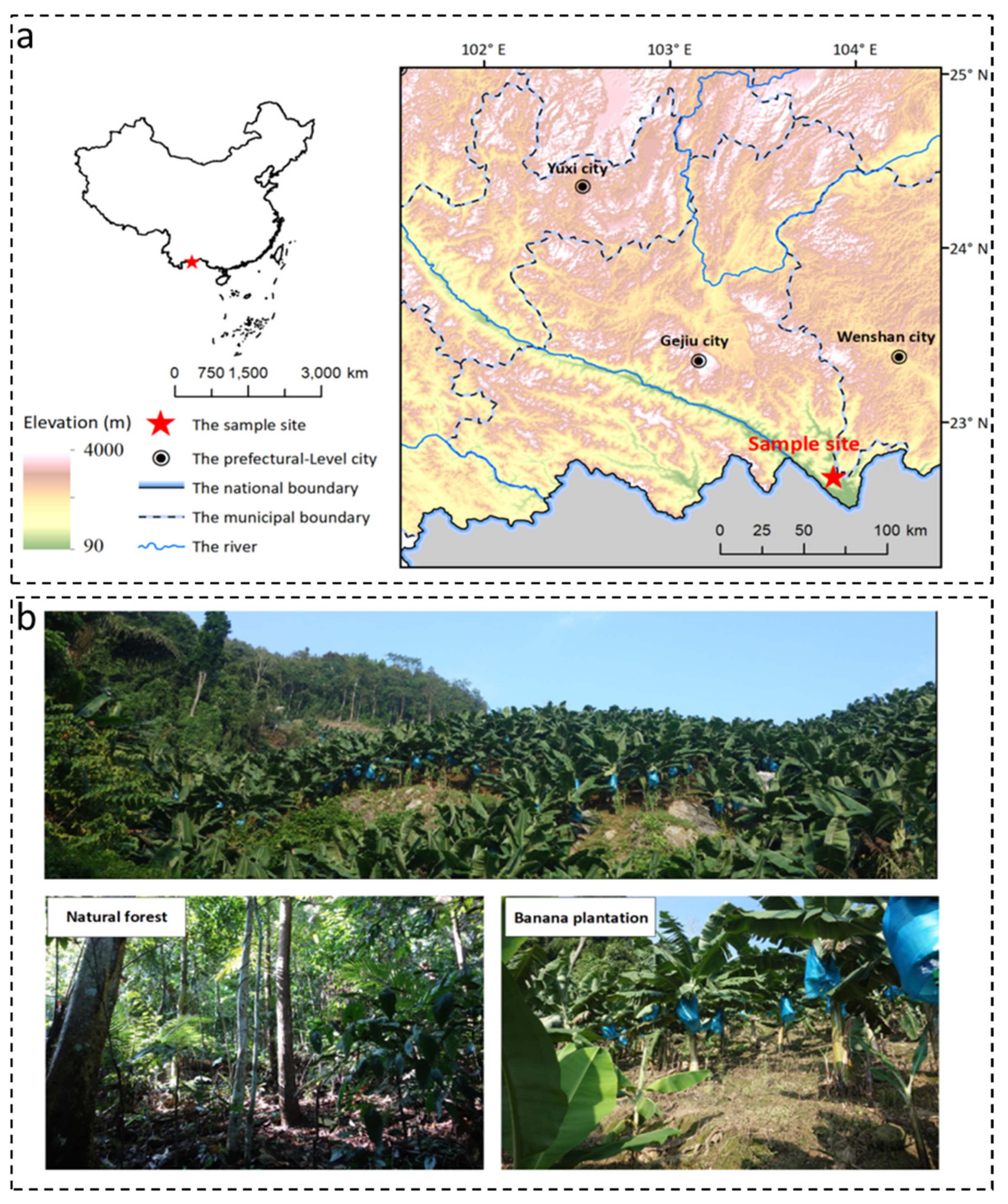
2.1. Site Description and Sample Collection
2.2. Gross Mineralization and Nitrification Rates
2.3. Analyses
2.4. Bacterial or Archaeal amoA Abundance Analysis
2.5. Data and Statistical Analyses
3. Results
3.1. Soil Physical and Chemical Properties, AOA and AOB Abundances
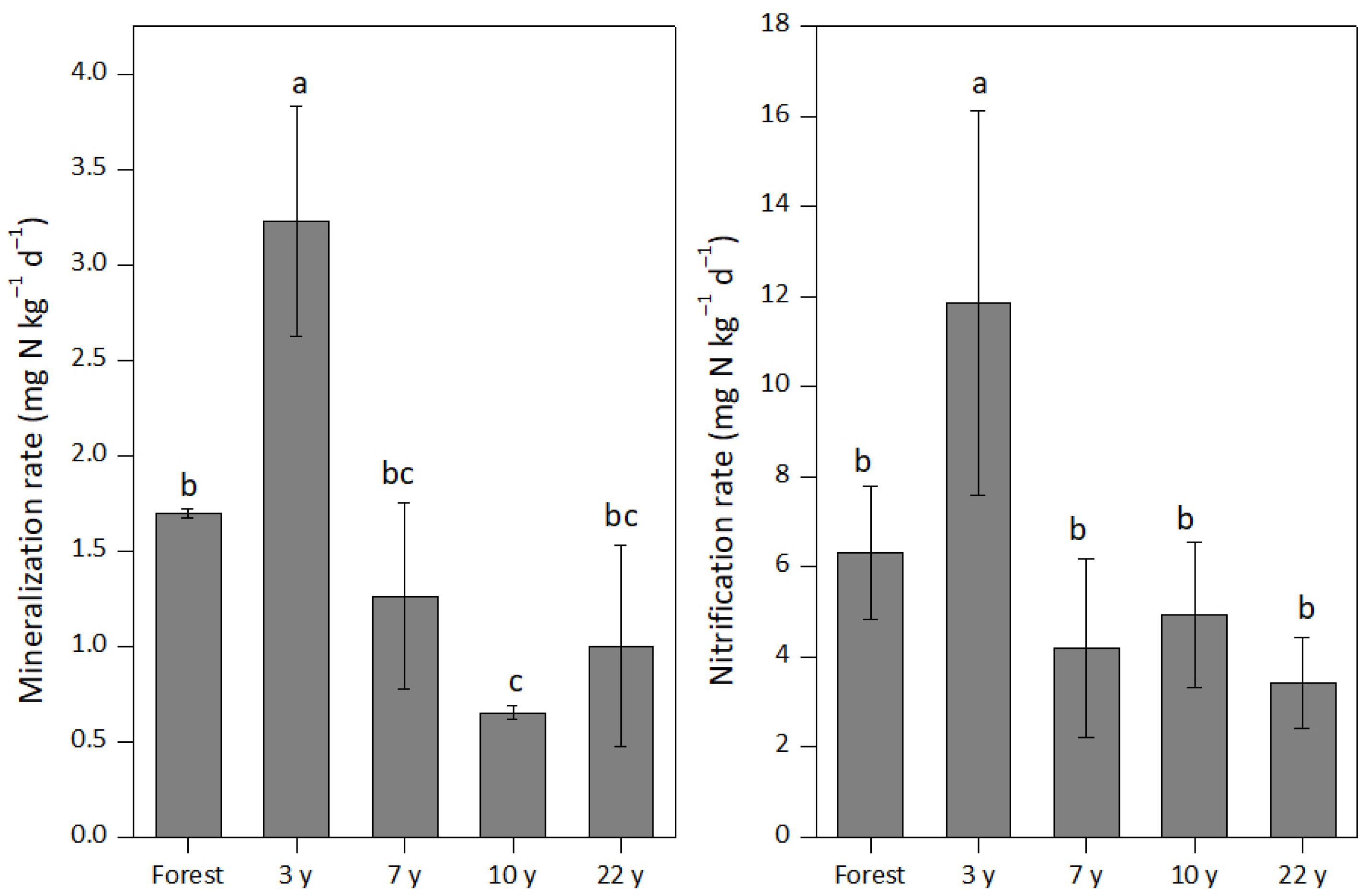
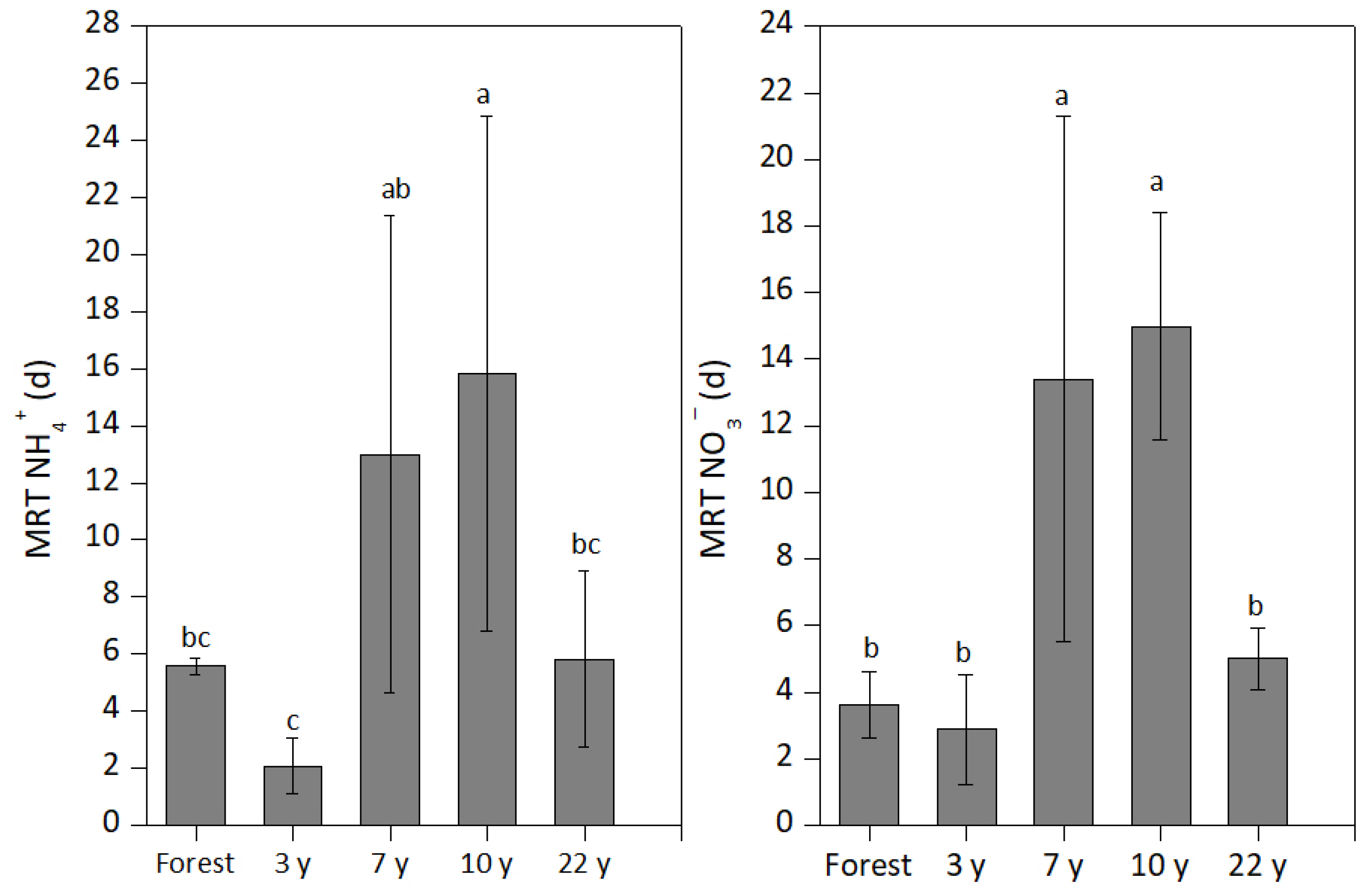
3.2. Gross Mineralization (MNorg) and Nitrification (ONorg) Rates
4. Discussion
4.1. Low Supply Capacity of Inorganic N in Soils under Natural Forest Conditions
4.2. Response of MNorg and ONorg Rates to Banana Cultivation
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, J.Z.; Bei, S.K.; Li, B.S.; Zhang, J.L.; Christie, P.; Li, X.L. Organic fertilizer, but not heavy liming, enhances banana biomass, increases soil organic carbon and modifies soil microbiota. Appl. Soil Ecol. 2019, 136, 67–79. [Google Scholar] [CrossRef]
- Meya, A.I.; Ndakidemi, P.A.; Mtei, K.M.; Swennen, R.; Merckx, R. Optimizing soil fertility management strategies to enhance banana production in volcanic soils of the northern highlands, Tanzania. Agronomy 2020, 10, 289. [Google Scholar] [CrossRef] [Green Version]
- FAO. FAO Statistical Databases. 2020. Available online: http://www.fao.org/faostat/zh/?#data/QC (accessed on 1 January 2021).
