
Abstract
Key message
Local climate exerted a stronger effect on leaf litter decomposition of deciduous Quercus acutissima Carruth. and evergreen Quercus glauca Thunb. than initial leaf litter quality: the decomposition was much faster for leaves of the two species in warm-temperate forest than in the cool-temperate one.
Context
Litter decomposition mainly depends on initial litter quality (e.g., physical features and nutrient content). However, the effects of litter quality on decomposition remain inconclusive in different climatic zones due to complex interaction with climatic factors.
Aims
This study investigated the effect of litter quality and climatic factors on litter decomposition of deciduous Quercus acutissima Carruth. and evergreen Quercus glauca Thunb. in warm-temperate forests in Jeju Island (Cheongsu; Seonheul) and cool-temperate forest at Chungnam National University Experimental Forest (CNU).
Methods
Through an inter-site experiment and litterbag technique, variations in remaining mass and decomposition coefficients (k) were measured from the two Quercus species in Korea.
Results
The initial mass loss was rapid during the wettest months in summer season. Annual k varied significantly by site: Seonheul (0.77 year−1 ± 0.03) > Cheongsu (0.62 year−1 ± 0.02) > CNU (0.42 year−1 ± 0.01). Leaf litter decomposition was similar for the two species in the warm-temperate sites and 36% higher in the deciduous species in the cool-temperate site. Based on the principal component analysis, overall k values were strongly correlated (rho = 0.55–0.60) with climatic factors, and no significant correlation was detected in leaf litter quality.
Conclusion
In conclusion, our results highlighted that the climatic condition in the study sites predominantly controlled the litter decomposition of the deciduous and evergreen oak species, with nevertheless an interspecific difference in the coolest site.
Similar content being viewed by others

1 Introduction
In terrestrial ecosystems, plant litter decomposition is one of the prominent processes linking aboveground and belowground carbon and nutrient cycling via litterfall (Zhou et al. 2008; Cornwell et al. 2008; Tomlinson et al. 2013). Through this process, nutrients are released to the soil and part of the carbon fixed during photosynthesis is returned to the atmosphere, and the rest is stored in the soil as organic matter (Moretto et al. 2001). Litter decomposition is influenced by initial litter traits and climate factors (Ge et al. 2017; Waring 2012). Under similar macroclimate and soil conditions, the rate of litter decomposition is expected to depend mainly on initial litter traits due to the presence of inhibitory compounds in litter and contrasting feeding preferences among different communities of decomposers (Cassart et al. 2020; Cordova et al. 2018). The carbon and nutrient chemistry and stoichiometry of the litter and its physical features can have a strong effect on the abundance and activity of decomposers leading to different rates of decomposition (Taylor et al. 1989). Increased leaf decomposition rates in the early stages of decay have generally been linked to high initial leaf N and P concentrations (McClaugherty et al. 1985), but Ribeiro et al. (2002) observed a contrasting pattern in several plant species. A high predictive power of specific leaf area (SLA) to litter decomposition has also been illustrated in an evergreen broadleaf (Huang et al. 2007), semi-evergreen (Sanchez-Silva et al. 2018), and tropical rain forests (Esquivel et al. 2020). However, several studies did not confirm those results as the effect of litter quality on decomposition may become inconsistent among different climatic zones due to complex interactions with climatic factors (Zhang and Wang 2015; Zhou et al. 2018). Both temperature and precipitation can directly affect litter decomposition by regulating the activity of decomposers and indirectly by their effects on leaf litter quality (Zhou et al. 2018; Alvarez-Clare and Mack 2011). In many studies, temperature was a strong predictor of leaf litter decomposition in evergreen and deciduous forests (Ge et al. 2017; Portillo-Estrada et al. 2016). For example, leaf litter decomposition rates were a positive linear function of mean annual temperature, such that the remaining time of leaf litter declined by ~31 days for every 1°C increase in temperature (Bothwell et al. 2014).
At global scale, an increasing number of inter-site decomposition experiments have been carried out to effectively investigate the influence of litter quality and macroclimate factors on litter decomposition (Bonan et al. 2013; Prescott 2010; Makkonen et al. 2012). Such a decomposition experiment involves exchanging of litters from different tree species and sites, allowing effective comparisons of the initial litter quality traits and climate variables (Harmon et al. 2009; Gholz et al. 2000). Most of the inter-site experiments have shown that litter decomposition was more responsive to environmental conditions (especially climate), explaining more than 50% of the variation in decomposition (Zhou et al. 2008; Cusack et al. 2009). However, other variables such as plant species-decomposer interactions via specific plant-soil interactions may also come into picture during the decomposition process, in which litter quality may play a significant role. Based on the “home-field advantage (HFA)” theory, litter decomposes more rapidly when incubated in the environment from which it is derived than in its outside environment, which can be explained by the specialization of decomposer communities to their particular substrate type (Gholz et al. 2000; Freschet et al. 2012). Moreover, litter decomposition rate generally decreases with the species diversity and abundance of decomposer communities (Liu et al. 2019).
