
Drought and Pathogen Effects on Survival, Leaf Physiology, Oxidative Damage, and Defense in Two Middle Eastern Oak Species

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growth Conditions
2.2. Fungal Isolates and Artificial Inoculation
2.3. Experimental Design and Greenhouse Experiment
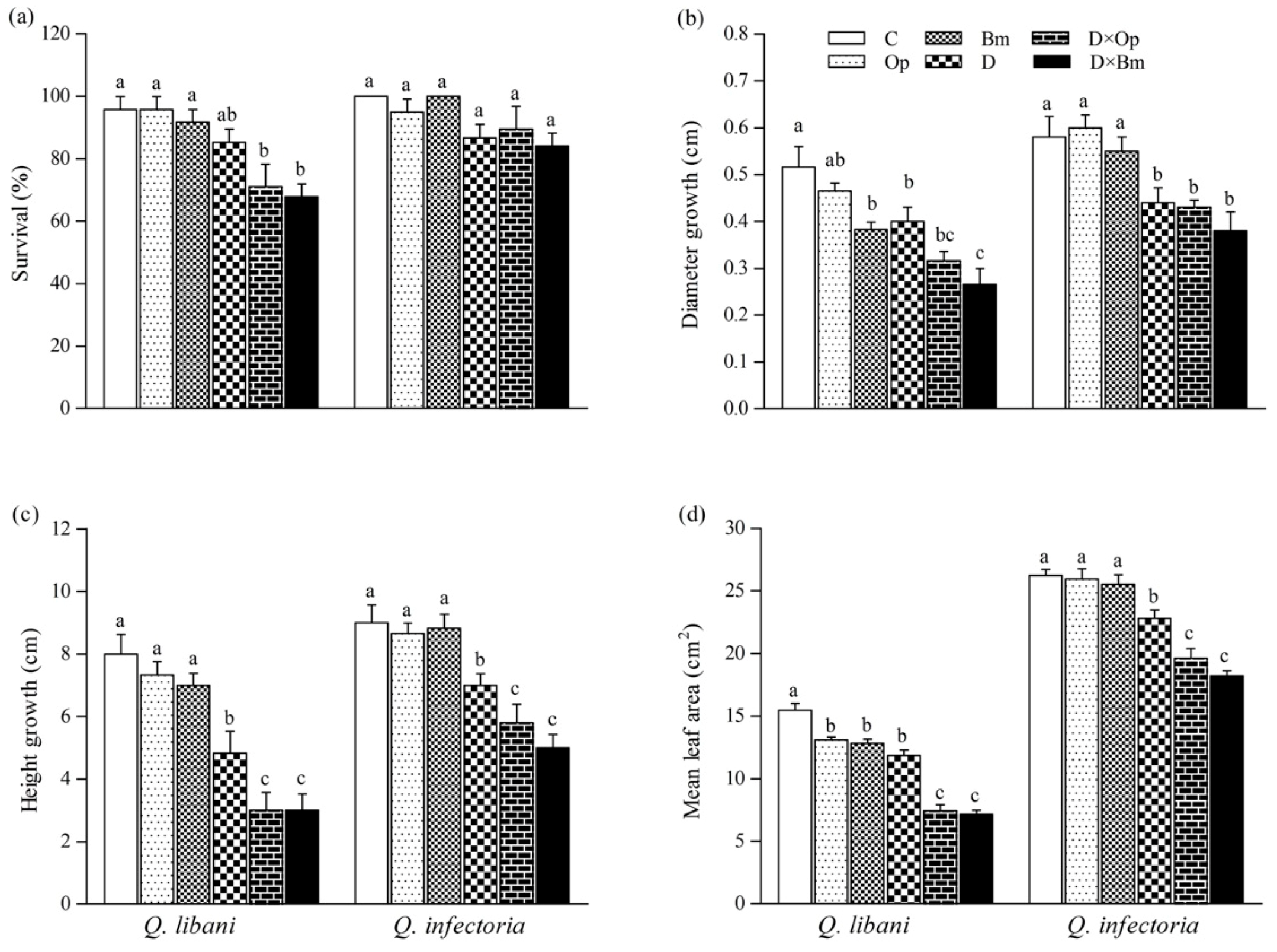
2.4. Survival and Plant Growth
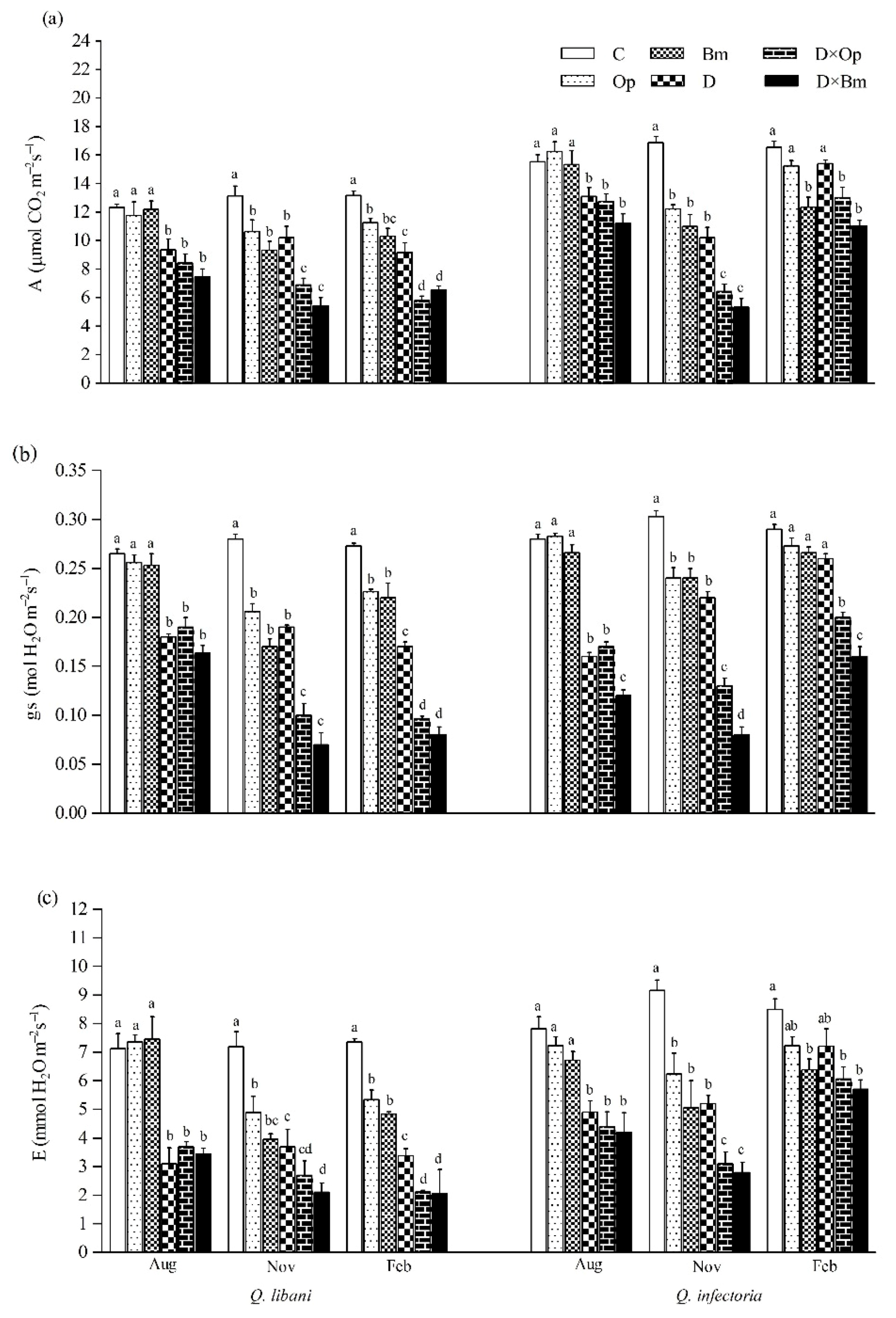
2.5. Gas-Exchange, RWC, and WUEi
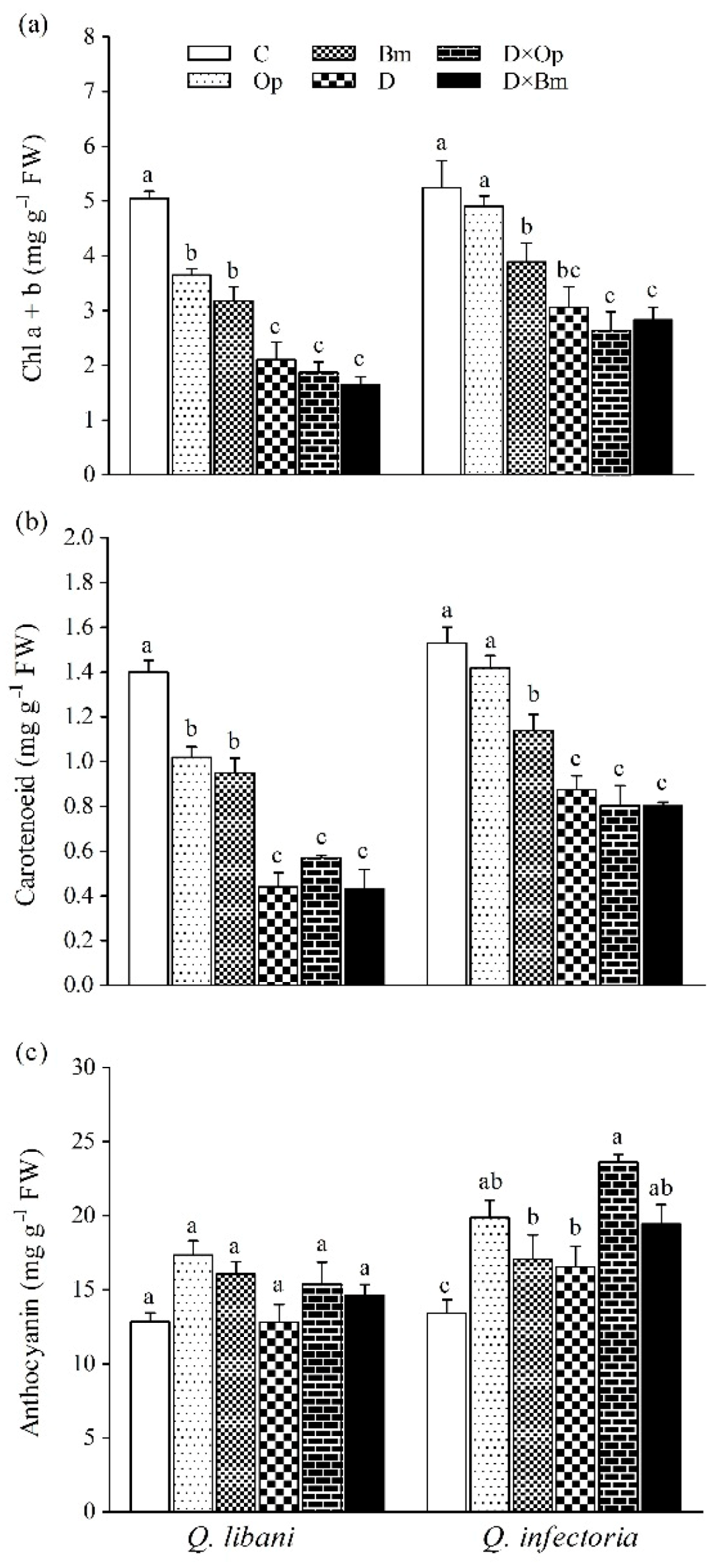
2.6. Pigment Content
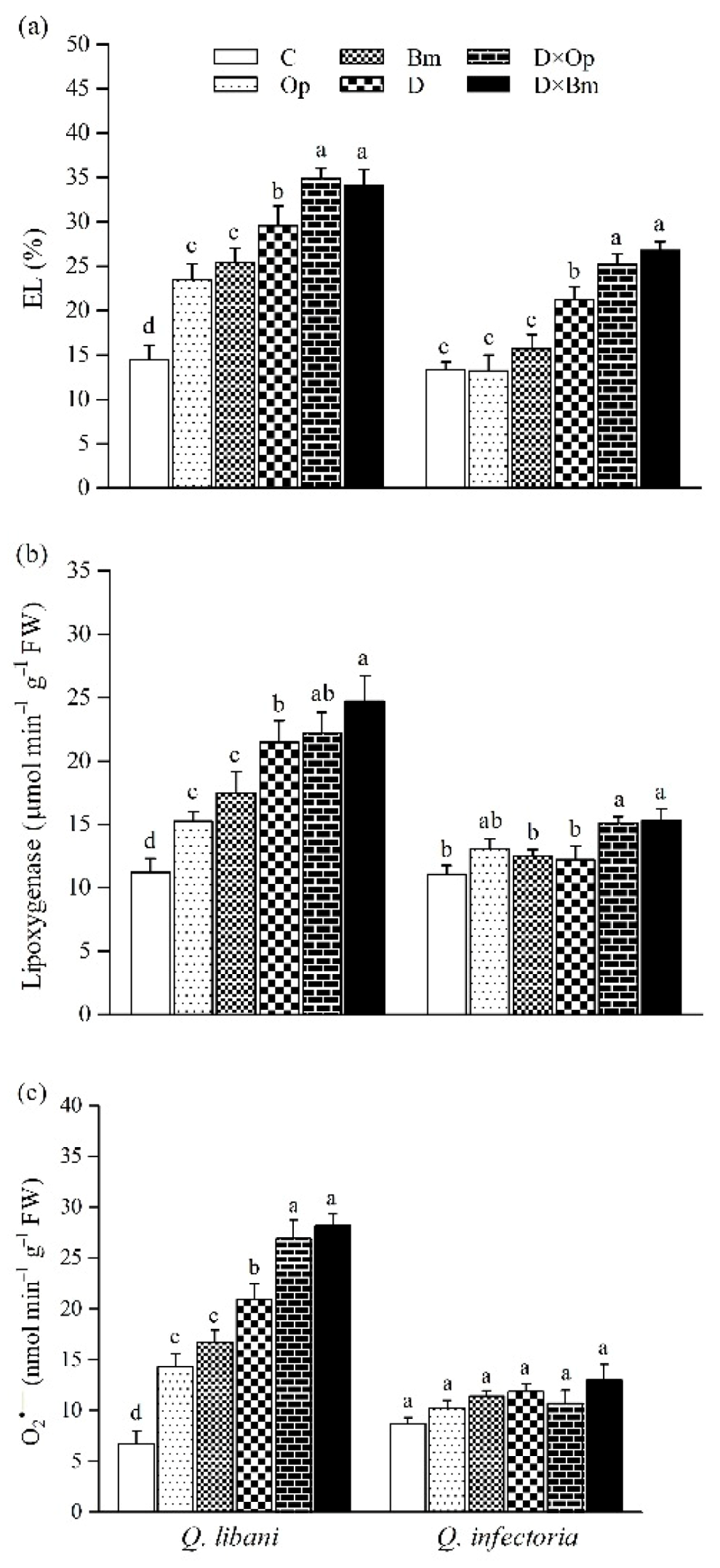
2.7. Electrolyte Leakage, LOX, and ROS Determination
2.8. Soluble Protein and Non-Enzymatic Antioxidants Content
2.9. Statistical Analysis
3. Results
3.1. Visible Symptoms
3.2. Survival and Plant Growth
3.3. Gas-Exchange, RWC, and WUEi
3.4. Pigment Content
3.5. Electrolyte Leakage, LOX, and ROS Determination
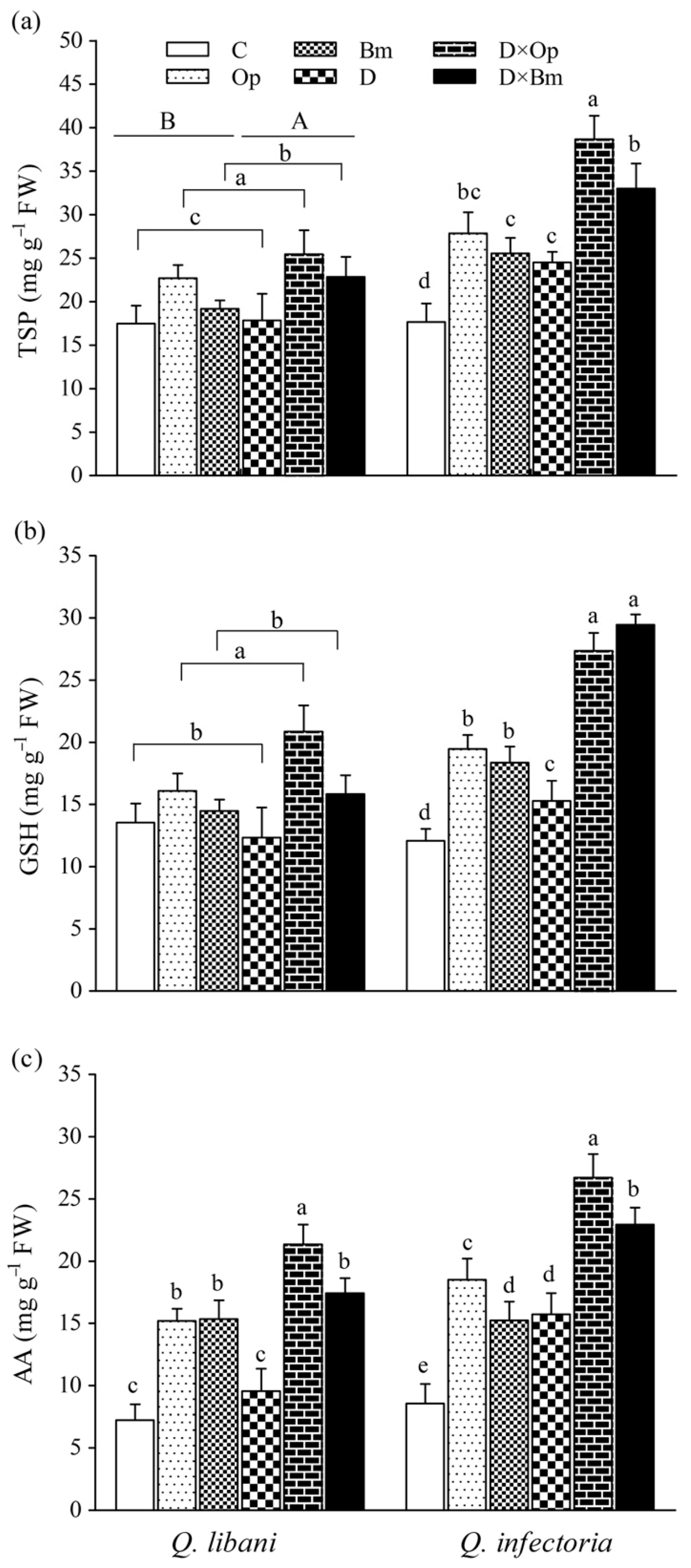
3.6. Soluble Protein and Non-Enzymatic Antioxidants Content
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Holopainen, J.K.; Gershenzon, J. Multiple stress factors and the emission of plant VOCs. Trends Plant Sci. 2010, 15, 176–184. [Google Scholar] [CrossRef]
- Niinemets, Ü. Responses of forest trees to single and multiple environmental stresses from seedlings to mature plants: Past stress history, stress interactions, tolerance and acclimation. For. Ecol. Manag. 2010, 260, 1623–1639. [Google Scholar] [CrossRef]
- Ramegowda, V.; Senthil-Kumar, M. The interactive effects of simultaneous biotic and abiotic stresses on plants: Mechanistic understanding from drought and pathogen combination. J. Plant Physiol. 2015, 176, 47–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Copolovici, L.; Väärtnõu, F.; Estrada, M.P.; Niinemets, Ü. Oak powdery mildew (Erysiphe alphitoides)-induced volatile emissions scale with the degree of infection in Quercus robur. Tree Physiol. 2014, 34, 1399–1410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dikilitas, M.; Karakas, S.; Hashem, A.; Allah, E.A.; Ahmad, P. Oxidative stress and plant responses to pathogens under drought conditions. In Water Stress and Crop Plants; Wiley: Hoboken, NJ, USA, 2016; pp. 102–123. [Google Scholar]
- Vannini, A.; Lucero, G.; Anselmi, N.; Vettraino, A. Response of endophytic Biscogniauxia mediterranea to variation in leaf water potential of Quercus cerris. For. Pathol. 2009, 39, 8–14. [Google Scholar] [CrossRef]
- Cobb, R.C.; Ruthrof, K.X.; Breshears, D.D.; Lloret, F.; Aakala, T.; Adams, H.D.; Anderegg, W.R.L.; Ewers, B.E.; Galiano, L.; Grünzweig, J.M.; et al. Ecosystem dynamics and management after forest die-off: A global synthesis with conceptual state-and-transition models. Ecosphere 2017, 8, e02034. [Google Scholar] [CrossRef]
- Wood, J.D.; Knapp, B.O.; Muzika, R.-M.; Stambaugh, M.C.; Gu, L. The importance of drought–pathogen interactions in driving oak mortality events in the Ozark Border Region. Environ. Res. Lett. 2018, 13, 015004. [Google Scholar] [CrossRef]
- Freeman, A.J.; Hammond, W.M.; Dee, J.R.; Cobb, R.C.; Marek, S.M.; Adams, H.D. The effect of prescribed fire on Biscogniauxia infection and δ13C in an upland oak-pine forest. Forest Ecol. Manag. 2019, 451, 117525. [Google Scholar] [CrossRef]
- Mirabolfathy, M.; Ju, Y.-M.; Hsieh, H.-M.; Rogers, J.D. Obolarina persica sp. nov., associated with dying Quercus in Iran. Mycoscience 2013, 54, 315–320. [Google Scholar] [CrossRef]
- Safaee, D.; Khodaparast, S.A.; Mirabolfathy, M.; Mousanejad, S. A multiplex PCR-based technique for identification of Biscogniauxia mediterranea and Obolarina persica causing charcoal disease of oak trees in Zagros forests. For. Pathol. 2017, 47, e12330. [Google Scholar] [CrossRef]
