
Differences in Growth and Log Quality of Douglas-Fir (Pseudotsuga menziesii (Mirb.) Franco) Provenances

1
Department of Forest Protection, Slovenian Forestry Institute, Večna pot 2, 1000 Ljubljana, Slovenia
2
Department of Forestry and Renewable Forest Resources, Biotechnical Faculty, University of Ljubljana, Večna pot 83, 1000 Ljubljana, Slovenia
*
Author to whom correspondence should be addressed.
Forests 2021, 12(3), 287; https://doi.org/10.3390/f12030287
Submission received: 26 January 2021
/
Revised: 16 February 2021
/
Accepted: 25 February 2021
/
Published: 2 March 2021
(This article belongs to the Special Issue Rare, Non-native and Invasive Plant Species: Structure, Properties and Potential Applications)
Abstract
:Douglas-fir (Pseudotsuga menziesii (Mirb.) Franco) is a non-native conifer from western North America that was introduced into European forests at the end of the 19th century. Plantations of Douglas-fir in Europe have shown good performance, quality, and resilience to exacerbating climatic conditions. However, all these qualities strongly depend on provenance. A total of 1061 surviving trees of fifteen different Douglas-fir provenances were measured in a Slovenian provenance trial that was established within the framework of the 1966/1967 IUFRO seed collection program. We found significant differences among provenances with respect to survival rate, growth performance, and log quality. The total recorded yield of the 46-year-old stand was 602.9 m3/ha, and the average survival rate was 43%. The correlation of juvenile tree heights in 1985 and their average breast height diameters in 2017 is positive and significant. Based on vitality and diameter, the best performing provenances were Yelm and Cathlamet. The provenance with the best log quality assessed through branchiness is Jefferson (Olympic Peninsula, western Washington). All the most promising provenances for western Slovenia (Central Europe) originate from the low-altitude western coast of Washington (WACO), with the Cathlamet provenance showing the best combination of good growth, survival rate, and log quality.
1. Introduction
Climatic fluctuations, along with pests and diseases outbreaks, have a considerable impact on forest ecosystems [1,2,3]. Reduced water availability caused by extremely warm and dry conditions is expected to become a major threat to the productivity and stability of forests (especially Norway spruce plantations) in Europe in the coming decades [4,5,6,7,8,9]. Increasing global demand for wood and rising interest in the green economy are likely to lead to changes in forest management [10]. Coping with these challenges will necessitate the new selection of tree species, including economically valuable non-native tree species [1,11,12], with the ability to maintain growth rate in a drier and warmer future climate with more extreme events such as freezing rain, hail, and windthrow [13].
Douglas-fir (Pseudotsuga menziesii (Mirb.) Franco) has been found to be a very useful species for afforestation and reforestation in Central and Western Europe: it originates from the western part of the United States and Canada and was introduced to Europe in the 19th century [14,15]. Currently, Douglas-fir is one of the most important non-native timber species in Western and Central Europe [12,14,16,17]. It has a high growth potential, even exceeding that of Norway spruce [18]. At the same time, Douglas-fir is a relatively undemanding species that copes well with chronic droughts and exhibits relatively high increment rates even when other conifers do not [19,20].
Douglas-fir currently covers an area of 830,707 ha in Europe [21], and this area is expected to increase [16,22]. This is not surprising given its impressive growth performance. In Germany, Douglas-fir is one of the most productive tree species in terms of growth, economic output, and carbon sequestration [16]. On comparable (but not extreme) sites, Douglas-fir’s growth capacity is much higher (15% to 50%) than that of Norway spruce and beech, and even higher (50% to 60% and more) compared to pine and oak [12]. The variation in growth of Douglas-fir in Europe largely (40–50%) depends on its origin [23]; different races are tested in field experiments. Forestry experts were aware of provenances in early 1910, but in 1967 the most important international IUFRO (International Union of Forest Research Organizations) project was set, aiming to preserve and establish genetic resources for supplying nurseries with best seed material [15]. For provenance tests, parameters that indicate age-related dimensions in the development stage are very desirable. Height growth of Douglas-firs at an early development stage is a good predictor for growth in older stages [24]. However, there are still provenances that show good performance in pole stage but later exhibit a drop in performance [24,25].
In its natural habitat, Douglas-fir grows in an extremely wide range of site conditions and accordingly displays high adaptive genetic variability. These facts are reflected in different flushing dates, susceptibility to late or early frost damage, and susceptibility to different pests and diseases [1]. Coastal Douglas-fir (P. menziesii (Mirb.) var. menziesii) grows better in Europe than the interior variety (P. menziesii (Mirb.) var. glauca) and is also more resistant to needle cast (Rhabdocline pseudotsugae) [14]. In Germany, the best growth performance has been found in provenances from Oregon, western Washington, and southeastern British Columbia, all originating from elevations of less than 600 m above sea level (a.s.l.) [1,14]. France has the largest number of Douglas-fir plantations at mid-elevation regions, predominating with provenances from lower altitudes (<450 m a.s.l.) of the western side of the Cascade Range [17,26]. A major challenge for European forestry is therefore to target the most appropriate genetic material (provenance) for selected sites under future climatic conditions [1,12,24,27,28].
In addition to growth performance, which is important for timber production, wood properties for pulp production and log quality for wood processing industry are also very important [26,29]. The quality of coniferous wood depends primarily on the number and diameter of knots [30]. Ramicorns on the lower half of the trunk area also have a detrimental effect on log quality [31]. The number of branches primarily depends on genetics, while branch diameter primarily depends on stand density [32], so branch size could be controlled with planting density [1]. Based on branchiness, the best provenances in Europe are those from the coastal areas of Washington, and those that have the worst branching habit are from southwestern Oregon [33]. Most research related with planting density on Douglas-fir log quality was made in young plantations (<25 years old) [32,33]; thus, research results from older Douglas-fir plantations are rare or lacking.
The aim of our study was to test whether Douglas-fir provenances grown in a 46-year-old IUFRO provenance trial in Brkini (Slovenia) differ with respect to tree survival, growth performance, yield, and log quality. We wished to identify the most suitable or promising provenances for the western part of Slovenia (Central Europe). In addition, one of our research goals was to determine whether the better height growth of a certain provenance in the young stage indicates its better diameter growth in the adult stage. This could be useful in evaluating the future potential of young plantations. Conifers generally have a strong and significant height–diameter correlation [34,35], and our further analysis derives from this correlation. The data on the average provenance heights in 1985 were taken from Breznikar (1991) [36]. Our hypothesis was that the superior height growth rates of individual provenances in the young stages are maintained in the following decades and after, expressed with above-average diameter growth.
2. Materials and Methods
2.1. Study Site and Origin of Provenances
Studied Douglas-fir trees are grown in a provenance trial site named Padež I and belong to the forest district of Sežana, Slovenia (45°36′13′′ N; 14°3′21′′ E). The climate is inland sub-Mediterranean [37] with an average annual temperature of 10.4 °C, average January temperature of 1.3 °C, and average July temperature of 20.1 °C (period 1980–2010). The average annual rainfall is 1306 mm. Precipitation is fairly favorably distributed within the vegetation period, with a slight dip in July and August (climate data from the meteorological station in Ilirska Bistrica (424 m a.s.l.), 16 km from the study site, reference period 1980–2010 [38]). The study site is at 530–580 m a.s.l., the relief is smooth with 5% outcrops, and the soil is a distric brown soil on non-carbonate flysch and decalcified marl.
