
Abstract
Sequential effects are prominent and pervasive phenomena that exist in most perceptual judgments. Of importance, these effects reflect dynamic aspects in our judgment bias induced by the recent context. When making successive judgments in response to a sequence of stimuli, two opposing consequences have frequently been observed: assimilation effects – current stimuli judged as being closer to preceding stimuli than they actually are, and contrast effects – current stimuli judged as being further from preceding stimuli than they actually are. Although several cognitive accounts have been previously proposed, there is still a lack of consensus on the underlying mechanism, particularly regarding the insights of the temporal dynamics. Building upon accumulating human M/EEG findings, I propose a framework to explain how sequential bias is generated, unfolded over time, and eventually incorporated into the formation of current biased judgment. By bringing sequential effects closer to a biologically plausible framework, this synthetic view could account for how the opposing consequences of sequential effects differentially evolve, distinguish the effects from other perceptual phenomena with similar behavioral outcomes (such as aftereffects and priming), and illuminate how perceptual judgment is adaptively adjusted under the impact of temporal context.
Similar content being viewed by others

Introduction
In a world that continually evolves and changes, humans constantly have to adapt their judgments in response to changing opportunities and threats in nature. Such adaptations to a dynamic environment may result in privileged access to recent information, particularly given that, often, little information in the past is available that could be predictive of current judgments (Gold & Stocker, 2017). In other words, more recent information is more likely to be relevant and may provide a strong clue for optimizing current judgments. However, this strategy is inevitably accompanied by systematic biases, which can be manifested by sequential effects.
For some time, sequential effects have been one of the more prominent and pervasive phenomena that have been reported in the literature to exist in most perceptual judgments, especially when absolute information of current stimuli is not readily available (DeCarlo & Cross, 1990; Garner, 1953; Holland & Lockhead, 1968; Jesteadt, Luce, & Green, 1977; Lockhead, 2004; Ward & Lockhead, 1971). These effects are further viewed as having practical implications, such as for uncovering learning mechanisms (Jones, Curran, Mozer, & Wilder, 2013) and diagnosing neurodevelopmental disorders (Lieder et al., 2019). When encountering a sequence of stimuli along a given dimension (Fig. 1A, left panel) and when human observers have to respond to randomly presented stimuli one at a time (a typical setting in laboratory-based experimental tasks or in more naturalistic real-world situations, such as during online dating (Taubert, Van der Burg, & Alais, 2016) and sport performance evaluation (Damisch, Mussweiler, & Plessner, 2006)), the judgments regarding the current stimuli are biased by the local sequential context provided by the immediately preceding trials and, to some extent, by the preceding trials presented two or more trials earlier in the trial sequence. Such sequential effects hold even when a task is designed such that observers make choices based only on the sensory evidence at hand and are not explicitly aware of the sequential context.

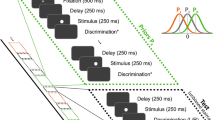
(A) Schematic illustration of the behavioral signatures of sequential effects. Sequential effects can be manifested in a simple binary choice task on a sequence of stimuli, such as a continuum of facial expressions morphed from fear to disgust (S1 to S5, left panel). The psychometric curve at the bottom of the left panel shows the classic responses to the stimuli at the current trial n. As shown in the right panel, when responses to stimuli (for example, S3) are analyzed according to the preceding context, current stimuli are more likely to be judged as “fear” or ”disgust” following preceding stimuli S2 or S4 (assimilation) or less likely to be judged as “fear” or ”disgust” following preceding stimuli S1 or S5 (contrast) than they actually are (i.e., a mean collapsed response (fear + disgust) of 50% across preceding stimulus types for S3). S, stimulus; R, response. (B) The neural signatures of sequential effects. Sequential context-dependent power modulations occur before the onset of the current stimulus (Time = 0) and in the low-frequency band. Assimilation is manifested by increased power (8–30 Hz, prominent at 20–28 Hz; left panel), whereas contrast is manifested by decreased power (10–30 Hz; right panel). The insets represent the topographies of the effects (the left inset serves for illustration only). Adapted from Hsu (2015) and de Lange et al. (2013), with permission
Two opposing consequences have frequently been observed either simultaneously or independently (Fig. 1A, right panel). On the one hand, assimilation effects have been obtained in which current stimuli are judged as being closer to preceding stimuli than they actually are. On the other hand, contrast effects have also been observed in which current stimuli are judged as being further from preceding stimuli than they actually are. However, despite recent renewed interest and a long history of research into these phenomena, the mechanism underlying sequential effects remains inconclusive. In this article, I propose an account of sequential effects as framed primarily by recent human electroencephalography (EEG) and magnetoencephalography (MEG) evidence and inspired by the previously existing and influential accounts that are briefly introduced below.
Existing accounts of sequential effects
Relative-judgment account
It has been thought that sequential effects reflect the result of a comparison strategy in which relative difference information between successive items is used to inform judgments (Stewart, Brown, & Chater, 2002). Because the memories for earlier stimuli interfere with the judgment of the distance between two successive stimuli, when these two stimuli are perceptually similar, participants may believe that the current and the preceding stimulus are the same (assimilation effect). In contrast, when facing two successive stimuli that are quite perceptually dissimilar, participants tend to judge these two stimuli as distinct (contrast effect).
