
Abstract
A river otter hemimandible has been rediscovered during the revision of the historical collections of G.A. Blanc from Grotta Romanelli, complementing the ongoing multidisciplinary research fieldwork on the site. The specimen, recovered from the level G (“terre rosse”; early Late Pleistocene or late Middle Pleistocene), is here assigned to Lutra lutra. Indeed, morphological and morphometric comparisons with other Quaternary Lutrinae fossils from Europe allow to exclude an attribution to the relatively widespread and older Lutra simplicidens, characterized by distinctive carnassial proportions. Differences with Cyrnaonyx antiqua, which possessed a more robust, shellfish-feeding dentition, support the view of a successful niche repartition between the two species during the late Middle to Late Pleistocene of Europe. The occurrence of Lutra lutra from the “terre rosse” of Grotta Romanelli suggests deep modifications of the landscapes due to the ecological adaptation of the taxon, and indicates that the Eurasian otter spread into Europe at the Middle–Late Pleistocene transition.
Similar content being viewed by others

Introduction
Otters (Lutrinae), subfamily of Mustelidae, constitute a widely distributed carnivoran group (13 extant species), which evolved morphological adaptations for living and hunting in aquatic environments (Wilson and Mittermeier 2009; Nowak 1999). Despite earlier insights from morphological analysis (Van Zyll de Jong 1972,1987), river otters have been traditionally ascribed only to Lutra, until genetic data highlighted the substantial molecular divergence between the Eurasian and American branches of Lutrinae evolutionary tree, now assigned to Lutra and Lontra, respectively (Koepfli and Wayne 1998; Koepfli et al. 2008).
However, if Lontra occurred in North American deposits since the Pliocene (Prassack 2016), placing the first appearance of Lutra in a chronological framework is on the contrary complicated by the use of the genus either as a waste-basket taxon, or in sensu lato, to allocate specimens of uncertain affinities and/or Lutra-like morphology (Willemsen 1992; Geraads et al. 2015; Cherin et al. 2016). In fact, the earliest “Lutra” species, Lutra affinis Gervais 1859, is only known from three sites close to the Miocene–Pliocene boundary: Venta del Moro (Spain), Maramena (Greece) and Montpellier (France). However, the whereabouts of the never-figured type material from Montpellier are unknown (Willemsen 1992). Furthermore, the fragmentary remains from the other localities markedly differ in size (Koufos 2011; Montoya et al. 2011) and have been suggested to represent different taxa (Cherin et al. 2016). Several putative Lutra species have been described from the Pliocene to the Early Pleistocene of Europe (Gervais 1859; Pohle 1919; Kurtén 1968; Willemsen 1992; Fejfar and Sabol 2004), but none of them provides conclusive evidence of their generic status (Cherin et al. 2016).
A long gap characterized the Lutrinae record during the late Pliocene and the beginning of the Early Pleistocene, until the earliest occurrence of Lutra simplicidens and Lutraeximia umbra from the late Villafranchian Upper Valdarno (Tuscany) and Pantalla (Umbria), respectively (Cherin and Rook 2014; Cherin et al. 2016). The latter taxon is only known from the holotype cranium, whereas the former is relatively widespread in the early Middle Pleistocene and is regarded as ancestral to several endemic otters of the Mediterranean islands (Willemsen 2006): “Lutra” euxena from Malta, Lutraeximia trinacriae from Sicily, and Sardolutra ichnusae, “Lutra” castiglionis, Algarolutra majori and Megalenhydris barbaricina from Sardinia and Corsica, even though these fossil samples often include few and fragmentary remains (Willemsen 1992; Cherin et al. 2016). Nonetheless, the extant Eurasian river otter, Lutra lutra, is considered not related to these European species and its origin is placed in the Asian continent, probably deriving from the fossil species Lutra palaeoindica (Willemsen 1992; Raghavan et al. 2007). This scenario has received indirect support from the first description of L. simplicidens maxillary dentition, which led Cherin (2017) to doubt of the species generic attribution, remarking the similarities with Lutraeximia umbra (Cherin et al. 2016). Alternatively, Fejfar and Sabol (2004) argued for a derivation of the extant Lutra sumatrana from Lutra bravardi Pomel 1843, but excluding a close relationship with L. lutra of the latter species, which is difficult to reconciliate with molecular data that consistently find a close relation between L. lutra and L. sumatrana (Koepfli et al. 2008; Hung and Law 2016).
L. lutra is the most widely distributed otter species, with extant populations occurring throughout Asia, Europe and northern Africa. Historically, this carnivoran was also widespread in Japan, Portugal, arctic region of Asia and Europe and southern Indonesia (Foster-Turley et al. 1990). Up to now, fossils of the species are mainly known from Holocene deposits (Sommer and Benecke 2004), even if it was also reported from the late Middle Pleistocene of Grotta Maggiore di San Bernardino (Cassoli and Tagliacozzo 1994c) and the Late Pleistocene sites of Geographical Society Cave (Russian Far East) (Baryshnikov 2015), Pilisszánto 1 (Jánossy 1986) and Skjonghelleren Cave (Larsen et al. 1987). Indeed, Baryshnikov and Puzachenko (2012) suggested that the river otter could have survived during the cold phases of the Last Glacial in southern European refugia, from where it redispersed during the Early Holocene. Another otter species, Cyrnaonyx antiqua, is reported from several late Middle to Late Pleistocene European localities and is characterized by a more robust, shellfish-feeding type of dentition (Kurtén 1968; Willemsen 1992).
The otter fossil from Grotta Romanelli (southern Italy), belonging to the historical collection of Gian Alberto Blanc and never figured or described, is here presented for the first time. In addition, morphological and morphometric comparisons with other Lutrinae fossils coming from the Early Pleistocene to Middle Holocene sites of Europe and with extant specimens of European otter are taken into account to evaluate their morphological and body size variability.
Materials and methods
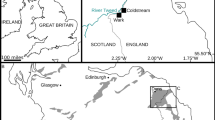
Grotta Romanelli (40° 00′ 58.33″ N, 18° 25′ 59.36″ E) is a costal cave located in the administrative territory of the Castro (Lecce) municipality, at the south-eastern extremity of Apulia (southern Italy) (Fig. 1a–d). The cave lies within the area of the Regional Natural Park “Costa Otranto—Santa Maria di Leuca and Bosco di Tricase”. From the beginning of the twentieth century, Grotta Romanelli was considered a key site for Mediterranean Quaternary studies, due to its remarkable importance as the first evidence of the Upper Paleolithic period in the Italian Peninsula and the first record of rock art in the Euro-Mediterranean area (Graziosi 1973; Sardella et al. 2018, 2019). The deposit was discovered in 1874 by Ulderigo Botti (Botti 1874), but only during the earlier 1900s, Paolo Emilio Stasi realized the first excavation campaigns (Stasi and Regalìa 1904). In 1914, Gian Alberto Blanc began a pioneering excavation campaign, during which the first systematic archeological/paleontological and stratigraphic study using scientific methods was carried out (Blanc 1920, 1928) (Fig. 1e–g). Nowadays, Grotta Romanelli is recognized as a key site for the Mediterranean Pleistocene for its archeological and paleontological content and the relative stratigraphical, geomorphological and radiometric data (Sardella et al. 2018, 2019).