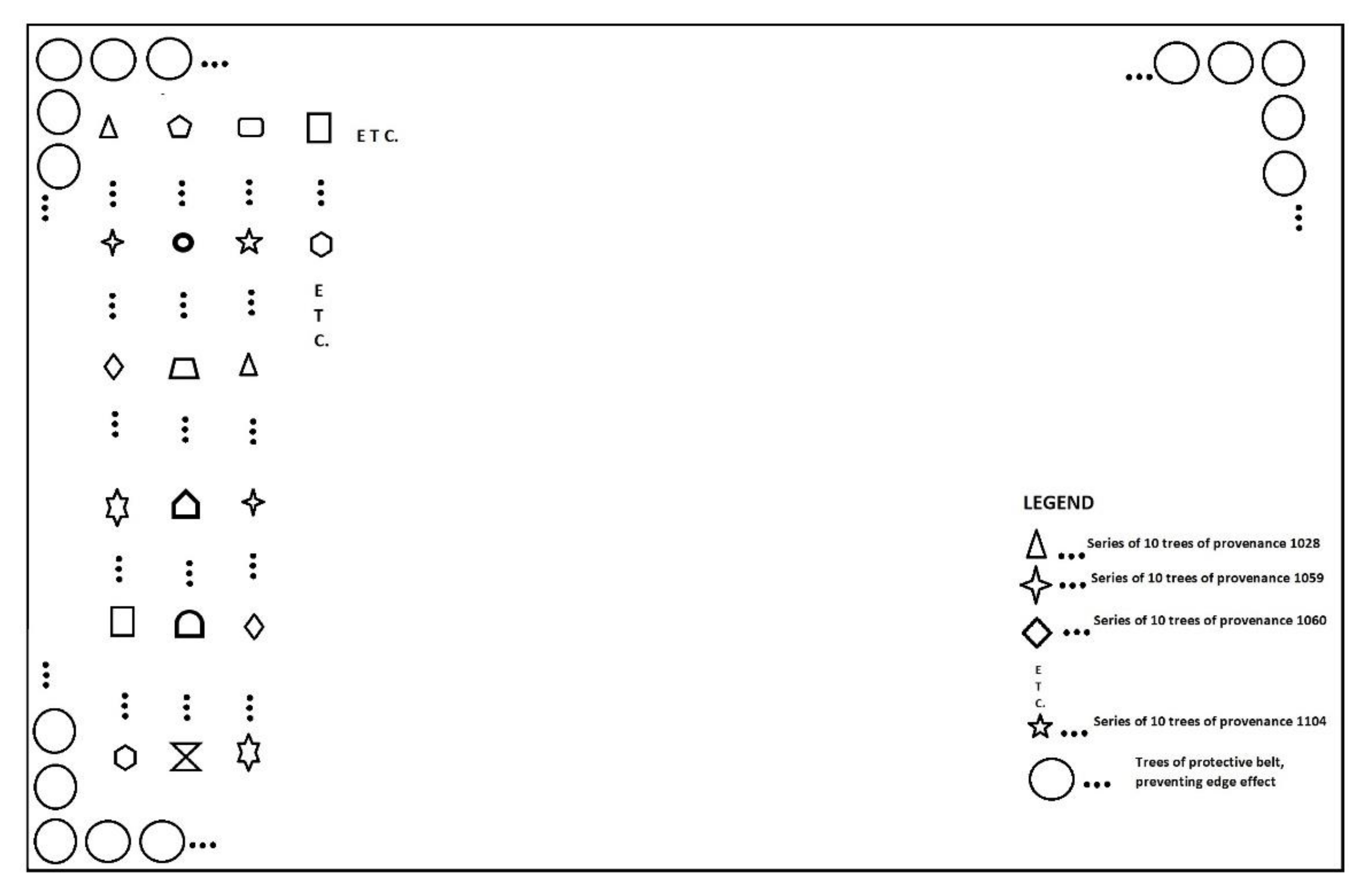
The provenance trial is part of an extensive IUFRO program in which seeds from the natural range of Douglas-fir were collected and distributed to 20 European countries [15]. The provenance trial in Slovenia was established in 1971 with the planting of 15 coastal Douglas-fir (P. menziesii var. menziesii) provenances. The experiment plot was rectangular with an area of 1.56 ha, where 2460 trees of various provenances were planted in rows with 2.5-m spacing. Rows consisted of multiple series of 10 trees per provenance, and there were 11–20 repetitions per provenance, depending on available number of seedlings (Table 1, Figure 1). A protective belt of Douglas-fir trees surrounding the plantation was to prevent edge effects, and provenances were planted in a systematic distribution to exclude environmental factors (small differences in soil, slope). In the establishment phase, the trial was fenced, and planting success was above 90% [39]. Prior to this study, data was collected in 1985 [36], and the trial plantation has never been thinned.
2.2. Field Measurements
Data was collected in April and May of 2017. We performed measurements on all living Douglas-fir trees. Trunk diameter was measured at breast height (dbh1.3, hereafter dbh) and the degree of precision was 1 mm. All trees were classified in vitality classes (3: good vitality, 2: medium vitality, 1: low vitality) according to Leibundgut (1956) [40]. For all trees, several special features that can potentially affect log quality were recorded (multiple trunks and shriveled shoots at a sharp angle (ramicorns)).
We assessed branch number and measured the diameter of branches with small calipers as close as possible to the trunk. All branches with a diameter greater than 8.0 mm that were in a 100–160 cm band above the ground level were measured. The data was used to objectively estimate log quality [32]. Branchiness was not evaluated on all trees but was systematically sampled on the 3rd and 5th live tree in each provenance series. If the number of remaining living trees in a series was less than five, we performed a draw to ensure randomness. Douglas-fir trees with more space for growth (trees along forest trails) were excluded from the analysis of branchiness.
2.3. Statistical Analyses
In the calculations of timber volume, each trunk on multi-trunked trees (forking below breast height) was treated as an autonomous tree. For other analyses (survival, vitality, and dbh), average dbh and average estimates of the vitality of all trunks were used for calculation. Since we were not able to measure tree heights due to the high stand density, for wood stock calculations the 4th tariff class (E4) for even-aged forests were used. Tarif class was determined in forest management plan [41] by the Slovenian Forestry service according to procedure, described in Kotar (2003) [42].
Because certain conditions for homogeneity of variance were not met, a non-parametric Kruskal–Wallis test was used to determine differences among provenances in average dbh and vigor. In the posterior analysis, the Mann–Whitney U-test was used. Due to multiple comparisons, Bonferroni’s correction of the critical p-value [43] was performed.
The differences in branchiness at band 100–160 cm above the ground were checked according to the following criteria: the number of branches (NOB), the average diameter of branches (ADB), the maximum diameter of branches (MDB), and the average diameter of the four thickest branches at breast height (AFB) [32]. The Kruskal–Wallis test was used to test for differences according to individual branchiness criteria, and the pairing between the provenances was done with the Mann–Whitney U-test. When considering the potential impact of provenance and dbh on the number and diameter of branches, an analysis of covariance (ANCOVA) was designed where “provenance” was a fixed factor and dbh was used as a covariate. Growth trend comparisons were tested with the Pearson and Spearman correlation coefficients between the height data from 1985 and diameter data from 2017. All analyzes and computations were performed with IBM SPSS Statistics 25.0 software (IBM Corp.; Armonk, NY, USA), data editing and charts plotting were done using Microsoft office Excel (Microsoft Corp.; Redmond, WA, USA).
