
Drainage and Stand Growth Response in Peatland Forests—Description, Testing, and Application of Mechanistic Peatland Simulator SUSI

 , , , , , , and
, , , , , , and
Abstract
:1. Introduction
1.1. Background
1.2. Connections between WT and Stand Growth
- (i)
- High : Low oxygen () availability disturbs root metabolism and reduces photosynthesis.
- (ii)
- Low : Limited water supply leads to stomatal closure and reduces photosynthesis.
- (iii)
- Nutrient disorders: Limited nutrient supply reduces biomass accumulation.
- (a)
- Is more pronounced and occurs faster in fertile than infertile sites [22];
- (b)
- (c)
- Correlates with the late summer , so that growth is better the deeper is the [25];
- (d)
- Is not affected by high in the spring and early summer [25];
- (e)
- Is delayed in the sense that deep in the late summer increases tree growth during the following growing season [26];
- (f)
1.3. WT and Forest Management
1.4. Call for a Mechanistic Model and the Aims of the Study
2. Materials and Methods
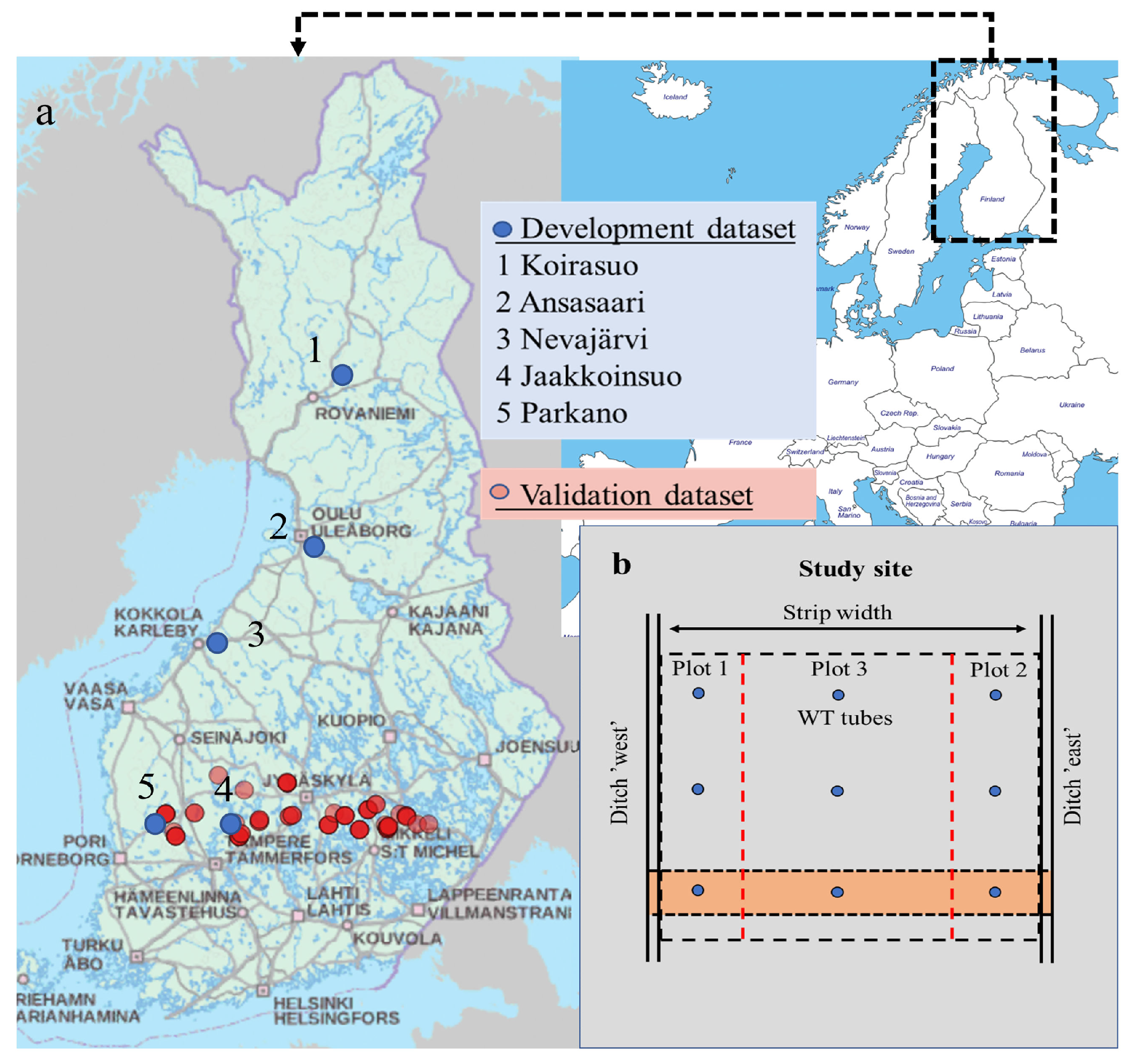
2.1. Data for Model Development
2.2. Validation Data
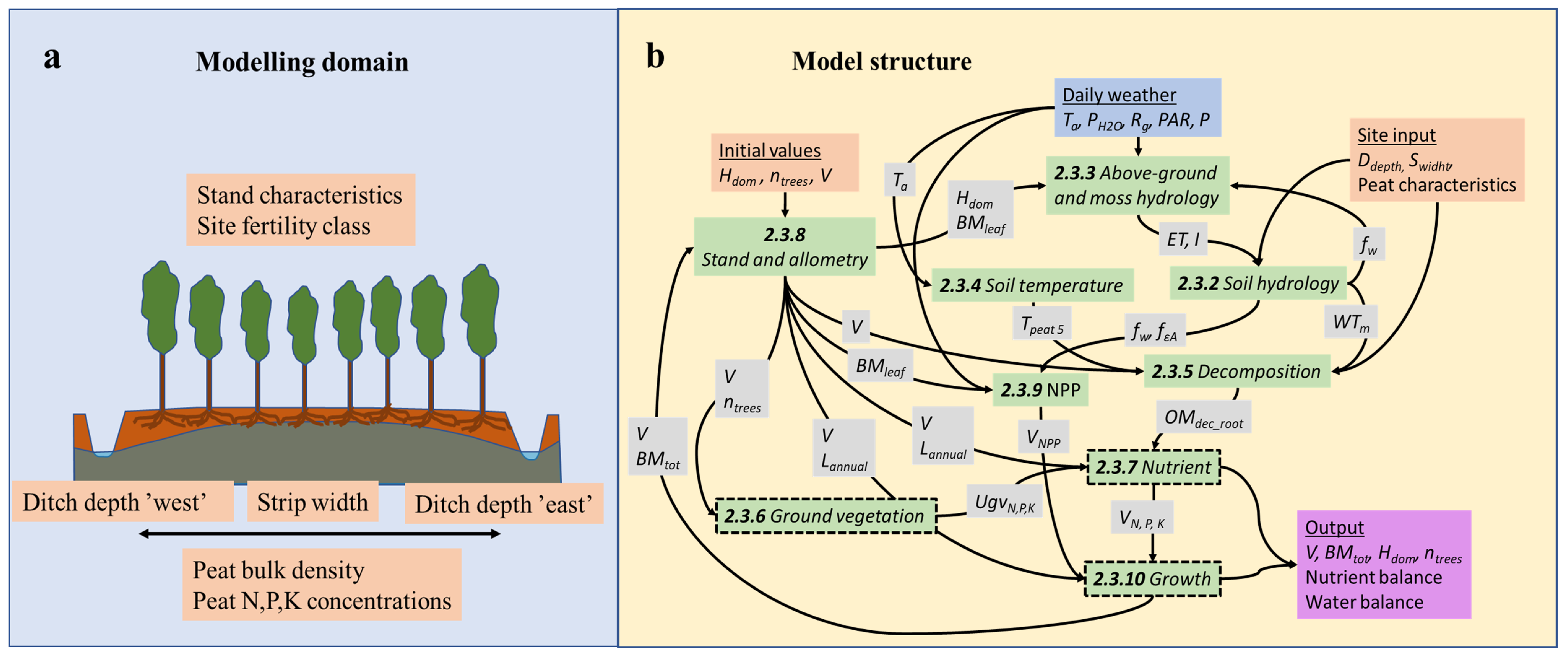
2.3. Peatland Simulator SUSI
2.3.1. General Description
2.3.2. Soil Hydrology Module: Dynamics
2.3.3. Aboveground Hydrology Module: Infiltration and Evapotranspiration
2.3.4. Peat Temperature Module
2.3.5. Organic Matter Decomposition Module
2.3.6. Ground Vegetation Module: Nutrient Demand
2.3.7. Nutrient Module: Release, Allocation and Uptake of Nutrients
2.3.8. Stand and Allometry Module: Allometry and Growth Path
2.3.9. Net Primary Production Module:
2.3.10. Growth Module: Biomass Growth and Yield
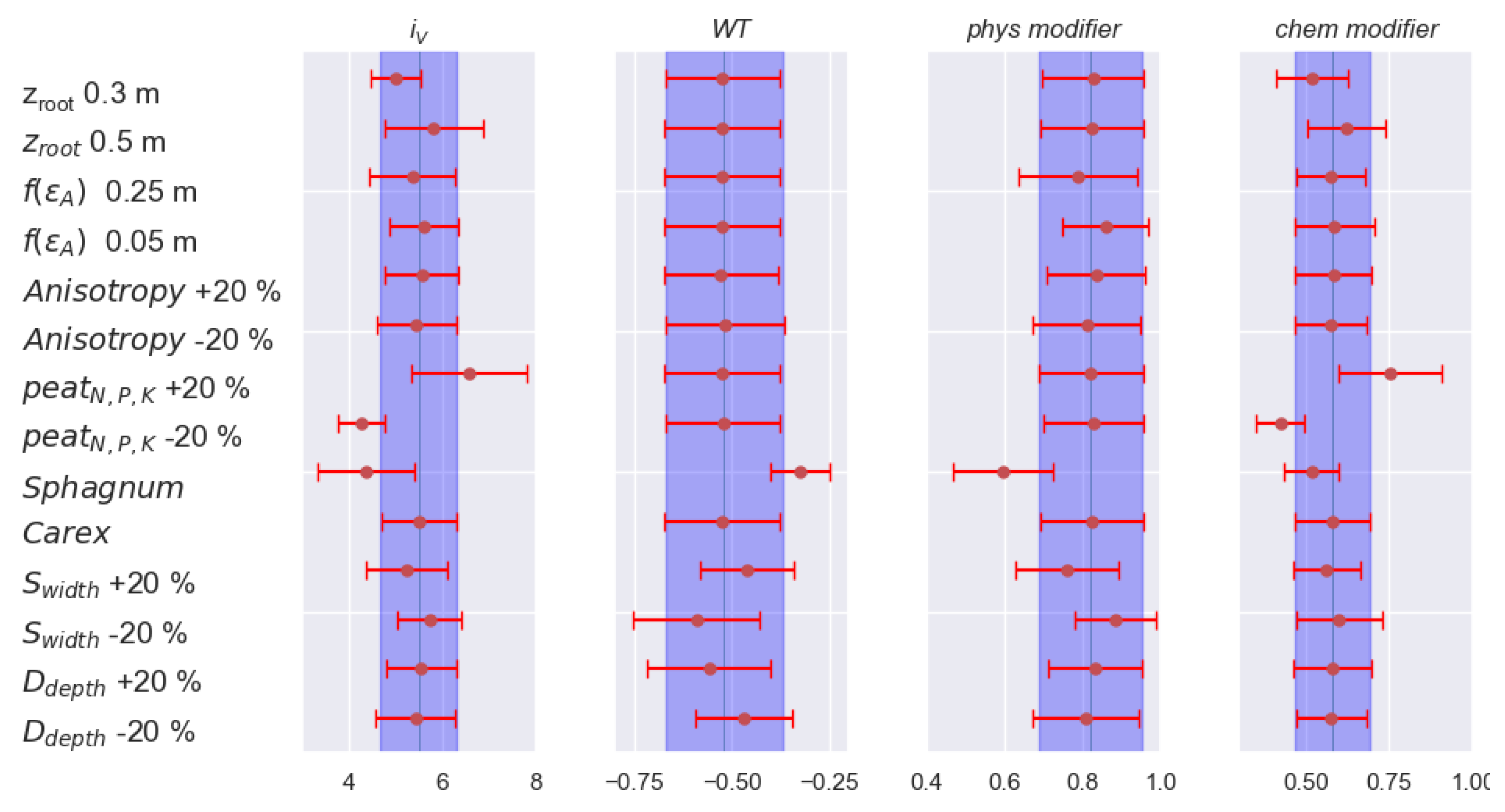
2.4. Sensitivity Analysis
2.5. Model Application to Ditch Network Maintenance
3. Results
3.1. Model Development and Sensitivity
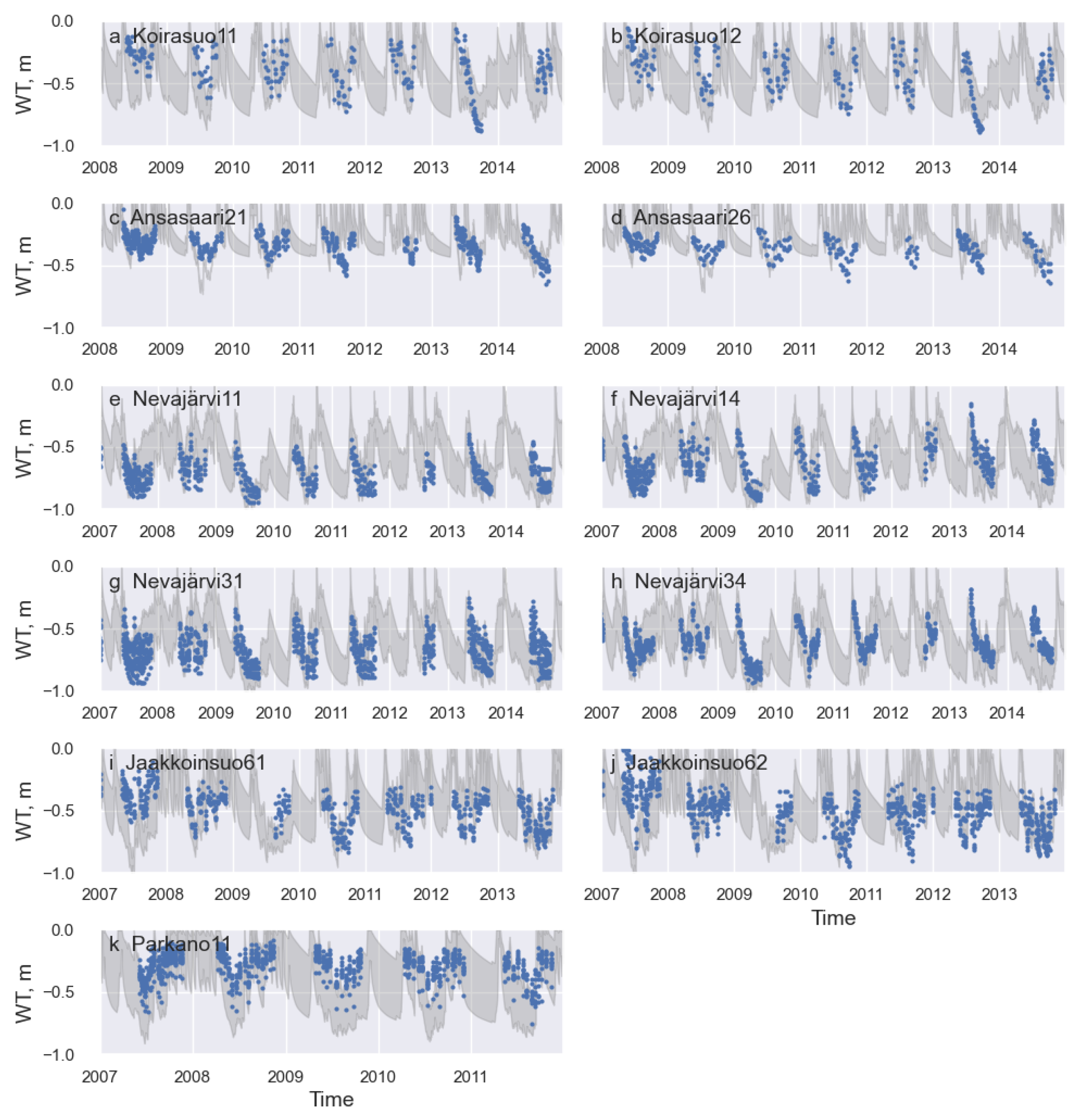
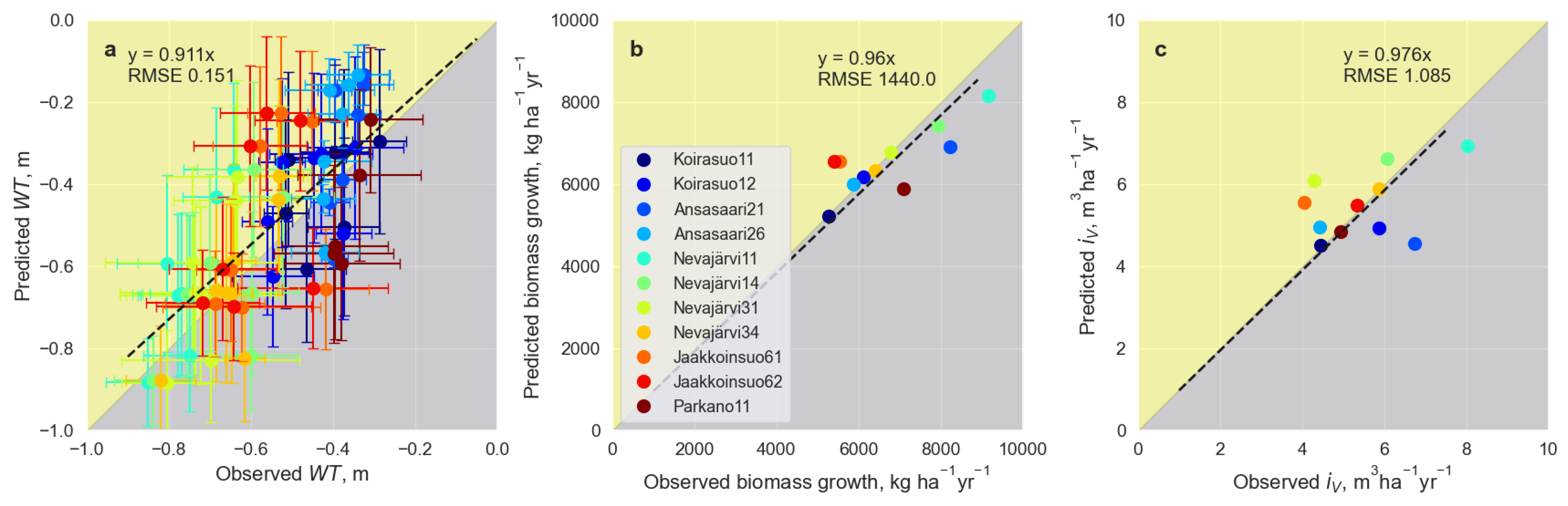
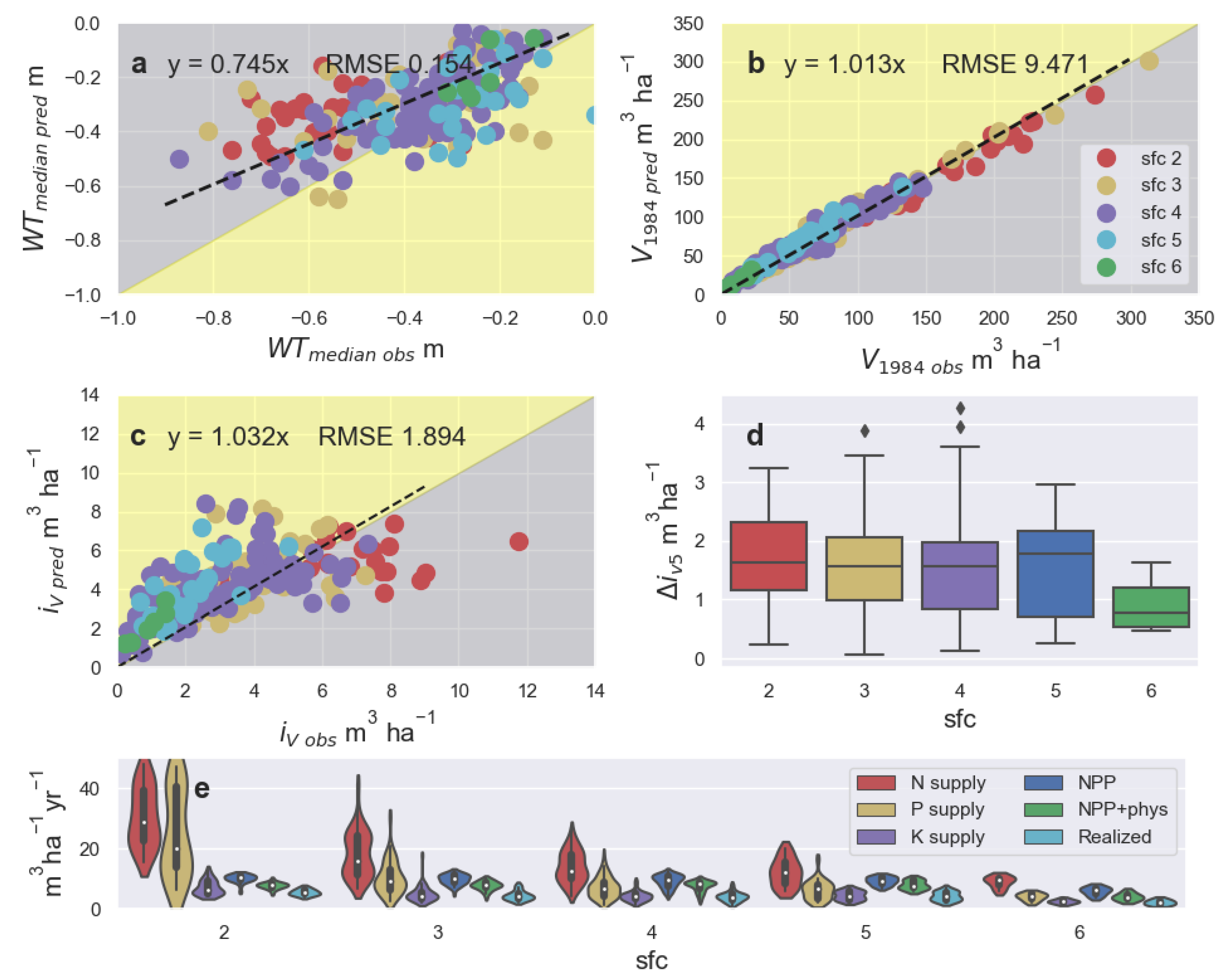
3.2. Model Validation and Application to Ditch Network Maintenance
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| Parameters and variables specific to field data | |
| Initial age of the stand, years | |
| Ditch depth in the validation data, m, negative down | |
| Ditch depth in the validation data, m, negative down | |
| Degree of decomposition of peat, class variable [1,10] | |
| Number of growing seasons | |
| Number of ground water tubes | |
| Number of plots | |
| Number of sites | |
| Annual precipitation, mm | |
| Temperature sum, 5 °C threshold, degree days | |
| Stand volume at the end of the measuring period, m3 ha−1 | |
| Initial stand volume, m3 ha−1 | |
| Median of measured growing season water tables, m, negative down | |
| Mean observed water table, m, negative down | |
| Weather variables | |
| P | Precipitation, mm day−1 |
| Photosynthetically active radiation, W m−2 | |
| Partial pressure of water vapor, hPa | |
| Global radiation, W m−2 | |
| Air temperature, °C | |
| Stand parameters and variables | |
| A | Stand age, years |
| Basal area, m2 ha−1 | |
| Stand biomass in component i: leaf, bark, branch, stem, stump, roots, kg ha−1 | |
| Stand total biomass, kg ha−1 | |
| Biomass yield between time points and , kg ha−1 yr−1 | |
| Tree breast height diameter, cm | |
| h | Tree height, m |
| Dominant height, m | |
| Stand volume growth, m3 ha−1 yr−1 | |