- Capretti, P.; Battisti, A. Water stress and insect defoliation promote the colonization of Quercus cerris by the fungus Biscogniauxia mediterranea. For. Pathol. 2007, 37, 129–135. [Google Scholar] [CrossRef]
- Olson, J. Biscogniauxia (Hypoxylon) canker and dieback of trees. In Oklahoma Cooperative Ex-Tenison Service EPP-7620; O.S. University: Stillwater, OK, USA, 2013. [Google Scholar]
- Alidadi, A.; Kowsari, M.; Javan-Nikkhah, M.; Salehi Jouzani, G.R.; Ebrahimi Rastaghi, M. New pathogenic and endophytic fungal species associated with Persian oak in Iran. Eur. J. Plant Pathol. 2019, 155, 1017–1032. [Google Scholar] [CrossRef]
- Mazzaglia, A.; Anselmi, N.; Gasbarri, A.; Vannini, A. Development of a Polymerase Chain Reaction (PCR) assay for the specific detection of Biscogniauxia mediterranea living as an endophyte in oak tissues. Mycol. Res. 2001, 105, 952–956. [Google Scholar] [CrossRef]
- Collado, J.; Platas, G.; Pelaez, F. Identification of an endophytic Nodulisporium sp. from Quercus ilex in central Spain as the anamorph of Biscogniauxia mediterranea by rDNA sequence analysis and effect of different ecological factors on distribution of the fungus. Mycologia 2001, 93, 875–886. [Google Scholar] [CrossRef]
- Linaldeddu, B.T.; Sirca, C.; Spano, D.; Franceschini, A. Variation of endophytic cork oak-associated fungal communities in relation to plant health and water stress. For. Pathol. 2010, 41, 193–201. [Google Scholar] [CrossRef] [Green Version]
- Guo, R.; Wang, Z.; Zhou, C.; Huang, Y.; Haijuan, F.; Wang, Y.; Zhihua, L. Biocontrol potential of Trichoderma asperellum mutants T39 and T45 and their growth promotion of poplar seedlings. J. For. Res. 2020, 31, 1035–1043. [Google Scholar] [CrossRef]
- Safaee, D.; Khodaparast, S.A.; Mirabolfathy, M.; Sheikholeslami, M. Some aspects of biology and host range of Biscogniauxia mediterranea, one of the causal agent of oak charcoal disease. Mycol. Iran. 2017, 4, 121–129. [Google Scholar]
- Nouri, A.; Kiani, B.; Hakimi, M.H.; Mokhtari, H.M. Estimating oak forest parameters in the western mountains of Iran using satellite-based vegetation indices. J. For. Res. 2020, 31, 541–552. [Google Scholar] [CrossRef]
- Bucci, S.J.; Goldstein, G.; Scholz, F.G.; Meinzer, F.C. Physiological Significance of Hydraulic Segmentation, Nocturnal Transpiration and Capacitance in Tropical Trees: Paradigms Revisited. Tree Physiol. 2016, 6, 205–225. [Google Scholar] [CrossRef]
- Hossain, M.; Veneklaas, E.J.; Hardy, G.E.S.J.; Poot, P. Tree host–pathogen interactions as influenced by drought timing: Linking physiological performance, biochemical defense and disease severity. Tree Physiol. 2019, 39, 6–18. [Google Scholar] [CrossRef]
- Ghanbary, E.; Tabari Kouchaksaraei, M.; Mirabolfathy, M.; Modarres Sanavi, S.A.M.; Rahaei, M. Growth and physiological responses of Quercus brantii seedlings inoculated with Biscogniauxia mediterranea and Obolarina persica under drought stress. For. Pathol. 2017, 47, e12353. [Google Scholar] [CrossRef]
- Hajji, M.; Dreyer, E.; Marçais, B. Impact of Erysiphe alphitoides on transpiration and photosynthesis in Quercus robur leaves. Eur. J. Plant Pathol. 2009, 125, 63–72. [Google Scholar] [CrossRef] [Green Version]
- Karakaya, A.; Dikilitas, M. Biochemical, Physiological and molecular defense mechanisms of tea plants against pathogenic agents under changing climate conditions. In Stress Physiology of Tea in the Face of Climate Change; Springer: Singapore, 2018; pp. 241–268. [Google Scholar]
- Dinis, L.-T.; Peixoto, F.; Zhang, C.; Martins, L.; Costa, R.L.; Gomes-Laranjo, J. Physiological and biochemical changes in resistant and sensitive chestnut (Castanea) plantlets after inoculation with Phytophthora cinnamomi. Physiol. Mol. Plant Pathol. 2011, 75, 146–156. [Google Scholar] [CrossRef]