3. Results
3.1. Survival Rate, Vitality, and Growth of Provenances
Data analysis showed that the average survival of provenances in the 46-year-old, never thinned, plantation was 43.0% (1061 trees survived out of 2460 planted). The Denny creek (1070) and Cle Elum (1078) provenances had the highest survival rates (55.9% and 53.6%, respectively) (Table 2). The Brookings (1104) provenance from the south coast of Oregon had the lowest survival rate. Total volume according to tariffs was 602.9 m3/ha.
With the Kruskal–Wallis test, we confirmed the effect of provenance on average dbh (H (15) = 40.165, p < 0.001). Yelm (1080) had the highest average dbh (36.1 cm), significantly differing from most of the other provenances (Table A1). In contrast, Merritt (1028) had the lowest average dbh (27.0 cm), also significantly differing from most of the other provenances.
The Kruskal–Wallis test was also used to confirm the effect of provenance on vitality (H (15) = 31.242, p < 0.01). The most vital provenance was Yelm (1080) with an average grade of 2.49, followed by Cathlamet (1089) and Perry Creek (1059) with grades of 2.37 and 2.30, respectively. The Merritt (1028) provenance had the lowest vitality with a grade of 1.87. The Spearman correlation of vitality with average dbh was significant (p < 0.01) and high (rs = 0.873).
3.2. Does Height Growth in Youth Indicate Greater Age-Related Diameter?
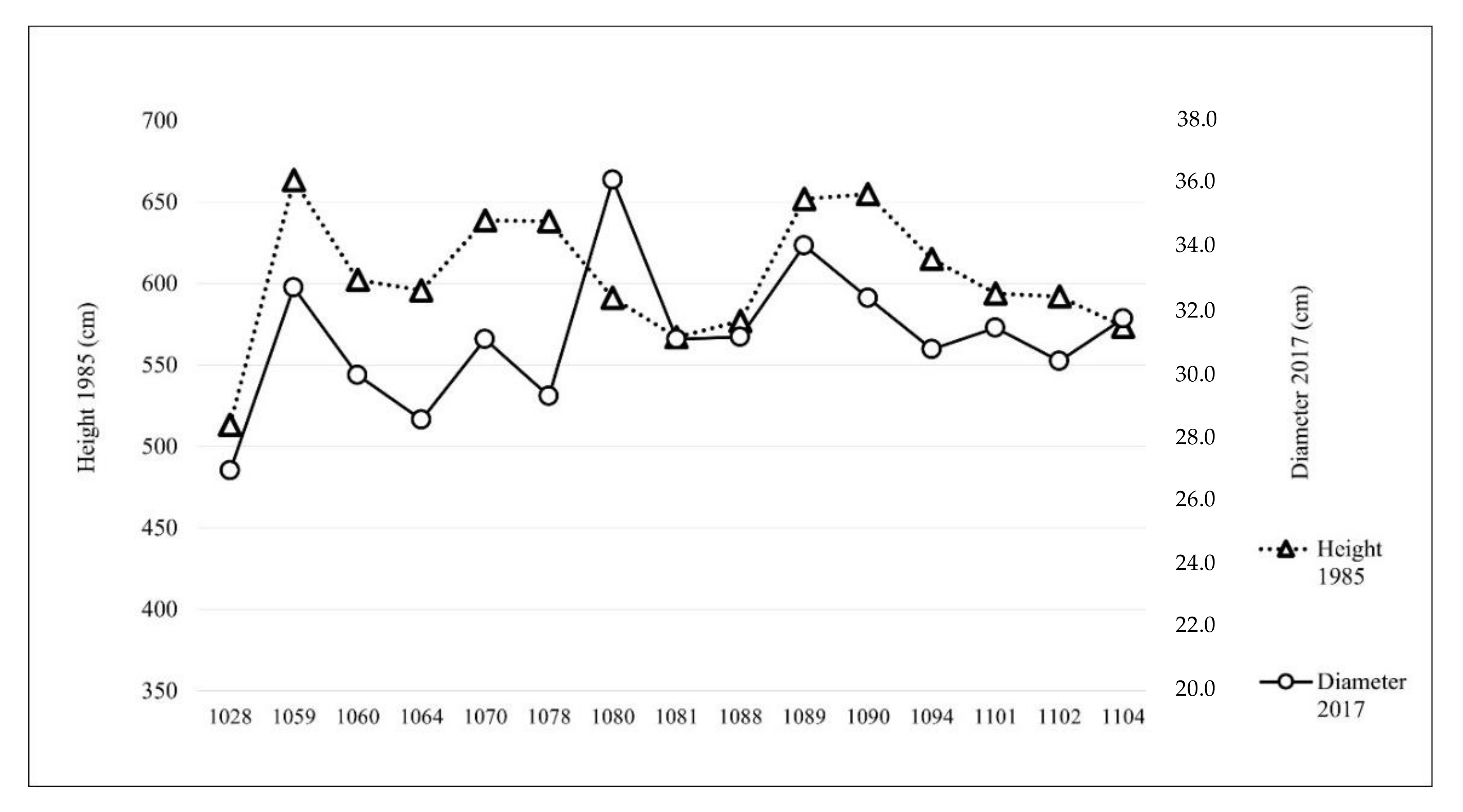
Correlation coefficients (Pearson and Spearman) rp = 0.973 and rs = 0.941 (both significant (p < 0.01)) between tree heights in 1985 and average dbh in 2017 confirmed a similar growth trend for individual provenances in the period 1985–2017. This is also illustrated in Figure 2, where provenances with the highest average tree heights in 1985 dominate with the highest average dbh in 2017. The Yelm (1080) provenance deviates from this trend; its average dbh in 2017 was above average, while in 1985 its average height was merely average.
