EvoDevo: An Ongoing Revolution?


Department of Cognitive Sciences, University of Messina, Via Concezione 8, 98121 Messina, Italy
Philosophies 2020, 5(4), 35; https://doi.org/10.3390/philosophies5040035
Submission received: 27 September 2020
/
Revised: 2 November 2020
/
Accepted: 2 November 2020
/
Published: 5 November 2020
(This article belongs to the Special Issue Renegotiating Disciplinary Fields in the Life Sciences)
{kind=link}
Abstract
:Since its appearance, Evolutionary Developmental Biology (EvoDevo) has been called an emerging research program, a new paradigm, a new interdisciplinary field, or even a revolution. Behind these formulas, there is the awareness that something is changing in biology. EvoDevo is characterized by a variety of accounts and by an expanding theoretical framework. From an epistemological point of view, what is the relationship between EvoDevo and previous biological tradition? Is EvoDevo the carrier of a new message about how to conceive evolution and development? Furthermore, is it necessary to rethink the way we look at both of these processes? EvoDevo represents the attempt to synthesize two logics, that of evolution and that of development, and the way we conceive one affects the other. This synthesis is far from being fulfilled, but an adequate theory of development may represent a further step towards this achievement. In this article, an epistemological analysis of EvoDevo is presented, with particular attention paid to the relations to the Extended Evolutionary Synthesis (EES) and the Standard Evolutionary Synthesis (SET).
1. Introduction: The House That Charles Built
Evolutionary biology, as well as science in general, is characterized by a continuous genealogy of problems, i.e., “a kind of dialectical sequence” of problems that are “linked together in a continuous family tree” [1] (p. 148).
Let’s start with Charles Darwin, whose masterpiece On the Origin of Species still has a fundamental influence on biological thinking [2]. Darwin specified three foundational problems on which every evolutionary theory must be based: Variation, selection, and inheritance. The process of descent with modification needs some source able to produce variation in organisms. This variation must be reliably passed on to subsequent generations. Finally, variation will be maintained within a lineage if it confers an adaptive advantage.
However, as observed by Darwin himself, it is not natural selection that induces variation, but its role is limited to maintaining favorable variations and eliminating harmful ones [2] (p. 63). “Laws of variations” and “laws of heredity” are the most notable absentees within Darwin’s long argument. “The laws governing inheritance are quite unknown; no one can say why a peculiarity in different individuals of the same species, or in individuals of different species, is sometimes inherited and sometimes not so; why the child often reverts in certain characters to its grandfather or grandmother or other more remote ancestor; […].” [2] (p. 13).
“I have hitherto sometimes spoken as if the variations […] had been due to chance. This, of course, is a wholly incorrect expression, but it serves to acknowledge plainly our ignorance of the cause of each particular variation.” [2] (p. 102).
Anyway, Evolution (Natural Selection), Variation, and Heredity represent the three foundational issues from which are derived the genealogy of problems that have long affected evolutionary biological thought. These three issues represent the conceptual triad to be taken into account in any evolutionary theory [3,4].
Nevertheless, biological thinking did not address these foundational problems in the same way, and the phenomena and roles attributed to each element of the conceptual triad have always been different.
Within the Modern Synthesis (MS), particular emphasis was placed on the role of natural selection and genes, the latter being conceived both as unit of variation and unit of inheritance. In this framework, evolution was defined as a change in the frequency of an allele within a gene pool [5] or, in a simpler way, “biological evolution consists of changes in the genetic constitution of populations” [6] (p. 21). However, as said by Scott Gilbert, reported in [7], “The modern synthesis is remarkably good at modeling the survival of the fittest, but not good at modeling the arrival of the fittest” [7] (p. 281).
This is one of the reasons for dissatisfaction with the Modern Synthesis. Another reason is that MS has black-boxed all the processes and mechanisms that are between genotype and phenotype or between genetic mutation and natural selection [8]. As stated by one of the greatest critics of MS, Conrad Hal Waddington: “It has been clear at least since von Baer’s day that a theory of evolution requires, as a fundamental part of it, some theory of development. Evolution is concerned with changes in animals, and it is impossible profitably to discuss changes in a system unless one has some picture of what the system is like. Since every aspect of an animal is a product of development, or rather is a temporary phase of a continuous process of development, a model of the nature of animal organization can only be given in developmental terms” [9] (p. 11).
Evolutionary Developmental Biology (EvoDevo) is the theoretical proposal able to unlock the black box of development, and also able to investigate the influence of development on evolution and the influence of evolution on development [10,11]. So, one of the aims of EvoDevo is to investigate the causal-mechanistic interactions which exist between the processes of individual development and the processes of evolutionary change [12], and to identify the generative mechanisms responsible for the variations presented by organisms (the arrival of the fittest). However, as we will see in Section 3, the conceptual framework of EvoDevo is anything but homogeneous. This conceptual framework is characterized by a set of core concepts, such as: Novelty [13,14]; homology [15,16]; genetic regulatory networks [17,18]; evolvability [19,20]; developmental bias [21]; modularity [22]; and others. Evodevo makes use of a combination of “[…] tools, techniques, and findings of molecular biology, anatomy, physiology, functional morphology, cell biology, embryology, developmental genetics, paleontology, comparative genomics, and population genetics, […]” [23] (p. 1). It is composed of a variety of research programs, such as comparative embryology and morphology programs, evolutionary developmental genetics programs, experimental epigenetic programs, and theoretical and computational programs [12,24]. Finally, it is guided by several questions concerning the relationships between evolution, development, and environment [10,12]. For these reasons, this pluralistic framework has been rightly conceived as a “[…] loose conglomeration of research programs […]” [25] (p. 265).
Anyway, EvoDevo represents a hard opponent for the MS. In fact, it was presented as: A paradigm [24,26]; a research program [27]; a revolution or revolutionary science [17,28]; or a new synthesis or a revolutionary synthesis [29,30]. These terms, far from being linguistic labels, represent theoretical and conceptual tools useful for a theoretical comparison between different perspectives; such as MS and EvoDevo. Nevertheless, the relationship between these two perspectives is far from clear. Is EvoDevo an expansion, an extension, or a revolution compared to the Modern Synthesis? EvoDevo “[…] is a discipline still in search of its identity” [27] (p. 213) whose conceptual and theoretical foundation has not yet been properly founded [27,31].
Section 2 describes the pluralistic framework of EvoDevo, with particular attention paid to how the various accounts conceive the processes of evolution and development.
In Section 3, we highlight the relationship between EvoDevo with the Extended Evolutionary Synthesis (EES) and Standard Evolutionary Theory (SET).
In the last section, we present some theoretical proposals to make EvoDevo take a few steps forwards.