- Vasques, A.R.; Pinto, G.; Dias, M.C.; Correia, C.M.; Moutinho-Pereira, J.M.; Vallejo, V.R.; Santos, C.; Keizer, J.J. Physiological response to drought in seedlings of Pistacia lentiscus (mastic tree). New For. 2016, 47, 119–130. [Google Scholar] [CrossRef]
- Senthilkumar, P.; Thirugnanasambantham, K.; Mandal, A.K.A. Suppressive Subtractive Hybridization Approach Revealed Differential Expression of Hypersensitive Response and Reactive Oxygen Species Production Genes in Tea (Camellia sinensis (L.) O. Kuntze) Leaves during Pestalotiopsis thea Infection. Appl. Biochem. Biotechnol. 2012, 168, 1917–1927. [Google Scholar] [CrossRef] [PubMed]
- Smirnoff, N. The role of active oxygen in the response of plants to water deficit and desiccation. New Phytol. 1993, 125, 27–58. [Google Scholar] [CrossRef]
- Shukla, N.; Awasthi, R.; Rawat, L.; Kumar, J. Biochemical and physiological responses of rice (Oryza sativa L.) as influenced by Trichoderma harzianum under drought stress. Plant Physiol. Biochem. 2012, 54, 78–88. [Google Scholar] [CrossRef]
- Allen, C.D.; Breshears, D.D.; McDowell, N.G. On underestimation of global vulnerability to tree mortality and forest die-off from hotter drought in the Anthropocene. Ecosphere 2015, 6, art129. [Google Scholar] [CrossRef]
- Garrett, K.A.; Dendy, S.P.; Frank, E.E.; Rouse, M.N.; Travers, S.E. Climate Change Effects on Plant Disease: Genomes to Ecosystems. Annu. Rev. Phytopathol. 2006, 44, 489–509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hubbart, J.A.; Guyette, R.; Muzika, R.-M. More than Drought: Precipitation Variance, Excessive Wetness, Pathogens and the Future of the Western Edge of the Eastern Deciduous Forest. Sci. Total Environ. 2016, 566, 463–467. [Google Scholar] [CrossRef] [Green Version]
- Ghanbary, E.; Kouchaksaraei, M.T.; Guidi, L.; Mirabolfathy, M.; Etemad, V.; Sanavi, S.A.M.M.; Struve, D. Change in biochemical parameters of Persian oak (Quercus brantii Lindl.) seedlings inoculated by pathogens of charcoal disease under water deficit conditions. Trees 2018, 32, 1595–1608. [Google Scholar] [CrossRef]
- Ghanbary, E.; Tabari Kouchaksaraei, M.; Zarafshar, M.; Bader, M.K.-F.; Mirabolfathy, M.; Ziaei, M. Differential physiological and biochemical responses of Quercus infectoria and Q. libani to drought and charcoal disease. Physiol. Plant. 2020, 168, 876–892. [Google Scholar] [CrossRef]
- Linaldeddu, B.T.; Sirca, C.; Spano, D.; Franceschini, A. Physiological responses of cork oak and holm oak to infection by fungal pathogens involved in oak decline. For. Pathol. 2009, 39, 232–238. [Google Scholar] [CrossRef]
- Oguchi, R.; Hikosaka, K.; Hiura, T.; Hirose, T. Leaf anatomy and light acclimation in woody seedlings after gap formation in a cool-temperate deciduous forest. Oecologia 2006, 149, 571–582. [Google Scholar] [CrossRef]
- Baltzer, J.L.; Thomas, S.C. Determinants of whole-plant light requirements in Bornean rain forest tree saplings. J. Ecol. 2007, 95, 1208–1221. [Google Scholar] [CrossRef]
- Parad, G.A.; Tabari Kouchaksaraei, M.; Striker, G.G.; Sadati, S.E.; Nourmohammadi, K. Growth, morphology and gas exchange responses of two-year-old Quercus castaneifolia seedlings to flooding stress. Scand. J. For. Res. 2015, 35, 458–466. [Google Scholar]
- Almeida, T.; Pinto, G.; Correia, B.; Santos, C.; Gonçalves, S. QsMYB1 expression is modulated in response to heat and drought stresses and during plant recovery in Quercus suber. Plant Physiol. Biochem. 2013, 73, 274–281. [Google Scholar] [CrossRef] [PubMed]