In this study, we aimed at investigating the effect of litter quality and climatic conditions on litter decomposition of Quercus acutissima Carruth. and evergreen Quercus glauca Thunb. in a cool-temperate forest at CNU and a warm-temperate forest in Jeju Island in Korea through an inter-site experiment (reciprocal transplant experiment). The study sites differed in mean annual temperature (i.e., 15.5 °C, 14.8 °C, and 13.6 °C in Cheongsu, Seonheul, and CNU, respectively), annual precipitation (i.e., 1569 mm, 1883 mm, and 1299 mm in Cheongsu, Seonheul, and CNU, respectively), and dominant species, which are Q. acutissima (deciduous broadleaf) in the cool-temperate site and Q. glauca (evergreen broadleaf) at the warm-temperate sites. Evergreen leaves have a longer life span compared to deciduous ones, and this implies also differences in the physical structure and chemical composition of their leaves that can lead to contrasting temporal trend of decomposition (Cornwell et al. 2008). Leaf litter of deciduous species usually decomposes faster than evergreen leaves due to their more diverse leaf structures and chemical characteristics. However, results of previous studies are inconsistent across and within forest types due to the interacting controls of biotic and abiotic factors (e.g., Gessner et al. 2010; Petraglia et al. 2019). Consequently, two hypotheses were tested in this study: (1) the rate of litter decomposition is higher in sites with higher mean annual temperature (MAT) and mean annual precipitation (MAP), and (2) it is higher in deciduous species due to its leaf traits, which are more favorable to a faster rate of decay compared with the evergreen one.
2 Material and methods
2.1 Research site descriptions
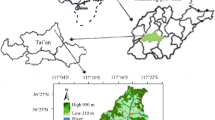
Two sites on Jeju Island [i.e., Cheongsu (CS) and Seonheul (SH) Gotjawal located at the southwest and northeast part of the island, respectively] and one site in Daejeon [i.e., Chungnam National University Experimental Forest (CNU)] were established in August 2015 (Fig. 1). These sites differ in species composition and dominance, stand structure, and climate (Table 1, Park et al. 2020). CS and SH sites are both warm-temperate broadleaf evergreen forests dominated by evergreen species (i.e., Q. glauca). In the former, Q. glauca displays a mean basal area (BA) of 30.3 m2 ha−1 and a mean height of 9.5 m, growing in association with the sub-canopy species (i.e., 11% of the total BA). At SH, the dominant species, Q. glauca, displays a mean basal area (BA) of 44.4 m2 ha−1 and a mean height of 13.0 m, growing in association with the canopy and sub-canopy tree species (i.e., 12% of the total BA). In contrast, CNU is a cool-temperate broadleaf deciduous forest dominated by deciduous Quercus acutissima with 18.5 m2 ha−1 mean BA and 13.8 m mean height in association with the other woody vegetation in the sub-canopy which accounted for 44% of the total BA.

The location of the research sites and plots in this study
The highest mean annual temperature (MAT) was observed at CS (15.5 °C) and intermediate at SH (14.8 °C), and the lowest was observed at CNU (13.6 °C). Winter is about 5 °C colder at CNU than Jeju Gotjawal sites, but all sites have similar summer temperature (i.e., 25–26 °C in June–August) (Fig. 2). Mean annual precipitation (MAP) also differed among the sites, i.e., SH (1883 mm) > CS (1569 mm) > CNU (1299 mm) (Fig. 2). In all stands, the amount of rainfall during 2015–2017 was the highest during mid-summer season (July) to mid-fall (September), while the lowest concentration occurred during winter (December–February).

A 3-year climate pattern at the three contrasting research sites in Jeju Island [Cheongsu (CS) and Seonheul (SH)] and in Daejeon [Chungnam National University (CNU) Experimental Forest]. Data were provided by the meteorological center near CS, SH, and CNU, i.e., Seogwang Station, Seonheul Station, and Daejeon Station, respectively as cited in Park et al. (2020)
Across the three sites, soils are generally acidic (pH values ranging from 4.0 to 4.5) but had contrasting soil textures, i.e., clay, loam, and sandy loam for CS, SH, and CNU, respectively (Table 2). The organic matter (OM), total nitrogen (TN), cation exchange capacity (CEC), and exchangeable cations (i.e., K+, Ca2+, Mg2+, and Na+) at Jeju Gotjawal are higher than CNU by at least 90%. Also, there is no much difference in available phosphorus between SH and CNU, but almost four-fold higher in CS.
2.2 Experimental design
An inter-site leaf litter decomposition experiment was carried out from August 2015 to November 2017 (2 years and 3 months). Freshly fallen leaves from two plant species were used: Q. acutissima, a deciduous species from CNU, and Q. glauca, an evergreen species from the two sites in Jeju Gotjawal. These tree species were chosen based on leaf functional traits and high dominance in the study sites. In this study, we used the leaf for the decomposition experiment because this plant organ can represent 50–80% of the total aboveground litter production in terms of biomass (Martinez-Yrizar 1995) and has relatively higher nutrient concentrations compared to the other plant organs (Alvarez et al. 1992).
Litterbags (256 cm2) were made from nylon (thermoplastic material) with 1 mm mesh size, and each litterbag received a total mass of 6 g of air-dried leaf litter samples of each species. They were then sealed and tagged for further collection. In August 2015, we established four study plots (100–144 m2) in each research site, and the distance between two plots was approximately 20 m. In each plot, we used four 3-m parallel lines with nine points each for litterbag placement. Without altering or removing any vegetation or litter layer on the forest floor, a total of 432 bags were placed on the top of the ground in CS, SH, and CNU for the succeeding nine collections at 3 months intervals (3 sites × 4 plots × 2 species × 2 bags species−1 × 9 collection times = 432 bags). As an inter-site experiment, litter from the two species was used in the three sites, following the site × species level design. The plot within each site was considered as the observational unit.