- Yao, L.X.; Li, G.L.; Yang, B.M.; Tu, S.H. Optimal fertilization of banana for high yield, quality, and nutrient use efficiency. Better Crops Plant Food 2009, 93, 10–11. [Google Scholar]
- Vitousek, P.M.; Howarth, R.W. Nitrogen limitation on land and in the sea: How can it occur? Biogeochemistry 1991, 13, 87–115. [Google Scholar] [CrossRef]
- Memon, N.U.N.; Memon, K.S.; Anwar, R.; Ahmad, S.; Nafees, M. Status and response to improved NPK fertilization practices in banana. Pak. J. Bot. 2010, 42, 2369–2381. [Google Scholar]
- Gonçalves, A.L.; Kernaghan, J.R. Banana Production Methods: A comparative Study; Centro Ecológico: Ipê da Serra, Brazil, 2014; 40p. [Google Scholar]
- Hobbie, E.A.; Högberg, P. Nitrogen isotopes link mycorrhizal fungi and plants to nitrogen dynamics. New Phytol. 2012, 196, 367–382. [Google Scholar] [CrossRef] [PubMed]
- Schimel, J.P.; Bennett, J. Nitrogen mineralization: Challenges of a changing paradigm. Ecology 2004, 85, 591–602. [Google Scholar] [CrossRef]
- Hart, S.C.; Nason, G.E.; Myrold, D.D.; Perry, D.A. Dynamics of gross nitrogen transformations in an old-growth forest: The carbon connection. Ecology 1994, 75, 880–891. [Google Scholar] [CrossRef]
- Zhang, J.B.; Cai, Z.C.; Zhu, T.B.; Yang, W.Y.; Müller, C. Mechanisms for the retention of inorganic N in acidic forest soils of southern China. Sci. Rep. 2013, 3, 2342. [Google Scholar] [CrossRef] [PubMed]
- Zhu, T.B.; Meng, T.C.; Zhang, J.B.; Yin, Y.; Cai, Z.C.; Yang, W.Y.; Zhong, W.H. Nitrogen mineralization, immobilization turnover, heterotrophic nitrification, and microbial groups in acid forest soils of subtropical China. Biol. Fert. Soils 2013, 49, 323–331. [Google Scholar] [CrossRef]
- Xie, Y.; Yang, L.; Zhu, T.B.; Yang, H.; Zhang, J.B.; Yang, J.L.; Cao, J.H.; Bai, B.; Jiang, Z.C.; Liang, Y.M.; et al. Rapid recovery of nitrogen retention capacity in a subtropical acidic soil following afforestation. Soil Biol. Biochem. 2018, 120, 171–180. [Google Scholar] [CrossRef]
- Wang, C.H.; Wang, N.N.; Zhu, J.X.; Liu, Y.; Xu, X.F.; Niu, S.L.; Yu, G.R.; Han, X.G.; He, N.P. Soil gross N ammonification and nitrification from tropical to temperate forests in eastern China. Funct. Ecol. 2018, 32, 83–94. [Google Scholar] [CrossRef] [Green Version]
- Shan, Z.J.; Yin, Z.; Yang, H.; Zuo, C.Q.; Zhu, T.B. Long-term cultivation of fruit plantations decreases mineralization and nitrification rates in calcareous soil in the karst region in southwestern China. Forests 2020, 11, 1282. [Google Scholar] [CrossRef]
- Knops, J.M.H.; Bradley, K.L.; Wedin, D.A. Mechanisms of plant species impacts on ecosystem nitrogen cycling. Ecol. Lett. 2002, 5, 454–466. [Google Scholar] [CrossRef] [Green Version]
- Baijukya, F.P.; de Ridder, N.; Masuki, K.F.; Giller, K.E. Dynamics of banana-based farming systems in Bukoba District, Tanzania: Changes in land use, cropping and cattle keeping. Agric. Ecosyst. Environ. 2005, 106, 395–406. [Google Scholar] [CrossRef]