- Arnon, D.I. Copper Enzymes in Isolated Chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wagner, G.J. Content and Vacuole/Extravacuole Distribution of Neutral Sugars, Free Amino Acids, and Anthocyanin in Protoplasts. Plant Physiol. 1979, 64, 88–93. [Google Scholar] [CrossRef] [Green Version]
- Nayyar, H. Accumulation of osmolytes and osmotic adjustment in water-stressed wheat (Triticum aestivum) and maize (Zea mays) as affected by calcium and its antagonists. Environ. Exp. Bot. 2003, 50, 253–264. [Google Scholar] [CrossRef]
- Axelrod, B.; Cheesbrough, T.M.; Laakso, S. Lipoxygenases from soybeans. In Methods in Enzymology; Lowenstein, J.M., Ed.; Academic Press: New York, NY, USA, 1981; pp. 441–451. [Google Scholar]
- Bai, T.; Li, C.; Ma, F.; Feng, F.; Shu, H. Responses of growth and antioxidant system to root-zone hypoxia stress in two Malus species. Plant Soil 2009, 327, 95–105. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- De Vos, C.H.R.; Vonk, M.J.; Vooijs, R.; Schat, H. Glutathione Depletion Due to Copper-Induced Phytochelatin Synthesis Causes Oxidative Stress in Silene cucubalus. Plant Physiol. 1992, 98, 853–858. [Google Scholar] [CrossRef] [Green Version]
- Singh, N.; Lena, Q. India (host institution) Lucknow 226001 Rana Pratap Marg National Botanical Research Institute; Srivastava, M.; Rathinasabapathi, B. Metabolic adaptations to arsenic-induced oxidative stress in Pteris vittata L. and Pteris ensiformis L. Plant Sci. 2006, 170, 274–282. [Google Scholar] [CrossRef]
- Nowakowska, J.A.; Stocki, M.; Stocka, N.; Ślusarski, S.; Tkaczyk, M.; Caetano, J.M.; Tulik, M.; Hsiang, T.; Oszako, T. Interactions between Phytophthora cactorum, Armillaria gallica and Betula pendula Roth. Seedlings Subjected to Defoliation. Forests 2020, 11, 1107. [Google Scholar] [CrossRef]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.H.; et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef] [Green Version]
- Atkinson, N.J.; Urwin, P.E. The interaction of plant biotic and abiotic stresses: From genes to the field. J. Exp. Bot. 2012, 63, 3523–3543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bilgin, D.D.; Zavala, J.A.; Zhu, J.; Clough, S.L.; Ort, D.R.; De Lucia, E.H. Biotic stress globally down regulates photosynthesis genes. Plant Cell Environ. 2010, 33, 1597–1613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogaya, R.; Penuelas, J. Comparative field study of Quercus ilex and Phillyrea latifolia: Photosynthetic response to experimental drought conditions. Environ. Exp. Bot. 2003, 50, 137–148. [Google Scholar] [CrossRef]
- Corcobado, T.; Cubera, E.; Juárez, E.; Moreno, G.; Solla, A. Drought events determine performance of Quercus ilex seedlings and increase their susceptibility to Phytophthora cinnamomi. Agric. For. Meteorol. 2014, 192, 1–8. [Google Scholar] [CrossRef]
- Oszako, T.; Orlikowski, L.B. First data on the occurrence of Phytophthora cinnamomi on pedonculate oak in Poland. Sylwan 2005, 149, 47–53. [Google Scholar]
- Oszako, T.; Voitka, D.; Stocki, M.; Stocka, N.; Nowakowska, J.A.; Linkiewicz, A.; Hsiang, T.; Belbahri, L.; Berezovska, D.; Malewski, T. Trichoderma asperellum efficiently protects Quercus robur leaves against Erysiphe alphitoides. Eur. J. Plant Pathol. 2021, 159, 295–308. [Google Scholar] [CrossRef]
- Anderson, L.J.; Harley, P.C.; Monson, R.K.; Jackson, R.B. Reduction of isoprene emissions from live oak (Quercus fusiformis) with oak wilt. Tree Physiol. 2000, 20, 1199–1203. [Google Scholar] [CrossRef] [Green Version]
- Luque, J.; Cohen, M.; Savé, R.; Biel, C.; Álvarez, I.F. Effects of three fungal pathogens on water relations, chlorophyll fluorescence and growth of Quercus suber L. Ann. Sci. For. 1999, 56, 19–26. [Google Scholar] [CrossRef] [Green Version]