Criterion-setting account
The criterion-setting account posits that sequential effects are mediated by two simultaneous but opposing mechanisms acting on the criterion for trial-by-trial perceptual judgment (Treisman & Williams, 1984). These two mechanisms are tracking and stabilization. On the one hand, the tracking mechanism assumes that the best evidence for the current judgment is the occurrence of recent judgments because, by nature, the recent evidence is likely to persist and thereby should be highly weighted. Accordingly, the criterion tends to be lowered so that when the same stimulus appears again, it is more likely to be correctly identified. This consequence produces assimilation to preceding responses in the current trial. On the other hand, the stabilization mechanism assumes that incoming sensory information can be used to appropriately position the criterion and keep it stable to maintain balanced responding in the long run. For example, if a series of sensory inputs all lie well above a criterion, the criterion may be too low in relation to the distribution of inputs. Thus, the criterion is elevated, resulting in contrast to previous stimuli.
Selective attention, mapping, ballistic accumulator (SAMBA) account
According to the SAMBA account, attention to the stimulus range, such as a continuum of facial expressions (Fig. 1A), is assumed to be maintained, thus yielding a limit on performance when the stimulus range is increased, for example. When a stimulus is presented, the relative position of the stimulus within the stimulus range is used to judge its magnitude. Specifically, given that attention is randomly distributed across the stimulus range, stimulus magnitude is estimated by ΣL/(ΣL +ΣU), where ΣL or ΣU, respectively, represents total activity between the current stimulus and the lower or upper end of the stimulus set (e.g., for the current stimulus S3: ΣL = total activity from S1 to S3, ΣU = total activity from S3 to S5).
Contrast effects arise through reallocation of activity in the selective attention stage. If attention is selectively directed to a preceding stimulus (S1 in Fig. 1A as an example) that falls within the lower range relative to the current stimulus (S3), this will increase ΣL (e.g., increased ΣL ≈ total activity from S1 to S4) on the current trial because additional activity is assigned around the location of the preceding stimulus. As a result, the current stimulus will be estimated as if it were the stimulus in the upper range (e.g., S4 because of similar stimulus magnitudes between S4 and adjusted S3 after activity reallocation). In other words, the current stimulus is judged as further from the preceding stimulus. In a similar vein, if attention is selectively directed to a preceding stimulus that falls within the upper range, this will increase ΣU and thereby the magnitude estimate of the current stimulus will shift in a direction closer to that of the stimuli in the lower range. In this case, the current stimulus is also judged as further from the preceding stimulus.
Assimilation effects arise through biased evidence accumulation on the current trial. Specifically, after the selective attention stage, the magnitude estimate of the current stimulus serves as input to a mapping process. This process transforms stimulus magnitude into a set of response strengths, which then drive ballistic accumulators, one for each possible response, with the first accumulator to reach a threshold determining the final choice. Here, the ballistic accumulators are evidence accumulators that accrue activation at a rate determined by the response strengths from the mapping process. Importantly, the starting point for each accumulator on the current trial is affected by residual activity from a previous trial. Assimilation could be accounted for by assuming the accumulator that won the race on the previous trial will begin the current trial with the highest level of the starting point. This benefits the same accumulator to reach the threshold, thus increasing the probability that the previous response will be produced again.
Continuity-field account
Recent renewed interest in sequential effects has postulated the “continuity field,” a specific mechanism responsible for assimilative bias (Fischer & Whitney, 2014). Specifically, assimilation reflects the nature of perceptual stability through which our visual system exploits the temporal continuity of past evidence because it tends to persist over time, making the recent past a good predictor of the present. This account stands in sharp contrast to widely documented negative aftereffects that represent neural adaptation that prepares the brain for the uptake of new information. In this view, assimilation and aftereffects reflect how the visual system balances the need to optimize sensitivity to new changes while capitalizing on the temporal continuity of past information. Therefore, this newly emerging line of research has frequently equated contrast effects to negative aftereffects.
Taken together, the aforementioned individual accounts emphasize distinct processes to account for sequential effects, as each process is built upon different cognitive constructs (Table 1). Moreover, the relative-judgment, criterion-setting, and continuity-field accounts regard assimilation and contrast as two sides of a single process. In contrast, the SAMBA account conceptualizes these two opposing biases as two somewhat separate processes. Although each account has been successful at explaining these two sequential consequences to different extents, there is still a lack of insight because most accounts, except for the SAMBA account, do not have a temporal component to track exactly how sequential bias is generated, reached over time, and eventually incorporated into the formation of current biased judgments. Furthermore, better insight into the temporal component could also help address a recent debate surrounding whether sequential effects operate at an early perceptual or a late decision stage (Cicchini, Mikellidou, & Burr, 2017; Fritsche, Mostert, & de Lange, 2017). More importantly, without bringing these accounts closer to a more biologically plausible framework, it will remain unknown how such temporal bias is implemented and constrained in the human brain.
A proposed neural-based account of sequential effects
Based on a synthesis of recent relevant behavioral and M/EEG (which offers excellent temporal resolution of brain activity) research, with a strong focus on the results from binary choice tasks in the absence of performance feedback, I propose a neural-based account of sequential effects to account for the temporal dynamics of sequential bias. The account can be broadly disentangled into three stages to describe (1) how neural dynamics manifest sequential effects; (2) how sequential context per se produces such context-dependent neural dynamics; and (3) how context-dependent neural dynamics interact with the current stimulus, leading to assimilative and contrastive response biases.