| Site parameters | |
| Peat bulk density, kg m−3 | |
| Ditch depth, m, negative down | |
| N, P, K concentration in peat, mg g−1 | |
| Site fertility class, decreasing fertility from class 1 to 6, class variable [1,6] | |
| Strip width, m | |
| Hydrology and soil parameters and variables | |
| Volumetric air content in peat, m3 m−3 | |
| Soil water potential, m H2O | |
| Volumetric water content in peat, m3 m−3 | |
| Horizontal /vertical ratio applied in the surface peat | |
| Water storage coefficient, m m−1 | |
| Thermal diffusivity of peat, m2 s−1 | |
| Snow depth, m | |
| Evapotranspiration, m day−1 | |
| Physical growth modifying factor, [0,1] | |
| Soil moisture feedback function in computation | |
| Elevation of saturated water column (elevation of water table), m | |
| I | Infiltration, m day−1 |
| Depth of impermeable bottom in peat profile, m | |
| Saturated hydraulic conductivity, m s−1 | |
| S | Sink/source, m s−1 |
| t | Time in simulation, s |
| Temperature in peat in 0.05 m layers, °C | |
| Temperature in peat in 0.05 m depth, °C | |
| Transmissivity of soil profile, m−1 s−1 | |
| W | Water storage in the peat profile, m |
| Water table depth, m, negative down | |
| Mean growing season (May–Sep) water table, m, negative down | |
| Depth in soil, m | |
| Decomposition variables | |
| Decomposed organic matter in the whole peat profile, kg ha−1 yr−1 | |
| Decomposed organic matter in the rooting zone, kg ha−1 yr−1 | |
| CO2 released in heterotrophic respiration, g CO2 m−2 d−1 | |
| Heterotrophic respiration in reference temperature, kg ha−1 day−1 | |
| Refference soil temperature at 0.05 m depth, °C | |
| Temperature in which heterotrophic respiration is zero, °C | |
| Rooting zone depth, m | |
| NPP parameters and variables | |
| Potential daily light use efficiency, g C mol−1 | |
| Absorbed PAR, mol m−2 day−1 | |
| Modifying factors for suboptimal conditions, [0,1] | |
| NPP/GPP ratio, [0,1] | |
| Beer–Lambert light extinction parameter | |
| Net primary production, kg ha−1 day−1 | |
| Annual net primary production, kg ha−1 yr−1 | |
| Ground vegetation parameters and variables | |
| Ground vegetation component i: [dwarf shrub, herbs and sedges, mosses] | |
| Biomass in component i, kg ha−1 | |
| Leaf / green biomass in ground vegetation, kg ha−1 | |
| Content of N,P,K in component i, kg ha−1 | |
| Annual litterfall from component i, kg ha−1 yr−1 | |
| Concentration of N,P,K in component i, mg g−1 | |
| Retranslocation of N, P, K in component i, kg kg−1 | |
| Annual turnover of BM in component i, yr−1 | |
| Uptake demand of N, P, K for component i, kg ha−1 yr−1 | |
| Nutrient parameters and variables | |
| Content of N,P, K in the stand, kg ha−1 | |
| Atmospheric deposition of N, P, K, kg ha-−1 yr−1 | |
| Chemical growth modifying factor, [0,1] | |
| Parameter allocating nutrient supply to ground vegetation, kg kg−1 | |
| Immobilization of N,P, K to microbes, kg kg−1 | |
| N,P, and K lost in litterfall, kg ha−1 yr−1 | |
| Leaching of N, P, K to watercourse, kg ha−1 yr−1 | |
| Parameter allocating nutrient suppy to litter, kg kg−1 | |
| Retranslocation of N,P, K, kg kg−1 | |
| Supply of N,P, K, kg ha−1 yr−1 | |
| Supply of nutrients available for stand growth, kg ha−1 yr−1 | |
| Realized ground vegetation nutrient uptake, kg ha−1 yr−1 | |
References