- Valladares, F.; Sanchez-Gomez, D. Ecophysiological traits associated with drought in Mediterranean tree seedlings: Individual responses versus interspecific trends in eleven species. Plant Biol. 2006, 8, 688–697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gómez-Gallego, M.; Bader, M.K.-F.; Scott, P.M.; Leuzinger, S.; Williams, N.M. Phytophthora pluvialis Studies on Douglas-fir Require Swiss Needle Cast Suppression. Plant Dis. 2017, 101, 1259–1262. [Google Scholar] [CrossRef] [Green Version]
- Kalmatskaya, O.A.; Karavaev, V.A. The fluorescent indices of bean leaves treated with sodium fluoride. Biophysics 2015, 60, 843–848. [Google Scholar] [CrossRef]
- Ranjbar, A. Comparative study on the effect of water stress and rootstock on photosynthetic function in pistachio (Pistacia vera L). Trees 2017, 8, 151–159. [Google Scholar]
- Sun, J.; Zhang, Q.; Tabassum, M.A.; Ye, M.; Peng, S.; Li, Y. The inhibition of photosynthesis under water deficit conditions is more severe in flecked than uniform irradiance in rice (Oryza sativa) plants. Funct. Plant Biol. 2017, 44, 464–472. [Google Scholar] [CrossRef] [PubMed]
- Evidente, A.; Andolfi, A.; Maddau, L.; Franceschini, A.; Marras, F. Biscopyran, a phytotoxic hexasubstituted pyranopyran produced by Biscogniauxia mediterranea, a fungus pathogen of cork oak. J. Nat. Prod. 2005, 68, 568–571. [Google Scholar] [CrossRef] [PubMed]
- Farouk, S.; Abdul Qados, A.M.S. Osmotic adjustment and yield of cowpea in response to drought stress and chitosan. Indian J. Appl. Res. 2013, 3, 1–6. [Google Scholar] [CrossRef]
- Fernandez-Martinez, J.; Zacchini, M.; Elena, G.; Fernandez-Marin, B.; Fleck, I. Effect of environmental stress factors on ecophysiological traits and susceptibility to pathogens of five Populus clones throughout the growing season. Tree Physiol. 2013, 33, 618–627. [Google Scholar] [CrossRef] [Green Version]
- González, L.; González-Vilar, M. Determination of relative water content. In Handbook of Plant Eco-Physiology Techniques; Reigosa, M.J., Ed.; Springer: Dordrecht, The Netherlands, 2001; pp. 207–212. [Google Scholar]
- Hutchinson, T.C. Lime Chlorosis as a Factor in Seedling Establishment on Calcareous Soils. New Phytol. 1970, 69, 143–157. [Google Scholar] [CrossRef]
- Fassnacht, F.E.; Stenzel, S.; Gitelson, A.A. Non-destructive estimation of foliar carotenoid content of tree species using merged vegetation indices. J. Plant Physiol. 2015, 176, 210–217. [Google Scholar] [CrossRef] [PubMed]
- Simon, U.K.; Polanschütz, L.M.; Koffler, B.E.; Zechmann, B. High Resolution Imaging of Temporal and Spatial Changes of Subcellular Ascorbate, Glutathione and H2O2 Distribution during Botrytis cinerea Infection in Arabidopsis. PLoS ONE 2013, 8, e65811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heuser, T.; Zimmer, W. Quantitative analysis of phytopathogenic ascomycota on leaves of pedunculate oaks (Quercus robur L.) by real-time PCR. FEMS Microbiol. Let. 2002, 209, 295–299. [Google Scholar] [CrossRef]
- Chalker-Scott, L. Environmental Significance of Anthocyanins in Plant Stress Responses. Photochem. Photobiol. 1999, 70, 1–9. [Google Scholar] [CrossRef]
- Karageorgou, P.; Manetas, Y. The importance of being red when young: Anthocyanins and the protection of young leaves of Quercus coccifera from insect herbivory and excess light. Tree Physiol. 2006, 26, 613–621. [Google Scholar] [CrossRef] [Green Version]
- Sivankalyani, V.; Feygenberg, O.; Diskin, S.; Wright, B.; Alkan, N. Increased anthocyanin and flavonoids in mango fruit peel are associated with cold and pathogen resistance. Postharvest Biol. Technol. 2016, 111, 132–139. [Google Scholar] [CrossRef]