2. One, No One, and Hundred Thousand EvoDevo
Vitangelo Moscarda is the protagonist of the novel ‘One, no one, and hundred thousand’, written by the Sicilian Nobel laureate Luigi Pirandello. In this novel, the protagonist realizes how his identity, contrary to what he believed, is not something certain, stable, and consistent for everyone, but each person has their own view of the personality and characteristics of Moscarda. The result is that his identity becomes questionable: From the certainty of a single initial identity to an identity distributed over a multiplicity of individuals, and finally to the impossibility of any form of identity. Although not afflicted by a comparable existential drama, we could say that the situation of the EvoDevo is similar, in some respects, to that of Vitangelo Moscarda.
2.1. One EvoDevo
Frequently, EvoDevo is defined as the study of how Evolution and Development are interrelated or, to put it another way, as the “evolution’s influence on development and development’s influence on evolution” [32] (p. 9). Moreover, how to conceive EvoDevo, that is, what kind of problems, themes, or aspects must be considered fundamental and worthy of investigation, “depends on whether the scientist is a developmental biologist, a paleontologist or an evolutionary biologist”, or something else [30] (p. 75). Although it is a discipline that is more than 30 years old, EvoDevo, from a theoretical point of view, is (still) a discipline in search of its identity [27,32].
EvoDevo would like to be “[…] a synthesis of those processes operating during ontogeny with those operating between generations (during phylogeny)” [33] (p. xiii) (Hall&Olson 2003, p. xiii), and also an integration of several research areas such as genetics, ecology, paleontology, behavior, cognition, physiology, and so on [34] (p. 499). This framework is characterized by several core concepts (from life cycles evolution, modularity, plasticity, to evolvability, environmental induction, canalization, and more) [34] (p. 505), and by a set of problems agenda, whose purpose is to investigate the questions born at the interface between evolution, development (EvoDevo questions; DevoEvo questions), and also environment (EcoEvoDevo questions) [12].
So, we can say that the general aim of EvoDevo is “[…] to provide a mechanistic explanation of how developmental mechanisms have changed during evolution, and how these modifications are reflected in changes of organismal form”, in such a way as to make possible “[…] to determine the mechanisms behind the ‘arrival of the fittest’” [35] (p. 2). The problems begin when we enter in the “[…] incredibly intricate multi-level domains, mechanisms, processes, structures […]” [36] (p. 259) that characterize development and start looking for the mechanisms responsible for the generation and variation of forms.
However, “the way we perceive the field today is often reflected in the way we reconstruct its history and, similarly, the way we reconstruct the history can reveal a lot about current assumptions” [37] (p. 2).
2.2. No One EvoDevo
A revolutionary event in the history of developmental genetics, so prominent that for some authors it represents the birth of the EvoDevo [17], has been the discovery of the homeobox [38,39] and of the role that this plays in the evolution and development of animal forms. The discovery was made in the 1980s. In the following years emerged what has been called the logic that controls development, based on a regulatory system in which a set of genes and transcription factors acts on other genes, activating or inhibiting them, in this way determining “[…] variations in the level, pattern, or timing of gene expression […]” [40] (p. 579) which result in the construction of forms during development. This set of genes is highly conserved in the course of evolution, and it is held responsible for the construction of body plans and different body structures [17].
The discovery of the homeobox has been described as the Rosetta Stone of developmental genetics; on this foundation, it would be possible to achieve an understanding of the “[…] deep unity about pattern formation […] which underlies the apparent superficial complexity and diversity of animal development” [41] (p. 365). Both homology and evolutionary novelty are thus explained in terms of modifications based on the combinatory logic of activation/inhibition of the cis-regulatory systems; what has been called by Gould “Hoxology” [42].
The central role played by this gene regulation system, both in evolution and development, is expressed by what we may call Davidson’s Syllogism: “Since the morphological features of an animal are the product of its developmental process, and since the developmental process in each animal is encoded in its species-specific regulatory genome, then change in animal form during evolution is the consequence of change in genomic regulatory programs for development” [43] (p. 27–28).
What characterizes the Hoxological approach is an explicit gene-centrism, according to which the great variety of animal forms is encoded in the combinatory logic of the cis-regulatory system, conceived as “[…] the essence of animal development” [44] (p. 25).
What is more interesting is that, according to some exponents of this approach [17,45], the EvoDevo/Hoxology represents a revolutionary expansion of Modern Synthesis; a third act (the first one was the Darwinian theorization, the second one was the MS) that complemented the MS with a hitherto missing piece: A mechanistic explanation for the evolution and development of phenotypes. As stated in [45], both phyletic and morphological evolution are characterized by the same genetic and developmental mechanisms, even suggesting an equation between Macroevolution = Microevolution. If so, EvoDevo would not represent “[…] any fundamental conceptual problem for evolutionary biology” [46] (p. 386); rather, it would be the completion of the conceptual framework of MS through the addition of causal mechanisms able to explain the origins of variation. However, this perspective was accepted with both cautious optimism [47] and skepticism [48] by exponents of the Standard Evolutionary Theory.
But as pointed out by [37], “[…] evolutionary developmental biology is not as uniform as the image of a new scientific discipline and the powerful icons of “the genetic toolkit for development” and the almost magical qualities of “Hox genes” seems to suggest.” (p. 1). The study of the evolution and development of the phenotype “[…] requires thinking both within and outside the paradigm of transcription encoding factors” [49] (p. 129). So, EvoDevo must be something more than a “mere” Hoxology.
2.3. One Hundred Thousand EvoDevo
Marta Linde Medina introduces a distinction between “two versions of EvoDevo” [50], which reflects internalist and externalist traditions—as presented by Ronald Amundson [51], and which represents two ways of conceptualizing evolution and development. One version, EvoDevo1, is linked to the discovery of Hox genes in the 1980s [52,53], and the consequent progress of developmental genetics [17,43]. This EvoDevo perspective is closely based on a gene-centric conception of both evolution and development, in which are often used metaphors such as those of the genetic program and of genes as depositories of ontogenetic and phylogenetic information. The development of organismal form is based on switches, regulatory sequences, transcription factors that make up a complex genetic regulatory network on which depends the construction, as well as the evolution, of body plans. The Natural Selection will act on variations in the wiring of the networks able to produce both microevolutionary and macroevolutionary phenomena. Thus, we are faced with a “perfect” synthesis of mechanisms acting during ontogeny and the mechanism of natural selection [17,43], in harmony with the externalist tradition. The other version, EvoDevo2, is linked to an internalist perspective, in which a fundamental role is recognized to the inherent properties of living matter [54], the self-organizing phenomena and events [55], and the overall set of interactions that occur during ontogeny and across several levels of organization [56]. In this perspective, the genetic circuitry, even if it plays an important role, is conceived as a post-hoc effect, i.e., a stabilizing factor of forms, rather than an exclusive generating factor. This last perspective is seen as “[…] a subdiscipline of an extended evolutionary synthesis (EES) […]” [50] (p. 9). On the other hand, the version presented by [17], for example, is more a genetic theory of morphological evolution [46], and therefore it is only one component, one aspect, of a more general theory of evolution where development is finally and adequately taken into consideration.