(modified by Blanc 1920). Fossil Lutra lutra from Grotta Romanelli in labial (h), lingual (i) and occlusal (j) views
Geographical position of Grotta Romanelli (a–c). External (d) and internal (e) views of the cave. Stratigraphy of the deposit as described by Blanc (f), the level where the fossil otter was found is highlighted in light orange. Schematic log of the sedimentary succession
The infilling deposit was described by Blanc (1920) which identified two main stratigraphical complexes: the upper complex, called “terre brune” (levels A–E), bearing Upper Paleolithic artifacts and vertebrate fauna; the lower complex (G–K) formed by a deposit called “terre rosse” (level G), bearing Middle Paleolithic artifacts and vertebrate fauna, a stalagmitic layer (level H), a bone breccia (level I), and a beach deposit (level K) referred to the Tyrrhenian Stage (Marine Isotopic Stage 5) (Blanc 1920, 1928) (Fig. 1f, g). These two complexes were separated by a thick stalagmitic layer (called level F) (Blanc 1920, 1928) (Fig. 1g). The upper complex was referred to the end of the Late Pleistocene and Early Holocene on the basis of radiocarbon datings performed since the end of 1950 (Bella et al. 1958; Vogel and Waterbolk 1963; Alessio et al. 1964, 1965). The two speleothems (levels H and F) were also dated with the 230Th/234Th method: level H yielded an apparent age of < 69,000 years BP, and level F was dated to 40,000 ± 3250 years BP (Fornaca-Rinaldi 1968a, b; Fornaca-Rinaldi and Radmilli 1968; Sardella et al. 2018). During the 1950s and 1960s, the excavation campaigns were directed by Cardini with the support of the Italian Institute of Human Paleontology (IsIPU) (Sardella et al. 2018).
In 2015, new multidisciplinary research fieldwork began, coordinated by Sapienza University of Rome together with other research institutions and with the authorization of Soprintendenza Archeologia Belle Arti e Paesaggio di Brindisi, Lecce e Taranto, with the aim to provide an updated stratigraphic and chronological scheme as well as to describe the fossil remains and artifacts (Giustini et al. 2018; Sardella et al. 2018, 2019), which is also triggering a revision of the large mammal faunas from the region (Sardella et al. 2014; Mecozzi et al. 2017, 2018, 2019; Iannucci et al. 2020; Iurino et al. 2020). The research team had to face the consequences of more than 40 years of inactivity in the site and the combined effects of erosion and illegal excavations that took place during last decades (Conti et al. 2017; Giustini et al. 2018; Sardella et al. 2018). Recently, the age of the “terre brune” has been reassessed, with new fossil remains submitted for 14C dating using the accelerator mass spectrometry radiocarbon (AMS14C) dating techniques (Calcagnile et al. 2019; Sigari et al. 2021). The results expand and refine the previous chronology, with a time span for levels D and B, respectively, from 13,976–13,545 14C cal BP to 9135–8639 14C cal BP (Calcagnile et al. 2019), whereas the level E was dated between 13,087–12,839 and 13,408–13,153 (Sigari et al. 2021). As reported by Sardella et al. (2018), one of the main unexplained aspects, which needs to be investigated, concerns the age of the lower deposits due to the apparent contradiction between the supposed archaic nature of the Mousterian artifacts and the position of the vertebrate fauna being situated above the supposed MIS 5.5 terrace.
New analyses on the fauna and lithic tools from the lower complex suggest a possible late Middle Pleistocene to early Late Pleistocene age (Sardella et al. 2018), but more data need to define the updated chronostratigraphy of the infilling deposit.
The fossil remains described in this work were collected from the “terre rosse” (level G), but no information about the exact stratigraphical position is available (Blanc 1920, 1928) (Fig. 1h–j). Other mammal species found in the level G are: Elephas antiquus (= Palaeoloxodon antiquus), Rhinoceros merckii (= Stephanorhinus kirchbergensis), Hippopotamus sp. amphibius, Hippopotamus sp. pentlandi, Bos taurus var. primigenius (= Bos primigenius), Cervus elaphus, Cervus elaphus var. corsicanus (= Cervus elaphus corsicanus), Dama dama, Capreolus capreolus, Equus caballus, Hyaena crocuta var. spelaea (= Crocuta crocuta spelaea), Canis lupus, Canis aureus, Meles taxus (= Meles meles), Oryctolagus cuniculus and Pelagius monachus (= Pelagius monachus) (Blanc 1920). The wolf and rhino remains from level G have recently been revised by Sardella et al. (2014) and Pandolfi et al. (2017), who referred them to Canis lupus and Stephanorhinus hemitoechus, respectively.
The fossil material from Grotta Romanelli is housed in the collection of the “Museo delle Civiltà” (ex. Museo Preistorico ed etnografico Luigi Pigorini), labeled with a progressive number MUCIV-GR1935 (Fig. 1h–j).
According to von den Driesch (1976) and Willemsen (1992), 16 mandibular and dental variables have been considered: total length (Tl), length from the anterior border of the C1 to posterior border of the M2 (LC1M2), height of the mandibular corpus above M1 (HM1), length and breadth of the lower teeth (C1L, C1W, P2L, P2W, P3L, P3W, P4L, P4W, M1L, M2L, M2W) length of trigonid (M1Ltri), breadth of the trigonid (M1Wtri) and talonid (M1Wtal). Craniodental morphometric measurements were recorded to the nearest 0.1 mm with a digital caliper.
Literature data on fossil river otters from the Early Pleistocene to Early Holocene (Late Villafranchian to Aurelian) sites (Table 1) as well as extant specimens of L. lutra have been used. Comparisons are also based on fossil material found in the late Neolithic site of Tosina di Monzambano (Mantova—Italy) (Bona 2014; Poggiani Keller 2014) and extant specimens housed in the PaleoFactory laboratory, Department of Earth Sciences, Sapienza, University of Rome.
We performed body mass estimates of the fossil sample following regression equations based on the length of the lower carnassial (Van Valkenburg 1990) (Table 1). Size and proportions of the lower carnassial of the specimen from Grotta Romanelli were compared with literature data on Pleistocene–Holocene otters through standard bivariate plot. In mustelids, these teeth enable determining masticatory performance and bite force and their morphology and size are linked to diet (e.g., Baryshnikov et al. 2003; Madurell-Malapeira et al. 2011a, b).
Systematic Paleontology
Class Mammalia Linnaeus, 1758
Order Carnivora Bowdich, 1821
Family Mustelidae Fischer von Waldheim, 1817
Subfamily Lutrinae Bonaparte, 1838
Genus Lutra Brisson, 1762
Lutra lutra Linnaeus, 1758
Figure 1h-j
Material. Right hemimandible (MUCIV-GR1935; measurements in Table 2).
Locality and level. Grotta Romanelli; “terre rosse”—G.