- Renou-Wilson, F.; Bolger, T.; Bullock, C.; Convery, F.; Curry, J.; Ward, S.; Wilson, D.; Müller, C. BOGLAND - Sustainable Management of Peatlands in Ireland; Environmental Protection Agency: Washington, DC, USA, 2011; p. 157. [Google Scholar]
- Päivänen, J.; Hånell, B. Peatland Ecology and Forestry—A Sound Approach; Helsingin yliopiston metsätieteiden laitoksen julkaisuja; Helsingin Yliopiston Metsätieteiden Laitos: Helsinki, Finland, 2012. [Google Scholar]
- Xu, J.; Morris, P.J.; Liu, J.; Holden, J. PEATMAP: Refining estimates of global peatland distribution based on a meta-analysis. CATENA 2018, 160, 134–140. [Google Scholar] [CrossRef] [Green Version]
- Minkkinen, K.; Laine, J. Long-term effect of forest drainage on the peat carbon stores of pine mires in Finland. Can. J. For. Res. 1998, 28, 1267–1275. [Google Scholar] [CrossRef]
- Ojanen, P.; Minkkinen, K.; Alm, J.; Penttilä, T. Soil–atmosphere CO2, CH4 and N2O fluxes in boreal forestry-drained peatlands. For. Ecol. Manag. 2010, 260, 411–421. [Google Scholar] [CrossRef]
- Jauhiainen, J.; Hooijer, A.; Page, S.E. Carbon dioxide emissions from an Acacia plantation on peatland in Sumatra, Indonesia. Biogeosciences 2012, 9, 617–630. [Google Scholar] [CrossRef] [Green Version]
- Nieminen, M.; Sallantaus, T.; Ukonmaanaho, L.; Nieminen, T.M.; Sarkkola, S. Nitrogen and phosphorus concentrations in discharge from drained peatland forests are increasing. Sci. Total Environ. 2017, 609, 974–981. [Google Scholar] [CrossRef]
- Boggie, R.; Miller, H.G. Growth of Pinus contorta at Different Water-Table Levels in Deep Blanket Peat. For. Int. J. For. Res. 1976, 49, 123–131. [Google Scholar] [CrossRef]
- Dunn, S.; Mackay, R. Modelling the hydrological impacts of open ditch drainage. J. Hydrol. 1996, 179, 37–66. [Google Scholar] [CrossRef]
- Holden, J. Peatland hydrology and carbon release: Why small-scale process matters. Philos. Trans. R. Soc. A Math. Phys. Eng. Sci. 2005, 363, 2891–2913. [Google Scholar] [CrossRef] [Green Version]
- Lukkala, O.J. Tutkimuksia Soiden metsätaloudellisesta ojituskelpoisuudesta erityisesti kuivatuksen tehokkuutta silmälläpitäen. Referat: Untersuchungen über die Waldwirtschaftliche entwässerrungsfähigkeit der Moore mit besonderer rücksicht auf den trocknungssefekt. Commun. Instituti For. Fenn. 1929, 15, 1–278. [Google Scholar]
- Seppälä, K. Ditch spacing as a regulator of post-drainage stand development in spruce and in pine swamps. Acta For. Fenn. 1972, 125, 1–25. [Google Scholar] [CrossRef] [Green Version]
- Hånell, B. Postdrainage forest productivity of peatlands in Sweden. Can. J. For. Res. 1988, 18, 1443–1456. [Google Scholar] [CrossRef]
- Heikurainen, L. Skogsdikning; P.A. Norstedt & Söners förlag: Stockholm, Sweden, 1973; pp. 1–444. [Google Scholar]
- Tóth, J.; Gillard, D. Experimental design and evaluation of a peatland drainage system for forestry by optimization of synthetic hydrographs. Can. J. For. Res. 1988, 18, 353–373. [Google Scholar] [CrossRef]
- Braekke, F.H. Nutrient relationships in forest stands: Effects of drainage and fertilization on surface peat layers. For. Ecol. Manag. 1987, 21, 269–284. [Google Scholar] [CrossRef]
- Vompersky, S.; Rubtsov, V.V.; Dudorov, A. The method for determining a forest drainage network parameters by modelling the ground water. In Socio-Economic Impacts of the Utilization of Peatlands in Industry and Forestry, Proceedings of IPS Symposium in Oulu, Finland; CRC Press, Lewis Publishers: Boca Raton, FL, USA, 1986; pp. 119–126. [Google Scholar]