This first distinction is useful because it allows making a coarse theoretical distinction between two alternative conceptions of EvoDevo [25]: EvoDevo in a narrow sense, including mainly Hoxology; and EvoDevo in a broad sense, including a wider set of perspectives. This distinction can be further developed into a typological classification of positions, based on [57].
Callebaut, Müller, and Newman [57] identify various accounts of Evo-Devo or, as they call them, twelve evo-devo packages, “[…]which in part reinforce one another, but in part also compete” (p. 34). These accounts are the result of the combination of different theoretical, methodological, and epistemological perspectives. We consider it useful to present an overview of these accounts to highlight the plurality of perspectives that characterizes EvoDevo. In the end, we will make some general remarks.
Core EvoDevo (1) is described as a perspective “[…] encompassing both the program to explain evolution through changes in development and the program to reframe evolutionary biology more drastically along developmental lines” [57] (p. 35).
Gene Regulatory Evolution (2) basically corresponds to the Hoxology presented in the previous section.
Epigenetic EvoDevo (3) has as its main target morphological evolution; notably the processes and mechanisms that characterize the generation, fixation, and variation of structural building elements of organisms [58]. Within this perspective, a further problem is represented by the genotype–phenotype relation, that is, how genes and morphologies are related. On the one hand, genetic and developmental pathways may change during phylogeny while morphologies remain relatively constant; on the other hand, similar gene expression patterns may correspond to different morphologies [15]. In order to describe the processes of form generation, it is not enough to give a description of the genetic control systems: Our account must include “[…] the generic material properties of, and interaction dynamic among, cells, tissues and their environments”[59] (p. 66). The latter factors constitute the epigenetic properties of development, such as: “(i) interactions of cell metabolism with physicochemical environment within and external to the organism; (ii) interactions of tissues masses with the physical environment on the basis of physical laws inherent to condensed materials, and (iii) interaction among tissue themselves, according to an evolving set of rules” [56] (pp. 305–306). The relationship between genes and forms is indirect, and epigenetic mechanisms represent the principal factor directing morphological evolution; meanwhile genetic networks are co-opted subsequently to play a stabilizing role rather than a generative one. In the Epigenetic account, homology has an important role, insofar as the main clue of a common descent and of organismic evolution. Indeed, homologues represent attractors of morphological design, i.e., “autonomous organizer of the phenotype in an evolutionary lineage” [59] (p. 70–71), and that during the phylogeny provides the basis for further modifications. Therefore, there is an attempt to recognize a relatively autonomous role of the morphological level with respect to the underlying mechanisms, especially the genetic ones.
Process Structuralism (4) “[…] assumes that there is a logical order to the biological realm and that organisms are generated according to rational dynamic principles” [60] (p. 91). So, a theory of evolution must include a theory of biological form, which explains how organisms are structured and by which transformations are characterized. This account puts center stage organisms conceived as “dynamically transforming systems” described as fields, i.e., “[…] domains of spatial order, defined by internal relationships, that change in time according to well-defined principles or rules” [61] (p. 129), and moves against the Neo-Darwinian assumptions according to which, to explain evolutionary phenomena, it is enough the action of natural selection on random genetic mutations [62]. So, what is proposed is a theory of morphogenetic fields defined as complex dynamic systems in which “[…] genetic and environmental factors determine parametric values in the equations describing the field and therefore act to select or stabilize one manifest form from the set of forms which are possible for that type of field” [61] (p. 121). In this perspective, the patterns of gene expression alone cannot provide neither a causal explanation of the developmental processes nor an understanding of the generative principles underlying the development and evolution of forms. On the contrary, development is conceived as a hierarchical process along which various factors can alter the dynamics of the system. In this way, it is possible to define the space of possible biological forms (generic forms), i.e., “the natural kinds that are revealed in evolution, the basic biological forms that are all transformations of one another under changes in the detailed dynamics of morphogenetic fields” [61] (p. 130).
The morphogenetic field represents the generative unit both in development and evolution. Evolution is conceived as a “time-dependent exploration of a set of possibilities under internal (genetic) and external (environmental) parametric variation” [60] (p. 96); these possibilities are given by “generic states of forms […] whose distribution in the space of developmental trajectories defines the set of possible forms” (p. 96). What emerges is a perspective on evolution and development in which: “The hierarchical nature of this generative process leads naturally to a hierarchical taxonomy of biological forms: ontogenesis provides the logical foundation for understanding phylogenesis” [60] (p. 130).
Self-Organization of Biological Complexity (5) explains the complexity of the organization and forms of biological systems as due to intrinsic dynamics and patterns, rather than to the external action of natural selection. This account, as was Process Structuralism, was inspired by the tradition of Rational Morphology (Goethe, Cuvier, Geoffroy Saint-Hilaire, Owen, and more recently, D’Arcy Thompson), according to which “[…] organisms were built up by combinatorial variations of a small number of principles”[55] (p. 4). The aim of Rational Morphology, as well as of the Self-Organization account, is to discover the laws of form underlying biological organization. We can define Self-Organization as “[…] a process in which pattern emerges at the global (collective) level through interactions among the components of the system at the individual level, without these interactions explicitly specifying the global pattern […]” [63] (p. 79).
This means that the order performed by the organisms is not due to the action of natural selection, which does not have the ability to generate order, but to the inherent properties of complex systems which constitute the organisms themselves [55]. The main processes occurring during development, endowed with self-organizing properties, are cellular differentiation and morphogenesis. Cellular differentiation is the result of the dynamical behaviors of genetic regulatory networks, in which cell types represent “[…] a recurrent pattern of gene activity” [55] (p. 442). By contrast, morphogenesis is defined as “[…] the consequence in time and space of the structural and catalytic proprieties of proteins encoded in time and space by the genome, acting in concert with nonprotein materials and with physical and chemical forces to yield reliable forms” [55] (p. 410). Within this account, a significant problem is represented by the relationship between natural selection and self-organization. There are different possible hypotheses: A primacy of natural selection over self-organization; self-organization as a constraint or an auxiliary of natural selection; natural selection and self-organization as two sides of the same coin [64]. The last hypothesis is suggested by Kauffman: “Selection achieves and maintains complex systems poised on the boundary, or edge, between order and chaos. This systems are best able to coordinate complex tasks and evolve in a complex environment. The typical, or generic, properties of such poised systems emerge as potential ahistorical universals in biology” [55] (p. xv).