Description. MUCIV-GR1935 is a finely preserved mandible, lacking only of a portion of the vertical ramus. In labial view, it shows three mental foramina: one located to the mid length of the C1; one posteriorly to the distal border of the P2; and one to the mid length of the P3. The anterior portion of the masseteric fossa reaches the level of the distal portion of the M1. The ramus is high and straight with respect to the corpus axis and the angular process is elongated dorsally. In dorsal view, the mandibular condyle has a straight posterior border and is inclined in posterolateral direction. The ventral outline of the mandibular corpus is almost straight, but anteriorly, at the level of the alveolus of the P2, it sharply rears upwards. The toothrow is relatively straight and the longitudinal axis of each tooth diverges, with the C1, P3 and M1 axes almost aligned. The posterior part of the P4 crown slightly overlaps with the anterior margin of the M1. No incisors are preserved. The C1 is mesodistally elongated, with a smooth crown. No diastema separates the C1 from the P2. The P2 is a two-rooted and buccolingually compressed tooth with a small and mesially projected protoconid, and with a crest connecting the weak distal cingulum to the protoconid. The P3 is buccolingually compressed and has the high, straight and anteriorly located protoconid, and a weak crest connecting the mesial cingulum to the protoconid. It has weak lingual and labial cingula. The P4 is also buccolingually compressed, with the high protoconid, a marked distal accessory cusp, and a marked basal cingulum.
In the lower carnassial (M1) the talonid is wider than the trigonid. The paraconid is well developed and high as the metaconid. The protoconid is the highest cusp and two distinct grooves separate it from the paraconid and the entoconid. The entoconid and hypoconid are reduced and aligned with the metaconid and the paraconid, respectively. Along the distal rim of the talonid, there are two very reduced cusps, buccally the entoconulid and lingually the hypoconulid. The talonid basin is concave. The basal distal cingulum is well developed, whereas anteriorly tends to be less marked.
The M2 displays a circular outline, with four cusps. The less developed cuspids, the paraconid and the hypoconid, occupy the anterior and posterior portion of the toot,h respectively; whereas, the metaconid and the protoconid are more developed.
Comparison. MUCIV-GR1935 possesses a typical Lutra-like appearance. Morphologically, the lower carnassial of the specimen from Grotta Romanelli shares several features with L. lutra (Fig. 2f–l): talonid wider than trigonid; concave talonid; hypoconulid present; strong and continuous lingual cingulum (Willemsen 1992). Opposite characters were observed in L. simplicidens from Hundsheim, Sussenborn, Mosbach, East Runton and West Runton (Fig. 2a–d): trigonid wider than talonid; flat talonid; hypoconulid absent; less developed and incomplete lingual cingulum (Willemsen 1992). However, the specimen NHM M29688 from the Upper Valdarno ascribed to L. simplicidens possesses a continuous cingulum, which extends all around the paraconid, and the trigonid and talonid are equal in width (Cherin and Rook 2014) (Fig. 2a). Similarly, in the specimen AMZ/OP-966 from the Chumbur Kosa (Azov Sea area), classified as L. simplicidens tamanensis, trigonid and talonid are equal in width (Sotnikova and Titov 2009) (Fig. 2d). The studied specimen differs from L. palaeindica from the Upper Siwalik in that it possesses a larger trigonid than talonid (Fig. 2e). MUCIV-GR1935 shares several features with the extant L. sumatrana (Fig. 2m, n), such as the talonid larger than trigonid and the concave talonid basin.

Comparative morphological sketch of the lower carnassial of the fossil and extant otter taxa in occlusal view (drawn not in scale). Lutra simplicidens: a NHM M29688 from Upper Valdarno (Cherin and Rook 2014); b BM 6089 from West Runton (Willemsen 1992); c SMF PA/F 8718 from Mosbach (Willemsen 1992); d KP-27075/OP-966 from Chumbur Rosa (Sotnikova and Titov 2009). Lutra palaeindica: e BM37152 from Upper Siwalk (Matthew 1929). Fossil Lutra lutra: f MUCIV-GR1935 from Grotta Romanelli; g ZIN 37,285–1 from Geographical Society Cave (Baryshnikov 2015); h, i Tos 6813 and Tos 7128 from Tosina di Monzambano (Mantova—Italy). Extant Lutra lutra: j, k 882 and 883; l n/a (Pohle, 1919). Lutra sumatrana: m n/a (Pohle 1919); n ZM CN4494 (Zyll de Yong, 1972). Cyrnaonyx antiqua: o–q BM M 34,378 and BM M 50,709 from Tornetown Cave (Willemsen 1992); p Ke RM 226 from Maasvlakte (Willemsen 1992)
According to Willemens (1992), the lower carnassial of Cyrnaonyx antiqua differs from L. lutra in the following features: larger paraconid; metaconid more closely connected to the other two cuspids of the trigonid and smaller than the other two cuspids; talonid wider than trigonid; labial cingulum more developed; larger hypoconid (Fig. 2o, p). MUCIV-GR1935 from Grotta Romanelli resembles C. antiqua for the large paraconid and the talonid wider than trigonid.
In the extant specimens of L. lutra, the morphology of the M2 is highly variable, with three main morphotypes recognized, characterized by a progressive complication of crown morphology: A1, with only protoconid and metaconid; A2, with protoconid, metaconid, paraconid and hypoconid; A3, with all the four cuspids on the grinding surface and an additional element, a projection on the lingual side of tooth crown (Gimranov and Kosintsev 2012). Following this, MUCIV-GR1935 can be referred to the A2 morphotype. The only M2 of L. simplicidens from the Upper Valdarno differs from MUCIV-GR1935 in the absence of the paraconid and in possessing a more centrally placed protoconid (Cherin and Rook 2014).
Results
Morphometric comparison
Standard bivariate plots of talonid width (M1WTal) vs length (M1L) of the lower carnassial show a distinctively small size for the genus Lutra (Fig. 3a). Within the Lutra group, the specimen from Grotta Romanelli and those of L. simplicidens fall in the broader variability of L. lutra. The C. antiqua material is instead characterized by a large size of the lower carnassial, well distinct in the M1WTal (Fig. 3a). Considering the M1WIndex (calculated as M1Wtal−M1Wtri), other differences can be observed (Fig. 3b). Two groups can be distinct based on the M1WIndex: the material with positive values (above the dotted line), which has a larger talonid; and those specimens with negative values (below the dotted line), which have a larger trigonid. Generally, C. antiqua and L. lutra occupy the upper part of the graphs, whereas L. simplicidens occupies the lower part (Fig. 3b). However, a few specimens of Lutra show a talonid and trigonid equal in width (located on the dotted line) (Fig. 3b).

Plot of lower carnassial: a length of M1 vs. width of M1 talonid; b length of M1 vs. width of M1 W index (calculated as M1 W talonid−M1 W trigonid)
Body mass
Body size in extant otters is hugely variable, reflecting the diverse ecological niches this group occupies (Conroy et al. 2000; Kruuk 2006): small-sized piscivore (c. 5–10 kg), medium-sized cancrivore (c. 11–12 kg) and large-sized molluscivore (c. 22–25 kg) (Raghavan et al. 2007). Therefore, in the fossil record, reconstructing the body size could represent an important proxy to evaluate possible ecological niche differentiation (Conroy et al. 2000). The Eurasian otter is a dimorphic species, with males 50% larger than females (Larivière and Jennings 2009). According to Conroy et al. (2000), the body mass of males is of 5.45–11.4 kg, whereas that of females is of 3.36–7.6 kg. Conversely, values reported by Kruuk (2006) range from 5.0 to 14.0 kg, but no sex distinction was made. Considering the fossil sample, the M1L predicts a reduced body-size variation for the Early–Middle Pleistocene L. simplicidens, with values between 5.2 and 7.0 kg (Table 1). The L. lutra sample possesses slightly larger, but not different body mass, with estimated values ranging from 5.0 to 8.2 kg (Table 1). Finally, the highest body size values are obtained for the sample of C. antiqua, whose estimates are between 7.8 and 10.8 kg (Table 1).