- Wesseling, J.; van Wijk, W. Soil physical conditions in relation to drain depth. In Drainage of Agricultural Lands; American Society of Agronomy: Madison, WI, USA, 1957; pp. 461–504. [Google Scholar]
- Paavilainen, E. Männyn juuriston suhteesta turpeen ilmatilaan. Summary: Relationship between the root system of Scots pine and the air content of peat. Commun. Instituti For. Fenn. 1967, 63, 1–20. [Google Scholar]
- Paavilainen, E.; Päivänen, J. Forest Management on Peatlands; Springer: Berlin/Heidelberg, Germany, 1995; Volume 111, pp. 143–179. [Google Scholar] [CrossRef]
- Fabrika, M.; Pretzsch, H. Forest Ecosystem Analysis and Modelling; Technical University in Zvolen, Faculty of Forestry, Department of Forest Management and Geodesy: Zvolen, Slovakia, 2013; pp. 1–620. [Google Scholar]
- Seppälä, K. Kuusen ja männyn kasvun kehitys ojitetuilla turvemailla. Acta For. Fenn. 1969, 93, 1–89. [Google Scholar] [CrossRef]
- Hökkä, H.; Ojansuu, R. Height development of Scots pine on peatlands: Describing change in site productivity with a site index model. Can. J. For. Res. 2004, 34, 1081–1092. [Google Scholar] [CrossRef]
- Sarkkola, S.; Nieminen, M.; Koivusalo, H.; Laurén, A.; Ahti, E.; Launiainen, S.; Nikinmaa, E.; Marttila, H.; Laine, J.; Hökkä, H. Domination of growing-season evapotranspiration over runoff makes ditch network maintenance in mature peatland forests questionable. Mires Peat 2013, 11, 1–11. [Google Scholar]
- Pelkonen, E. Vuoden eri aikoina korkealla olevan pohjaveden vaikutus männyn kasvuun. Summary: Effects on Scots pine growth of ground water adjusted to the ground surface for periods of varying length during different seasons of the year. Suo 1975, 26, 25–32. [Google Scholar]
- Huikari, O.; Paarlahti, K. Results of field experiments on the ecology of pine, spruce, and birch. Commun. Instituti For. Fenn. 1968, 64, 1–135. [Google Scholar]
- Moilanen, M.; Hytönen, J.; Hökkä, H.; Ahtikoski, A. Fertilization increased growth of Scots pine and financial performance of forest management in a drained peatland in Finland. Silva Fenn. 2015, 49, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Gliński, J.; Stępniewski, W. Soil Aeration and Its Role for Plants; CRC Press: Boca Raton, FL, USA, 1985; pp. 1–229. [Google Scholar] [CrossRef]
- Kozlowski, T.T. Soil aeration and growth of forest trees (review article). Scand. J. For. Res. 1986, 1, 113–123. [Google Scholar] [CrossRef]
- Bartholomeus, R.P.; Witte, J.P.M.; van Bodegom, P.M.; van Dam, J.C.; Aerts, R. Critical soil conditions for oxygen stress to plant roots: Substituting the Feddes-function by a process-based model. J. Hydrol. 2008, 360, 147–165. [Google Scholar] [CrossRef]
- Verry, E. Hydrological processes of natural, northern forested wetlands. In Northern Forested Wetlands. Ecology and Management; CRC Press, Lewis Publishers: Boca Raton, FL, USA, 1997; pp. 163–188. [Google Scholar] [CrossRef]
- Kaunisto, S.; Paavilainen, E. Nutrient stores in old drainage areas and growth of stands. Comminicationes Instituti For. Fenn. 1988, 145, 1–39. [Google Scholar]
- Moilanen, M.; Saarinen, K.; Silfverberg, K. Foliar nitrogen, phosphorus and potassium concentrations of Scots pine in drained mires in Finland. Silva Fenn. 2010, 44, 583–601. [Google Scholar] [CrossRef] [Green Version]