- Hatier, J.H.B.; Gould, K.S. Anthocyanin Function in Vegetative Organs. In Anthocyanins; Winefield, C., Davies, K., Gould, K., Eds.; Springer: New York, NY, USA, 2008; pp. 1–19. [Google Scholar]
- Iriti, M.; Rossoni, M.; Borgo, M.; Faoro, F. Benzothiadiazole Enhances Resveratrol and Anthocyanin Biosynthesis in Grapevine, Meanwhile Improving Resistance to Botrytis cinerea. J. Agric. Food Chem. 2004, 52, 4406–4413. [Google Scholar] [CrossRef] [PubMed]
- Chiwetalu, U.J.; Mbajiorgu, C.C.; Ogbuagu, N.J. Remedial ability of maize (Zea mays) on lead contamination under potted condition and non-potted field soil condition. J. Bioresour. Bioprod. 2020, 5, 51–59. [Google Scholar] [CrossRef]
- Hughes, N.M.; Neufeld, H.S.; Burkey, K.O. Functional role of anthocyanins in high-light winter leaves of the evergreen herb Galax urceolata. New Phytol. 2005, 168, 575–587. [Google Scholar] [CrossRef]
- Coram, T.E.; Wang, M.; Chen, X. Transcriptome analysis of the wheat–Puccinia striiformis f. sp. tritici interaction. Mol. Plant Pathol. 2008, 9, 157–169. [Google Scholar] [CrossRef] [PubMed]
- Del Río, L.A. ROS and RNS in plant physiology: An overview. J. Exp. Bot. 2015, 66, 2827–2837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torres, M.A.; Jones, J.D.; Dangl, J.L. Reactive Oxygen Species Signaling in Response to Pathogens. Plant Physiol. 2006, 141, 373–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef] [Green Version]
- Asada, K. Production and Scavenging of Reactive Oxygen Species in Chloroplasts and Their Functions. Plant Physiol. 2006, 141, 391–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.; Li, Y.; Wang, M.; Wang, C.; Ling, N.; Mur, L.A.; Shen, Q.; Guo, S. Redox imbalance contributed differently to membrane damage of cucumber leaves under water stress and Fusarium infection. Plant Sci. 2018, 274, 171–180. [Google Scholar] [CrossRef]
- Henmi, T.; Miyao, M.; Yamamoto, Y. Release and Reactive-Oxygen-Mediated Damage of the Oxygen-Evolving Complex Subunits of PSII during Photoinhibition. Plant Cell Physiol. 2004, 45, 243–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishiyama, Y.; Allakhverdiev, S.I.; Murata, N. A new paradigm for the action of reactive oxygen species in the photoinhibition of photosystem II. Biochim. Biophys. Acta BBA Bioenerg. 2006, 1757, 742–749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Q.; Su, L.-J.; Chen, J.-W.; Zeng, X.-Q.; Sun, B.-Y.; Peng, C.-L. The antioxidative role of anthocyanins in Arabidopsis under high-irradiance. Biol. Plant. 2012, 56, 97–104. [Google Scholar] [CrossRef]
- Geat, N.; Singh, D.; Khirbat, S. Effect of Non-conventional Chemicals and Synthetic Fungicide on Biochemical Characteristics of Chilli against Fruit Rot Pathogen Colletotrichum capsici. J. Plant Pathol. Microbiol. 2016, 7, 605–613. [Google Scholar] [CrossRef]
- Ali, M.B.; Hahn, E.-J.; Paek, K.-Y. Effects of temperature on oxidative stress defense systems, lipid peroxidation and lipoxygenase activity in Phalaenopsis. Plant Physiol. Biochem. 2005, 43, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Kabiri, R.; Nasibi, F.; Farahbakhsh, H. Effect of exogenous salicylic acid on some physiological parameters and alleviation of drought stress in Nigella sativa plants under hydroponic hulture. Plant Protec. Sci. 2014, 50, 43–51. [Google Scholar] [CrossRef] [Green Version]
- Costa, M.; Civell, P.M.; Chaves, A.R.; Martinez, G.A. Effects of ethephon and 6-benzylaminopurine on chlorophyll degrading enzymes and peroxidase-linked chlorophyll bleaching during post-harvest senescence of broccoli (Brassica oleracea L.) at 20 °C. Postharvest Biol. Technol. 2005, 35, 191–199. [Google Scholar] [CrossRef]