From each plot, we started collecting litterbags at the end of November 2015. The collected litterbags were carefully packed to avoid the loss of fine particles, stored in an icebox, and transported to the laboratory. The soil particles, fine roots, and other extraneous materials attached to the bags were carefully removed using a brush. Then, litterbags were dried at room temperature for several days and oven-dried down to a constant mass at 80 °C.
2.3 Measurement of leaf litter traits
In each site, a total of five fully mature leaves from four trees of the two species were used to determine SLA (specific leaf area, cm2 g−1); the leaf area was measured using LI-3000 (LI-COR Inc., Lincoln, NE, USA), and the weight was determined from the oven-dried samples at 65 °C. Composite subsamples of leaves from each oak species were air-dried and ground to determine leaf chemical traits following a standard protocol in tissue nutrient analysis. Total nitrogen, phosphorus (P), potassium (K), calcium (Ca), and magnesium (Mg) were determined before the inter-site decomposition experiment. Determination of N was done using the micro-Kjeldahl digestion method. Atomic absorption spectrophotometry (AA280FS, Varian, USA) and ascorbic acid method were used to determine the total Ca, Mg, and K and P, respectively (Park et al. 2020).
2.4 Data and statistical analysis
Mean mass loss rate across plots at each site and collection time was calculated (Park et al. 2021), and the percentage of dry mass remaining (RM) of each species at time t was computed following Eq. (1) (Bragazza and Iacumin 2009). The dry mass remaining of the two species collected from all sites (site × species) were fitted to a single negative exponential model, i.e., Eq. (2) (Olson 1963) using ordinary least square (OLS) regression to compute decomposition constant. Negative exponential decomposition models are commonly used in many studies to describe the decomposition of litter bags (Patricio et al. 2012; Rahman and Tsukamoto 2013).
Eq. (1) RM (%) = 100 – [X0 − Xi/X0] x 100
Eq. (2) y = X0e−kt
where X0 is the initial dry weight of the leaf litter, Xi is the weight of the residual leaf litter after time t (year), k is the decomposition constant, y is the litter mass remaining, and t is the time interval of sampling expressed in years.
All statistical analyses were performed using SAS 9.5 at p<0.05 significance level. One-way ANOVA was run to determine the significant difference in litter quality traits between species. Two-way ANOVA followed by Tukey’s post hoc test was employed to examine any statistical differences in k values across different sites and species and their interaction. To assess the relationships of decomposition rates among litter traits and climate variables, a principal component analysis (PCA) was done with seven litter traits (SLA, C, N, P, K, Ca, Mg, and C:N), two site factors (MAP and MAT), and k-values under different sites and species.
3 Results
3.1 Litter quality traits
Litter traits differed significantly between the two Quercus species, except N, K, and Ca concentrations (Table 3). SLA was significantly higher in Q. acutissima than Q. glauca by 39%. For initial leaf litter nutrients, Q. acutissima had significantly higher C and C/N ratio than Q. glauca, whereas P and Mg concentrations were significantly higher in Q. glauca (Table 3).
3.2 Decomposition rate by site and species
Leaf litter mass loss followed a similar pattern through time (Fig. 3). Initial mass loss from mid-August to mid-November in 2015 was large (20–22% at CS, 30–32% at SH, and 15–18% at CNU) for both species. The initial decomposition rate of Q. acutissima in 2015 was similar between CS and CNU, whereas that between CS and SH differed considerably. Leaf litter mass loss increased faster during summer (mid-May to mid-August 2016) than other seasons, but we could not find the same pattern in 2017. Similarly, rapid mass loss was also observed at the wettest months from July to August 2016.

Remaining mass (in percentage of the initial mass) of (a) Quercus acutissima and (b) Q. glauca leaf litter in Cheongsu (CS) and Seonheul (SH) Gotjawal in Jeju Island and Chungnam National University (CNU) Experimental Forest in Daejeon in Korea from August 2015 to November 2017. Vertical bars represent standard errors (n=4)
The annual decomposition constant differed significantly among the sites and species (Table 4 and Table 5 in Appendix). The k (average of the two species) at SH evergreen forest was the highest (0.77 year−1) across the sites, followed by another evergreen forest at CS (0.62 year−1), and the lowest (0.42 year−1) was observed at CNU deciduous forest. At the species level, there was no significant difference in k-values between Q. acutissima and Q. glauca placed at CS and SH, but varied significantly at CNU (p = 0.015) in which Q. acutissima had higher k than Q. glauca by 36% (Table 4 and Table 5 in Appendix). The half-life (t0.5) periods of decomposing leaf litter samples were found to be the shortest for Q. acutissima at SH and longest for Q. glauca at CNU (Table 4).
The potential influence of initial leaf litter traits of the two Quercus species and climatic factors on the decomposition rates across the three sites is illustrated through a PCA (Fig. 4). The first two axes (component 1 and component 2) accounted for 97.5% of the total variation; in particular, about 73.1% was explained by the first component. On component 1, high positive loadings were observed for initial leaf N, P, K, Ca, and Mg concentrations, whereas high negative loadings were observed for SLA, C/N ratio, and C concentration (Table 6 in Appendix). MAT (mean annual temperature), MAP (mean annual precipitation), and k displayed high positive loadings on component 2. Overall, initial litter traits varied by species as shown by the clear separation of Q. acutissima and Q. glauca in the PCA. Initial P, K, and Mg concentrations showed moderate negative correlation (rho = 0. 35) with initial C, C/N ratio, and SLA (Fig. 4, Table 7 in Appendix). The PCA also revealed that the overall k values were strongly correlated (rho = 0.55–0.60) with climatic factors (MAT and MAP).