Structural Modeling (6) is a computational and theoretical approach to the study of RNA folding, that can be presented as “minimal model of a genotype-phenotype relation” [65] (p. 1164), but it cannot represent the entire process of organismal development. Through this model, it is possible to explore what kind of molecular phenotypes (RNA secondary structure, or RNA shape) can be reached from gradual modifications of the genotype. Moreover, it is possible to understand how genotype and phenotype influence each other, that is, how particular phenotypes are produced from particular genotypes, how these phenotypes are targets of selection, and, finally, how this selection feeds back on the genotypes and developmental mechanisms producing the initial phenotypes. The evolution of the phenotype depends on the topological structure of the phenotypic space as result of the genotype–phenotype map. What emerges from the simulations is that the possible phenotypes accessible from various genotype sequences are limited; there is a redundant mapping that maps many genotypes onto a single phenotype [66]. By gradually varying the sequences of the genotype, it is possible to observe the result on the phenotypic space. Many variations will be neutral, that is, will not lead to a significant change in the process of folding of RNA and, consequently, in the final shape of RNA. So, a Neutral Network can be defined as “[…] a mutationally connected set of genotypes that map to the same phenotype” [66] (pp. 79–80). Therefore, it is possible to distinguish between continuous and discontinuous phenotypic changes: The former results from the exploration within the boundary of a neutral network, while the latter results from crossing the boundary between two adjacent neutral networks, mapping onto two different phenotypes. Although this perspective does not provide a general representation of the development of organisms, it allows us to study, in a limited and controlled manner, phenomena such as epistasis, phenotypic plasticity, constraints on variation, canalization, modularity, phenotypic robustness, and evolvability, whose large-scale study would require considering a large and complex set of factors and variables such as to make investigation intractable.
Dialectical Account (7) rejects the ideology of biological/genetic determinism, which reduces properties and characteristics of complex biological systems to the organization and processes of their simplest constituent components (genes), to which causal and ontological priority is also attributed [67,68]. Conversely: “Dialectical explanations, on the contrary, do not abstract properties of parts in isolation from their associations in wholes but see the properties of parts as arising out of their associations. […] according to the dialectical view, the properties of parts and wholes codetermine each other” [68] (p. 11).
This means that in the explanation of development, there is no place for genetic determinism or program metaphors; neither for a naive dualistic conception according to which some traits are caused by genes while the environment plays the role of filling the remaining causal gaps. On the contrary, development is a process in which there is constant interaction between genes and environment where “[…] random variation in growth and division of cell during development […]” represents an important source of variation (developmental noise) during ontogeny [67,69]. Moreover, the environment is not conceived as something external and independent from organisms and to which organisms must adapt, but environment and organism are engaged in a process of co-determination [69] in which: “[…] organisms determine which elements of external world are put together to make their environments and what are the relations among the elements that are relevant to them” [69] (p. 51); moreover, “[…] organisms not only determine what aspects of the outside world are relevant to them by peculiarities of their shape and metabolism, but they actively construct […] a world around themselves” (p. 54); and finally, “[…] organisms […] are in a constant process of altering their environment” (p. 55). In this account, both evolution and development must be conceived in a more complex causal perspective, according to which “[…] the genome, the proteome, the traitome, the behaviorome, and the societome […]” [70] (p. 334), and the environment in general, are characterized by complex nonlinear and bidirectional dynamics.
Systems Biology (8) is an emerging approach of which it is difficult to provide a comprehensive definition; within this field are present various traditions, such as a physiological [71] and a computational one [72]. Anyway, to provide a general definition, systems biology “is the study of the behavior of complex biological organization and processes in terms of the molecular constituents” [73] (p. 504). The aim is to obtain a “system-level understanding of biological systems” [72], and to achieve this goal, it is necessary to acquire and integrate many kinds of data; especially from the ‘-omics’ disciplines [74]. From the analysis of these complex biological networks, it may be possible to understand the interaction between DNA, phenotype, and environment [71]. The principal aim of this approach is a multi-level integration [75]: “Identifying all genes and proteins in an organism is like listing all the parts of an airplane. While such a list provides a catalog of the individual components, by itself it is not sufficient to understand the complexity underlying the engineered object” [76] (p. 1622).
This means that systems biology moves beyond the single level of analysis method to adopt an integrative perspective, including both top-down and bottom-up approaches. Likewise, exclusive knowledge of genetic regulatory networks and biochemical interaction is not enough to explain the organization of organisms. What must be pursued is an understanding of the organizing principles and the behaviors of the various levels of hierarchical organization. In this sense, systems biology looks for “[…] multiscale, multilevel explanations of organismal properties” [77] (p. 8), in which there is no privileged level of explanation to which to reduce biological phenomena. The complex framework presented by systems biology also forces us to reconsider the causal relationships among levels of organization. Namely, there is not only upward causation as in reductionist models, from genes to organismal behavior, but between levels of organization there are feedback circuits through which higher levels constrain the behavior of the lower levels [78]. This perspective requires a rethinking of development, which cannot be conceived of as a simple, linear, and unidirectional process directed by a program or a single level of organization, but as a complex system of interactions; and afterward a rethinking of the factors and aspects that are relevant in heredity and evolution [75,79].
Cybernetic Synthesis (9) is based on Jean Piaget’s conception of evolution, closely based on the role of phenotypes and behaviors as main actors of evolutionary phenomena [80]. In his conception, behaviors are viewed not only as a result of evolution, but also as one of its determinants [80] (p. xi). Piaget’s theory of organismic development and evolution is influenced by Paul Weiss and Conrad Waddington. If we want to understand the role of behavior in evolution, we must consider the hierarchical system in which the organisms are located. If we conceive the organism as a system of concentric shells, in which the innermost corresponds to genes while the outermost corresponds to the environment. What characterizes organism’s dynamics is a complex network of bidirectional relations [81]. Thus, organisms are characterized by a system of cybernetic interactions, in which a single level (e.g., genetic) cannot cause or control, unilaterally and simply, the highest levels (ex. morphology and behavior). Rather, the highest levels can feed back, altering the dynamics of the lower levels.
Therefore, behavior is conceived “[…] as the expression of the overall dynamics of organization in its interaction with environment and as a source of supersessions and innovations for as long as the environment or environments continue to contain any elements creating obstacles for the organism” [80] (pp. 140–141). In this way, the behavior and other environmental factors can stress a response (in the hierarchical system) of the organism to produce a modified phenotype (phenocopy), which will be stabilized by the genetic level in successive generations [80] (p. 73–83). “The Organism is an open system, a necessary precondition of whose functioning is behavior; and […] it is of the essence of behavior that it is forever attempting to transcend itself and that it thus supplies evolution with its principal motor” [80] (p. 139).