- Sarkkola, S.; Ukonmaanaho, L.; Nieminen, T.; Laiho, R.; Laurén, A.; Finér, L.; Nieminen, M. Should harvest residues be left on site in peatland forests to decrease the risk of potassium depletion? For. Ecol. Manag. 2016, 374, 136–145. [Google Scholar] [CrossRef]
- Finér, L. Biomass and nutrient dynamics of Scots pine on a drained ombrotrophic bog. Finn. For. Res. Inst. Res. Pap. 1992, 420, 1–43. [Google Scholar]
- Westman, C.; Laiho, R. Nutrient dynamics of drained peatland forests. Biogeochemistry 2003, 63, 269–298. [Google Scholar] [CrossRef]
- Reddy, K.; Delaune, R. Biogeochemistry of Wetlands: Science and Applications; CRC Press: Boca Raton, FL, USA, 2008; pp. 1–781. [Google Scholar] [CrossRef]
- Holmen, H. De skogliga våtmarkernas växtnäringsförhollande. In Skog på Våtmarker; Skogs-och Jordbrokets Forskningsråd: Stockholm, Sweden, 2008; pp. 29–42. [Google Scholar]
- Hytönen, J.; Silfvenberg, K. Effect of drainage on thermal conditions in peat soils. Kuivatustehon vaikutus turvemaan lämpöoloihin. Folia For. 1991, 780, 1–24. [Google Scholar]
- Vompersky, S.; Sirin, A. Hydrological Processes of Natural, Northern Forested Wetlands: Ecology and Management. In Northern Forested Wetlands. Ecology and Management; CRC Press, Lewis Publishers: Boca Raton, FL, USA, 1997; pp. 189–205. [Google Scholar] [CrossRef]
- Heikurainen, L. Metsäojien syvyyden ja pintaleveyden muuttuminen sekä ojien kunnon säilyminen. Acta For. Fennica. 1957, 65, 1–45. [Google Scholar] [CrossRef] [Green Version]
- Hökkä, H.; Stenberg, L.; Laurén, A. Modeling depth of drainage ditches in forested peatlands in Finland. Balt. For. 2020, 26, 1–9. [Google Scholar] [CrossRef]
- Sikström, U.; Hökkä, H. Interactions between soil water conditions and forest stands in boreal forests with implications for ditch network maintenance. Silva Fenn. 2016, 50. [Google Scholar] [CrossRef] [Green Version]
- Laasasenaho, J. Taper curve and volume functions for pine, spruce and birch. Commun. Instituti For. Fenn. 1982, 108, 1–74. [Google Scholar]
- Jaakko, R. Biomass equations for Scots pine and Norway spruce in Finland. Silva Fenn. 2009, 43, 625–647. [Google Scholar] [CrossRef] [Green Version]
- Hynynen, J.; Ojansuu, R.; Hökkä, H.; Siipilehto, J.; Salminen, H.; Haapala, P. Models for predicting stand development in MELA system. Finn. For. Res. Inst. Res. Pap. 2002, 835, 1–116. [Google Scholar]
- Muukkonen, P.; Mäkipää, R. Empirical biomass models of understorey vegetation in boreal forests according to stand and site attributes. Boreal Environ. Res. 2005, 11, 355–369. [Google Scholar]
- Von Post, L. Sveriges geologiska undersöknings torvinventering och några av dess hittills vunna resultat. Sven. Mosskulturföreningens Tidskr. 1922, 37, 1–27. [Google Scholar]
- Venäläinen, A.; Tuomenvirta, H.; Pirinen, P.; Drebs, A. A Basic Finnish Climate Data Set 1961-200—Description and Illustrations; Finnish Meteorological Institute: Helsinki, Finland, 2005. [Google Scholar] [CrossRef]
- Laine, J.; Vanha-Majamaa, I. Vegetation ecology along a trophic gradient on drained pine mires in southern Finland. Ann. Bot. Fenn. 1992, 29, 213–233. [Google Scholar]