Principle component analysis (PCA) based on the correlation matrix of initial leaf litter traits (SLA, specific leaf area; carbon and macro nutrient concentrations), decomposition constant (k, year−1), and climate conditions (MAT, mean annual temperature; MAP, mean annual precipitation) in Cheongsu (CS) and Seonheul (SH) Gotjawal in Jeju Island and Chungnam National University (CNU) Experimental Forest in Daejeon in Korea
4 Discussion
4.1 Variation in litter decomposition rate across sites
In this study, the k of both deciduous and evergreen Quercus species was found the highest at SH, intermediate at CS, and the lowest at CNU. The results can be ascribed to the effect of climate variables, which showed a strong correlation with k values based on the PCA result (Fig. 4). Specifically, although SH had lower temperature than CS, the precipitation is several-fold higher in the former site by 264 mm year−1 than in the latter site, suggesting that precipitation may have predominantly controlled the decomposition of the two species at SH and CS. This result is consistent with a large number of studies, which revealed that precipitation is among the most important determinant of litter decomposition rate. For example, several studies have concluded that litter decomposition rate would increase with the amount of precipitation in temperate forests (e.g., Cassart et al. 2020; Salah and Scholes 2011; Santonja et al. 2017). In a warm-temperate forest on Jeju Island in Southern Korea, the annual decay rate of evergreen broadleaf trees was also positively influenced by precipitation (MAP: 1533 mm) (Lee et al. 2015). A rainfall manipulation study revealed also that the mass losses of deciduous Q. acutissima and O. serrata and evergreen Pinus densiflora in fully covered treatment (i.e., less precipitation) were reduced by up to 26% compared with fully exposed litter (Salamanca et al. 2003).
In our experiment, we also found that the initial decomposition rate between CS and CNU for Q. acutissima during the first 3 months in 2015 was similar, whereas that between CS and SH differed considerably. Prior to the placement of litterbags on the ground, the precipitation at SH in July 2015 was significantly the highest compared with those of CS and CNU (Fig. 2). Further, although the monthly precipitation declined in all sites from August to December 2015, the highest monthly precipitation was maintained at SH from August to September, while those of CS and CNU significantly declined below 200 mm. Thus, it could be asserted that the precondition of the climate, particularly precipitation, prior to the conduct of the experiment may have also influenced the litter mass loss of Q. acutissima at CS and SH. The conditions of the forest floor at SH may have become more favorable for a faster decay rate of the species by having wetter soil when the litterbags were placed on the ground, exhibiting a soil moisture–driven phase of decomposition. Soil moisture availability, particularly at warmer regions, would affect the abundance and activity of soil microbes and soil fauna, thus exerting a significant influence on litter decomposition rate (Petraglia et al. 2019; Peterjohn et al. 1994). It was also reported that increasing the soil moisture favored mass loss, particularly in high-quality litter or those types rich in organic and inorganic compounds (Liu et al. 2005; Cotrufo et al. 2015).
4.2 Variation in litter decomposition across species
Here, we found no significant difference in leaf litter decomposition between Q. acutissima and Q. glauca for leaves placed in warm-temperate sites despite their contrasting leaf litter qualities, but it varied for leaves placed at the cool-temperate site (i.e., Q. acutissima decomposed faster than Q. glauca). We could further imply that the climatic factors predominantly controlled the mass loss of the two species regardless of litter quality (i.e., deciduous vs. evergreen) as revealed by the weak correlation of k values with the initial leaf litter traits. Here, both species showed a rapid and huge early mass loss regardless of the contrasting litter qualities and their origin, and the order of the amount of loss corresponded to the order of the amount of precipitation and temperature in each site, that is, SH > CS. Further, the pattern of the change in precipitation (i.e., high to low amount) from the first year to last year of the experiment, particularly in the summer season, tended to be similar in CS and SH sites. This sudden decrease in summer precipitation may have slowed down the decomposition process in both SH and CS, thus resulting in a similar decomposition rate. Contrarily, the significantly higher decay rate of Q. acutissima than Q. glauca at the cool-temperate site could be ascribed to a sudden large increase in summer precipitation in July 2016 and 2017, possibly through activation of the activity of decomposers and thermal balance change in the soil. Studies have already shown that seasonal increased or changed in climate have a significant effect on litter decomposition in both temperate and tropical forests (Anaya et al. 2012; Chadwick et al. 2016). For example, it was found that suddenly increased precipitation accelerated the decomposition of high-quality litter (Liu et al. 2006). We, therefore, suggest that the increased precipitation (i.e., from < 200 mm in July 2015 to as high as 380–440 mm in July 2016–2017) at CNU site may have increased the sensitivity of Q. acutissima to increased precipitation due to its higher initial litter quality, particularly SLA, C content, and C/N ratio. This is because, in contrast to low-quality litter, the high-quality ones contain large amounts of water-soluble materials which are prone to the leaching effect of precipitation and can therefore increase litter mass loss with precipitation (Lee et al. 2015; Bakker et al. 2011; Cornwell et al. 2008). Further, broadleaved evergreens generally have a longer life span and, thus, have thicker leaves, a greater number of cells, and higher tensile strength than deciduous ones (Cornelissen et al. 1999; Navas et al. 2003). These traits may have resulted in higher resistance to leaching effect of rainfall and attack of decomposers for the evergreen Q. glauca, thereby resulting in slower decomposition compared with the deciduous Q. acutissima. Although we showed that the litter traits had a weak correlation with k values, our result agrees with the findings reported in several studies, such that deciduous leaf litter decomposed faster than evergreen one (Rahman and Tsukamoto 2013; Salamanca et al. 2003; Sun and Zhao 2016; You et al. 2000).