Sequential context-related neural dynamics as indexed by low-frequency prestimulus power
A few studies (de Lange, Rahnev, Donner, & Lau, 2013; Hsu, 2015) have directly examined the temporal neural dynamics pertaining to sequential context effects, as captured by time-frequency representations of brain activity. Although these studies were conducted to respectively investigate assimilation or contrast effects, a converging and coherent pattern surprisingly emerges despite different experimental settings in those electrophysiological studies (Fig. 1B). First, the overall results have shown that sequential context could elicit context-dependent power activity that precedes the onset of the current stimulus and slightly extends to the peristimulus periods. In addition, such prestimulus activity primarily occurs in the low-frequency band wherein assimilation is manifested by increased power (8–30 Hz, prominent at 20–28 Hz) and contrast is manifested by decreased power (10–30 Hz). Moreover, the context-dependent prestimulus power alone did not ultimately determine the current judgments, as the power is independent of the strength of current stimuli (de Lange et al., 2013; Hsu, 2015) and the current judgments rely on the coupling strength between the context-dependent prestimulus power and subsequent decision-related activity (Hsu, 2015).
Notably, the context-related prestimulus power is manifested by a broad frequency range, mainly spanning alpha and beta bands. This phenomenon is in line with recent evidence reporting a temporal and spatial co-modulation of alpha and beta rhythms (Michalareas et al., 2016) and studies indicating the concurrent contribution of alpha and beta prestimulus power in subsequent perceptual reports (Iemi, Chaumon, Crouzet, & Busch, 2017; Samaha, Switzky, & Postle, 2019). Moreover, these two rhythms may share similar functional roles and coordinate with each other, as both are thought to stabilize/maintain ongoing processing via top-down control (Clayton, Yeung, & Cohen Kadosh, 2018; Engel & Fries, 2010). In this view, reduction in these power activities reflect a change of ongoing states, facilitating shifts toward alternative states, whereas enhancement leads to intended states. In accordance with this notion, the context-dependent prestimulus alpha in tandem with beta power might participate in carrying cognitive information from previous trials to current trials, such that the magnitude of power activity may signal different states of this preserved information. Stronger power indicates the maintenance of such carry-over information, thereby leading to increased likelihood for eventual assimilation effects (i.e., response repetition), whereas reduced power indicates destabilization, leading to increased likelihood for eventual contrast effects (i.e., alternative choices). It is worth emphasizing that although the current proposal suggests that both alpha and beta powers subserve in maintaining ongoing cognitive information from previous trials, this does not contradict the presence of subfunctions in this processing. Alpha might maintain ongoing processing in response to upcoming perceptual evidence, whereas beta might maintain ongoing processing in response to upcoming sensorimotor activity (Clayton et al., 2018; Samaha, Gosseries, & Postle, 2017). In the next section, I further suggest that a change of internal criterion may serve as a potential construct that gives rise to the fluctuation of the state of carry-over information, as indexed by the context-dependent low-frequency prestimulus power.
The association between the context-dependent prestimulus power and the internal criterion
In past research, low-frequency prestimulus power has been linked to several cognitive variables, especially attention (Van Diepen, Foxe, & Mazaheri, 2019) or expectation (Summerfield & de Lange, 2014). However, in a standard experimental setting during sequential effects, stimuli are randomly presented, and levels of attention or expectation are not specifically manipulated. Thus, these two factors can be reasonably assumed to show little fluctuation during the prestimulus period. In addition, no evidence to date has indicated that assimilation and contrast effects reflect the results of different levels of these factors. In this proposal, I take a step further and suggest that the internal criterion might serve as one potential construct that produces the fluctuation of the context-dependent prestimulus power. In support of this hypothesis, recent studies on near-threshold stimulus detection have suggested that the low-frequency prestimulus power, diversely ranging from alpha to beta ranges, corresponded to the criterion used to commit perceptual judgment (Benwell et al., 2017; Craddock, Poliakoff, El-Deredy, Klepousniotou, & Lloyd, 2017; Iemi et al., 2017; Kloosterman et al., 2019; Limbach & Corballis, 2016; Wostmann, Waschke, & Obleser, 2019). This line of evidence has consistently demonstrated that the reduced or enhanced prestimulus power is related to a liberal (i.e., high false-alarm rate: reporting target presence even when no target is shown) or a conservative criterion, respectively (Fig. 2A). This view is further corroborated by a recent behavioral study that linked the fluctuations in alpha rhythms to the propagation of criteria through a perceptual history (Ho, Burr, Alais, & Morrone, 2019). From this viewpoint, a sequential context is likely to shift the internal criterion, as indexed by a change in the context-dependent prestimulus power, which in turn, is used to evaluate against current evidence.

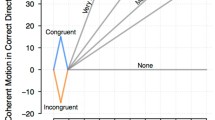
(A) Criterion-related neural signatures. A liberal criterion elicits decreased power with a prestimulus time window from -610 ms to stimulus onset at frequencies of 6–22 Hz. The inset represents the topography of the effect. The right panel depicts how the peak activity (10-Hz power) varies according to the criterion. The red line indicates the time points before which oscillatory activity is not contaminated by poststimulus activity. Adapted from Iemi et al. (2017), with permission. (B) A liberal criterion biases the drift rate at which evidence for “target-present” choice is accumulated even when no stimulus is present. z starting point
Nevertheless, the neural networks underlying the context-dependent prestimulus power remain obscure. On the one hand, the neural signatures reflecting assimilation have been found across sensory, motor to frontal regions (Akaishi, Umeda, Nagase, & Sakai, 2014; de Lange et al., 2013; St John-Saaltink, Kok, Lau, & de Lange, 2016). On the other hand, contrast-related prestimulus power has been identified over parietal-frontal topography (Hsu, 2015). This discrepancy may be attributed to several factors. First, for the MEG findings (de Lange et al., 2013; Hsu, 2015), only topography was reported without additional localization of the underlying sources, whereas for the neuroimaging findings (Akaishi et al., 2014; St John-Saaltink et al., 2016), it is unclear to which stage of sequential bias the observed brain regions correspond due to poor temporal resolution. Moreover, given that different stimuli and tasks were employed in those studies, in the future, simultaneously examining the two biases under the same experimental setting would better reveal the underlying neural networks.