The estimated values on fossil otters reveal a reduced body size variation among groups, with slight difference in the average values and wide overlap in distribution ranges. Taking into account the body mass range of the extant Eurasian otter, the difference among the fossil groups is even less significant, since all their body size values fall into the extant variation range.
Discussion
Eurasian quaternary otters
Blanc (1920, 1928) reported for the first-time the presence of L. lutra from the level G of Grotta Romanelli. Since then, the Eurasian otter from this archeological site was cited by paleontologists (e.g., Willemens 1992; Pennacchioni and Cassola 1986), but no operational diagnosis was available, thus preventing its morphological and/or biometric comparison. The lack of these data sometimes sparked debates on taxonomy of some specimens belonging to these collections, as for instance happened for the canid collection recovered from the level G of Grotta Romanelli (see Sardella et al. 2014 for discussion). Morphological analysis reveals that the features of the fossil specimen from Grotta Romanelli clearly fit with an attribution to the extant L. lutra, confirming what Blanc (1920, 1928) reported.
Despite several fossil otters have been referred to Lutra over the years, many of them are now reassigned to other genera, whereas “Lutra” is retained mainly for ease of reference for taxa of uncertain taxonomy (Willemsen 1992; Cherin et al. 2016), recognizing its paraphyletic nature (Geraads et al. 2015). L. simplicidens has a relatively abundant fossil record, compared to other Pleistocene Lutrinae. However, it is almost entirely known from isolated mandibular and postcranial remains (Willemsen 1992), which are not directly comparable with the recently described Lutraeximia umbra, only known for the holotype cranium. It is worth noticing that Cherin (2017), providing the first description of the maxillary dentition of L. simplicidens, recognized a close relationship between this species, Lutraeximia umbra, Lutraeximia trinacriae and other Mediterranean Quaternary otters likely forming a monophyletic clade (Cherin et al. 2016). If new discoveries are needed to substantiate the attribution of L. simplicidens to Lutraeximia (Cherin 2017), nonetheless there is general agreement in recognizing as hardly conceivable a derivation of L. lutra from L. simplicidens (Willemsen 1992, 2006). Our data agree in evidencing morphometric differences between the two species, providing further support for a different derivation of L. lutra.
Willemsen (1992) hypothesized an Asian origin for L. lutra, probably from L. palaeindica. However, the latter taxon is only known from the type sample (Falconer 1868; Pohle 1919; Mattthew 1929), and possibly from another specimen (Raghavan et al. 2007), recovered from the Upper Siwalik. Similarly, no cranial material of L. simplicidens has been described so far, which highlights the strongly restricted knowledge on the morphological variability of these species. Nevertheless, in dental morphology, L. palaeindica shares many features with L. simplicidens. In the P4: the metacone lower than the paracone with a crest that connects the two cusps curved lingually, the parastyle well defined and the talonid mesiodistally elongated, triangular-shaped in occlusal view (though more pronounced in L. palaeindica) (Fig. 4); and in the M1: the trigonid larger the talonid and a relatively reduced metaconid (Fig. 2). Compared to C. antiqua, these two taxa instead possess marked differences in the upper and lower carnassial morphology, especially in the P4, where the profile in occlusal view is trapezoidal (Fig. 4j–m).

Upper fourth premolar of fossil and extant otter in occlusal view (drawn not in scale). Lutra simplicidens: a IQW 1966/7313 (Voi. 3525) from Voigtstedt; b IQW 1966/7314 (Voi. 3532) from Voigtstedt (Cherin 2017). Lutra palaeindica: c BM37151 from Upper Siwalik (Matthew 1929). Lutra sp.: d BM4936 from Hoxne (Willemsen 1992). Lutra lutra: e n/a (Zyll de Jong 1972). f n/a (Pohle 1919). Lutra sumatrana: g CN4494 (Zyll de Jong 1972); h n/a (Pohle 1919). Lutraeximia umbra: i SBAU 337,654 from Pantalla (Cherin et al. 2016). Cyrnaonyx antiqua: j 7 M from Cengelle (Pasa 1947); k–m BM34371, BM50707 and BM34373 from Tornetown Cave (Willemsen 1992)
Setting our results in chronological framework, it seems that L. lutra dispersed into Europe from Asia at the beginning of the Late Pleistocene or, possibly, during the late Middle Pleistocene, replacing L. simplicidens. In this scenario, the taxonomic attribution of two Middle Pleistocene specimens needs to be considered. The first is the hemimandible from Cengelle BB classified as L. lutra by Pasa (1947). According to his description, the specimen displayed clear Lutra features, among which is a trigonid larger than talonid in the lower carnassial. It is worth noting that L. simplicidens was instituted by Thenius (1965), and thus the name was not available to Pasa (1947). According to Willemsen (1992), the trigonid larger than talonid character has a high taxonomic value, which allows an unquestionable attribution of the specimen from Cengelle BB to L. simplicidens. The second specimen was a P4 recovered from Hoxne and ascribed to Lutra sp. by Willemsen (1992). The author recognized the strong resemblance with L. lutra, but given the absence of coeval maxillary Lutrinae material, he could not rule out an attribution to L. simplicidens, preferring an evolutionary hypothesis in which L. lutra appears during the Early Holocene. Recently, Cherin (2017) described for the first time the morphology of the maxillary dentition of L. simplicidens. The comparison of the P4 shows slightly but significative morphological differences between the Hoxne specimen and Voigtstedt ones (Fig. 4a, b, d; Table 3). Notably, L. simplicidens possesses a characteristic talon profile, with a convexity along the distal margin, about half length, in occlusal view (Cherin 2017) (Fig. 4a, b). In the P4 from Hoxne, the profile of the distal margin is straight, not convex, which allows to refer it to L. lutra.
Paleoecological implications
Despite its fossil record is less abundant if compared to other Mustelidae taxa (e.g., Meles meles, Mecozzi et al. 2019), otters are largely distributed across the Eurasian continent over a long stratigraphical interval. Currently, this carnivoran inhabits a large variety of aquatic habitats, including highland and lowland lakes, rivers, streams, marshes, swamp forests and coastal areas independent of their size, origin or latitude (Mason and Macdonald 2009; Hung and Law 2016). It can be found in brackish waters from sea level up to 4120 m (Mason and Macdonald 2009). Similarly to other European carnivorans, the extant populations of L. lutra suffer the strong human pressure, which mainly reflects the habitat loss (Toschi 1965; Prigioni 2003). In the Italian Peninsula, the presence of the Eurasian otter is nowadays confined in the southern regions, with stable populations reported also in northern Apulia (Toschi 1965; Prigioni 2003). Thus, the occurrence of L. lutra from Grotta Romanelli is surprisingly, since the geographical position of the site falls outside the range of the extant species. Moreover, its ecological profile indicates a strong relation to large freshwater bodies (rivers or lakes), which are no longer present in the southern part of the Apulia (Mastronuzzi and Sansò 2017). Therefore, its presence strongly supports a more diversified hydrographic network in this territory during the late Quaternary.