3.3. Differences in Log Quality among Provenances
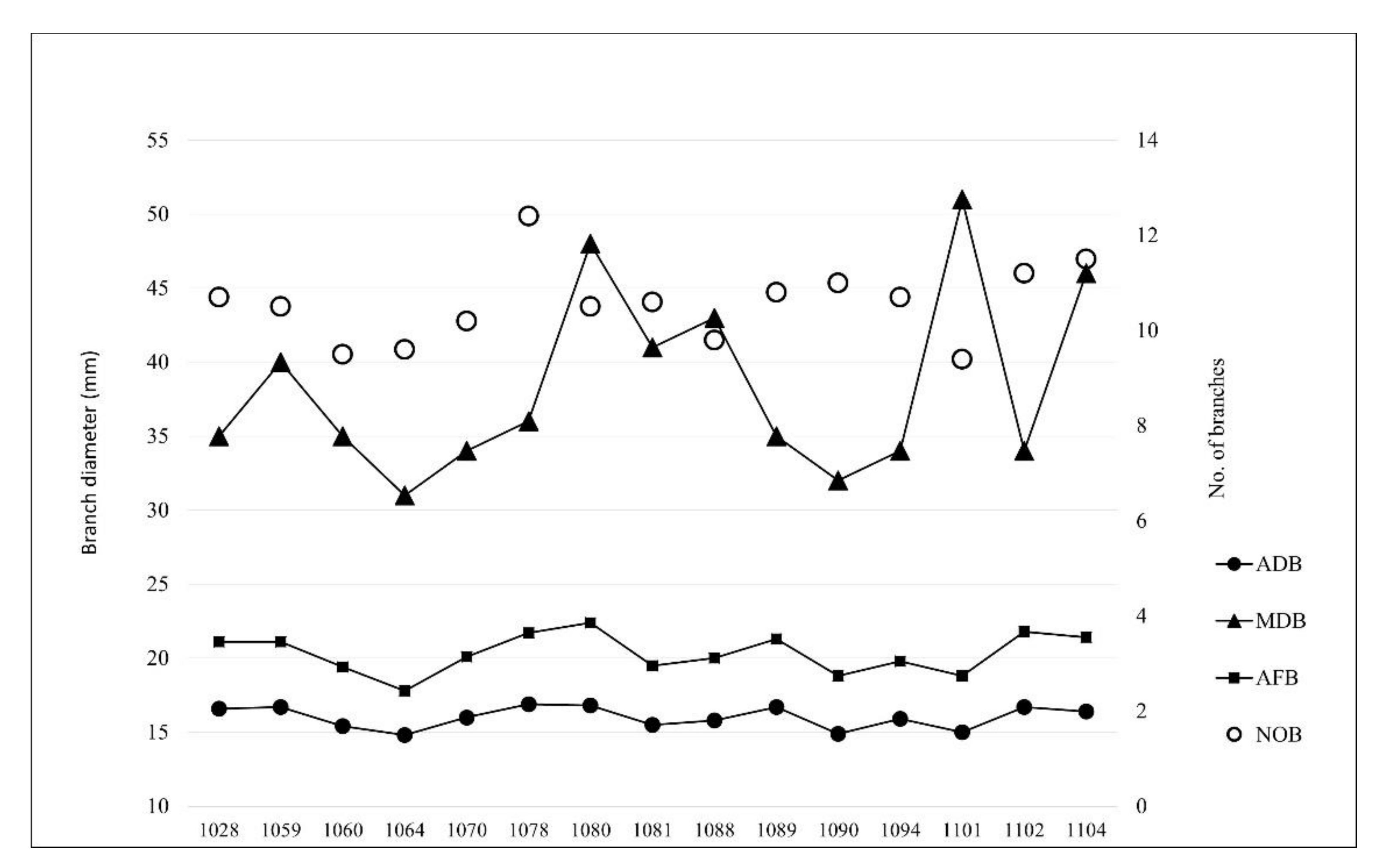
We found statistically significant differences among provenances with respect to all four measured parameters of branchiness (Table 3). The average number of branches per tree (NOB) was between 9.4 and 12.4, and the average diameter of branches (ADB) was 14.8 to 16.9 mm (Figure 3). The diameter of the thickest branch (MDB) ranged from 31 to 51 mm, while the average diameter of the thickest four branches (AFB) was 17.8–22.4 mm. Jefferson (1064) had the lowest number of branches as well as the thinnest branches, while Cle Elum (1078) had the largest number of branches. Yelm (1080) had the thickest branches accordingly (AFB in ADB) while Waldport (1101) had thickest branch (MDB) overall (Figure 3). Ramicorns appeared on 11.6% of trees on average, with Clallam (1060) having the share of ramicorns that was the highest at 24.3% (Table 2).
The analysis of covariance showed that the covariate, dbh, was significantly related to the number of the branches, F = 7.95, p < 0.01. Moreover, a positive value of b for the covariate (b = 0.041, p < 0.01) means that the number of branches increases with dbh. However, provenance also had a significant effect on the number of branches after controlling for the effect of dbh, F = 2.21, p < 0.01. Provenance and dbh also had a significant effect on the other studied traits (Table 3).
4. Discussion
Our analysis and some other foreign studies from Serbia, the Netherlands, and Bulgaria similarly confirm differences in growth among provenances [14,24,44,45,46]. In our experiment, Yelm (1080) showed the best growth performance, it had the largest average dbh, and performed well against most other provenances (Table 2). Cathlamet (1089) and Perry Creek (1059) also exhibited large average dbh.
After 46 years, the growing stock of the Douglas-fir provenance trial was 602.9 m3/ha, and the average dbh was 31.2 cm, which is comparable to other studies (e.g., 30.5 cm after 41 years) [24]. In comparable site conditions, other coniferous tree species in such stand type (coniferous plantations on silicate) have a comparable annual increment to Douglas-fir [41]. Average survival at pole stage is comparable with other studies [24].
All provenances in our provenance trial with above-average dbh originated from a lower or (in one case) similar altitude to that of the Padež I trial site (580 m a.s.l.). All provenances with above-average growth performance and superb vitality in the experimental trial originated from the coastal range of Washington (WACO region; [33]), which was previously known to be the most suitable provenance source region for plantation establishment in Europe for sites not experiencing a strong continental climate [14,24,44,45,46,47]. Guidelines for provenance selection in Germany recommend coastal subspecies (P. menziesii var. menziesii) from an altitude of up to 600 m a.s.l. [12,14,33,48]. On the other hand, coastal provenances from Oregon and Northern California are better suited to a dry climate and are thus usually recommended for drier parts of Europe, such as Apennines (Italy) [14]. Recent findings have revealed that several old Douglas-fir stands in Austria originated from areas outside the recommended regions (e.g., Northern California), indicating that additional seed sources may be suitable for Douglas-fir in Central Europe [28]. The provenance with the lowest average dbh in our trial was Merrit (1028), which originates from higher altitudes and latitudes in British Columbia; other provenances from these areas also grow poorly in Serbia [44] and Eastern Austria [49]. In contrast, in northern countries (e.g., the Netherlands), provenances from higher latitudes grow better, and growth decreases with decreasing geographical latitude of origin [24]. Over all provenance trials, breeding programs, and planned afforestation in the past, current mature Douglas-fir stands in Europe maintain high genetic diversity, which can (under certain conditions) benefit adaptive forest management under climate change [50]. It is also possible that the future suitability of certain provenances changes so that the coastal provenances from current altitudes that are currently optimal for Europe could be replaced with southern range or more drought-tolerant interior subspecies in the future [27,51,52]. While some models predict an improvement in conditions for Douglas-fir growth in the Alps, others predict a 10–36% decrease in growth on today’s optimal sites [53]. It is also important to consider the effect of climate change on Douglas-fir pathogens. Higher temperatures and moisture levels in spring could promote pathogenic fungi such as Rhabdocline pseudotsugae and Nothophaeocryptopus gaeumannii, which can infect resistant provenances, resulting in poor growth or death of a Douglas-fir tree [1].