Despite awaiting future clarification, the frontoparietal network might constitute one of the crucial substrates mediating the context-dependent prestimulus power. This proposition is formulated based on the following evidence. In animal studies, the frontoparietal network is engaged in representing the “decision trace” from previous trials (Hwang, Dahlen, Mukundan, & Komiyama, 2017; Tsunada, Cohen, & Gold, 2019). From the perspective of the temporal dynamics in sequential effects, the frontoparietal network has been found to associate with internal criterion (White, Mumford, & Poldrack, 2012) and to be involved in biasing evidence accumulation in current trials (Mulder, van Maanen, & Forstmann, 2014), one essential temporal component in shaping sequential effects, as is discussed in the next section. From the perspective of the frequency-band mechanism underlying the context-dependent power, beta power in the frontoparietal network is prominently linked to the active representation of maintained information (Antzoulatos & Miller, 2016; Spitzer & Haegens, 2017), while alpha recruits this network for top-down influences (Clayton et al., 2018) and may reflect the degree of biasedness of human choices (Grabot & Kayser, 2020). Overall, the frontoparietal network might be able to mediate the state of criterion in terms of low-frequency prestimulus power and the interaction between criterion and current evidence, as is formalized in the next section.
The effect of context-dependent prestimulus power on current perceptual judgment
The influential diffusion decision model provides one valuable hint at revealing the temporal mechanism by which the context-dependent prestimulus power can be incorporated into the formation of current judgments (Ratcliff, Smith, Brown, & McKoon, 2016). According to the model, perceptual judgment is characterized by the temporal accumulation of sensory evidence from a starting point to a boundary, and there is one boundary for each choice. A choice is made when the evidence accumulation, as determined by its drift rate, reaches one of the two decision boundaries. Sequential bias can be explained by (i) shifting the starting point (i.e., initial position of z in Fig. 2B) of accumulation toward one of the boundaries or (ii) modulating the drift rate (i.e., black arrows in Fig. 2B) at which evidence for a given choice is accumulated.
An accumulating number of findings now seem to support the drift-bias account (Fig. 2B). Diffusion modeling of participant choice behavior has indicated that assimilation can be captured by a bias in the rate of evidence accumulation toward the boundary reflecting the previous choice (Urai, de Gee, Tsetsos, & Donner, 2019). Complementing the above, by manipulating participants’ internal criteria, research (Kloosterman et al., 2019) has shown that a liberal criterion, accompanied by prestimulus alpha suppression, modulates the drift-bias parameter toward target-present choices. Notably, these results are compatible with prior findings showing that context-dependent prestimulus power may subsequently interact with poststimulus gamma or beta activity related to current evidence or current judgments (de Lange et al., 2013; Hsu, 2015). By referencing evidence from different domains as outlined above, I suggest a synthetic view that through the modulation of evidence accumulation by the internal criterion, which is indexed by the fluctuation of context-dependent prestimulus power, perceptual interpretation of current stimuli could be biased toward one of the two stimulus categories.
The generative source of context-dependent prestimulus power
Both conceptually and intuitively, preceding stimuli and preceding responses have long been considered essential elements in guiding the directionality of sequential effects (DeCarlo & Cross, 1990; Jesteadt et al., 1977). Because responses are inextricably intertwined with the characteristics of the stimuli, it is challenging to parse the relationship between the two. However, when the respective roles of preceding stimuli and preceding responses can be analytically separated, behavioral results do indicate that preceding responses are often associated with assimilation effects (Hsu & Wu, 2019; Jones, Love, & Maddox, 2006; Pegors, Mattar, Bryan, & Epstein, 2015), whereas preceding stimuli seem to involve contrast effects (Hsu & Wu, 2019; Pegors et al., 2015). Neural evidence further indicates that preceding stimuli/responses are able to reduce/elevate the low-frequency prestimulus power (de Lange et al., 2013; Hsu, 2015), although the effects of two preceding elements are somewhat conflated in these findings. Notably, contrast effects would be prevalent if current stimuli are preceded by stimuli from a different stimulus dimension (e.g., different facial identities between successive stimuli when performing a judgment on a continuum of facial expressions) (Hsu & Yang, 2013; Taubert, Alais, & Burr, 2016) or by stimuli at the ends of the stimulus range (i.e., large stimulus dissimilarity between preceding and current stimuli) (Hampton, Estes, & Simmons, 2005). Altogether, when the recent evidence provides little or ambiguous information, it is likely that observers may adopt a conservative criterion in an idiosyncratic manner, such that the best estimate for the current judgment is the occurrence of recent judgments. As a result, preceding responses may induce observers to repeat the responses in current trials, leading to assimilative sequential bias. However, when deviant preceding stimuli are present, for example, which signal the fluctuation of recent evidence, observers may need to adopt a more liberal criterion. As a result, such criterion readjustment in response to preceding stimuli facilitates shifts toward alternative choices, leading to contrastive sequential bias. In particular, all the influences from preceding stimuli and responses are likely to be carried by the low-frequency prestimulus power.