- Chakhchar, A.; Wahbi, S.; Lamaoui, M.; Ferradous, A.; El Mousadik, A.; Ibnsouda-Koraichi, S.; Filali-Maltouf, A.; El Modafar, C. Physiological and biochemical traits of drought tolerance in Argania spinosa. J. Plant Interact. 2015, 10, 252–261. [Google Scholar] [CrossRef]
- Liang, C.; Liu, T.; Zhao, Y.; Feng, Y.; Wan, T.; Cai, Y. Defense Responses of Cherry Rootstock ‘Gisela 6’ Elicited by Agrobacterium tumefaciens Infection. J. Plant Growth Regul. 2019, 38, 1082–1093. [Google Scholar] [CrossRef]
- Montillet, J.-L.; Agnel, J.-P.; Ponchet, M.; Vailleau, F.; Roby, M.; Triantaphylidès, C. Lipoxygenase-mediated production of fatty acid hydroperoxides is a specific signature of the hypersensitive reaction in plants. Plant Physiol. Biochem. 2002, 40, 633–639. [Google Scholar] [CrossRef]
- Fontaine, F.; Pinto, C.; Vallet, J.; Clément, C.; Gomes, A.C.; Spagnolo, A. The effects of grapevine trunk diseases (GTDs) on vine physiology. Eur. J. Plant Pathol. 2015, 144, 707–721. [Google Scholar] [CrossRef]
- Ge, Y.; Bi, Y.; Guest, D.I. Defense responses in leaves of resistant and susceptible melon (Cucumis melo L.) cultivars infected with Colletotrichum lagenarium. Physiol. Mol. Plant Pathol. 2013, 81, 13–21. [Google Scholar] [CrossRef]
- Koç, E.; Üstün, A.S.; Islek, C.; Arıcı, Y.K.; Arici, Y.K. Defence responses in leaves of resistant and susceptible pepper (Capsicum annuum L.) cultivars infected with different inoculum concentrations of Phytophthora capsici Leon. Sci. Hortic. 2011, 128, 434–442. [Google Scholar] [CrossRef]
- Liu, B.; Li, M.; Cheng, L.; Liang, N.; Zou, Y.; Ma, F. Influence of rootstock on antioxidant system in leaves and roots of young apple trees in response to drought stress. Plant Growth Regul. 2012, 67, 247–256. [Google Scholar] [CrossRef]
- Mohammadi, H.; Moradi, F. Effects of growth regulators on enzymatic and non-enzymatic antioxidants in leaves of two contrasting wheat cultivars under water stress. Braz. J. Bot. 2016, 39, 495–505. [Google Scholar] [CrossRef]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Prochazkova, D.; Sairam, R.; Srivastava, G.; Singh, D. Oxidative stress and antioxidant activity as the basis of senescence in maize leaves. Plant Sci. 2001, 161, 765–771. [Google Scholar] [CrossRef]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive Oxygen Species, Oxidative Damage, and Antioxidative Defense Mechanism in Plants under Stressful Conditions. J. Bot. 2012, 2012, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Guha, A.; Sengupta, D.; Rasineni, G.K.; Reddy, A.R. Non-enzymatic antioxidative defense in drought-stressed mulberry (Morus indica L.) genotypes. Trees 2012, 26, 903–918. [Google Scholar] [CrossRef]
- Noctor, G.; Foyer, C.H. Intracellular redox compartmentation and ROS-related communication in regulation and signaling. Plant Physiol. 2016, 171, 1581–1592. [Google Scholar] [CrossRef] [Green Version]
- Mullineaux, P.M.; Rausch, T. Glutathione, photosynthesis and the redox regulation of stress-responsive gene expression. Photosynth. Res. 2005, 86, 459–474. [Google Scholar] [CrossRef]
- Nahar, K.; Hasanuzzaman, M.; Alam, M.M.; Fujita, M. Glutathione-induced drought stress tolerance in mung bean: Coordinated roles of the antioxidant defense and methylglyoxal detoxification systems. Aob Plants 2015, 7, plv069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choudhary, A.; Pandey, P.; Senthil-Kumar, M. Tailored Responses to Simultaneous Drought Stress and Pathogen Infection in Plants. In Drought Stress Tolerance in Plants; Hossain, M., Wani, S., Bhattacharjee, S., Burritt, D., Tran, L.S., Eds.; Springer: Cham, Germany, 2016; Volume 1, pp. 427–438. [Google Scholar]