- Dita, M.; Barquero, M.; Heck, D.; Mizubuti, E.S.G.; Staver, C.P. Fusarium wilt of banana: Current knowledge on epidemiology and research needs toward sustainable disease management. Front. Plant Sci. 2018, 9, 1468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradshaw, C.J.A. Little left to lose: Deforestation and forest degradation in Australia since European colonization. J. Plant. Ecol. 2012, 5, 109–120. [Google Scholar] [CrossRef]
- Zhu, T.B.; Zhang, J.B.; Huang, P.; Suo, L.; Wang, C.; Ding, W.X.; Meng, L.; Zhou, K.X.; Hu, Z.W. N2O emissions from banana plantations in tropical China as affected by the application rates of urea and a urease/nitrification inhibitor. Biol. Fert. Soils 2015, 51, 673–683. [Google Scholar] [CrossRef]
- Grünzweig, J.M.; Sparrow, S.D.; Chapin, F.S. Impact of forest conversion to agriculture on carbon and nitrogen mineralization in subarctic Alaska. Biogeochemistry 2003, 64, 271–296. [Google Scholar] [CrossRef]
- Zhu, T.B.; Zhang, J.B.; Meng, T.Z.; Zhang, Y.C.; Yang, J.J.; Müller, C.; Cai, Z.C. Tea plantation destroys soil retention of NO3− and increases N2O emissions in subtropical China. Soil Biol. Biochem. 2014, 73, 106–114. [Google Scholar] [CrossRef]
- Monkai, J.; Goldberg, S.D.; Hyde, K.D.; Harrison, R.D.; Mortimer, P.E.; Xu, J.C. Natural forests maintain a greater soil microbial diversity than that in rubber plantations in Southwest China. Agr. Ecosyst. Environ. 2018, 265, 190–197. [Google Scholar] [CrossRef]
- Ryals, R.; Kaiser, M.; Torn, M.S.; Berhe, A.A.; Silver, W.L. Impacts of organic matter amendments on carbon and nitrogen dynamics in grassland soils. Soil Biol. Biochem. 2014, 68, 52–61. [Google Scholar] [CrossRef]
- Zhang, J.B.; Cai, Z.C.; Yang, W.Y.; Zhu, T.B.; Yu, Y.J.; Yan, X.Y.; Jia, Z.J. Long-term field fertilization affects soil nitrogen transformations in a rice-wheat-rotation cropping system. J. Plant Nutr. Soil Sci. 2012, 175, 939–946. [Google Scholar] [CrossRef]
- Hwang, S.; Hanaki, K. Effects of oxygen concentration and moisture content of refuse on nitrification, denitrification and nitrous oxide production. Bioresour. Technol. 2000, 71, 159–165. [Google Scholar] [CrossRef]
- Bai, J.H.; Gao, H.F.; Deng, W.; Yang, Z.F.; Cui, B.S.; Xiao, R. Nitrification potential of marsh soils from two natural saline–alkaline wetlands. Biol. Fert. Soils 2010, 46, 525–529. [Google Scholar] [CrossRef]
- Huygens, D.; Boeckx, P.; Templer, P.; Paulino, L.; Van Cleemput, O.V.; Oyarzún, C.; Müller, C.; Godoy, R. Mechanisms for retention of bioavailable nitrogen in volcanic rainforest soils. Nat. Geosci. 2008, 1, 543–548. [Google Scholar] [CrossRef]
- Zhu, T.B.; Zhang, J.B.; Cai, Z.C. The contribution of nitrogen transformation processes to total N2O emissions from soils used for intensive vegetable cultivation. Plant Soil 2011, 343, 313–327. [Google Scholar] [CrossRef]