Among the provenances themselves, and even among the descendants of a single Douglas-fir tree, there are differences in the rhythm of growth within the lifespan of a tree [54]. We compared growth trends of the individual provenances for the period from 1985 to 2017 and found a strong correlation between tree heights in 1985 and their average dbh in 2017. Similar results were obtained in the Netherlands, where most provenances remained in the same ranking after a 24-year period [24]. Similarly, juvenile growth was shown to have a positive and significant correlation with the heights [55]. A significant correlation indicates that juvenile growth could be a good predictor or indicator of future radial growth. This strong correlation suggests that it is possible to make a satisfactory selection in younger stages. Despite being a relatively long-lived species, commonly reaching an age of 750 years [48], coastal Douglas-fir exhibits rapid juvenile growth and reaches a dimension suitable for economic exploitation at a young age. Consequently, Douglas-fir trunks are full of dead and living branches, which affect log quality. Douglas-fir is generally known as a species with poor self-pruning ability, with provenance having some influence on branch characteristics [1]. Our analysis confirmed an influence of a provenance on the number and diameter of branches, which was also found in experimental plots in the species “native” range [32] and in provenance trials in the UK [33]. Regarding branch number and size, Jefferson (1064) was shown as the best, having small number and small diameter of branch, but the Yelm (1080) contrarily having the worst (Figure 3). On the other hand, thick branches and the presence of lateral shoots (ramicorns) are common issues for the most productive trees and provenances [48]. Brookings (1104) originates from the southern shores of Oregon (SOCO region) and has a poor branching habit, but the remaining coastal provenances from Washington (WACO), such as Clallam (1060), Jefferson (1064), and Cathlamet (1089), show below-average branchiness, which corresponds to other findings [33]. The number of branches primarily depends on genetics, while the diameter of branches primarily depends on planting density [32]. Spacing can be used to control branch size; at a planting density of 1000 trees per ha, branch diameters are thicker than 40 mm. A density of 1000–2000 trees per ha is recommended for smaller branch diameters [56], but even at a density of 4000 trees per ha, self-pruning never results in branch-free timber [1]. Therefore, production of high-quality logs in a short period of time (up to 80 years) is only possible with artificial pruning [1,12,56]. In our case, the trial plantation was never thinned or pruned, and the trees are heavily branched. The quality of coniferous logs depends on the straightness and shape of the trunk, but it primarily depends on the abundance and diameter of knots. According to log and product grading rules, all logs in the trial would be classified in the C and D quality classes, because of knots larger than 50 mm [30,57]. Quality is obviously very poor at this age, but we expect that quality will start to improve with increasing age. Side shoots (ramicorns) also detract from log quality, as they result in twisted fiber and undesirable trunk shapes. However, ramicorn formation depends more on ecological factors other than genetics [58], including site productivity, distance to the coast [59], and frost and pest damage [60]. In our case, ramicorns were not a decisive factor in the poor log quality grades.
5. Conclusions
In this study, we analyzed a provenance trial established within the IUFRO framework in 1970. We confirmed significant differences in growth performance among analyzed Douglas-fir provenances. In general, provenances from the WACO region exhibited better growth performance than the others. We confirmed that juvenile height is correlated to age-related radial growth. Significant differences among provenances in branching habit were also found. Despite dense planting and no thinnings, our results and field observations showed poor self-pruning at 46 years, and consequently low log quality. We were unable to identify the best provenance based on both high growth rate and superior branching habit. Under current conditions, disregarding predicted future climate change and invasive alien pest species threats, the Cathlamet provenance of Douglas-fir could be an optimal choice.
Author Contributions
Conceptualization: R.B. and K.J.; design of the work: P.S., R.B. and K.J.; the acquisition, analysis, and interpretation of data: P.S., R.B. and K.J.; original draft: P.S., R.B. and K.J.; review and editing: P.S. and R.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable (study does not involve human or animals).
Informed Consent Statement
Not applicable (study does not involve human).
Data Availability Statement
Datasets generated and/or analyzed during the current study are available from the corresponding author on request.
Acknowledgments
This article was created within the research project CRP V4-1818, which was financed by the Slovenian Research Agency and the Ministry of Agriculture, Forestry and Food. We thank to the Pahernik Foundation for the scholarship during study, and supporting the scientific work.
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A

{kind=link}
{kind=link}
{kind=link}
Table A1.
Matrix of pair comparisons of provenances average dbh with the Mann–Whitney U-test (p values). Significance levels are adjusted using sequential Bonferroni according to Rice (1989)49 (** 0.001 < p < 0.01; *** p < 0.001). See Table 1 for provenance names.
Table A1.
Matrix of pair comparisons of provenances average dbh with the Mann–Whitney U-test (p values). Significance levels are adjusted using sequential Bonferroni according to Rice (1989)49 (** 0.001 < p < 0.01; *** p < 0.001). See Table 1 for provenance names.
| Provenance | 1028 | 1059 | 1060 | 1064 | 1070 | 1078 | 1080 | 1081 | 1088 | 1089 | 1090 | 1094 | 1101 | 1102 | 1104 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1028 | 0.001 | 0.107 | 0.329 | 0.007 | 0.074 | 1.4 × 10−6 *** | 0.064 | 0.026 | 9.6 × 10−5 ** | 0.007 | 0.051 | 0.018 | 0.029 | 0.034 | |
| 1059 | 0.019 | 0.378 | 0.049 | 0.043 | 0.437 | 0.402 | 0.406 | 0.888 | 0.386 | 0.468 | 0.199 | 0.732 | |||
| 1060 | 0.509 | 0.461 | 0.78 | 0.001 | 0.539 | 0.515 | 0.025 | 0.248 | 0.665 | 0.432 | 0.801 | 0.387 | |||
| 1064 | 0.129 | 0.606 | 6.5 × 10−5 ** | 0.246 | 0.215 | 0.002 | 0.071 | 0.311 | 0.137 | 0.292 | 0.138 | ||||
| 1070 | 0.235 | 0.003 | 0.98 | 0.973 | 0.069 | 0.558 | 0.847 | 0.961 | 0.555 | 0.729 | |||||
| 1078 | 6.3 × 10−5 ** | 0.364 | 0.294 | 0.003 | 0.119 | 0.493 | 0.289 | 0.679 | 0.254 | ||||||
| 1080 | 0.026 | 0.009 | 0.273 | 0.070 | 0.016 | 0.013 | 0.001 | 0.074 | |||||||
| 1081 | 0.850 | 0.180 | 0.606 | 0.939 | 0.858 | 0.783 | 0.774 | ||||||||
| 1088 | 0.091 | 0.559 | 0.901 | 0.894 | 0.575 | 0.682 | |||||||||
| 1089 | 0.363 | 0.113 | 0.130 | 0.037 | 0.381 | ||||||||||
| 1090 | 0.496 | 0.629 | 0.305 | 0.791 | |||||||||||
| 1094 | 0.792 | 0.844 | 0.713 | ||||||||||||
| 1101 | 0.557 | 0.928 | |||||||||||||
| 1102 | 0.566 | ||||||||||||||
| 1104 |
References
- Spiecker, H.; Lindner, M.; Schuler, J.K. What Science Can Tell Us 9—Douglas-Fir: An Option for Europe; European Forest Institute: Joensuu, Finland, 2019; 121p. [Google Scholar]
- Lopatka, J. Climate Change to Blame as Bark Beetles Revage Central Europe Forests; Reuters (Prague): Nové Město, Czech Republic, 2019; Available online: https://www.reuters.com/article/us-centraleurope-environment-barkbeetle-idUSKCN1S21LA (accessed on 12 February 2021).