- Hökkä, H.; Repola, J.; Laine, J. Quantifying the interrelationship between tree stand growth rate and water table level in drained peatland sites within Central Finland. Can. J. For. Res. 2008, 38, 1775–1783. [Google Scholar] [CrossRef]
- Skaggs, R.W. A water management model for artificially drained soils. Tech. Bull. North Carol. Agric. Exp. Stn. 1980, 267, 1–54. [Google Scholar]
- Van Genuchten, M.T. A Closed-form Equation for Predicting the Hydraulic Conductivity of Unsaturated Soils. Soil Sci. Soc. Am. J. 1980, 44, 892–898. [Google Scholar] [CrossRef] [Green Version]
- Päivänen, J. Hydraulic conductivity and water retention in peat soils. Acta For. Fennica. 1973, 129, 1–70. [Google Scholar] [CrossRef] [Green Version]
- Koivusalo, H.; Ahti, E.; Laurén, A.; Kokkonen, T.; Karvonen, T.; Nevalainen, R.; Finér, L. Impacts of ditch cleaning on hydrological processes in a drained peatland forest. Hydrol. Earth Syst. Sci. 2008, 12, 1211–1227. [Google Scholar] [CrossRef]
- Haahti, K.; Warsta, L.; Kokkonen, T.; Younis, B.A.; Koivusalo, H. Distributed hydrological modeling with channel network flow of a forestry drained peatland site. Water Resour. Res. 2016, 52, 246–263. [Google Scholar] [CrossRef] [Green Version]
- Launiainen, S.; Guan, M.; Salmivaara, A.; Kieloaho, A.J. Modeling boreal forest evapotranspiration and water balance at stand and catchment scales: A spatial approach. Hydrol. Earth Syst. Sci. 2019, 23, 3457–3480. [Google Scholar] [CrossRef] [Green Version]
- Kuusisto, E. Snow accumulation and snowmelt in Finland. Publ. Water Res. Inst. 1984, 55, 1–149. [Google Scholar] [CrossRef]
- Korhonen, L.; Korhonen, K.; Stenberg, P.; Maltamo, M.; Rautiainen, M. Local Models for Forest Canopy Cover with Beta Regression. Silva Fenn. 2007, 41, 671–685. [Google Scholar] [CrossRef] [Green Version]
- Härkönen, S.; Lehtonen, A.; Manninen, T.; Tuominen, S.; Peltoniemi, M. Estimating forest leaf area index using satellite images: Comparison of k-NN based Landsat-NFI LAI with MODIS-RSR based LAI product for Finland. Boreal Environ. Res. 2015, 20, 181–195. [Google Scholar]
- De Vries, D. Heat Transfer in Soils; Springer: Dordrecht, The Netherlands, 1975; pp. 5–28. [Google Scholar]
- Palviainen, M.; Finér, L.; Mannerkoski, H.; Piirainen, S.; Starr, M. Responses of ground vegetation species to clear-cutting in a boreal forest: Aboveground biomass and nutrient contents during the first 7 years. Ecol. Res. 2005, 20, 652–660. [Google Scholar] [CrossRef]
- Mälkönen, E. Growth, suppression, death, and self-pruning of branches of Scots pine in southern and central Finland. Commun. Instituti For. Fenn. 1974, 838, 1–87. [Google Scholar]
- Ruoho-Airola, T.; Alaviippola, B.; Salminen, K.; Varjoranta, R. An investigation of base cation deposition in Finland. Boreal Environ. Res. 2003, 8, 83–95. [Google Scholar]
- Palviainen, M.; Finér, L. Estimation of nutrient removals in stem-only and whole-tree harvesting of Scots pine, Norway spruce, and birch stands with generalized nutrient equations. Eur. J. For. Res. 2012, 131, 945–964. [Google Scholar] [CrossRef]
- Chertov, O.; Komarov, A.; Nadporozhskaya, M.; Bykhovets, S.; Zudin, S. ROMUL—A model of forest soil organic matter dynamics as a substantial tool for forest ecosystem modeling. Ecol. Model. 2001, 138, 289–308. [Google Scholar] [CrossRef]
- Laiho, R.; Laine, J. Nitrogen and phosphorus stores in Peatlands drained for forestry in Finland. Scand. J. For. Res. 1994, 9, 251–260. [Google Scholar] [CrossRef]