So, according to Piaget, behavior can be considered responsible “[…] for the far-reaching morphogenetic changes of macroevolution” [80] (p. 140).
Reproducer Perspective (10) represents a radical way to think about a unifying theory of heredity, development, and evolution “[…] that accounts for the way heredity and development are entwined in reproduction processes” [82] (p. 253). Units of heredity and units of development are intertwined in units of reproduction [83], and the latter, the reproducers, are the units of evolutionary transition. Reproducers are defined as entities that have the capacity to make more reproducers, i.e., [84] development is presented as “[…] the recursive acquisition (over a compositional hierarchy of parts and wholes) of a capacity of reproduce” [85] (p. 187). In other words, “[…] reproduction is the recursive propagation of the organized capacity to develop and development is the ordered realization of the capacity to reproduce” [86] (p. 267).
In this processual perspective, development is a complex process that does not concern a single level of organization, but involves an organized mechanism of reproduction, spanning over several levels. Meanwhile, evolution is conceived as a “[…] descent with modification of a population of reproducers” [85] (p. 187).
Developmental Systems Perspective (11) is defined as “[…] a general theoretical perspective on development, heredity and evolution” [87] (p. 1), representing an attempt to go beyond classical dichotomies such as nature–nurture, genes–environment, and gene-centric approaches. In this perspective, the fundamental unit is the developmental system conceived “[…] as a complex of interacting influences, some inside the organism’s skin, some external to it, and including its ecological niche in all its spatial and temporal aspects, many of which are typically passed on in reproduction […]. It is in this ontogenetic crucible that form appears and is transformed […]” [88] (p. 39). The stability and variation of forms are not explained by the presence of a genetic program, however complex it may be, but by the contingent process of construction that involves several factors during ontogeny.
Developmental Systems Theory has been regarded as a key partner of EvoDevo, although problematic [3,89]. The major tenets of DST present a serious challenge to traditional evolutionary concepts. This includes [88] (p. 2): (i) The presence of multiple causes involved in developmental processes; (ii) the contingency of development processes towards the context and the other states of the system; (iii) an extended conception of heredity; (iv) a conception of development as a construction (there is not a preconceived plan); (v) there is not a privileged level of control, but this is distributed over multiple factors; (vi) evolution is conceived as changes in the system of interactions between organism and environment.
So, Development Systems Theory conceives “[…] both development and evolution as processes of construction and reconstruction in which heterogeneous resources are contingently but more or less reliably reassembled for each life cycle” [88] (p. 1).
Core Configuration Model (12) represents the attempt to sketch an evo-devo perspective to human cognition, able to explain, in particular, the behaviors and organization of social cognition. Here too, the Neo-Darwinian dichotomy between genes and environment and the gene-centric perspective are refuted and replaced by the notion of recurrence or repeated assembly, conceived as “recurrent entity-environment relations composed of hierarchically organized, heterogeneous components (which may themselves be repeated assemblies) having differing temporal scales and cycles of replication” [90] (p. 59). Biological entities present a hierarchical organization, spanning from DNA, cells, and tissues to organisms, groups, and populations. This hierarchical organization implies that organisms are “[…] the contingently developmental result of various genetic and epigenetic resources […]” [90] (p. 59), and their traits are “the result of a relational linkage, contingently and concretely situated in a specific context of interacting elements spanning between the various level of hierarchical organization characterizing biological entities” [91] (pp. 278–279). In this way, the elements found on a level of organization represent the context for elements of another level. All the elements which constitute that hierarchical system are characterized by both upward and downward causation [91] (p. 278).
Thus, we can define repeated assembly as recurrent constructive relations composed of heterogeneous and hierarchical components. Therefore, development is a non-programmed process, which emerges from the interactions among various resources that must be repeatedly assembled over generations and which must be inherited in a stable manner [91] (p. 279).
2.4. The Search for Unity
This overview of EvoDevo approaches, based on the typological classification presented in [57], is not necessarily complete. In fact, it is possible to identify further theoretical perspectives, which we present here in a concise manner, but whose complexity would require a broader discussion. The Developmental Plasticity perspective (13) is an account that considers the role of developmental plasticity as a leader, not necessarily follower, in phenotypic evolution. The latter is conceived more as a complex reorganizational phenomenon rather than due to a genetic mutation [92,93]. The Ontophylogenetic perspective (14) represents a radical holistic account in which evolution and development “[…] are the two inseparable sides of a single reality produced by a unique process” [94] (p. 5). These two mechanisms are joined in a genealogical line (lignée généalogique) that is the result of a continuous production process of organisms. The unit of analysis of phylogeny, i.e., the species, and the unit of analysis of ontogeny, i.e., the individual, are two aspects of a continuous phenomenon that encompasses both: Ontophylogenesis [95]. The Probabilistic Epigenesis perspective (15) represents a meta-theoretical model of development which emphasizes the role of developmental manifold, conceived as a set of bidirectional influences within and between levels of organization/analysis (including genetic activity, neural activity, behavior, and physical, social and cultural environment) [96,97]. The Eco-Evo-Devo perspective (16) is a complex approach in which “[…] the environment is a source and inducer of genotypic and phenotypic variation at multiple levels of biological organization, while development act as regulator that can mask, release, or create new combination of variation” [98] (p. 107). This perspective aims to discover “[…] the rules by which an organism’s genes, environment, and development interact to create the variation and selective pressures needed for evolution” [99] (p. XIV). The Behavioral/Cognitive perspective (17) investigates the development of behavior and cognition and how these two phenomena can contribute to modify and determine the dynamics of development and evolution [100,101]. The Character Identity Networks perspective (17) is a genetic approach to morphological evolution, which considers the role of genes and genetic networks in determining the identity and variational modalities of a morphological character, also taking into account the developmental context (species-specific mode of development) [18].
However, this does not mean that it is possible to indefinitely extend the set of theoretical proposals related to EvoDevo. The risk is to take an inflationary attitude that identifies more differences in theoretical proposals than necessary. For this reason, it is appropriate to associate a deflationary attitude, which, instead of multiplying the possible perspectives, brings them together, smoothing out the differences. For example, it is possible to combine in a single perspective those proposals (9, 12, 17) that focus on the role of behavior and cognition in the light of ethology (Bateson [100]) and current cognitive theories (Stotz [101]). Moreover, it is possible to amend those Hoxological perspectives by taking into consideration the fundamental role of genetic networks within a broader and more complex developmental context, as in [18,102]. The various accounts presented previously represent theoretical perspectives, that is, particular points of view from which it is possible to investigate evolution and development [103,104]. Each perspective focuses on a particular aspect of the phenomena investigated, highlighting certain characteristics, and omitting others. So, the creation of a typology is a necessary step that must be followed by further elaboration and integration of the various perspectives to develop a general conceptual framework of EvoDevo. This is a complex task of theoretical synthesis, which cannot be done here.