Body mass estimates reveal a large overlap between L. lutra and L. simplicidens (Fig. 5), which also share a fish-feeding type of dentition, making difficult to envision a possible coexistence. On the contrary, C. antiqua has similar but slightly larger values of estimated body mass but occupied a different trophic niche. In fact, given its robust dentition with broader teeth, several authors aligned it with Aonyx (e.g., Malatesta 1978; Pennacchioni and Cassola 1986), whose paleobiological interpretation points to a more important role of shellfish in its diet (Willemsen 1992). Following this, a sympatric coexistence scenario of the fish-feeding L. simplicidens and L. lutra with C. antiqua could be possible, with different craniodental features which reflect the diversification of ecological niches. The other fossil European otter, Lutraeximia umbra, according to Cherin et al. (2016) had a strong preference for hard prey (molluscs), but possessed intermediate features between fish specialist and hard invertebrates dentition.

Geographical distribution of the body size estimates
Finally, several scholars detected geographic-related body size differences in extant otters, which have been explained according to Bergmann’s ecogeographical rule (Yom-Tov et al. 2006; Meiri et al. 2007), or advocating a detrimental effect of environmental pressures and food availability (Yom-Tov et al. 2010). Despite the paucity of the otter sample prevents definitive conclusions, body size estimates of the fossil specimens would show no dimensional clines across Europe, neither geographic, nor chronological (Fig. 5).
Homo-otter interactions
The finding of the fossil otter in the deposit of Grotta Romanelli opens another question that requires attention. Although the studied specimen lacks both exact stratigraphical and taphonomic information, in level G lithic artifacts referred to Mousterian (Middle Paleolithic) were also found, which document the human occupation of the cave. The potential Homo-Lutra interactions during Pleistocene are indeterminate, and in need of an in-depth emphasis from both paleoecological and archaeozoological points of view. Man exploited carnivorans for a wide range of purposes, among which food and fur, but even for bone artifact industry, symbolism, and ornamental purposes (Mallye 2007, 2011). The Homo medium-to-small-sized carnivoran interactions are difficult to investigate due to the fragility of the skeletal bones (Kurtén 1968). Indeed, the majority of the carnivoran records from archeological deposits are represented by fragmentary craniodental, metapodial and phalange remains. This is probably due to selective human exploitation, since the skull and distal portion of the limbs have lesser nutritional return. In addition, the carnivoran species are usually rare in archeological deposits, as for instances Riparo Mochi (2.9%, Tagliacozzo et al. 2012), Arene Candide (27.7%, Cassoli and Tagliacozzo 1994a), Riparo Fumane (16.8%, Cassoli and Tagliacozzo 1994b), Grotta Maggiore di San Bernardino (26.5%, Cassoli and Tagliacozzo 1994c) and upper complex (level E–A) of Grotta Romanelli (26.9%, Tagliacozzo 2003). However, the presence of carnivorans and Homo in the same deposit not necessarily indicates that these taxa were hunted. Cassoli and Tagliacozzo (1994a), analyzing the fossil remains from the long sedimentary succession of Arene Candide, suggested that the majority of the carnivoran fossil remains do not show cut mark and often were found in anatomical connection. The authors also identified bite marks in a few carnivoran specimens, as well as some ungulate ones, suggesting therefore that this cave was also seasonally frequented by large carnivorans (leopard or hyaena) or more stable occupied by bear, where the prevalence of young individuals has been interpreted as its possible den. Recently, a natural accumulation of an abundant sample of L. lutra from Bliznets Cave (Russian Far East) has been described, testifying that otters can visit caves as well as perish in them (Alekseeva and Baryshnikov 2020). In this scenario, who hunted the L. lutra from Grotta Romanelli cannot be ascertained, even if the abundance of the Middle Paleolithic artifacts from level G points to the Neanderthal as possible responsible, which could have included otter in its diet.
Conclusion
The Eurasian otter from the late Middle Pleistocene of Grotta Romanelli represents one of the earliest evidences of the species, previously known only since the Holocene onwards. The comparison analyses allow to exclude an attribution to L. simplicidens and C. antiqua. L. simplicidens and L. lutra have a similar size and share a fish-feeding type of dentition, but they are distinct by the proportions of their lower carnassial teeth and morphological features of the upper carnassial. C. antiqua has a shellfish-feeding dentition, broader and more robust than that of L. lutra. However, here, we recognize that the two species overlap in chronological distribution, which urges to exercise caution in the attribution of isolated remains.
The occurrence of the Eurasian otter from the “terre rosse” of Grotta Romanelli provides valuable paleoecologial information. L. lutra, nowadays, is mainly living in fresh and clear running waters or lakes, and a certain degree of adaptation to brackish water is possible. Most likely, a hydrographic network could have been occurred in the area in front of the cave, at present under the sea level.
Unfortunately, taphonomic data are not enough to reconstruct who buried the mandible and how, at present the only bone referable to the species, in the “terre rosse”. Anyway, the occurrence of L. lutra at Grotta Romanelli suggests that the taxon spreads into Europe earlier than supposed before, at least at the beginning of the Late Pleistocene or during the late Middle Pleistocene, replacing L. simplicidens and coexisting with C. antiqua.
Genetic evidence resume L. lutra as closely related with the extant L. sumatrana (Hung and Law 2016), and it has been suggested that both derived from the Upper Siwalik L. palaeindica (Willemsen 1992). However, despite the lack of cranial material prevents conclusive comparisons, our analyses reveal strong morphological affinities between L. palaeindica and L. simplicidens.
Further discoveries will shed new light on systematics and evolution of these carnivorans in the Quaternary of Europe.
References
Alekseeva, E., and G.F. Baryshnikov. 2020. Late Pleistocene and Holocene remains of carnivorous mammals (Carnivora) from the Bliznets Cave in southern part of Russian Far East. Proceedings of Zoological Institute, Russian Academy of Sciences 324(3): 388–444. (in Russian, with English summary).
Alessio, M., F. Bella, and C. Cortesi. 1964. University of Rome carbon14 dates II. Radiocarbon 6: 77–90.
Alessio, M., F. Bella, F. Bachecchi, and C. Cortesi. 1965. University of Rome carbon14 dates III. Radiocarbon 7: 213–222.
Baryshnikov, G.F. 2015. Late Pleistocene Ursidae and Mustelidae remains (Mammalia, Carnivora) from Geographical Society Cave in the Russian Far East. Proceedings of the Zoological Institute RAS 319(1): 3–22.
Baryshnikov, G.F., and A.Y. Puzachenko. 2012. Craniometrical variation of river otter (Lutra lutra: Carnivora: Mustelidae) in Northern Eurasia. Proceedings of Zoological Institute, Russian Academy of Sciences 316(3): 203–222. (in Russian, with English summary).
Baryshnikov, G.F., A.Y. Puzachenko, and A.V. Abramov. 2003. New analysis of variability of cheek teeth in Eurasian badgers (Carnivora, Mustelidae, Meles). Russian Journal of Theriology 1: 133–149.
Bella, F., A.C. Blanc, G.A. Blanc, and C. Cortesi. 1958. Una prima datazione con il carbonio 14 della formazione pleistocenica di Grotta Romanelli (Terra d’Otranto). Quaternaria 5: 87–94.
Blanc, G.A. 1920. Grotta Romanelli I. Stratigrafia dei depositi e natura e origine di essi. Archivio per l’Antropologia e la Etnologia 50: 1–39.