Overall, our findings are consistent with a large number of studies which emphasized that climate consistently showed stronger effects on decomposition rates compared with litter quality traits across species and climatic zones (e.g., Alvarez-Clare and Mack 2011; Chen et al. 2013; Lu and Liu 2012; Zhang and Wang 2015; Zhang et al. 2008). In an oak forest plantation, for instance, it was reported that litter decomposition was much higher during the rainy season than the other seasons due to the physical effects of high rainfall such as leaching (Perez-Suarez et al. 2012). The usual explanation for this pattern of litter decomposition is the effect of warmer temperature and higher precipitation on microbial activity or abundance and stimulation of activities of soil macro-detritivores (Cruz-Martinez et al. 2012; Jasinska et al. 2019; van Geffen et al. 2011), while others attributed it to the synergistic actions of microbes and microarthropods (Riutta et al. 2012; Salamanca et al. 2003). The leaching effect of rainfall can also increase the palatability of litter by washing out tannins and lignin, which tended to increase the diversity of decomposers, thereby increasing litter mass loss (Dilly et al. 2001).
5 Conclusion
Our study revealed that, across sites, leaf litter decomposition of both deciduous and evergreen Quercus species was the fastest at SH with the highest annual and summer precipitation, intermediate at CS, and the lowest at CNU with the lowest annual and summer precipitation. Leaf litter decomposition between species tended to vary significantly at the site with an evident abrupt increase in summer precipitation throughout the experimental period. Therefore, leaf litter decomposition of both oak species was predominantly controlled by climatic variables, particularly precipitation, and not by the initial litter quality. Our findings should improve our understanding of how precipitation controls leaf litter decomposition of deciduous and evergreen species in warm- and cool-temperate forest stands. While still incomplete, our study is relevant to understanding the effects of increased precipitation changes caused by global warming to litter decomposition and nutrient cycling in oak-dominated forests in warm- and cool-temperate zones.
Data availability
The datasets generated during and/or analyzed during the current study are available in the Harvard Dataverse repository (https://doi.org/10.7910/DVN/2VDERV) and also from the corresponding author on reasonable request.
References
Alvarez S, Sánchez G, Sánchez-Gallén I, González-Iturbe J (1992) Methods for the study of primary productivity and decomposition in terrestrial communities. Universidad Nacional Autónoma de México, Mexico
Alvarez-Clare S, Mack MC (2011) Influence of precipitation on soil and foliar nutrients across nine Costa Rican forests. Biotropica 43:433–441. https://doi.org/10.1111/j.1744-7429.2010.00732.x
Anaya CA, Jaramillo VJ, Martinez-Yrizar A, Garcia-Oliva F (2012) Large rainfall pulses control litter decomposition in a tropical dry forest: evidence from an 8-year study. Ecosystems 15:652–663. https://doi.org/10.1007/s10021-012-9537-z
Bakker MA, Carreno-Rocabado G, Poorter L (2011) Leaf economics traits predict litter decomposition of tropical plants and differ among land use types. Funct Ecol 25:473–483
Bonan GB, Hartman MD, Parton WJ, Wieder WR (2013) Evaluating litter decomposition in earth system models with long-term litterbag experiments: an example using the Community Land Model version 4 (CLM4). Global Change Biol 19:957–974. https://doi.org/10.1111/gcb.12031
Bothwell LD, Selmants PC, Giardina CP, Litton CM (2014) Leaf litter decomposition rates increase with rising mean annual temperature in Hawaiian tropical montane wet forests. Peerj 2:e685. https://doi.org/10.7717/peerj.685
Bragazza L, Iacumin P (2009) Seasonal variation in carbon isotopic composition of bog plant litter during 3 years of field decomposition. Biol Fert Soils 46:73–77. https://doi.org/10.1007/s00374-009-0406-7
Cassart B, Basia AA, Jonard M, Ponette Q (2020) Average leaf litter quality drives the decomposition of single-species, mixed-species and transplanted leaf litters for two contrasting tropical forest types in the Congo Basin (DRC). Ann Forest Sci 77. https://doi.org/10.1007/s13595-020-00942-4
Chadwick R, Good P, Martin G, Rowell DP (2016) Large rainfall changes consistently projected over substantial areas of tropical land. Nat Clim Change 6:177–181. https://doi.org/10.1038/nclimate2805
Chen H, Dong SF, Liu L, Ma CA, Zhang T, Zhu XM, Mo JM (2013) Effects of Experimental Nitrogen and Phosphorus Addition on Litter Decomposition in an Old-Growth Tropical Forest. Plos One 8:e84101. https://doi.org/10.1371/journal.pone.0084101