- Williams, D.W.; Liebhold, A.M. Climate change and the outbreak ranges of two North American bark beetles. Agric. For. Entomol. 2002, 4, 87–99. [Google Scholar] [CrossRef]
- IPCC/Working Group. The physical science basis: Summary for policymakers and technical summary and frequently asked questions; Part of the Working Group I contribution to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. In Proceedings of the Alpine Snow Workshop, Munich, Germany, 5–6 October 2007; p. 142. [Google Scholar]
- Eilmann, B.; Zweifel, R.; Buchmann, N.; Fonti, P.; Rigling, A. Drought-induced adaptation of the xylem in Scots pine and pubescent oak. Tree Physiol. 2009, 29, 1011–1020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eilmann, B.; Buchmann, N.; Siegwolf, R.; Saurer, M.; Cherubini, P.; Rigling, A. Fast response of Scots pine to improved water availability reflected in tree-ring width and δ13C. Plant Cell Environ. 2010, 33, 1351–1360. [Google Scholar] [CrossRef]
- Eilmann, B.; Zweifel, R.; Buchmann, N.; Pannatier, E.G.; Rigling, A. Drought alters timing, quantity, and quality of wood formation in Scots pine. J. Exp. Bot. 2011, 62, 2763–2771. [Google Scholar] [CrossRef] [Green Version]
- Rigling, A.; Bigler, C.; Eilmann, B.; Feldmeyer-Christe, E.; Gimmi, U.; Ginzler, C.; Graf, U.; Mayer, P.; Vacchiano, G.; Weber, P.; et al. Driving factors of a vegetation shift from Scots pine to pubescent oak in dry Alpine forests. Glob. Change Biol. 2013, 19, 229–240. [Google Scholar] [CrossRef] [PubMed]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.H.; et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef] [Green Version]
- Ince, P.J.; Kramp, A.D.; Skog, K.E.; Yoo, D.-I.; Sample, V.A. Modeling future U.S. forest sector market and trade impacts of expansion in wood energy consumption. J. For. Econ. 2011, 17, 142–156. [Google Scholar] [CrossRef]
- Bolte, A.; Ammer, C.; Löf, M.; Madsen, P.; Nabuurs, G.-J.; Schall, P.; Spathelf, P.; Rock, J. Adaptive forest management in central Europe: Climate change impacts, strategies and integrative concept. Scand. J. For. Res. 2009, 24, 473–482. [Google Scholar] [CrossRef]
- Konnert, M.; Alizoti, P.; Bastien, J.C.; Chakraborty, D.; Cvjetkovic, B.; Klisz, M.; Kroon, J.; Mason, B.; Neophytou, C.; Schueler, S.; et al. European Provenance Recommendations for Selected Non-Native Tree Species—WG2 Report; University of Natural Resources and Life Sciences: Vienna, Austria, 2018. [Google Scholar]
- Beniston, M.; Stephenson, D.B.; Christensen, O.B.; Ferro, C.A.T.; Frei, C.; Goyette, S.; Halsnaes, K.; Holt, T.; Jylhä, K.; Koffi, B.; et al. Future extreme events in European climate: An exploration of regional climate model projections. Clim. Change 2007, 81, 71–95. [Google Scholar] [CrossRef] [Green Version]
- Bastien, J.C.; Sanchez, L. Chapter 7 Douglas-Fir (Pseudotsuga menziesii (Mirb.) Franco) in Forest Tree Breeding in Europe: Current State-of-the-Art and Perspectives; Springer: Dordrecht, The Netherlands, 2013; Volume 25, pp. 325–369. [Google Scholar]
- Kleinschmit, J.; Bastien, J.C. IUFRO’s Role in Douglas-Fir (Pseudotsuga menziesii (Mirb.) Franco) Tree improvement. Silvae Genet. 1992, 41, 161–173. [Google Scholar]
- Krumm, F.; Vitkova, L. Introduced tree species in European forests: Opportunities and challenges. In Focus—Managing Forests in Europe; European Forest Institute: Joensuu, Finland, 2016. [Google Scholar]
- Curt, T.; Bouchaud, M.; Agrech, G. Predicting site index of Douglas-Fir plantations from ecological variables in the Massif Central area of France. For. Ecol. Manag. 2001, 149, 61–74. [Google Scholar] [CrossRef]
- Jasser, C. Douglasie in Oberösterreich: Möglichkeiten und Grenzen. BFW Prax. 2008, 16, 19–20. [Google Scholar]
- Eilmann, B.; Rigling, A. Tree-growth analyses to estimate tree species’ drought tolerance. Tree Physiol. 2012, 32, 178–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montwé, D.; Spiecker, H.; Hamann, A. Five decades of growth in a genetic field trial of Douglas-fir reveal trade-offs between productivity and drought tolerance. Tree Genet. Genomes 2015, 11, 1–11. [Google Scholar] [CrossRef]
- Brus, R.; Pötzelsberger, E.; Lapin, K.; Brundu, G.; Orazio, C.; Straigyte, L.; Hasenauer, H. Extent, distribution and origin of non-native forest tree species in Europe. Scand. J. For. Res. 2019, 34, 533–544. [Google Scholar] [CrossRef]
- Pulkrab, K.; Sloup, M.; Zeman, M. Economic Impact of Douglas-fir (Pseudotsuga menziesii (Mirb.) Franco) production in the Czech Republic. J. For. Sci. 2014, 60, 297–306. [Google Scholar] [CrossRef] [Green Version]
- Institut National de la Recherche Agronomique (INRA). Genetically Improved Douglas-Fir for European Forestry. 1993. Available online: https://cordis.europa.eu/project/id/MA2B900010 (accessed on 11 February 2021).
- Eilmann, B.; De Vries, S.M.; Ouden, J.D.; Mohren, G.M.; Sauren, P.; Sass-Klaassen, U. Origin matters! Difference in drought tolerance and productivity of coastal Douglas-fir (Pseudotsuga menziesii (Mirb.)) provenances. For. Ecol. Manag. 2013, 302, 133–143. [Google Scholar] [CrossRef]
- Nagamitsu, T.; Nagasaka, K.; Yoshimaru, H.; Tsumura, Y. Provenance tests for survival and growth of 50-year-old Japanese larch (Larix kaempferi) trees related to climatic conditions in central Japan. Tree Genet. Genomes 2013, 10, 87–99. [Google Scholar] [CrossRef]
- Chantre, G.; Rozenberg, P.; Baonza, V.; Macchioni, N.; Le Turcq, A.; Rueff, M.; Petit-Conil, M.; Heois, B. Genetic selection within Douglas fir (Pseudotsuga menziesii) in Europe for papermaking uses. Ann. For. Sci. 2002, 59, 583–593. [Google Scholar] [CrossRef] [Green Version]
- Isaac-Renton, M.G.; Roberts, D.R.; Hamann, A.; Spiecker, H. Douglas-fir plantations in Europe: A retrospective test of assisted migration to address climate change. Glob. Change Biol. 2014, 20, 2607–2617. [Google Scholar] [CrossRef] [PubMed]
- Hintsteiner, W.J.; Van Loo, M.; Neophytou, C.; Schueler, S.; Hasenauer, H. The geographic origin of old Douglas-fir stands growing in Central Europe. Eur. J. For. Res. 2018, 137, 447–461. [Google Scholar] [CrossRef] [Green Version]
- Généré, B. Five-year field performance of two types of Douglas fir mini-plug transplants in three forest sites in France. Ann. Sci. For. 1998, 55, 885–897. [Google Scholar] [CrossRef] [Green Version]
- Forstwirtschaftsrat, D.; Holzwirtschaftsrat, D. Rahmenvereinbarung für den Rohholzhandel in Deutschland (RVR); Forstliche Versuchs und Forschungsanstalt Baden-Württemberg (FVA): Freiburg, Germany, 2015; p. 56. [Google Scholar]
- Dvorak, W.; Kietzka, E.; Hodge, G.; Nel, A.; Dos Santos, G.; Gantz, C. Assessing the potential of Pinus herrerae as a plantation species for the subtropics. For. Ecol. Manag. 2007, 242, 598–605. [Google Scholar] [CrossRef]
- Briggs, D.; Ingaramo, L.; Turnblom, E. Number and diameter of breast-height region branches in a Douglas-fir spacing trial and linkage to log quality. For. Prod. J. 2007, 57, 28–34. [Google Scholar]
- Fletcher, M.; Samuel, C.J.A. Choice of Douglas Fir Seed Origins for Use in British Forests; Forestry Commission: Edinburgh, UK, 2010. [Google Scholar]
- Wonn, H.T.; O’Hara, K.L. Height: Diameter Ratios and Stability Relationships for Four Northern Rocky Mountain Tree Species. West J. Appl. For. 2001, 16, 87–94. [Google Scholar] [CrossRef] [Green Version]
- Hanus, M.L.; Marshall, D.D.; Hann, D.W. Height-Diameter Equations for Six Species in the Coastal Regions of the Pacific Northwest; Forest Research Laboratory, Oregon State University: Corvallis, OR, USA, 1999; 11p. [Google Scholar]
- Breznikar, A. Mednarodno Proučevanje Duglazije (Pseudotsuga menziesii (Mirb) Franco) v Sloveniji; University of Ljubljana: Ljubljana, Slovenia, 1991. [Google Scholar]
- Ogrin, D. Podnebni tipi v Sloveniji. Geogr. Vestn. 1996, 68, 39–56. [Google Scholar]
- Agencija Republike Slovenije za Okolje. Meteorološka Postaja Ilirska Bistrica. 2014. Available online: http://meteo.arso.gov.si/uploads/probase/www/climate/table/sl/by_location/ilirska-bistrica/climate-normals_81-10_Ilirska-Bistrica.pdf (accessed on 22 April 2018).