Summary
In the framework of the current proposed account (Fig. 3), I suggest that preceding stimuli and responses may compete and differentially modulate the low-frequency prestimulus power. As a result, the power would be either elevated or reduced, possibly representing fluctuations in the internal criterion. Subsequently, in the face of the current stimulus, this modulated power activity serves as one of the sources in forming the current perceptual judgment, as it could interact with current evidence by biasing the rate of evidence accumulation toward the boundary reflecting a given choice. Specifically, when the power activity is low due to strong influence from preceding stimuli, possibly reflecting a liberal criterion, for example, this will boost evidence accumulation drifting toward the opposite choice as indicated in the previous section (solid blue line at the bottom of the right inset in Fig. 3). In contrast, when the power activity is high due to a strong influence from preceding responses, possibly reflecting a conservative criterion, for example, the rate of evidence accumulation either facilitates the previous choice (solid red line at the top of the right inset) or impedes the opposite choice (solid red line at the bottom) – whichever is more likely requires further investigation. Notably, due to its idiosyncratic nature, assimilation relative to contrast effects appears to be robust, as assimilation could be induced by earlier preceding trials (Hsu & Wu, 2019). In addition, when current evidence conveys strong information (dashed orange line), robust evidence accumulation might override the drift bias induced by the preceding context, leading to reduced sequential effects (de Lange et al., 2013; Hsu & Yang, 2013).

The neural-based account of sequential effects. According to the account, preceding stimuli (dashed blue line) and responses (dashed red line) compete and differentially modulate the low-frequency prestimulus power. Subsequently, this modulated power activity interacts with the current evidence by biasing the rate of evidence accumulation toward the boundary reflecting a given choice. Increased power yields a tendency to repeat previous choices by facilitating the previous choice (solid red line at the top of the lower right inset) or impeding the opposite choice (solid red line at the bottom), thus producing assimilation; decreased power yields a tendency not to repeat previous choices by facilitating the opposite choice (solid blue line at the bottom of the lower right inset), thus producing contrast. When current evidence conveys strong information (dashed orange line), robust evidence accumulation might override the drift bias. In addition, the preceding feedback or metacognition may act as an additional element, which could modulate the low-frequency prestimulus power (dashed green line)
Concluding remarks
Contrast versus aftereffect and assimilation versus priming
Notably, well-established aftereffects also resemble contrastive bias such that current stimuli are perceived as being repulsed by preceding adaptors (Chopin & Mamassian, 2012; Webster, 2015). In addition, perceptual priming effects could lead to more accurate current responses after a prior exposure to a related or identical stimulus (Wiggs & Martin, 1998), thereby resembling assimilation. Despite sharing similar behavioral outcomes, these phenomena exhibit several distinct characteristics compared with sequential effects. First, aftereffects or priming are typically assessed in a test phase after an adaptation or priming phase during which responses are often not given to interrupt the effects of preceding stimuli. In principle, taking face perception as an example (see Mueller, Utz, Carbon, & Strobach, 2020, for a review), the strength of an aftereffect grows stronger as a function of adaptation time (usually more than seconds) but weaker as a function of test time, whereas robust priming requires brief presentation (i.e., subsecond presentation) of both the prime and the test stimuli. Conversely, contrast effects could be induced with subsecond stimulus durations (Hsu & Lee, 2016; Hsu & Yang, 2013), whereas assimilation has been observed using seconds of presentation time (Pegors et al., 2015). Next, both aftereffects and priming effects could survive with a long delay between adaptation/priming and the test phase, even up to days or more, whereas sequential effects are relatively labile and gradually disappear when there is a gap of a few trials between the preceding and current stimuli. Last, aftereffects or priming seem to reflect stimulus-specific neural representations, as they have been respectively characterized by poststimulus increases (Tikhonov, Handel, Haarmeier, Lutzenberger, & Thier, 2007; Wang, Iliescu, Ma, Josic, & Dragoi, 2011) or decreases (Friese, Supp, Hipp, Engel, & Gruber, 2012; Gruber & Muller, 2002) in gamma power, which represents a neural signature distinct from that of sequential effects as discussed here.
Generalization of the current account and future perspectives
Although beyond the present scope, the proposed account is potentially applicable, despite being subject to future examination, to several noted sequential-effect phenomena in other task settings. For example, Mori and Ward (1995) found that once feedback was provided, the effect of the previous stimulus became strong, and the effect of the previous response was reduced. In addition, Samaha et al. (2019) found that metacognition during subjective confidence rating amplifies the size of sequential effects. Accordingly, the present account would predict that the preceding feedback or metacognition acts as an additional element, in addition to the preceding stimuli/responses, which could modulate the low-frequency prestimulus power (dashed green line in Fig. 3). As another example, in an absolute identification task with feedback where a set of stimuli that vary on a single physical dimension are assigned to a set of labels, the pattern of assimilation and contrast is distinct from that during the binary choice task. Typically, the response made on the current trial is assimilated toward that on the immediately preceding trials, while the current response tends to contrast with that from trials further back in the sequence (Brown, Marley, Donkin, & Heathcote, 2008). In this case, the account would predict high prestimulus power induced by the immediately preceding trials, possibly reflecting a dominant idiosyncratic assimilative bias given the increasing uncertainty resulting from multiple labels, but anticipate low prestimulus power induced by trials further back, possibly reflecting an enhanced effect of preceding stimuli due to the presence of feedback (Mori & Ward, 1995).