Blanc, G.A. 1928. Grotta Romanelli II. Dati ecologici e paletnologici. Archivio per l’Antropologia e la Etnologia 58: 1–49.
Bona, F. 2014. La fauna del sito di Tosina. In Contadini, allevatori e artigiani a Tosina di Monzambano tra V e IV millennio a.C. Una comunità neolitica nei circuiti padani e veneti, ed. R. Poggiani Keller, 137–148. Brescia: Grafiche Tagliani Stampa e Comunicazione per Acherdo Edizioni.
Bonaparte, C.L. 1838. Synopsis Vertebratorum Systematis. Nuovi Annali delle Scienze Naturali, Bologna 2: 105–133.
Botti, U. 1874. Sulla scoperta di ossa fossili in Terra d’Otranto. Bollettino del Regio Comitato Geologico 1: 7–8.
Bowdich, T.E. 1821. An analysis of the Natural Classification of Mammalia for the use of students and travellers. Paris: J. Smith.
Brisson, M.J. 1762. Regnum animale in classes IX distributuin sive synopsis distributuin sive synopsis methodica. Editio altera auction. Leiden: Th. Haak.
Calcagnile, L., R. Sardella, I. Mazzini, F. Giustini, M. Brilli, M. D’Elia, E. Braione, J. Conti, B. Mecozzi, F. Bona, D.A. Iurino, G. Lembo, B. Muttillo, and G. Quarta. 2019. New radiocarbon dating results from the Upper Paleolithic-Mesolithic levels in Grotta Romanelli (Apulia, Southern Italy). Radiocarbon 61(5): 1211–1220.
Cassoli, P.F., and A. Tagliacozzo. 1994a. I macromammiferi dei livelli tardopleistocenici delle Arene Candide (Savona, Italia): considerazioni paleontologiche e archeozoologiche. Quaternaria Nova 4: 101–262.
Cassoli, P.F., and A. Tagliacozzo. 1994b. I resti ossei di macromammiferi, uccelli e pesci della Grotta Maggiore di San Bernardino sui Colli Iberici (VI): considerazioni paleoeconomiche, paleoecologiche e cronologiche. Bullettino di Paletnologia Italiana 85: 1–71.
Cassoli, P.F., and A. Tagliacozzo. 1994c. Considerazioni Paleontologiche, paleoecologiche e archezoologiche sui macromammiferi e gli uccelli dei livelli del Pleistocene Superiore del Riparo Fumane (VR) (Scavi 1988–91). Bollettino del Museo Civico di Storia Naturale di Verona 18: 349–445.
Cherin, M. 2017. New material of Lutra simplicidens (Carnivora, Mustelidae, Lutrinae), a key taxon for understanding the evolution of European otters. Rivista Italiana di Paleontologia e Stratigrafia 123(3): 433–441.
Cherin, M., and L. Rook. 2014. First report of Lutra simplicidens (Carnivora, Mustelidae, Lutrinae) in the Early Pleistocene of the Upper Valdarno (Italy) and the origin of European otters. Italian Journal of Geosciences 133: 200–203.
Cherin, M., D.A. Iurino, G. Willemsen, and G. Carnevale. 2016. A new otter from the Early Pleistocene of Pantalla (Italy), with remarks on the evolutionary history of Mediterranean Quaternary Lutrinae (Carnivora, Mustelidae). Quaternary Science Reviews 135: 92–102.
Conroy, J.W.H., P. Yoxon, and A.C. Gutleb. 2000. Proceedings of the First Otter Toxicology Conference. Broadford: International Otter Survival Fund.
Conti, J., S. Tucci, L. Bellucci, F. Bona, M. Brilli, F. Giustini, D.A. Iurino, G. Lembo, M. Massussi, I. Mazzini, B. Mecozzi, B. Muttillo, D. Sigari, F. Strani, M. Voltaggio, and R. Sardella. 2017. Toward the virtual reconstruction of Grotta Romanelli, Apulia (Southern Italy). In IMEKO International Conference on Metrology for Archaeology and Cultural Heritage, Lecce, Italy, October 23–25, 2017, 577–582. Red Hook: Curran Associates, Inc.
Driesch, A. von den. 1976. A guide to the measurement of animal bones from archaeological sites. Bullettin of Peabody Museum of Natural History 1: 1–137.
Falconer, H. 1868. Fauna Antiqua Sivalensis. XVII. On Enhydriodon (Amyxodon), a fossil genus allied to Lutra, from the tertiary strata of the Siwalik Hills. Falconer Palaeontological Memoirs 1: 331–338.
Fejfar, O., and M. Sabol. 2004. Pliocene Carnivores (Carnivora, Mammalia) from Ivanovce and Hajnáčka (Slovakia). Courier Forschungsinstitut Senckenberg 246: 15–53.
Fischer von Waldheim, G. 1817. Adversaria Zoologica. Mémoires de la Société Impériale des Naturalistes de Moscou 5: 368–428.
Fornaca-Rinaldi, G. 1968a. Il metodo 230Th/238U per la datazione di stalattiti e stalagmite. Bollettino di Geofisica and Teoria Applicata 10: 3–14.
Fornaca-Rinaldi, G. 1968b. 230Th/234Th dating of cave concretions. Earth and Planetary Science Letters 5: 120–122.
Fornaca-Rinaldi, G., and A.M. Radmilli. 1968. Datazione con il metodo 230Th/ 238U di stalagmiti contenute in depositi musteriani. Atti della Socità Toscana di Scienze Naturali 75(1): 639–646.
Foster-Turley, P., S. Macdonald, and C. Mason. 1990. Otters: an action plan for their conservation. Brookfield: IUCN/SSC Otter Specialist Group, Chicago Zoological Society.
Geraads, D., Z. Alemseged, R. Bobe, and D. Reed. 2015. Pliocene Carnivora (Mammalia) from the Hadar Formation at Dikika, Lower Awash Valley, Ethiopia Lower Awash Valley, Ethiopia. Journal of African Earth Sciences 107: 28–35.
Gervais, P. 1859. Zoologie et palèontologie françaises. Nouvelles recherches sur les animaux vertébrés dont on trouve les ossements enfouis dans le sol de la France et sur leur comparaison avec les espèces propres aux autres régions du globe. Paris: A. Bertrand.
Gimranov, D.O., and P.A. Kosintsev. 2012. Geographic distribution of the tooth morphotypes of the river otter (Carnivora, Mustelidae, Lutra lutra L., 1758) in Northern Eurasia. Doklady Biological Sciences 443: 84–86.
Giustini, F., F. Bona, M. Brilli, J. Conti, A. D’Agostino, G. Lembo, I. Mazzini, B. Mecozzi, B. Muttillo, and R. Sardella. 2018. An introduction to the Early Holocene eolian deposits of Grotta Romanelli, Apulia, Southern Italy. Alpine and Mediterranean Quaternary 31: 135–139.
Graziosi, P. 1973. L’arte Preistorica in Italia. Firenze: Sansoni.
Hung, N., and C.J. Law. 2016. Lutra lutra (Carnivora: Mustelidae). Mammalian Species 48(940): 109–122.
Iannucci, A., R. Sardella, F. Strani, and B. Mecozzi. 2020. Size shifts in late Middle Pleistocene to Early Holocene Sus scrofa (Suidae, Mammalia) from Apulia (southern Italy): ecomorphological adaptations? Hystrix, the Italian Journal of Mammalogy 31(1): 10–20.