Moreover, it is possible to identify another issue in this typology, namely a theoretical categorization problem, according to which (as pointed out opportunely by one reviewer) different perspectives (specifically from 4 to 10) have been formulated outside the theoretical framework of EvoDevo and sometimes well before it emerged. If by EvoDevo perspectives we have to mean only those that explicitly refer to the same conceptual apparatus, then the solution to this problem would be trivial: It would be enough to identify all the theoretical proposals that use concepts like homology, novelty, genetic regulatory systems, modularity, and so on, to identify which can be labeled as EvoDevo and which cannot. But if the outsider perspectives contribute to a greater understanding of the processes, phenomena, and factors that characterize evolution and development through introducing new theoretical tools, then why not integrate these new tools within EvoDevo′s theoretical apparatus? For example, Self-organization is not a phenomenon that can be found only in the biological field, but also in the physical and chemical fields. Therefore, even if it is a concept that has not been formulated within EvoDevo, it can be a useful tool to understand and describe the development of organisms as a phenomenon that emerges from the organizational complexity of the organisms themselves.
In summary, we consider this typology useful because: (i) It makes explicit the pluralistic framework, or the conglomerate structure, [25] that characterizes EvoDevo; (ii) it allows us to isolate various proposals and positions concerning the three fundamental processes (evolution, development, and heredity); (iii) it provides a conceptual basis for comparing different theoretical proposals with each other; and (iv) it sets a task for the future, i.e., “[…] to probe how deeply entrenched the metacommitments of the advocates of the various perspectives on evo-devo really are” [57] (p. 40).
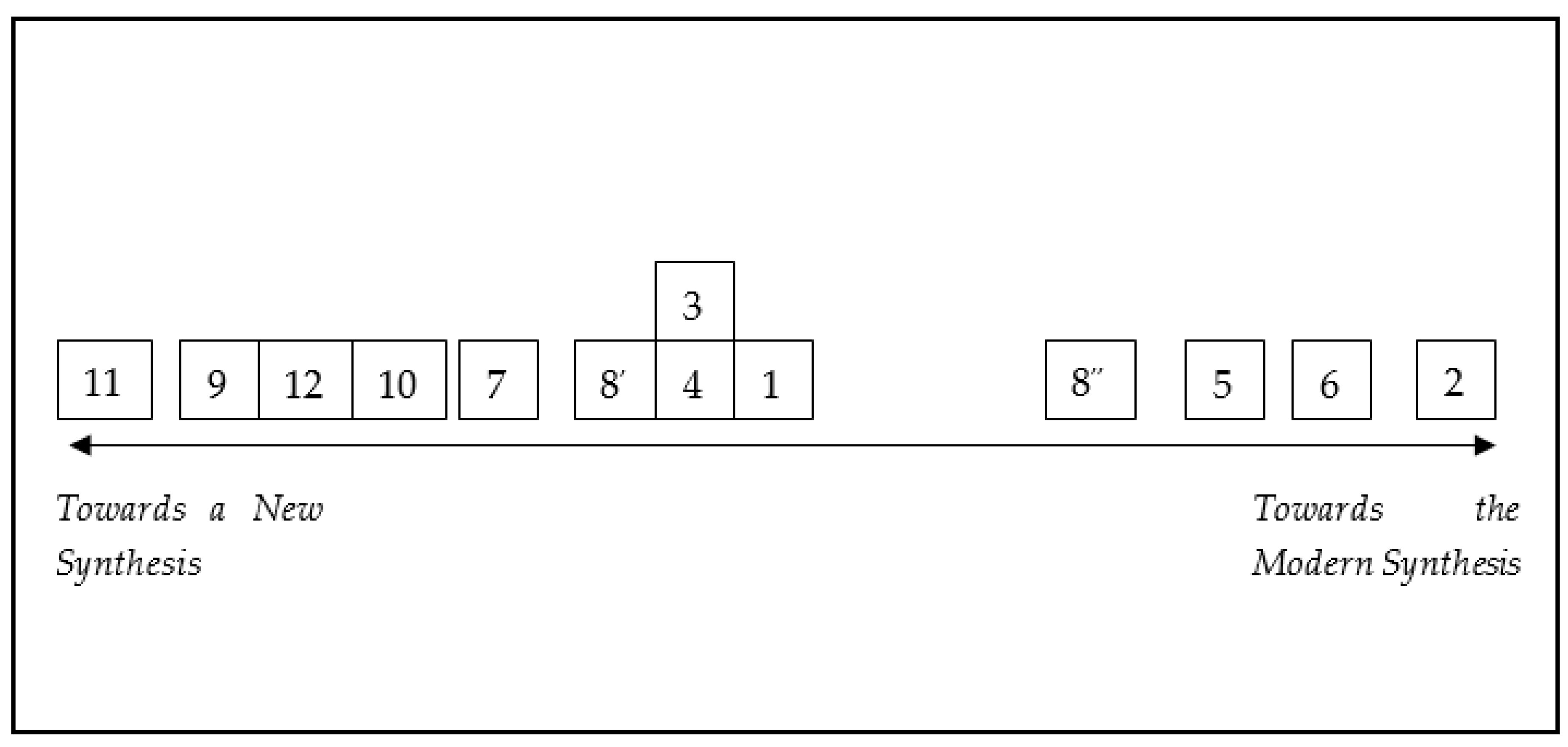
Inspired by Robert [105] (p. 596), we can graphically represent the various perspectives as points distributed on a continuum (Figure 1). We can define a criterion according to which the various positions can be arranged along this continuum according to their degree of inclusiveness. A criterion that takes into account which factors, mechanisms, and processes are involved in explaining and describing evolution and development. Simplifying, we could say that in this theoretical continuum, we can see reductionist to the right and holist to the left [106] (p. 41). On the right side, there are those approaches that reduce development processes to a single level and are based on notions of information, regulation, and control. On the left side, there are those perspectives that have a more global, dynamic, and inclusive approach.
At the right end of the continuum, there is Gene Regulatory Evolution/Hoxology, with its gene-centric and reductionistic approach that presents the DNA/genetic regulatory system as depositary of the instruction to build the organisms, the only hereditary factor is represented by genes, and morphological evolution is due to modification in the regulatory system. This EvoDevo account is closer than the others to the theoretical framework of the MS. At the left end, there is Developmental Systems Theory, conceived not only as a synthesis of evolution, development, and heredity, but also as a radical rethinking of evolutionary theory. In the middle we find the core EvoDevo, guiding the theoretical enterprise and setting the concepts and the problems that have to be addressed. All other accounts are distributed throughout the continuum, according to the criterion of inclusiveness.