- Laiho, R.; Laine, J. Changes in mineral element concentrations in peat soils drained for forestry in Finland. Scand. J. For. Res. 1995, 10, 218–224. [Google Scholar] [CrossRef] [Green Version]
- Nieminen, T.; Helmisaari, H.S. Nutrient retranslocation in the foliage of Pinus sylvestris L. growing along a heavy metal pollution gradient. Tree Physiol. 1996, 16, 825–831. [Google Scholar] [CrossRef] [Green Version]
- Salminen, H.; Lehtonen, M.; Hynynen, J. Reusing legacy FORTRAN in the MOTTI growth and yield simulator. Comput. Electron. Agric. 2005, 49, 103–113. [Google Scholar] [CrossRef]
- Lamppu, J.; Huttunen, S. Scots pine needle longevity and gradation of needle shedding along pollution gradients. Can. J. For. Res. 2001, 31, 261–267. [Google Scholar] [CrossRef]
- Mäkinen, H. Growth, suppression, death, and self-pruning of branches of Scots pine in southern and central Finland. Can. J. For. Res. 1999, 29, 585–594. [Google Scholar] [CrossRef]
- Mäkelä, A.; Pulkkinen, M.; Kolari, P.; Lagergren, F.; Berbinger, P.; Lindroth, A.; Loustau, D.; Nikinmaa, E.; Vesala, T.; Hari, P. Developing an empirical model of stand GPP with the LUE approach: Analysis of eddy covariance data at five contrasting conifer sites in Europe. Glob. Chang. Biol. 2008, 14, 92–108. [Google Scholar] [CrossRef]
- Valentine, H.T. Estimation of the net primary productivity of even-aged stands with a carbon-allocation model. Ecol. Model. 1999, 122, 139–149. [Google Scholar] [CrossRef]
- Hillel, D. Preface. In Introduction to Environmental Soil Physics; Hillel, D., Ed.; Academic Press: Burlington, VT, USA, 2003; pp. xiii–xvi. [Google Scholar] [CrossRef]
- Ahti, E. Fertilizer-induced leaching of phosphorus and potassium from peatlands drained for forestry. Commun. Instituti For. Fenn. 1983, 111, 1–20. [Google Scholar]
- Nieminen, M.; Laiho, R.; Sarkkola, S.; Penttilä, T. Whole-tree, stem-only and stump harvesting impacts on site nutrient capital of a Norway spruce dominated peatland forest. Eur. J. For. Res. 2016, 135, 531–538. [Google Scholar] [CrossRef]
- Hökkä, H.; Repola, J.; Moilanen, M. Modelling volume growth response of young Scots pine (Pinus sylvetris) stands to N, P, and K fertilization in drained peatland sites in Finland. Can. J. For. Res. 2012, 42, 1359–1370. [Google Scholar] [CrossRef]
- Huotari, N.; Tillman-Sutela, E.; Moilanen, M.; Laiho, R. Recycling of ash—For the good of the environment? For. Ecol. Manag. 2015, 348, 226–240. [Google Scholar] [CrossRef]
- Lauhanen, R.; Piiroinen, M.L.; Penttilä, T.; Kolehmainen, E. Kunnostusojitustarpeen arviointi Pohjois-Suomessa. Evaluation of the need for ditch network maintenance in Northern Finland. Mires Peat 1998, 49, 101–112. [Google Scholar]
- Ahti, E.; Kojola, S.; Nieminen, M.; Penttilä, T.; Sarkkola, S. The effect of ditch cleaning and complementary ditching on the development of drained Scots pine-dominated peatland forests in Finland. In Proceedings of the 13th International Peat Congress. After Wise Use—The Future of Peatlands. Volume 1, Oral Presentations; Farrel, C.F.J., Ed.; International Peat Society: Jyväskylä, Finland, 2008; Volume 1, pp. 457–459. Available online: https://peatlands.org/assets/uploads/2019/06/ipc2008p457-459-Ahti-The-effect-of-ditch-cleaning-and-complementary-ditching.pdf (accessed on 11 November 2020).