Iurino, D.A., J. Conti, B. Mecozzi, and R. Sardella. 2020. Braincase With Natural Endocast of a Juvenile Rhinocerotinae From the Late Middle Pleistocene Site of Melpignano (Apulia, Southern Italy). Frontiers in Earth Science 8: 94.
Jánossy, D. 1986. Pleistocene vertebrate faunas of Hungary. Development in Palaeontology and Stratigraphy 8: 1–201.
Koepfli, K.P., and R.K. Wayne. 1998. Phylogenetic relationships of otters (Carnivora: Mustelidae) based on mitochondrial cytochrome b sequences. Journal of Zoology 246: 401–416.
Koepfli, K.P., K.A. Deere, G.J. Slater, C. Begg, K. Begg, L. Grassman, M. Lucherini, G. Veron, and R.K. Wayne. 2008. Multigene phylogeny of the Mustelidae: resolving relationships, tempo and biogeographic history of a mammalian adaptive radiation. BMC Biology 6: 10.
Koufos, G.D. 2011. The Miocene carnivore assemblage of Greece. Estudios Geologicos 67: 291–320.
Kruuk, H. 2006. Otters: Ecology, Behavior and Conservation. New York: Oxford University Press.
Kurtén, B. 1968. Pleistocene mammals of Europe. Chicago: Aldine Publishing Co.
Larivière, S., and A.P. Jennings. 2009. The Mustelidae. In Handbook of the Mammals of the World. Volume 1. Carnivores, eds. D.E. Wilson, and R.A. Mittermeier, 564–656. Barcelona: Lynx Edicions
Larsen, E., S.E. Gulliksen, R. Lauritzen, R. Lie, R. Lövlie, and J. Mangerud. 1987. Cave stratigraphy in western Norway; multiple Weichselian glaciations and interstadial vertebrate fauna. Boreas 16: 267–292.
Linnaeus, C. 1758. Systema naturae per régna tria naturae, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Tomus I. Holmiae [=Stockholm]: L. Salvii.
Madurell-Malapeira, J., D.M. Alba, J. Marmi, J. Aurell, and S. Moyà-Solà. 2011a. The taxonomic status of European Plio-Pleistocene badgers. Journal of Vertebrate Paleontology 31(5): 885–894.
Madurell-Malapeira, J., B. Martínez-Navarro, S. Ros-Montoya, M.P. Espigares, I. Toro, and P. Palmqvist. 2011b. The earliest European badger (Meles meles), from the Late Villafranchian site of Fuente Nueva 3 (Orce, Granada, SE Iberian Peninsula). Comptes Rendus Palevol 10(8): 609–615.
Malatesta, A. 1978. On the genus Cyrnaonyx Helbing, 1935 (Mustelidae, Lutrinae) ant its type species. Quaternaria 20: 109–116.
Mallye, J.B. 2007. Les restes de Blaireau en contexte archéologique: Taphonomie, Archéozoologie et éléments de discussion des séquences préhistoriques. Unpublished PhD thesis. Bordeaux: Université Bordeaux 1.
Mallye, J.B. 2011. Badger (Meles meles) remains within caves as an analytical tool to test the integrity stratified sites: the contribution of Unikoté cave (Pyrénées-Altantiques, France). Journal of Taphonomy 9(1): 15–36.
Mason, C.F., and S.M. Macdonald. 2009. Otters: ecology and conservation. Cambridge: Cambridge University Press.
Mastronuzzi, G., and P. Sansò. 2017. The Salento Peninsula (Apulia, Southern Italy): a water-shaped landscape without rivers. In Landscapes and Landforms of Italy, 421–430. Cham: Springer.
Matthew, W.D. 1929. Critical observations upon Siwalik Mammals. Bulletin of American Museum of Natural History 56: 437–560.
Mecozzi, B., D.A. Iurino, D.F. Berté, and R. Sardella. 2017. Canis mosbachensis (Canidae, Mammalia) from the Middle Pleistocene of Contrada Monticelli (Putignano, Apulia, southern Italy). Bollettino della Società Paleontologica Italiana 56(1): 71–78.
Mecozzi, B., A. Chakroun, H. Baills, D.A. Iurino, R. Sardella, and D. Coppola. 2018. Late Pleistocene mammal faunal assemblage from Cava Spagnulo (Grottaglie, Apulia, SE Italy). Alpine and Mediterranean Quaternary 31(1): 5–19.
Mecozzi, B., D. Coppola, D.A. Iurino, R. Sardella, and A.M. De Marinis. 2019. The Late Pleistocene European badger Meles meles from Grotta Laceduzza (Brindisi, Apulia, Southern Italy): the analysis of the morphological and biometric variability. The Science of Nature 106: 13.
Meiri, S., Y. Yom-Tov, and E. Geffen. 2007. What determines conformity to Bergmann’s rule? Global Ecology and Biogeography 16: 788–794.
Montoya, P., J. Morales, and J. Abella. 2011. Musteloidea (Carnivora, Mammalia) from the Late Miocene of Venta del Moro (Valencia, Spain). Estudios Geologicos 67: 193–206.
Nowak, R.M. 1999. Walker’s mammals of the world, 6th ed. Baltimore: John Hopkins University Press.
Pandolfi, L., P. Boscato, J. Crezzini, M. Gatta, A. Moroni, M. Rolfo, and A. Tagliacozzo. 2017. Late Pleistocene last occurrence of the narrow-nosed rhinoceros Stephanorhinus hemitoechus (Mammalian, Perissodactyla) in Italy. Rivista Italiana di Paleontologia e Stratigrafia 123(2): 177–192.
Pasa, A. 1947. I mammiferi di alcune antiche brecce veronesi. Memorie del Museo Civico di Storia Naturale di Verona 1: 1–111.
Pennacchioni, G., and F. Cassola. 1986. Le lontre fossili d’Europa e del bacino mediterraneo: sintesi delle attuali conoscenze e considerazioni generali. La lontra in Italia. WWF, Ser. Atti e Studi 5: 127–132.
Poggiani Keller, R. 2014. Tosina di Monzambano: prime considerazioni su cronologia e aspetti culturali. In Contadini, allevatori e artigiani a Tosina di Monzambano tra V e IV millennio a.C. Una comunità neolitica nei circuiti padani e veneti, ed. R. Poggiani Keller, 167–174. Brescia: Grafiche Tagliani Stampa e Comunicazione per Acherdo Edizioni.
Pohle, H. 1919. Die Unterfamilie der Lutrinae. Archiv für Naturgeschichte 85A: 1–247.
Pomel, A. 1843. Note sur une espèce fossile du genre Loutre, dont les ossements ont été recueillis dans les alluvions volcaniques de 1’Auvergne. Bullettin de la Société Géologique de France 1: 168–171.
Prassack, K.A. 2016. Lontra weiri, sp. nov., a Pliocene River Otter (Mammalia, Carnivora, Mustelidae, Lutrinae) from the Hagerman Fossil Beds (Hagerman Fossil Beds National Monument), Idaho, USA. Journal of Vertebrate Paleontology 36(4): e1149075.
Prigioni, C. 2003. Lutra. In Fauna d’Italia 3, eds. L. Boitani, S. Lovari, and A. Vigna Talgianti, 168–178. Bologna: Calderini.