Conclusion
In the current account, sequential effects are conceived as autonomous mechanisms to update low-frequency prestimulus power. Especially when encountering ambiguous stimuli, the updated power may strongly bias current judgment toward or away from the direction of the previous choice, depending on its magnitude calibrated by preceding stimuli and responses in an opposing manner. Notably, the current account goes beyond existing findings or accounts and provides additional insights into sequential effects. First, by linking the varying magnitude of context-dependent low-frequency prestimulus power to the opposing consequences of sequential biases, these two biases may be conceptualized as two sides of a single process and possibly mediated by the frontoparietal network. Specifically, the context-dependent prestimulus power seems to participate in maintaining ongoing cognitive information from previous trials. A change of internal criterion, as differentially driven by preceding stimuli or responses, may serve as a potential construct that gives rise to the fluctuation of the state of such cognitive trace. In this view, the current neural-based account bears some resemblance to the existing criterion-setting account; however, distinct from this psychological-based account, the current neural account proposes a framework to explain the temporal dynamics that ultimately shape sequential effects. Better understanding how sequential effects unfold over time is essential, as this advance may help address the debate regarding at which processing stage sequential effects occur. From a neural dynamic viewpoint, sequential effects reflect a phenomenon resulting from a series of chain events. Accordingly, either early perceptual or late decisional bias constitute only parts of a bigger temporal profile of sequential effects. To summarize, this framework integrates research across behavioral and neurophysiological fields to offer an account of the mechanisms motivating sequential effects, laying the groundwork for future experiments that can illustrate how perceptual judgment is dynamically and systematically biased under the impact of temporal context.
References
Akaishi, R., Umeda, K., Nagase, A., & Sakai, K. (2014). Autonomous mechanism of internal choice estimate underlies decision inertia. Neuron, 81(1), 195-206. https://doi.org/10.1016/j.neuron.2013.10.018
Antzoulatos, E. G., & Miller, E. K. (2016). Synchronous beta rhythms of frontoparietal networks support only behaviorally relevant representations. Elife, 5. https://doi.org/10.7554/eLife.17822
Benwell, C. S. Y., Tagliabue, C. F., Veniero, D., Cecere, R., Savazzi, S., & Thut, G. (2017). Prestimulus EEG power predicts conscious awareness but not objective visual performance. Eneuro, 4(6). https://doi.org/10.1523/ENEURO.0182-17.2017
Brown, S. D., Marley, A. A., Donkin, C., & Heathcote, A. (2008). An integrated model of choices and response times in absolute identification. Psychological Review, 115(2), 396-425. https://doi.org/10.1037/0033-295X.115.2.396
Chopin, A., & Mamassian, P. (2012). Predictive properties of visual adaptation. Current Biology, 22(7), 622-626. https://doi.org/10.1016/j.cub.2012.02.021
Cicchini, G. M., Mikellidou, K., & Burr, D. (2017). Serial dependencies act directly on perception. Journal of Vision, 17(14). https://doi.org/10.1167/17.14.6
Clayton, M. S., Yeung, N., & Cohen Kadosh, R. (2018). The many characters of visual alpha oscillations. European Journal of Neuroscience, 48(7), 2498-2508. https://doi.org/10.1111/ejn.13747
Craddock, M., Poliakoff, E., El-Deredy, W., Klepousniotou, E., & Lloyd, D. M. (2017). Pre-stimulus alpha oscillations over somatosensory cortex predict tactile misperceptions. Neuropsychologia, 96, 9-18. https://doi.org/10.1016/j.neuropsychologia.2016.12.030
Damisch, L., Mussweiler, T., & Plessner, H. (2006). Olympic medals as fruits of comparison? Assimilation and contrast in sequential performance judgements. Journal of Experimental Psychology: Applied, 12(3), 166-178. https://doi.org/10.1037/1076-898x.12.3.166
de Lange, F. P., Rahnev, D. A., Donner, T. H., & Lau, H. (2013). Prestimulus oscillatory activity over motor cortex reflects perceptual expectations. Journal of Neuroscience, 33(4), 1400-1410. https://doi.org/10.1523/JNEUROSCI.1094-12.2013
DeCarlo, L. T., & Cross, D. V. (1990). Sequential effects in magnitude scaling - models and theory. Journal of Experimental Psychology: General, 119(4), 375-396.
Engel, A. K., & Fries, P. (2010). Beta-band oscillations--signalling the status quo? Current Opinion in Neurobiology, 20(2), 156-165. https://doi.org/10.1016/j.conb.2010.02.015
Fischer, J., & Whitney, D. (2014). Serial dependence in visual perception. Nature Neuroscience, 17(5), 738-743. https://doi.org/10.1038/nn.3689
Friese, U., Supp, G. G., Hipp, J. F., Engel, A. K., & Gruber, T. (2012). Oscillatory MEG gamma band activity dissociates perceptual and conceptual aspects of visual object processing: a combined repetition/conceptual priming study. Neuroimage, 59(1), 861-871. https://doi.org/10.1016/j.neuroimage.2011.07.073
Fritsche, M., Mostert, P., & de Lange, F. P. (2017). Opposite effects of recent history on perception and decision. Current Biology, 27(4), 590-595. https://doi.org/10.1016/j.cub.2017.01.006
Garner, W. R. (1953). An informational analysis of absolute judgments of loudness. Journal of Experimental Psychology, 48, 218-224.