- Zhu, T.B.; Zeng, S.M.; Qin, H.L.; Zhou, K.X.; Yang, H.; Lan, F.N.; Huang, F.; Cao, J.H.; Müller, C. Low nitrate retention capacity in calcareous soil under woodland in the karst region of southwestern China. Soil Biol. Biochem. 2016, 97, 99–101. [Google Scholar] [CrossRef]
- Garousi, F.; Shan, Z.J.; Ni, K.; Yang, H.; Shan, J.; Cao, J.H.; Jiang, Z.C.; Yang, J.L.; Zhu, T.B.; Müller, C. Decreased inorganic N supply capacity and turnover in calcareous soil under degraded rubber plantation in the tropical karst region. Geoderma 2021, 381, 114754. [Google Scholar] [CrossRef]
- Allen, K.; Corre, M.D.; Tjoa, A.; Veldkamp, E. Soil nitrogen-cycling responses to conversion of lowland forests to oil palm and rubber plantations in Sumatra, Indonesia. PLoS ONE 2015, 10, e0133325. [Google Scholar] [CrossRef] [Green Version]
- Li, H.M.; Ma, Y.X.; Liu, W.J.; Liu, W.J. Soil changes induced by rubber and tea plantation establishment: Comparison with tropical rain forest soil in Xishuangbanna, SW China. Environ. Manag. 2012, 50, 837–848. [Google Scholar] [CrossRef]
- Nawaz, M.F.; Bourrié, G.; Trolard, F. Soil compaction impact and modelling. A review. Agrono. Sustain. Dev. 2013, 33, 291–309. [Google Scholar] [CrossRef] [Green Version]
- Kirkham, D.; Bartholomew, W.V. Equations for following nutrient transformations in soil utilizing tracer data. Soil Sci. Soc. Am. Proc. 1954, 18, 33–34. [Google Scholar] [CrossRef]
- Murphy, D.V.; Recous, S.; Stockdale, E.A.; Fillery, I.R.P.; Jensen, L.S.; Hatch, D.J.; Goulding, K.W.T. Gross nitrogen fluxes in soil: Theory, measurement and application of 15N pool dilution techniques. Adv. Agron. 2003, 79, 69–118. [Google Scholar]
- Ross, D.S.; Ketterings, Q. Recommended methods for determining soil cation exchange capacity. In Cooperative Bulletin No. 493. Recommended Soil Testing Procedures for the Northeastern United States; University of Delaware Newark: Newark, DE, USA, 2011; pp. 75–85. [Google Scholar]
- Bremner, J.M.; Keeney, D.R. Determination and isotope-ratio analysis of different forms of nitrogen in soils: 3. Exchangeable ammonium, nitrate, and nitrite by extraction-distillation methods. Soil Sci. Soc. Am. J. 1966, 30, 577–582. [Google Scholar] [CrossRef]
- Rotthauwe, J.H.; Witzel, K.P.; Liesack, W. The ammonia monooxygenase structural gene amoA as a functional marker: Molecular fine-scale analysis of natural ammonia-oxidizing populations. Appl. Environ. Microb. 1997, 63, 4704–4712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Francis, C.A.; Roberts, K.J.; Beman, J.M.; Santoro, A.E.; Oakley, B.B. Ubiquity and diversity of ammonia–oxidizing archaea in water columns and sediments of the ocean. Proc. Natl. Acad. Sci. USA 2005, 102, 14683–14688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, T.B.; Yang, C.; Wang, J.; Zeng, S.M.; Liu, M.Q.; Yang, J.L.; Bai, B.; Cao, J.H.; Chen, X.Y.; Müller, C. Bacterivore nematodes stimulate soil gross N transformation rates depending on their species. Biolo. Fert. Soils 2018, 54, 107–118. [Google Scholar] [CrossRef]