- Gong, B.; Sun, S.; Yan, Y.; Jing, X.; Shi, Q. Glutathione Metabolism and Its Function in Higher Plants Adapting to Stress. In Antioxidant Enzymes in Higher Plants; Gupta, D., Palma, J., Corpas, F., Eds.; Springer: Cham, Germany, 2018; pp. 181–205. [Google Scholar]
- Großkinsky, D.K.; Koffler, B.E.; Roitsch, T.; Maier, R.; Zechmann, B. Compartment-Specific Antioxidative Defense in Arabidopsis Against Virulent and Avirulent Pseudomonas syringae. Phytopathology 2012, 102, 662–673. [Google Scholar] [CrossRef] [Green Version]
- Modarres, R.; Sarhadi, A.; Burn, D.H. Changes of extreme drought and flood events in Iran. Glob. Planet. Chang. 2016, 144, 67–81. [Google Scholar] [CrossRef]
- Bader, M.-F.; Leuzinger, S. Hydraulic Coupling of a Leafless Kauri Tree Remnant to Conspecific Hosts. iScience 2019, 19, 1238–1247. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Pathogen | Drought Stress | Pathogen × Drought Stress |
|---|---|---|---|
| Quercus libani | |||
| Survival | 1.500 ns | 21.125 ** | 3.896 * |
| Diameter growth | 11.295 ** | 31.118 ** | 4.235 * |
| Height growth | 6.816 * | 125.28 ** | 3.956 * |
| Average leaf area | 48.672 ** | 217.313 ** | 3.902 * |
| Net photosynthesis | 35.765 ** | 214.25 ** | 4.937 * |
| Stomatal conductance | 52.676 ** | 343.05 ** | 4.324 * |
| Transpiration | 56.468 ** | 433.24 ** | 4.850 * |
| WUEi | 4.758 ** | 21.330 ** | 5.614 * |
| RWC | 75.718 ** | 536.23 ** | 4.536 * |
| Chlorophyll a+b | 14.262 ** | 189.14 ** | 9.029 * |
| Carotenoid | 9.046 ** | 205.59 ** | 12.767 * |
| Anthocyanin | 59.614 ** | 7.501 * | 3.165 ns |
| EL | 34.495 ** | 190.84 ** | 4.778 * |
| LOX | 11.462 ** | 82.00 ** | 4.459 * |
| O2•− | 38.971 ** | 127.83 ** | 14.693 ** |
| AA | 78.780 ** | 51.660 ** | 5.514 * |
| GSH | 10.621 ** | 3.238 ns | 3.511 ns |
| Total soluble protein | 53.440 ** | 32.803 ** | 3.117 ns |
| Quercus infectoria | |||
| Survival | 0.167 ns | 20.167 ** | 1.163 ns |
| Diameter growth | 1.500 ns | 32.667 ** | 4.164 * |
| Height growth | 4.433 * | 90.133 ** | 3.833 * |
| Average leaf area | 14.873 ** | 200.587 ** | 8.444 ** |
| Net photosynthesis | 22.028 ** | 81.411 ** | 4.017 * |
| Stomatal conductance | 24.939 ** | 340.48 ** | 4.030 * |
| Transpiration | 7.563 * | 8.49 * | 4.180 * |
| WUEi | 1.238 ns | 31.069 ** | 4.076 * |
| RWC | 24.325 ** | 326.852 ** | 3.856 * |
| Chlorophyll a+b | 17.337 ** | 122.69 ** | 4.082 * |
| Carotenoid | 17.448 ** | 402.07 ** | 4.135 * |
| Anthocyanin | 20.330 ** | 17.590 ** | 5.208 * |
| EL | 25.739 ** | 317.53 ** | 13.450 ** |
| LOX | 4.529 * | 6.090 * | 3.945 * |
| O2•− | 3.586 ns | 393.21 ** | 3.85 ns |
| AA | 58.707 ** | 97.242 ** | 3.869 * |
| GSH | 60.050 ** | 97.110 ** | 8.871 * |
| Total soluble protein | 73.893 ** | 65.773 ** | 5.069 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ghanbary, E.; Fathizadeh, O.; Pazhouhan, I.; Zarafshar, M.; Tabari, M.; Jafarnia, S.; Parad, G.A.; Bader, M.K.-F. Drought and Pathogen Effects on Survival, Leaf Physiology, Oxidative Damage, and Defense in Two Middle Eastern Oak Species. Forests 2021, 12, 247. https://doi.org/10.3390/f12020247
Ghanbary E, Fathizadeh O, Pazhouhan I, Zarafshar M, Tabari M, Jafarnia S, Parad GA, Bader MK-F. Drought and Pathogen Effects on Survival, Leaf Physiology, Oxidative Damage, and Defense in Two Middle Eastern Oak Species. Forests. 2021; 12(2):247. https://doi.org/10.3390/f12020247
Chicago/Turabian StyleGhanbary, Ehsan, Omid Fathizadeh, Iman Pazhouhan, Mehrdad Zarafshar, Masoud Tabari, Shahram Jafarnia, Ghasem Ali Parad, and Martin Karl-Friedrich Bader. 2021. "Drought and Pathogen Effects on Survival, Leaf Physiology, Oxidative Damage, and Defense in Two Middle Eastern Oak Species" Forests 12, no. 2: 247. https://doi.org/10.3390/f12020247