Gold, J. I., & Stocker, A. A. (2017). Visual decision-making in an uncertain and dynamic World. Annual Review of Visual Science, 3, 227-250. https://doi.org/10.1146/annurev-vision-111815-114511
Grabot, L., & Kayser, C. (2020). Alpha activity reflects the magnitude of an individual bias in human perception. Journal of Neuroscience, 40(17), 3443-3454. https://doi.org/10.1523/JNEUROSCI.2359-19.2020
Gruber, T., & Muller, M. M. (2002). Effects of picture repetition on induced gamma band responses, evoked potentials, and phase synchrony in the human EEG. Cognitive Brain Research, 13(3), 377-392. https://doi.org/10.1016/s0926-6410(01)00130-6
Hampton, J. A., Estes, Z., & Simmons, C. L. (2005). Comparison and contrast in perceptual categorization. Journal of Experimental Psychology: Learning, Memory, and Cognition, 31, 1459-1476.
Ho, H. T., Burr, D. C., Alais, D., & Morrone, M. C. (2019). Auditory perceptual history is propagated through alpha oscillations. Current Biology, 29(24), 4208-4217 e4203. https://doi.org/10.1016/j.cub.2019.10.041
Holland, M. K., & Lockhead, G. R. (1968). Sequential effects in absolute judgment of loudness. Perception & Psychophysics, 3, 409-414.
Hsu, S. M. (2015). The neural mechanism underlying the effects of preceding contexts on current categorization decisions. Neuropsychologia, 66, 39-47. https://doi.org/10.1016/j.neuropsychologia.2014.11.003
Hsu, S. M., & Lee, J. S. (2016). Relative judgment in facial identity perception as revealed by sequential effects. Attention Perception & Psychophysics, 78(1), 264-277. https://doi.org/10.3758/s13414-015-0979-1
Hsu, S. M., & Wu, Z. R. (2019). The roles of preceding stimuli and preceding responses on assimilative and contrastive sequential effects during facial expression perception. Cogn Emot, 1-16. https://doi.org/10.1080/02699931.2019.1696752
Hsu, S. M., & Yang, L. X. (2013). Sequential effects in facial expression categorization. Emotion, 13(3), 573-586. https://doi.org/10.1037/A0027285
Hwang, E. J., Dahlen, J. E., Mukundan, M., & Komiyama, T. (2017). History-based action selection bias in posterior parietal cortex. Nature Communications, 8(1), 1242. https://doi.org/10.1038/s41467-017-01356-z
Iemi, L., Chaumon, M., Crouzet, S. M., & Busch, N. A. (2017). Spontaneous Neural Oscillations Bias Perception by Modulating Baseline Excitability. Journal of Neuroscience, 37(4), 807-819. https://doi.org/10.1523/JNEUROSCI.1432-16.2016
Jesteadt, W., Luce, R. D., & Green, D. M. (1977). Sequential effects in judgments of loudness. Journal of Experimental Psychology: Human Perception and Performance, 3(1), 92-104. https://doi.org/10.1037/0096-1523.3.1.92
Jones, M., Curran, T., Mozer, M. C., & Wilder, M. H. (2013). Sequential effects in response time reveal learning mechanisms and event representations. Psychological Review, 120(3), 628-666. https://doi.org/10.1037/a0033180
Jones, M., Love, B. C., & Maddox, W. T. (2006). Recency effects as a window to generalization: separating decisional and perceptual sequential effects in category learning. Journal of Experimental Psychology: Learning, Memory, and Cognition, 32, 316-332.
Kloosterman, N. A., de Gee, J. W., Werkle-Bergner, M., Lindenberger, U., Garrett, D. D., & Fahrenfort, J. J. (2019). Humans strategically shift decision bias by flexibly adjusting sensory evidence accumulation. Elife, 8. https://doi.org/10.7554/eLife.37321
Lieder, I., Adam, V., Frenkel, O., Jaffe-Dax, S., Sahani, M., & Ahissar, M. (2019). Perceptual bias reveals slow-updating in autism and fast-forgetting in dyslexia. Nature Neuroscience, 22(2), 256-264. https://doi.org/10.1038/s41593-018-0308-9
Limbach, K., & Corballis, P. M. (2016). Prestimulus alpha power influences response criterion in a detection task. Psychophysiology, 53(8), 1154-1164. https://doi.org/10.1111/psyp.12666
Lockhead, G. R. (2004). Absolute judgments are relative: A reinterpretation of some psychophysical ideas. Review of General Psychology, 8(4), 265-272. https://doi.org/10.1037/1089-2680.8.4.265
Michalareas, G., Vezoli, J., van Pelt, S., Schoffelen, J. M., Kennedy, H., & Fries, P. (2016). Alpha-beta and gamma rhythms subserve feedback and feedforward Influences among human visual cortical areas. Neuron, 89(2), 384-397. https://doi.org/10.1016/j.neuron.2015.12.018
Mori, S., & Ward, L. M. (1995). Pure feedback effects in absolute identification. Perception & Psychophysics, 57(7), 1065-1079. https://doi.org/10.3758/Bf03205465
Mueller, R., Utz, S., Carbon, C. C., & Strobach, T. (2020). Face adaptation and face priming as tools for getting insights into the quality of face space. Frontiers in Psychology, 11, 166. https://doi.org/10.3389/fpsyg.2020.00166
Mulder, M. J., van Maanen, L., & Forstmann, B. U. (2014). Perceptual decision neurosciences - a model-based review. Neuroscience, 277, 872-884. https://doi.org/10.1016/j.neuroscience.2014.07.031