- Mlinšek, D. Eksote na Krasu, Letno Poročilo Inštituta za Gozdno in Lesno Gospodarstvo Slovenije; Inštitut za Gozdno in Lesno Gospodarstvo Slovenije: Ljubljana, Slovenia, 1977. [Google Scholar]
- Koop, H. Forest Dynamics: Silvi-Star: A Comprehensive Monitoring System; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2012. [Google Scholar]
- Zavod za Gozdove Slovenije. Gozdnogospodarski Načrt Kraškega Gozdnogospodarskega Območja (2011–2020); Zavod za Gozdove Slovenije, OE Sežana: Sežana, Slovenia, 2012.
- Kotar, M. Gozdarski Priročnik; Oddelek za Gozdarstvo in Obnovljive Gozdne Vire: Ljubljana, Slovenia, 2003. [Google Scholar]
- Rice, W.R. Analyzing tables of statistical tests. Evolution 1989, 43, 223–225. [Google Scholar] [CrossRef]
- Lavadinovic, V.; Isajev, V.; Miletic, Z.; Krstic, M. Variability of nitrogen content in the needles of Douglas-fir (Pseudotsuga menziesii Mir/Franco) provenances. Genetika 2011, 43, 407–417. [Google Scholar] [CrossRef]
- Popov, E. Results of 20 years old Douglas-fir provenance experiment established on the northern slopes of Rila Mountain in Bulgaria. J. For. Sci. 2014, 60, 394–399. [Google Scholar] [CrossRef] [Green Version]
- Petkova, K. Investigation of Douglas-Fir provenance test in NW Bulgaria at Age 20. For. Ideas 2011, 17, 131–140. [Google Scholar]
- Perić, S.; Jazbec, A.M.; Tijardović, M.; Margaletić, J.; Ivanković, M.; Pilaš, I.; Medak, J. Provenance studies of Douglas fir in the locality of »Kontija« (Istria). Period. Biol. 2009, 111, 487–493. [Google Scholar]
- Lavender, D.P.; Hermann, R.K. Douglas-Fir the Genus Pseudotsuga; OSU College of Forestry: Corvallis, OR, USA, 2014; p. 352. [Google Scholar]
- Schultze, U.; Raschka, H.D. Douglasienherkünfte für den sommerwarmen Osten Österreichs. FBVA Berichte. 2002, 126, 1–9. [Google Scholar]
- Neophytou, C.; Van Loo, M.; Hasenauer, H. Genetic diversity in introduced Douglas-fir and its natural regeneration in Central Europe. Forestry 2019, 93, 535–544. [Google Scholar] [CrossRef]
- Chauvin, T.; Cochard, H.; Segura, V.; Rozenberg, P. Native-source climate determines the Douglas-fir potential of adaptation to drought. For. Ecol. Manag. 2019, 444, 9–20. [Google Scholar] [CrossRef]
- Sergent, A.S.; Bréda, N.; Sanchez, L.; Bastein, J.C.; Rozenberg, P. Coastal and interior Douglas-fir provenances differ in growth performance and response to drought episodes at adult age. Ann. For. Sci. 2014, 71, 709–720. [Google Scholar] [CrossRef] [Green Version]
- Chakraborty, D.; Wang, T.; Andre, K.; Konnert, M.; Lexer, M.J.; Matulla, C.; Weißenbacher, L.; Schueler, S. Adapting Douglas-fir forestry in Central Europe: Evaluation, application, and uncertainty analysis of a genetically based model. Eur. J. For. Res. 2016, 135, 919–936. [Google Scholar] [CrossRef]
- Göhre, K. Douglasie und ihr Holz; Akadamie: Berlin, Germany, 1958; 595p. [Google Scholar]
- Schuler, T.M. Survival, Growth, and Juvenile-Mature Correlations in a West Virginia Sugar Maple Provenance Test 25 Years after Establishment; US Department of Agriculture, Forest Service, Northeastern Forest Experiment Station: Radnor, PA, USA, 1994; 5p. [CrossRef] [Green Version]
- Makkonen-Spiecker, K. Douglasie: Leistungsträger mit Migrationshintergrund. AFZ Wald. 2010, 23, 33–35. [Google Scholar]
- Pravilnik o Merjenju in Razvrščanju Gozdnih Lesnih Sortimentov iz Gozdov v Lasti Republike Slovenije. Uradni List RS, št. 30/2017. 2017.
- Cornelius, J. Heritabilities and additive genetic coefficients of variation in forest trees. Can. J. For. Res. 1994, 24, 372–379. [Google Scholar] [CrossRef]
- Magalska, L.; Howe, G.T. Genetic and environmental control of Douglas-fir stem defects. For. Ecol. Manag. 2014, 318, 228–238. [Google Scholar] [CrossRef]
- Cline, M.G.; Harrington, C. Apical dominance and apical control in multiple flushing of temperate woody species. Can. J. For. Res. 2007, 37, 74–83. [Google Scholar] [CrossRef]
Figure 1.
Experimental plot design with legend. The distance between individual trees and rows is 2.5 m.
Figure 1.
Experimental plot design with legend. The distance between individual trees and rows is 2.5 m.

Figure 2.
Data on average tree heights in 1985 and average diameter at breast height (dbh) in 2017. See Table 1 for provenance names.
Figure 2.
Data on average tree heights in 1985 and average diameter at breast height (dbh) in 2017. See Table 1 for provenance names.

Figure 3.
Combined graph of branch criteria, representing the mean values for the individual characteristics studied. See Table 1 for provenance names.
Figure 3.
Combined graph of branch criteria, representing the mean values for the individual characteristics studied. See Table 1 for provenance names.

Table 1.