- Corre, M.D.; Brumme, R.; Veldkamp, E.; Beese, F.O. Changes in nitrogen cycling and retention processes in soils under spruce forests along a nitrogen enrichment gradient in Germany. Glob. Chang. Biol. 2007, 13, 1509–1527. [Google Scholar] [CrossRef]
- Rowley, M.C.; Grand, S.; Verrecchia, É.P. Calcium-mediated stabilization of soil organic carbon. Biogeochemistry 2018, 137, 27–49. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Y.; Wang, J.; Mary, B.; Zhang, J.B.; Cai, Z.C.; Chang, S.X. Soil pH has contrasting effects on gross and net nitrogen mineralizations in adjacent forest and grassland soils in central Alberta, Canada. Soil Biol. Biochem. 2013, 57, 848–857. [Google Scholar] [CrossRef]
- Kemmitt, S.; Wright, D.; Goulding, K.; Jones, D. pH regulation of carbon and nitrogen dynamics in two agricultural soils. Soil Biol. Biochem. 2006, 38, 898–911. [Google Scholar] [CrossRef]
- He, J.Z.; Hang, W.H.; Zhi, H.X. Chapter six-ammonia-oxidizing archaea play a predominant role in acid soil nitrification. Adv. Agron. 2014, 125, 261–302. [Google Scholar]
- Yao, H.Y.; Campbell, C.D.; Chapman, S.J.; Freitag, T.E.; Nicol, G.W.; Singh, B.K. Multi-factorial drivers of ammonia oxidizer communities: Evidence from a national soil survey. Environ. Microbiol. 2013, 15, 2545–2556. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.F.; Mulder, J.; Zhu, J.; Zhang, X.S.; Wang, Z.W.; Dörsch, P. Denitrification as a major nitrogen sink in forested monsoonal headwater catchments in the sub-tropics: Evidence from multi-site dual nitrate isotopes. Glob. Chang. Biol. 2019, 25, 1765–1778. [Google Scholar] [CrossRef]
- Canfield, D.E.; Glazer, A.N.; Falkowski, P.G. The evolution and future of Earth’s nitrogen cycle. Science 2010, 330, 192–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, K.C.; He, T.G.; Chen, H.; Peng, W.X.; Song, T.Q.; Wang, K.L.; Li, D.J. Impacts of vegetation restoration strategies on soil organic carbon and nitrogen dynamics in a karst area, southwest China. Ecol. Eng. 2017, 101, 247–254. [Google Scholar] [CrossRef]
- Wang, K.L.; Zhang, C.H.; Chen, H.S.; Yue, Y.M.; Zhang, W.; Zhang, M.Y.; Qi, X.K.; Fu, Z.Y. Karst landscapes of China: Patterns, ecosystem processes and services. Landsc. Ecol. 2019, 34, 2743–2763. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Z.H.; Liu, H.Y.; Wang, H.Y.; Peng, J.; Meersmans, J.; Green, S.M.; Quine, T.A.; Wu, X.C.; Song, Z.L. Bedrock geochemistry influences vegetation growth by regulating the regolith water holding capacity. Nat. Commun. 2020, 11, 2392. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Parameter i | Natural Forest | 3 y ii | 7 y | 10 y | 22 y |