Pegors, T. K., Mattar, M. G., Bryan, P. B., & Epstein, R. A. (2015). Simultaneous perceptual and response biases on sequential face attractiveness judgments. Journal of Experimental Psychology: General, 144(3), 664-673. https://doi.org/10.1037/xge0000069
Ratcliff, R., Smith, P. L., Brown, S. D., & McKoon, G. (2016). Diffusion decision model: Current issues and history. Trends in Cognitive Sciences, 20(4), 260-281. https://doi.org/10.1016/j.tics.2016.01.007
Samaha, J., Gosseries, O., & Postle, B. R. (2017). Distinct oscillatory frequencies underlie excitability of human occipital and parietal cortex. Journal of Neuroscience, 37(11), 2824-2833. https://doi.org/10.1523/JNEUROSCI.3413-16.2017
Samaha, J., Switzky, M., & Postle, B. R. (2019). Confidence boosts serial dependence in orientation estimation. Journal of Vision, 19(4), 25. https://doi.org/10.1167/19.4.25
Spitzer, B., & Haegens, S. (2017). Beyond the status quo: a role for beta oscillations in endogenous content (re)activation. Eneuro, 4(4). https://doi.org/10.1523/ENEURO.0170-17.2017
St John-Saaltink, E., Kok, P., Lau, H. C., & de Lange, F. P. (2016). Serial Dependence in Perceptual Decisions Is Reflected in Activity Patterns in Primary Visual Cortex. Journal of Neuroscience, 36(23), 6186-6192. https://doi.org/10.1523/JNEUROSCI.4390-15.2016
Stewart, N., Brown, G. D. A., & Chater, N. (2002). Sequential effects in categorization of simple perceptual stimuli. Journal of Experimental Psychology: Learning, Memory, and Cognition, 28, 3-11.
Summerfield, C., & de Lange, F. P. (2014). Expectation in perceptual decision making: neural and computational mechanisms. Nature Reviews Neuroscience, 15(11), 745-756. https://doi.org/10.1038/nrn3838
Taubert, J., Alais, D., & Burr, D. (2016). Different coding strategies for the perception of stable and changeable facial attributes. Scientific Reports, 6. https://doi.org/10.1038/srep32239
Taubert, J., Van der Burg, E., & Alais, D. (2016). Love at second sight: Sequential dependence of facial attractiveness in an on-line dating paradigm. Scientific Reports, 6. https://doi.org/10.1038/srep22740
Tikhonov, A., Handel, B., Haarmeier, T., Lutzenberger, W., & Thier, P. (2007). Gamma oscillations underlying the visual motion aftereffect. Neuroimage, 38(4), 708-719. https://doi.org/10.1016/j.neuroimage.2007.07.055
Treisman, M., & Williams, T. C. (1984). A theory of criterion setting with an application to sequential dependencies. Psychological Review, 91(1), 68-111.
Tsunada, J., Cohen, Y., & Gold, J. I. (2019). Post-decision processing in primate prefrontal cortex influences subsequent choices on an auditory decision-making task. Elife, 8. https://doi.org/10.7554/eLife.46770
Urai, A. E., de Gee, J. W., Tsetsos, K., & Donner, T. H. (2019). Choice history biases subsequent evidence accumulation. Elife, 8. https://doi.org/10.7554/eLife.46331
Van Diepen, R. M., Foxe, J. J., & Mazaheri, A. (2019). The functional role of alpha-band activity in attentional processing: the current zeitgeist and future outlook. Current Opinion in Psychology, 29, 229-238. https://doi.org/10.1016/j.copsyc.2019.03.015
Wang, Y., Iliescu, B. F., Ma, J., Josic, K., & Dragoi, V. (2011). Adaptive changes in neuronal synchronization in macaque V4. Journal of Neuroscience, 31(37), 13204-13213. https://doi.org/10.1523/JNEUROSCI.6227-10.2011
Ward, L. M., & Lockhead, G. R. (1971). Response system processes in absolute judgment. Perception & Psychophysics, 9, 73-78.
Webster, M. A. (2015). Visual Adaptation. Annual Review of Visual Science, 1, 547-567. https://doi.org/10.1146/annurev-vision-082114-035509
White, C. N., Mumford, J. A., & Poldrack, R. A. (2012). Perceptual criteria in the human brain. Journal of Neuroscience, 32(47), 16716-16724. https://doi.org/10.1523/JNEUROSCI.1744-12.2012
Wiggs, C. L., & Martin, A. (1998). Properties and mechanisms of perceptual priming. Current Opinion in Neurobiology, 8(2), 227-233. https://doi.org/10.1016/s0959-4388(98)80144-x
Wostmann, M., Waschke, L., & Obleser, J. (2019). Prestimulus neural alpha power predicts confidence in discriminating identical auditory stimuli. European Journal of Neuroscience, 49(1), 94-105. https://doi.org/10.1111/ejn.14226
Acknowledgements
This work was supported by the Ministry of Science and Technology, R.O.C. (MOST 107-2420-H-002-031-RE2). The author thanks Li Yu for commenting on an early version of the manuscript. None of the data or materials for the experiments reported here are available, and none of the experiments were preregistered.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Hsu, SM. A neural-based account of sequential bias during perceptual judgment. Psychon Bull Rev 28, 1051–1059 (2021). https://doi.org/10.3758/s13423-021-01894-0
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13423-021-01894-0