Cordova SC, Olk DC, Dietzel RN, Mueller KE, Archontouilis SV, Castellano MJ (2018) Plant litter quality affects the accumulation rate, composition, and stability of mineral-associated soil organic matter. Soil Biol Biochem 125:115–124. https://doi.org/10.1016/j.soilbio.2018.07.010
Cornelissen JHC, Perez-Harguindeguy N, Diaz S, Grime JP, Marzano B, Cabido M, Vendramini F, Cerabolini B (1999) Leaf structure and defence control litter decomposition rate across species and life forms in regional floras on two continents. New Phytol 143:191–200. https://doi.org/10.1046/j.1469-8137.1999.00430.x
Cornwell WK, Cornelissen JHC, Amatangelo K, Dorrepaal E, Eviner VT, Godoy O, Hobbie SE, Hoorens B, Kurokawa H, Perez-Harguindeguy N, Quested HM, Santiago LS, Wardle DA, Wright IJ, Aerts R, Allison SD, van Bodegom P, Brovkin V, Chatain A, Callaghan TV, Diaz S, Garnier E, Gurvich DE, Kazakou E, Klein JA, Read J, Reich PB, Soudzilovskaia NA, Vaieretti MV, Westoby M (2008) Plant species traits are the predominant control on litter decomposition rates within biomes worldwide. Ecol Lett 11:1065–1071. https://doi.org/10.1111/j.1461-0248.2008.01219.x
Cotrufo MF, Soong JL, Horton AJ, Campbell EE, Haddix ML, Wall DH, Parton AJ (2015) Formation of soil organic matter via biochemical and physical pathways of litter mass loss. Nat Geosci 8:776. https://doi.org/10.1038/Ngeo2520
Cruz-Martinez K, Rosling A, Zhang Y, Song MZ, Andersen GL, Banfield JF (2012) Effect of rainfall-induced soil geochemistry dynamics on grassland soil microbial communities. Appl Environ Microb 78:7587–7595. https://doi.org/10.1128/Aem.00203-12
Cusack DF, Chou WW, Yang WH, Harmon ME, Silver WL, Team L (2009) Controls on long-term root and leaf litter decomposition in neotropical forests. Global Change Biol 15:1339–1355. https://doi.org/10.1111/j.1365-2486.2008.01781.x
Dilly O, Bartsch S, Rosenbrock P, Buscot F, Munch JC (2001) Shifts in physiological capabilities of the microbiota during the decomposition of leaf litter in a black alder (Alnus glutinosa (Gaertn.) L.) forest. Soil Biol Biochem 33:921–930. https://doi.org/10.1016/S0038-0717(00)00239-X
Esquivel J, Park BB, Casanoves F, Delgado D, Park GE, Finegan B (2020) Altitude and species identity drive leaf litter decomposition rates of ten species on a 2950 m altitudinal gradient in Neotropical rain forests. Biotropica 52:11–21. https://doi.org/10.1111/btp.12730
Freschet GT, Aerts R, Cornelissen JHC (2012) Multiple mechanisms for trait effects on litter decomposition: moving beyond home-field advantage with a new hypothesis. J Ecol 100:619–630. https://doi.org/10.1111/j.1365-2745.2011.01943.x
Ge JL, Xie ZQ, Xu WT, Zhao CM (2017) Controls over leaf litter decomposition in a mixed evergreen and deciduous broad-leaved forest, Central China. Plant Soil 412:345–355. https://doi.org/10.1007/s11104-016-3077-9
Gessner MO, Swan CM, Dang CK, Mckie BG, Bardgett RD, Wall DH, Hattenschwiler S (2010) Diversity meets decomposition. Trends Ecol Evol 25:372–380. https://doi.org/10.1016/j.tree.2010.01.010
Gholz HL, Wedin DA, Smitherman SM, Harmon ME, Parton WJ (2000) Long-term dynamics of pine and hardwood litter in contrasting environments: toward a global model of decomposition. Global Change Biol 6:751–765. https://doi.org/10.1046/j.1365-2486.2000.00349.x
Harmon ME, Silver WL, Fasth B, Chen H, Burke IC, Parton WJ, Hart SC, Currie WS, Lidet (2009) Long-term patterns of mass loss during the decomposition of leaf and fine root litter: an intersite comparison. Global Change Biol 15:1320–1338. https://doi.org/10.1111/j.1365-2486.2008.01837.x
Huang JJ, Wang XH, Yan ER (2007) Leaf nutrient concentration, nutrient resorption and litter decomposition in an evergreen broad-leaved forest in eastern China. Forest Ecol Manag 239:150–158. https://doi.org/10.1016/j.foreco.2006.11.019
Jasinska J, Sewerniak P, Markiewicz M (2019) Links between slope aspect and rate of litter decomposition on inland dunes. Catena 172:501–508. https://doi.org/10.1016/j.catena.2018.09.025
Kim JS, Kim DS, Lee KC, Lee JS, King GM, Kang S (2018) Microbial community structure and functional potential of lava-formed Gotjawal soils in Jeju, Korea. Plos One 14. https://doi.org/10.1371/journal.pone.0211435
Lee KE, Cha S, Lee SH, Shim JK (2015) Decomposition of leaf litter of some evergreen broadleaf trees in Korea. J Ecology Environ 38:517–528. https://doi.org/10.5141/ecoenv.2015.054
Liu P, Huang J, Han X, Sun OJ, Zhou Z (2005) Differential responses of litter decomposition to increased soil nutrients and water between two contrasting grassland plant species of Inner Mongolia. China Appl Soil Ecol 34:266–275