This theoretical continuum is an artificial, simplistic, and limited way to represent the relationships between the various perspectives. It represents a theoretical exercise and with obvious limitations; for example, it is not possible to define a metric to quantify the distance between theoretical perspectives. However, we believe that it can be a useful visual tool to understand how some perspectives are more conceptually closer than others, and an initial basis for further and more complex processing.
So, because of this pluralistic framework, it is difficult to conceive EvoDevo as a unitary theory; rather, it seems to be “[…] the pursuit of specific epistemic goals, such as the explanation of evolvability, evolutionary novelty, or homology” [107] (p. 396), or a Trading Zone, i.e., “[…] a variety of disciplines, styles, and paradigms negotiating heavily with one another” [108] (p. 459). It is thanks to this pluralistic framework that EvoDevo plays an ambiguous role within evolutionary thinking. At times, it seems to be in continuity with Modern Synthesis, while at other times a challenge. In the next section, we will investigate the role that EvoDevo plays within evolutionary theory.
3. A Synthesis within a Synthesis
In the last few years, there has been a growing interest in a new evolutionary synthesis, not conceived however as a paradigm shift, or revolution, but as a “[…] series of complex developments that build on its three major predecessors: Darwinism, Neo-Darwinism, and the MS itself” [109] (p. 2748). This Extended Synthesis represents a multifaceted research program expanding the conceptual and theoretical boundaries of evolutionary theory [110].
If EvoDevo has been defined as a synthesis of evolutionary and developmental biology, that it is more than “[…] developmental biology grafted onto evolutionary biology […]” [27] (p. 214) or a modern synthesis plus EvoDevo [111] (p. 190), so, what is the role or status of EvoDevo within the general framework of evolutionary theory?
In 2014, an article was published in which several of biologists and philosophers of biology called for a rethinking of evolutionary theory (the Reformists) [112] contrasted others who did not consider it necessary (the Conservatives) [113]. The first group of scientists advocates of an Evolutionary Extended Synthesis (EES), while the second group has a perspective in continuity with MS, a Standard Evolutionary Theory (SET). The role played by EvoDevo within each framework is different.
EES represents an “alternative vision of evolution” organized around a broader framework with its structure, assumptions and predictions, which “[…] will shed new light on how evolution works” [112] (pp. 161–162). The Reformists present four core processes neglected by SET (developmental bias, plasticity, niche construction, extra-genetic inheritance), that represent the main weapons in what is “[…] a struggle for the very soul of the discipline” (evolutionary biology). The fundamental message that emerges from this skirmish is a notion of development as “[…] a direct cause of why and how adaptation and speciation occurs, and of the rates and patterns of evolutionary change” (p. 164), and also the view “[…] that variation is not random, that there is more to inheritance than genes, and that there are multiple routes to the fit between organisms and environments” (p. 164). Based on these assumptions, the EES represents “[…] a different framework for understanding evolution […]” [114] (p. 3) or “[…] a comprehensive new synthesis” [115] (p. 8).
Meanwhile, on the Conservatives side, all is well: Adaptation and speciation are “[…] two of the most fundamental evolutionary processes”; DNA is “[…] the material basis for heredity and trait variation”; the putatively neglected processes were known since Darwin, and “[…] are already well integrated into evolutionary biology” [113] (p. 163). Then, more than a rethinking of evolutionary theory, the conservatives are more willing to a modest expansion covering the supposed neglected phenomena. However, the central tenets of evolutionary biology remain valid [113].
Futuyma [47] further discusses these aspects. The SET has already expanded its contents to include “[…] transposable elements, exon shuffling and chimeric genes, gene duplication and gene families, whole-genome duplication, de novo genes, gene regulatory networks, intragenomic conflict, kin selection, multilevel selection, phenotypic plasticity, maternal effects, morphological integration, evolvability, coevolution and more […]” [47] (p. 2). This expresses the inclusive character of the ES, which adopted its heuristics to include and expand the range of phenomena it was able to explain. More interesting, according to Futuyma, EvoDevo “[…] described by some adherents to an EES is oddly different from the research literature that has made the most substantial progress in evolutionary developmental biology” [47] (p. 8). So, Futuyma endorses the hoxological approaches [Carroll, Davidson], stating that “mechanistic understanding of gene action, of regulatory circuits, of the conservation of elements in the “genetic toolkit,” and their association with different downstream genes are rapidly deepening our understanding of evolutionary changes in form”[116] (p. 54). According to this, EvoDevo/Hoxology implement and complete the ST with mechanical causes able to link macroevolution to microevolution.
On the Reformist side, the theoretical tenets that characterize Extended Evolutionary Synthesis [114] are organized around a structure and a series of assumptions and predictions. In this perspective, EvoDevo plays a key role in shaping or extending the Synthesis [109]. EvoDevo not only “[…] provides a causal-mechanistic understanding of evolution […]” (p. 3) involving concepts such as developmental bias, evolvability, phenotypic plasticity, or facilitated variation; but also affects the overall structure of the synthesis, with a hierarchical, constructive, decentered (from the genetic level), and causally complex perspective on development and evolution [115].
In the clash between these two paradigms, Evodevo assumes an ambiguous position: One side refers to EvoDevo in a narrow sense, as Hoxology; the other one takes a broader perspective.
On the Synthesis and Expansion of Theories
We can say that EvoDevo, depending on the framework we adopt, is (i) a synthesis (desired, but not yet achieved) between developmental and evolutionary biology; (ii) a piece of a more complete synthesis, based on the MS; (iii) a leading (but ancillary, in respect to the overall theory) component in a new Extended Evolutionary Synthesis. We believe there is the risk that “[…] the boundaries of evolutionary developmental biology are becoming blurred and harder to demarcate, and so its goals and questions” [117] (p. 178).
About the term synthesis we can move from the following sketch [118]: “(i) a synthesis is a unification of originally disparate scientific structures (models, sets of models, theories, or even disciplines), and (ii) in the synthesized structure there is epistemic parity between the structure so unified” (p. 1217).
We can further distinguish between disciplinary integration, disciplinary synthesis, conceptual integration, and conceptual synthesis [119]. We have a disciplinary integration when two disciplines are compared, and “the result is one disciplinary structure that integrates two previously distinct disciplines” in which “[…] the individuality of the original parts is lost or effaced” [119] (pp. 311–312). A conceptual synthesis is when “[…] the significance of concepts in one discipline is evaluated for another (and vice versa) […]” with the “[…] blending of one or more parts to produce a new entity where the individuality of the original parts is not dissolved, though potentially transformed” [119] (pp. 311–312). In the meantime, a disciplinary synthesis “[…] produces a new discipline without dissolving those from which it was synthesized[…]” [119] (p. 312). Finally, a conceptual integration “[…] refers to how more than one concept can be merged into a single new concept for various purposes” [119] (p. 312). EvoDevo has been conceived, perhaps too rigidly, as a disciplinary synthesis.