|---|---|---|---|---|---|
| SOC (g C kg−1) | 34.6 ± 6.29 a | 32.9±3.14 a | 21.4±1.47 b | 20.0±2.36 b | 19.7±3.34 b |
| TN (g C kg−1) | 3.25 ± 0.18 a | 2.92 ± 0.18 a | 2.06 ± 0.06 b | 1.88 ± 0.09 b | 1.99 ± 0.29 b |
| pH | 6.75 ± 0.09 a | 6.29 ± 0.08 b | 5.05 ± 0.39 c | 4.65 ± 0.22 cd | 4.30 ± 0.16 d |
| WHC | 0.92 ± 0.15 a | 0.73 ± 0.02 b | 0.69 ± 0.02 bc | 0.67 ± 0.01 c | 0.58 ± 0.03 d |
| CEC (cmol kg−1) | 18.4 ± 1.25 a | 15.6 ± 0.85 b | 11.3 ± 0.07 c | 10.6 ± 0.38 c | 11.5 ± 0.49 c |
| CaO (%) | 8.39 ± 3.13 a | 3.94 ± 0.52 b | 1.77 ± 0.48 c | 1.45 ± 0.46 c | 1.54 ± 0.53 c |
| AP (mg kg−1) | 3.27 ± 0.51 c | 173 ± 26.0 a | 136 ± 10.3 b | 197 ± 18.2 a | 159 ± 19.6 a |
| AK (mg kg−1) | 236 ± 103 b | 783 ± 227 a | 848 ± 331 a | 800 ± 87.1 a | 761 ± 74.8 a |
| NH4+ (mg N kg−1) | 9.45 ± 0.36 a | 6.35 ± 2.54 a | 15.0 ± 8.41 a | 10.2 ± 5.34 a | 4.85 ± 0.7 a |
| NO3− (mg N kg−1) | 21.9 ± 1.41 c | 29.7 ± 6.79 c | 46.2 ± 3.55 b | 70.9 ± 12.7 a | 17.1 ± 5.36 c |
| NO3−/NH4+ | 2.31 ± 0.07 b | 5.08 ± 1.63 a | 3.65 ± 1.53 ab | 8.15 ± 4.07 a | 3.47 ± 0.78 ab |
| AOA abundance ×107 amoA gene copies (g dry soil)−1 | 11.6 ± 1.23 a | 14.4 ± 2.45 a | 5.55 ± 3.23 b | 3.66 ± 1.45 b | 1.45 ± 0.29 b |
| AOB abundance ×105 amoA gene copies (g dry soil)−1 | 19.6 ± 3.20 a | 24.6 ± 4.98 a | 6.48 ± 3.13 b | 11.6 ± 2.64 b | 9.14 ± 3.78 b |
| NH4+ | NO3− | MNorg | ONH4 | MRT NH4+ | MRT NO3− | |
|---|---|---|---|---|---|---|
| SOC | –0.09 | –0.22 | 0.87 ** | 0.70 * | –0.39 | –0.39 |
| TN | –0.22 | –0.34 | 0.91 ** | 0.72 ** | –0.58 | –0.51 |
| pH | –0.21 | –0.14 | 0.87 ** | 0.83 ** | –0.53 | –0.48 |
| WHC | 0.30 | 0.37 | 0.51 | 0.57 | 0.06 | 0.08 |
| CEC | –0.34 | –0.47 | 0.96 ** | 0.87 ** | –0.64 * | –0.65 * |
| CaO | –0.10 | –0.33 | 0.84 ** | 0.78 ** | –0.33 | –0.42 |
| AP | –0.09 | 0.47 | –0.14 | 0.04 | 0.18 | 0.10 |
| AK | –0.19 | 0.23 | –0.01 | 0.01 | –0.18 | –0.18 |
| AOA | 0.04 | –0.15 | 0.83 ** | 0.79 ** | –0.25 | –0.29 |
| AOB | –0.31 | –0.19 | 0.80 ** | 0.87 ** | –0.43 | –0.54 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qin, X.; Yang, C.; Yang, L.; Ma, E.; Meng, L.; Zhu, T. Response of Gross Mineralization and Nitrification Rates to Banana Cultivation Sites Converted from Natural Forest in Subtropical China. Land 2021, 10, 376. https://doi.org/10.3390/land10040376
Qin X, Yang C, Yang L, Ma E, Meng L, Zhu T. Response of Gross Mineralization and Nitrification Rates to Banana Cultivation Sites Converted from Natural Forest in Subtropical China. Land. 2021; 10(4):376. https://doi.org/10.3390/land10040376
Chicago/Turabian StyleQin, Xinghua, Cheng Yang, Lin Yang, Erdeng Ma, Lei Meng, and Tongbin Zhu. 2021. "Response of Gross Mineralization and Nitrification Rates to Banana Cultivation Sites Converted from Natural Forest in Subtropical China" Land 10, no. 4: 376. https://doi.org/10.3390/land10040376