Raghavan, P., M. Pickford, R. Patnaik, and P. Gayathri. 2007. First fossil small-clawed otter, Amblonyx, with a note on some specimens of Lutra, from the Upper Siwaliks, India. Estudios Geológicos 63: 135–146.
Sardella, R., F.D. Bertè, D.A. Iurino, M. Cherin, and A. Tagliacozzo. 2014. The wolf from Grotta Romanelli (Apulia, Italy) and its implications in the evolutionary history of Canis lupus in the Late Pleistocene of Southern Italy. Quaternary International 328: 179–195.
Sardella, R., I. Mazzini, F. Giustini, B. Mecozzi, M. Brilli, D.A. Iurino, G. Lembo, B. Muttillo, M. Massussi, D. Sigari, S. Tucci, and M. Voltaggio. 2018. Grotta Romanelli (Southern Italy, Apulia): legacies and issues in excavating a key site for the Pleistocene of the Mediterranean. Rivista Italiana di Paleontologia e Stratigrafia 124(2): 247–264.
Sardella, R., D.A. Iurino, B. Mecozzi, D. Sigari, F. Bona, L. Bellucci, M. Coltorti, J. Conti, G. Lembo, B. Muttillo, and I. Mazzini. 2019. Grotta Romanelli (Lecce, Southern Italy) Between Past and Future: New Studies and Perspectives for an Archaeo-geosite Symbol of the Palaeolithic in Europe. Geoheritage 11(4): 1413–1432.
Serres, M. de, J. Dubreuil, and JeanJean B. 1839. Recherches sur les ossements humatiles des caverns de Lunel-Viel. Montpellier.
Sigari, D., I. Mazzini, J. Conti, L. Forti, G. Lembo, B. Mecozzi, B. Muttillo, and R. Sardella. 2021, in press. New parietal engravings in the Romanelli cave (Apulia, southern Italy). Towards a systematic review. Antiquity.
Sommer, R., and N. Benecke. 2004. Late- and Post-Glacial history of the Mustelidae in Europe. Mammal Review 34(4): 249–284.
Sotnikova, M., and V. Titov. 2009. Carnivora of the Tamanian faunal unit (the Azov Sea area). Quaternary International 201: 43–52.
Stasi, P.E., and E. Regalia. 1904. Grotta Romanelli stazione con faune interglaciali calde e di steppa. Nota preventiva. Società Italiana di Antropologia 1: 17–81.
Tagliacozzo, A. 2003. Archeozoologia dei livelli dell’Epigravettiano finale di Grotta Romanelli (Castro, Lecce) strategie di caccia ed economia di sussistenza. In Grotta Romanelli Nel Centenario Della Sua Scoperta (1900–2000), eds. P. Fabri, E. Ingravallo, and A. Mangia, 169–216. Lecce: Congedo Editore.
Tagliacozzo, A., F. Zeppieri, I. Fiore, E. Spinapolice, and A. Del Lucchese. 2012. Archaeozoological evidence of subsistence strategies during the Gravettian at Riparo Mochi (Balzi Rossi, Ventimiglia, Imperia-Italy). Quaternary international 252: 142–154.
Thenius, E. 1965. Die Carnivoren-Reste aus dem Altpleistozän von Voigtstedt bei Sangerhausen in Thüringen. Paläontologische Abhandlungen (A: Paläozoologie) 2: 539–564.
Toschi, A. 1965. Fauna d’Italia. Volume VII. Mammalia: Lagomorpha, Rodentia, Carnivora, Ungulata, Cetacea. Calderini: Bologna.
Van Valkenburgh, B. 1990. Skeletal and dental predictors of body mass in carnivores. In Body Size in Mammalian Paleobiology: Estimation and Biological Implications, ed. J. Damuth and B.J. MacFadden, 181–205. Cambridge: Cambridge University Press.
Vogel, J.C., and H.T. Waterbolk. 1963. Groningen radiocarbon dates IV. Radiocarbon 5: 63–202.
Willemsen, G.F. 1992. A revision of the Pliocene and Quaternary Lutrinae from Europe. Scripta Geologica 101: 1–115.
Willemsen, G.F. 2006. Megalenhydris and its relationships to Lutra reconsidered. Hellenic Journal of Geoscience 41: 83–87.
Wilson, D.E., and R.A. Mittermeier. 2009. Handbook of the Mammals of the World. Volume 1. Carnivores. Barcelona: Lynx Edicions.
Yom-Tov, Y., T.M. Heggberget, Ø. Wiig, and S. Yom-Tov. 2006. Body size changes in the Norwegian otter: The possible effects of food availability and global warming. Oecologia 150: 155–160.
Yom-Tov, Y., A. Roos, P. Mortensen, Ø. Wiig, S. Yom-Tov, and T.M. Heggberget. 2010. Recent Changes in Body Size of the Eurasian Otter Lutra lutra in Sweden. Ambio 39: 496–503.
Van Zyll de Jong, C.G. 1972. A systematic review of the nearctic and neotropical river otters (Genus Lutra, Mustelidae, Carnivora). Royal Ontario Museum Life Science Contributions 80: 1–104.
Van Zyll de Jong, C.G. 1987. A phylogenetic study of the Lutrinae (Carnivora; Mustelidae) using morphological data. Canadian Journal of Zoology 65: 2536–2544.
Acknowledgments
The authors are thankful for granting access to the material, the kindness and availability of Francesca Alhaique, director of Archaeozoology laboratory of the Museum of Civilizations (includes the National Prehistoric Ethnographic Museum “Luigi Pigorini”). We are deeply indebted to the Soprintendenza Archeologia, Belle Arti e Paesaggio delle province di Brindisi, Lecce e Taranto for the permission of the research and field activities (2015–2017 and 2018–2020, resp. R. Sardella) (Maria Piccarreta, Laura Masiello and Serena Strafella). Many people provided their competence and help during fieldwork: L. Bellucci, A. Billi, M. Brilli, J. Conti, L. Forti, G. Gaglianone, B. Giaccio, F. Giustini, D.A. Iurino, F. Strani, D. Sigari and many students. N. Ciccarese, T. De Santis, don P. Frisullo, A. Capraro and M. Rizzo for their precious logistic support. Special thanks Luca Forti for the photo of the Grotta Romanelli bay (Fig. 1d). Finally, we thank the reviewers, G.F. Baryshnikov and M. Sabol, who greatly improved this article with constructive criticisms, comments, and suggestions. This work was funded by the Grandi Scavi 2016 (Grant no: SA116154CD9592F3); Grandi Scavi 2017 (Grant no: SA11715C81468801); Grandi Scavi 2018 (Grant no: SA1181642D3B3C58); Grandi Scavi 2019 (Grant no: SA11916B513E7C4B); Grandi Scavi 2020 (Grant no: SA120172B2C05E68).
Funding
Open access funding provided by Università degli Studi di Roma La Sapienza within the CRUI-CARE Agreement..
Author information
Authors and Affiliations
Corresponding author
Additional information
Handling Editor: Irina Ruf.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Mecozzi, B., Iannucci, A., Bona, F. et al. Rediscovering Lutra lutra from Grotta Romanelli (southern Italy) in the framework of the puzzling evolutionary history of Eurasian otter. PalZ 96, 161–174 (2022). https://doi.org/10.1007/s12542-021-00553-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12542-021-00553-y