Provenances in Padež I trial plot: IUFRO code—international provenance IUFRO code; name—provenance name (nearby city); state—federal state; N (°) and W (°)—geographical coordinates; altitude (m)—altitude in meters above sea level; num. seed—total number of planted seedlings per provenance; num. series—number of repetitions [24,27,36].
Table 1.
Provenances in Padež I trial plot: IUFRO code—international provenance IUFRO code; name—provenance name (nearby city); state—federal state; N (°) and W (°)—geographical coordinates; altitude (m)—altitude in meters above sea level; num. seed—total number of planted seedlings per provenance; num. series—number of repetitions [24,27,36].
| IUFRO Code | Name | State | N (°) | W (°) | Altitude (m) | Num. Seed | Num. Series |
|---|---|---|---|---|---|---|---|
| 1028 | Merrit | Brit. Kolumbija (BC) | 50.07 | 120.85 | 870–950 | 180 | 18 |
| 1059 | Perry creek | Washington (WA) | 48.05 | 121.47 | 600–700 | 197 | 20 |
| 1060 | Clallam, Sequim | Washington (WA) | 48.03 | 123.03 | 60–90 | 159 | 16 |
| 1064 | Jefferson, Hoh River | Washington (WA) | 47.80 | 123.97 | 240–245 | 166 | 17 |
| 1070 | Denny creek | Washington (WA) | 47.40 | 121.53 | 540–550 | 170 | 17 |
| 1078 | Cle Elum | Washington (WA) | 47.22 | 121.12 | 630–700 | 179 | 18 |
| 1080 | Thurston, Yelm | Washington (WA) | 47.02 | 122.73 | 60 | 170 | 17 |
| 1081 | Alder Lake | Washington (WA) | 46.80 | 122.28 | 420–430 | 161 | 16 |
| 1088 | Cowlitz, Castle Rock | Washington (WA) | 46.32 | 122.87 | 150 | 160 | 16 |
| 1089 | Wahkiakum, Cathlamet | Washington (WA) | 46.30 | 123.27 | 195–200 | 160 | 16 |
| 1090 | Cougar | Washington (WA) | 46.08 | 122.30 | 500–550 | 139 | 14 |
| 1094 | Washington, Vernonia | Oregon (OR) | 45.77 | 123.22 | 210–215 | 110 | 11 |
| 1101 | Waldport | Oregon (OR) | 44.40 | 123.87 | 60–90 | 170 | 17 |
| 1102 | Upper Soda | Oregon (OR) | 44.38 | 122.20 | 980–3250 | 179 | 17 |
| 1104 | Brookings | Oregon (OR) | 42.12 | 124.20 | 300–365 | 160 | 16 |
Table 2.
Performance of 15 Douglas-fir provenances originating from the west coast of North America (British Columbia (BC), Washington (WA), and Oregon (OR)) and planted in the Slovenian provenance trial. Measurements on survival, ramicorns, vitality, and diameters were conducted on all trees per provenance. * Arithmetic means from ordinal estimates were calculated only for the sake of easier representation and comparison.
Table 2.
Performance of 15 Douglas-fir provenances originating from the west coast of North America (British Columbia (BC), Washington (WA), and Oregon (OR)) and planted in the Slovenian provenance trial. Measurements on survival, ramicorns, vitality, and diameters were conducted on all trees per provenance. * Arithmetic means from ordinal estimates were calculated only for the sake of easier representation and comparison.
| Provenance (IUFRO-Code) | Survival (%) 1985 | Survival (%) 2017 | Vitality 2017 | Height (cm) 1985 | Avg. dbh (cm) 2017 | SD (dbh) | % of Trees with Ramicorns 2017 | |
|---|---|---|---|---|---|---|---|---|
| BC | 1028 | 71.7 | 35.0 | 1.87 | 513.4 | 27.0 | 9.3 | 15.9 |
| WA | 1059 | 82.2 | 51.3 | 2.30 | 663.3 | 32.7 | 11.0 | 8.9 |
| 1060 | 76.1 | 46.5 | 2.20 | 602.1 | 30.0 | 10.9 | 24.3 | |
| 1064 | 67.5 | 44.6 | 2.16 | 595.8 | 28.6 | 9.8 | 13.5 | |
| 1070 | 83.5 | 55.9 | 2.25 | 638.8 | 31.1 | 9.7 | 12.6 | |
| 1078 | 88.3 | 53.6 | 2.04 | 638.3 | 29.3 | 8.8 | 7.3 | |
| 1080 | 77.6 | 40.6 | 2.49 | 591.3 | 36.1 | 10.5 | 14.5 | |
| 1081 | 61.5 | 31.1 | 2.18 | 567.0 | 31.1 | 11.6 | 12.0 | |
| 1088 | 71.3 | 41.3 | 2.23 | 576.9 | 31.2 | 10.7 | 9.1 | |
| 1089 | 75.6 | 47.5 | 2.37 | 651.9 | 34.1 | 10.6 | 9.2 | |
| 1090 | 70.5 | 44.6 | 2.24 | 654.7 | 32.4 | 11.9 | 6.5 | |
| OR | 1094 | 57.3 | 39.1 | 2.19 | 615.1 | 30.8 | 10.3 | 2.3 |
| 1101 | 64.1 | 41.8 | 2.26 | 594.0 | 31.5 | 11.0 | 18.3 | |
| 1102 | 68.2 | 40.2 | 2.07 | 592.0 | 30.4 | 9.3 | 6.9 | |
| 1104 | 48.1 | 30.6 | 2.25 | 573.6 | 31.7 | 12.7 | 12.2 | |
| average * | 70.9 | 43.0 | 2.21 | 604.5 | 31.2 | 10.4 | 11.6 |
Table 3.
Results of the ANCOVA test. F-values and significance level are shown for provenance as a main factor and dbh as a covariate (b-value and its significance refers to the covariate).
Table 3.
Results of the ANCOVA test. F-values and significance level are shown for provenance as a main factor and dbh as a covariate (b-value and its significance refers to the covariate).
| Provenance | sig. | Covariate dbh | sig. | b | sig. | |
|---|---|---|---|---|---|---|
| Number of branches (NOB) | F = 2.21 | 0.007 | F = 7.95 | 0.005 | 0.041 | 0.005 |
| Average diameter of branches (ADB) | F = 2.27 | 0.005 | F = 159.76 | 0.000 | 0.160 | 0.000 |
| Maximum diameter of branch (MDB) | F = 1.85 | 0.030 | F = 164.60 | 0.000 | 0.305 | 0.000 |
| Average diameter of 4 thickest branches (AFB) | F = 2.63 | 0.001 | F = 177.50 | 0.000 | 0.257 | 0.000 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Smolnikar, P.; Brus, R.; Jarni, K. Differences in Growth and Log Quality of Douglas-Fir (Pseudotsuga menziesii (Mirb.) Franco) Provenances. Forests 2021, 12, 287. https://doi.org/10.3390/f12030287
AMA Style
Smolnikar P, Brus R, Jarni K. Differences in Growth and Log Quality of Douglas-Fir (Pseudotsuga menziesii (Mirb.) Franco) Provenances. Forests. 2021; 12(3):287. https://doi.org/10.3390/f12030287
Chicago/Turabian StyleSmolnikar, Peter, Robert Brus, and Kristjan Jarni. 2021. "Differences in Growth and Log Quality of Douglas-Fir (Pseudotsuga menziesii (Mirb.) Franco) Provenances" Forests 12, no. 3: 287. https://doi.org/10.3390/f12030287
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.