Liu P, Huang JH, Han XG, Sun OJ, Zhou Z (2006) Differential responses of litter decomposition to increased soil nutrients and water between two contrasting grassland plant species of Inner Mongolia, China. Appl Soil Ecol 34:266–275. https://doi.org/10.1016/j.apsoil.2005.12.009
Liu Y, Shen X, Chen YM, Wang LF, Chen QM, Zhang J, Xu ZF, Tan B, Zhang L, Xiao JJ, Zhu P, Chen LH (2019) Litter chemical quality strongly affects forest floor microbial groups and ecoenzymatic stoichiometry in the subalpine forest. Ann Forest Sci 76. https://doi.org/10.1007/s13595-019-0890-3
Lu SW, Liu CP (2012) Patterns of litterfall and nutrient return at different altitudes in evergreen hardwood forests of Central Taiwan. Ann Forest Sci 69:877–886. https://doi.org/10.1007/s13595-012-0213-4
Makkonen M, Berg MP, Handa IT, Hattenschwiler S, van Ruijven J, van Bodegom PM, Aerts R (2012) Highly consistent effects of plant litter identity and functional traits on decomposition across a latitudinal gradient. Ecol Lett 15:1033–1041. https://doi.org/10.1111/j.1461-0248.2012.01826.x
Martinez-Yrizar A (1995) Biomass distribution and primary productivity of tropical dry forests. In: Bullock SH, Mooney HA, Medina E (eds) Seasonally Dry Tropical Forests. Cambridge University Press, Cambridge, pp 326–345
McClaugherty CA, Pastor J, Aber JD, Melillo JM (1985) Forest litter decomposition in relation to soil nitrogen dynamics and litter quality. Ecology 66:266–275. https://doi.org/10.2307/1941327
Moretto AS, Distel RA, Didone NG (2001) Decomposition and nutrient dynamic of leaf litter and roots from palatable and unpalatable grasses in a semi-arid grassland. Appl Soil Ecol 18:31–37. https://doi.org/10.1016/S0929-1393(01)00151-2
Navas ML, Ducout B, Roumet C, Richarte J, Garnier J, Garnier E (2003) Leaf life span, dynamics and construction cost of species from Mediterranean old-fields differing in successional status. New Phytol 159:213–228. https://doi.org/10.1046/j.1469-8137.2003.00790.x
Olson JS (1963) Energy Storage and the Balance of Producers and Decomposers in Ecological Systems. Ecology 44:322–331. https://doi.org/10.2307/1932179
Park BB, Rahman A, Han SH, Youn WB, Hyun HJ, Hernandez J, An JY (2020) Carbon and nutrient inputs by litterfall in evergreen and deciduous forests in Korea. Forests 11. https://doi.org/10.3390/f11020143
Park BB, Hernandez J, An JY, Youn WB, Choi HS; Jung SC, Han SH (2021) "Primary data for "Leaf litter decomposition of deciduous Quercus acutissima Carruth. and evergreen Quercus glauca Thunb. in an inter-site experiment in three contrasting temperate forest stands in South Korea”", Harvard Dataverse, V1, [dataset], https://doi.org/10.7910/DVN/2VDERV
Patricio MS, Nunes LF, Pereira EL (2012) Litterfall and litter decomposition in chestnut high forest stands in northern Portugal. Forest Syst 21:259–271. https://doi.org/10.5424/fs/2012212-02711
Perez-Suarez M, Arredondo-Moreno JT, Huber-Sannwald E (2012) Early stage of single and mixed leaf-litter decomposition in semiarid forest pine-oak: the role of rainfall and microsite. Biogeochemistry 108:245–258. https://doi.org/10.1007/s10533-011-9594-y
Peterjohn WT, Melillo JM, Steudler PA, Newkirk KM, Bowles FP, Aber JD (1994) Responses of trace gas fluxes and N availability to experimentally elevated soil temperatures. Ecol Appl 4:617–625. https://doi.org/10.2307/1941962
Petraglia A, Cacciatori C, Chelli S, Fenu G, Calderisi G, Gargano D, Abeli T, Orsenigo S, Carbognani M (2019) Litter decomposition: effects of temperature driven by soil moisture and vegetation type. Plant Soil 435:187–200. https://doi.org/10.1007/s11104-018-3889-x
Portillo-Estrada M, Pihlatie M, Korhonen JFJ, Levula J, Frumau AKF, Ibrom A, Lembrechts JJ, Morillas L, Horvath L, Jones SK, Niinemets U (2016) Climatic controls on leaf litter decomposition across European forests and grasslands revealed by reciprocal litter transplantation experiments. Biogeosciences 13:1621–1633. https://doi.org/10.5194/bg-13-1621-2016
Prescott CE (2010) Litter decomposition: what controls it and how can we alter it to sequester more carbon in forest soils? Biogeochemistry 101:133–149. https://doi.org/10.1007/s10533-010-9439-0
Rahman MM, Tsukamoto J (2013) Leaf traits, litter decomposability and forest floor dynamics in an evergreen- and a deciduous-broadleaved forest in warm temperate Japan. Forestry 86:441–451. https://doi.org/10.1093/forestry/cpt015
Ribeiro C, Madeira M, Araujo MC (2002) Decomposition and nutrient release from leaf litter of Eucalyptus globulus grown under different water and nutrient regimes. Forest Ecol Manag 171:31–41. https://doi.org/10.1016/S0378-1127(02)00459-0
Riutta T, Slade EM, Bebber DP, Taylor ME, Malhi Y, Riordan P, Macdonald DW, Morecroft MD (2012) Experimental evidence for the interacting effects of forest edge, moisture and soil macrofauna on leaf litter decomposition. Soil Biol Biochem 49:124–131. https://doi.org/10.1016/j.soilbio.2012.02.028