- Ahtikoski, A.; Kojola, S.; Hökkä, H.; Penttilä, T. Ditch network maintenance in peatland forest as a private investment: Short- and long-term effects on financial performance at stand level. Mires Peat 2008, 3, 1–11. [Google Scholar]
- Lauhanen, R.; Kaunisto, S. Effect of drainage maintenance on the nutrient status on drained Scots pine mires. Suo 1999, 50, 119–132. [Google Scholar]
- Lieffers, V.; Macdonald, S. Growth and foliar nutrient status of black spruce and tamarack in relation to depth of water table in some Alberta peatlands. Can. J. For. Res. 1990, 20, 805–809. [Google Scholar] [CrossRef]
- Silvola, J. Effect of drainage and fertilization on carbon output and nutrient mineralization of peat. Suo 1988, 1–2, 27–37. [Google Scholar]
- Wang, M.; Talbot, J.; Moore, T.R. Drainage and fertilization effects on nutrient availability in an ombrotrophic peatland. Sci. Total Environ. 2018, 621, 1255–1263. [Google Scholar] [CrossRef]
- Moilanen, M.; Silfverberg, K.; Hokkanen, T.J. Effects of wood-ash on the tree growth, vegetation and substrate quality of a drained mire: A case study. For. Ecol. Manag. 2002, 171, 321–338. [Google Scholar] [CrossRef]
- Ojanen, P.; Penttilä, T.; Tolvanen, A.; Hotanen, J.P.; Saarimaa, M.; Nousiainen, H.; Minkkinen, K. Long-term effect of fertilization on the greenhouse gas exchange of low-productive peatland forests. For. Ecol. Manag. 2019, 432, 786–798. [Google Scholar] [CrossRef]
- Piirainen, S.; Domisch, T.; Moilanen, M.; Nieminen, M. Long-term effects of ash fertilization on runoff water quality from drained peatland forests. For. Ecol. Manag. 2013, 287, 53–66. [Google Scholar] [CrossRef]
- Finér, L.; Lepistö, A.; Karlsson, K.; Räike, A.; Härkönen, L.; Huttunen, M.; Joensuu, S.; Kortelainen, P.; Mattsson, T.; Piirainen, S.; et al. Drainage for forestry increases N, P and TOC export to boreal surface waters. Sci. Total Environ. 2021, 762, 144098. [Google Scholar] [CrossRef] [PubMed]
- Nieminen, M.; Hökkä, H.; Laiho, R.; Juutinen, A.; Ahtikoski, A.; Pearson, M.; Kojola, S.; Sarkkola, S.; Launiainen, S.; Valkonen, S.; et al. Could continuous cover forestry be an economically and environmentally feasible management option on drained boreal peatlands? For. Ecol. Manag. 2018, 424, 78–84. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | sfc | Peat Type | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Koirasuo11 | 7 | 888 | 513 | 76 | 91 | 122 | 2 | LC | 4 | −0.85 | 37 | −0.39 | 3 |
| Koirasuo12 | 7 | 888 | 513 | 76 | 123 | 164 | 2 | LC | 4 | −0.85 | 37 | −0.42 | 3 |
| Ansasaari21 | 7 | 1106 | 497 | 39 | 140 | 187 | 3 | SC…C | 6 | −0.45 | 40 | −0.34 | 6 |
| Ansasaari26 | 7 | 1106 | 497 | 39 | 110 | 141 | 3 | SC…C | 6 | −0.45 | 40 | −0.37 | 3 |
| Nevajärvi11 | 8 | 1079 | 539 | 80 | 163 | 228 | 3 | SC…LC | 5 | −1.04 | 25 | −0.72 | 6 |
| Nevajärvi14 | 8 | 1079 | 539 | 80 | 143 | 191 | 3 | SC…LC | 5 | −1.04 | 25 | −0.62 | 6 |
| Nevajärvi31 | 8 | 1079 | 539 | 80 | 127 | 162 | 3 | C | 5 | −1.08 | 25 | −0.68 | 10 |
| Nevajärvi34 | 8 | 1079 | 539 | 80 | 100 | 147 | 3 | C | 5 | −1.08 | 25 | −0.60 | 10 |
| Jaakkoinsuo61 | 7 | 1195 | 523 | 86 | 144 | 173 | 5 | LS…ErS | 4–8 | −0.85 | 40 | −0.51 | 6 |
| Jaakkoinsuo62 | 7 | 1195 | 523 | 86 | 149 | 187 | 5 | LS…ErS | 4–8 | −0.85 | 40 | −0.53 | 6 |
| Parkano11 | 5 | 1176 | 614 | 130 | 168 | 192 | 5 | LS…S | 3–5 | −0.86 | 65 | −0.32 | 12 |
| Variable | Unit | |||||
|---|---|---|---|---|---|---|
| number of plots | 24 | 45 | 105 | 27 | 6 | |
| number of sites | 8 | 15 | 35 | 9 | 2 | |
| Used in in the initialization of SUSI-simulator validation runs | ||||||
| m | −0.55 (0.14) | −0.49 (0.16) | −0.58 (0.16) | −0.57 (0.17) | −0.66 (0.11) | |
| m | −0.61 (0.07) | −0.53 (0.16) | −0.59 (0.18) | −0.58 (0.15) | −0.65 (0.07) | |
| m | 48 (13) | 59 (18) | 53 (14) | 57 (10) | 50 (0) | |
| kg m−3 | 138.0 (29.0) | 110.0 (26.0) | 104.0 (28.0) | 88.0 (25.0) | 118.0 (11.0) | |
| mg g−1 | 19.27 (2.57) | 16.13 (3.75) | 14.54 (3.32) | 11.86 (2.69) | 15.16 (1.19) | |
| mg g−1 | 1.03 (0.33) | 0.79 (0.34) | 0.64 (0.15) | 0.56 (0.16) | 0.64 (0.08) | |
| mg g−1 | 0.39 (0.08) | 0.37 (0.1) | 0.37 (0.08) | 0.38 (0.08) | 0.29 (0.05) | |
| trees ha−1 | 2017 (644) | 1491 (547) | 1389 (600) | 1091 (321) | 806 (282) | |
| m | 13.5 (3.7) | 9.6 (4.2) | 7.8 (3.3) | 6.7 (2.8) | 5.1 (2.1) | |
| m3 ha−1 | 120.2 (58.2) | 64.4 (58.0) | 44.5 (31.8) | 42.0 (22.0) | 10.2 (4.7) | |
| Used as a measured reference in the SUSI-simulator validation | ||||||
| m3 ha−1 | 153.4 (65.5) | 83.5 (63.3) | 58.9 (39.0) | 52.9 (26.3) | 14.7 (7.0) | |
| m3 ha−1 yr−1 | 6.64 (2.03) | 3.82 (1.56) | 2.87 (1.73) | 2.18 (1.01) | 0.89 (0.49) | |
| m, negative down | −0.58 (0.12) | −0.38 (0.16) | −0.35 (0.14) | −0.3 (0.13) | −0.24 (0.06) | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Laurén, A.; Palviainen, M.; Launiainen, S.; Leppä, K.; Stenberg, L.; Urzainki, I.; Nieminen, M.; Laiho, R.; Hökkä, H. Drainage and Stand Growth Response in Peatland Forests—Description, Testing, and Application of Mechanistic Peatland Simulator SUSI. Forests 2021, 12, 293. https://doi.org/10.3390/f12030293
Laurén A, Palviainen M, Launiainen S, Leppä K, Stenberg L, Urzainki I, Nieminen M, Laiho R, Hökkä H. Drainage and Stand Growth Response in Peatland Forests—Description, Testing, and Application of Mechanistic Peatland Simulator SUSI. Forests. 2021; 12(3):293. https://doi.org/10.3390/f12030293
Chicago/Turabian StyleLaurén, Ari, Marjo Palviainen, Samuli Launiainen, Kersti Leppä, Leena Stenberg, Iñaki Urzainki, Mika Nieminen, Raija Laiho, and Hannu Hökkä. 2021. "Drainage and Stand Growth Response in Peatland Forests—Description, Testing, and Application of Mechanistic Peatland Simulator SUSI" Forests 12, no. 3: 293. https://doi.org/10.3390/f12030293