Salah YM, Scholes MC (2011) Effect of temperature and litter quality on decomposition rate of Pinus patula needle litter. Procedia Environmen Sci 6:180–193. https://doi.org/10.1016/j.proenv.2011.05.019
Salamanca EF, Kaneko N, Katagiri S (2003) Rainfall manipulation effects on litter decomposition and the microbial biomass of the forest floor. Appl Soil Ecol 22:271–281. https://doi.org/10.1016/S0929-1393(02)00153-1
Sanchez-Silva S, De Jong BHJ, Aryal DR, Huerta-Lwanga E, Mendoza-Vega J (2018) Trends in leaf traits, litter dynamics and associated nutrient cycling along a secondary successional chronosequence of semi-evergreen tropical forest in South-Eastern Mexico. J Trop Ecol 34:364–377
Santonja M, Rancon A, Fromin N, Baldy V, Haattenschwiler S, Fernandez C, Montes N, Mirleau P (2017) Plant litter diversity increases microbial abundance, fungal diversity, and carbon and nitrogen cycling in a Mediterranean shrubland. Soil Biol Biochem 111:124–134. https://doi.org/10.1016/j.soilbio.2017.04.006
Sun Y, Zhao SQ (2016) Leaf litter decomposition in urban forests: test of the home-field advantage hypothesis. Ann Forest Sci 73:1063–1072. https://doi.org/10.1007/s13595-016-0577-y
Taylor BR, Parkinson D, Parsons WFJ (1989) Nitrogen and lignin content as predictors of litter decay-rates - a microcosm test. Ecology 70:97–104. https://doi.org/10.2307/1938416
Tomlinson KW, Poorter L, Sterck FJ, Borghetti F, Ward D, de Bie S, van Langevelde F (2013) Leaf adaptations of evergreen and deciduous trees of semi-arid and humid savannas on three continents. J Ecol 101:430–440. https://doi.org/10.1111/1365-2745.12056
van Geffen KG, Berg MP, Aerts R (2011) Potential macro-detritivore range expansion into the subarctic stimulates litter decomposition: a new positive feedback mechanism to climate change? Oecologia 167:1163–1175. https://doi.org/10.1007/s00442-011-2051-8
Waring BG (2012) A meta-analysis of climatic and chemical controls on leaf litter decay rates in tropical forests. Ecosystems 15:999–1009. https://doi.org/10.1007/s10021-012-9561-z
You YH, Namgung J, Lee YY, Kim JH, Lee JY, Mun HT (2000) Mass loss and nutrients dynamics during the leaf litter decomposition in Kwangnung experimental forest. J Korean For Soc 89:41–48
Zhang XY, Wang W (2015) Control of climate and litter quality on leaf litter decomposition in different climatic zones. J Plant Res 128:791–802. https://doi.org/10.1007/s10265-015-0743-6
Zhang DQ, Hui DF, Luo YQ, Zhou GY (2008) Rates of litter decomposition in terrestrial ecosystems: global patterns and controlling factors. J Plant Ecol 1:85–93. https://doi.org/10.1093/jpe/rtn002
Zhou GY, Guan LL, Wei XH, Tang XL, Liu SG, Liu JX, Zhang DQ, Yan JH (2008) Factors influencing leaf litter decomposition: an intersite decomposition experiment across China. Plant Soil 311:61–72. https://doi.org/10.1007/s11104-008-9658-5
Zhou SX, Huang CD, Xiang YB, Tie LH, Han BH, Scheu S (2018) Effects of reduced precipitation on litter decomposition in an evergreen broad-leaved forest in western China. Forest Ecol Manag 430:219–227. https://doi.org/10.1016/j.foreco.2018.08.022
Acknowledgements
We thank our colleagues (Woo Bin Youn, Jeong Min Seo, Afroja Rahman, Youngtak Ko, Huong Thi Thuy Dao) for their assistance in the field and laboratory work.
Contributions of the co-authors
Conceptualization: B.B.P. and S.C.J.Methodology: B.B.P.Validation: B.B.P., J.H., and J.Y.A.Formal analysis: B.B.P., J.H., and J.Y.A.Investigation: A.R., W.B.Y., and S.H.H.Resources: B.B.P. and S.H.H.Data curation: B.B.P. and S.H.H.Writing—original draft preparation: B.B.P., S.H.H., and A.R.Writing—review and editing: B.B.P., J.H., and J.Y.A.Visualization: J.Y.A.Supervision: B.B.P.Project administration: B.B.P., H.S.C., and S.C.J.Funding acquisition: B.B.P. and H.S.C.
Funding
This research was funded by the Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education (2018R1D1A3B07045079).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflicts of interest
The authors declare no competing interests.
Additional information
Handling Editor: Marcus Schaub
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Appendix
Appendix
Rights and permissions
About this article

Cite this article
Park, B.B., Han, S.H., Hernandez, J.O. et al. Leaf litter decomposition of deciduous Quercus acutissima Carruth. and evergreen Quercus glauca Thunb. in an inter-site experiment in three contrasting temperate forest stands in South Korea. Annals of Forest Science 78, 34 (2021). https://doi.org/10.1007/s13595-021-01058-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s13595-021-01058-z