Another useful distinction is made by James Griesemer [120], who distinguishes between different modes in which we can conceive a synthesis (in particular, the EES). The first mode is Theory Extension, which is a conceptual theory-revision or theory-building that “[…] works by building out from (and maybe modifying) a core theory to incorporate new theoretical principles (perhaps from another theory)” [120] (p. 323). The second mode is Domain Expansion, which consists of adding new phenomena “[…] under the umbrella of a theory and guidance of its perspective” [120] (p. 324). Lastly, Practice Integration consists of incorporating “[…]a practice into a workflow from other lines of work, specialties, disciplines, possibly changing what work is produced or changing interpretations” [120] (p. 324). Obviously, these three types of synthesis can also be combined with each other, but what is important in a process of theory extension, such as EES, is that “[…] the mode (addition, subtraction, modification, recombination) and target (concept, principle, model, perspective) must be specified” [120] (p. 323).
Let’s go back to EvoDevo and the two evolutionary frameworks. EvoDevo/Hoxology represents mainly an instance of Practice Integration, because it adds to MS/SET new (old) conceptual tools and empirical methodologies, i.e., the possibility to describe the evolution and development of morphologies through concepts like homology, novelty, body plans, etc., and explain experimentally the processes of construction of forms with the tools of developmental genetics. From a theoretical point of view, it is not required a substantial modification of Hoxology and SET, rather a gradual process of accommodation and conceptual synthesis, for example, with population thinking.
On the contrary, the EES represents at the same time a domain extension, since new phenomena and mechanisms are added to explain the processes affecting evolution and development, and a theory extension, because it requires a conceptual revision of how it conceives evolutionary theory. However, it does not represent a revolution of the old conceptual structure of MS, but an alternative research program [117] (p. 173).
In conclusion, EvoDevo conceived as a synthesis could represent a radical rethinking of the evolutionary theory [121], a kind of theory-revision/theory-building. But there are several conceptual difficulties. One of these concerns the pluralist framework that characterizes EvoDevo, and which represents an obstacle to conceptual and disciplinary synthesis. Another conceptual difficulty concerns the nature of development itself, and the possibility to establish a theory of development [122].
We agree with [24], that the main challenge of EvoDevo is the integration/synthesis between the various approaches, perspectives, or accounts presented above. The greatest contribution of EvoDevo will not consist in an expansion of the empirical basis, but in a new theoretical framework able to coherently hold together a variety of processes, mechanisms, phenomena, levels of description, phenomenology, and ontologies [123]. So “[…]a detailed epistemological analysis, […] of these programs and their underlying assumptions is therefore crucial for the future of evo-devo as a synthetic enterprise” [24] (p. 359).
What is needed is a conceptual synthesis able ”[…] to provide a conceptual foundation for different research programs that will ultimately explore the fuzzy edges and areas beyond the core of the synthesis, then we advocate an open conception of synthesis rather than a closed view based on integration of existing paradigms” [24] (p. 359). We think that this synthesis must be based on a theory that takes development seriously [3]. The problem is that we don’t “[…] have a sense of a general theory or principle of what development is beyond the slogans of decades or centuries past” [120] (p. 321).
4. Conclusions: Towards a Theory of What?
Although EvoDevo has been defined as “[…] evolution’s influence on development and development’s influence on evolution” [32] (p.9), little attention has been devoted to the formulation of a theory of development that could be integrated within the EvoDevo Synthesis. We agree with [124] “[…] that current skepticism partly results from a failure to articulate evo-devo’s conceptual foundation properly” (p. 100), and in particular its developmental foundations. As stated by [31], “compared to evolutionary biology, developmental biology has much less elaborated theoretical foundations to the extent that its subject, development is rarely defined, and usually delimited in purely operational terms […]” (p. 3). From this follows that “[…] a revised (more flexible and more comprehensive) concept of development will require a strong revisitation of evo-devo’s research agenda” [31] (p. 3).
The real innovative aspect of EvoDevo is precisely its focus on development. Without an adequate theory of development, at best, EvoDevo “shapes the extended synthesis” [11] as in EES, at worst, it can be reduced to a more articulated version of developmental genetics. In the latter case, the EvoDevo, going from the main antagonist of the MS, is likely to become its greatest ally. In the former case, without a theory of development, EvoDevo would not exploit its full potential.
What does it mean to develop a theory of development? Elaborating a complete and comprehensive theory of development is like packing a suitcase for a trip: The clothes are too many, the suitcase is too small, and something will remain out. Moreover, a theory of development faces several problematic aspects [122,125,126].
The first problem is a definitory one. Every definition of development leaves something out [127,128]: Some processes are often overlooked (regeneration); some definitions are biased (adultocentric perspective); some other definitions are too restricted (excluding unicellular organisms), and so on.
However, we believe that several aspects may be useful in the elaboration of a theory of development, in particular, and of a more comprehensive and complete EvoDevo Synthesis, in general.
The first aspect concerns the boundaries, both temporal and spatial, of development. From a spatial point of view, development involves multiple resources distributed on multiple levels of organization, as we have seen in the various accounts of EvoDevo. From a temporal point of view, development does not end with entering the adult stage [129,130]. Furthermore, development can go further than the boundaries of generations involving the influences (physiological, behavioral, etc.) of one generation over another (parental effects) [130]. Development is also a scaffolded process, in which several factors act as informational resources in the developmental processes (holobiont scaffolding) [131] (also cultural and cognitive scaffolding) [132].
A second aspect is represented by a broader understanding of the processes and mechanisms affecting the genetic level [133]. The discovery of Hox genes and of the genetic regulatory systems represented a milestone for EvoDevo, and the importance of such mechanisms in development is beyond question. However, it is one thing to acknowledge the importance of a discovery and of a mechanistic explanation, it is another to attribute exclusive causal powers to such mechanisms; especially in the light of a post-genomic conception of the gene [134]. The gene has become a complex entity [135], and the genome, far from being a simple repository of developmental and evolutionary information, is conceived as a Read-Write data storage system [136]. So, genes and genomes must be conceived of in a more dynamic and reactive way, not as exclusive protagonists of development, but as main players always in interaction with their context [137,138].
It has been observed that EvoDevo “[…] by itself cannot represent an alternative to the classical paradigm because it has no independent theory of heredity and population dynamics” [139] (p.276). However, it is possible to respond to both observations. As for the absence of an independent theory of inheritance within the EvoDevo, we believe it is a false problem for two reasons. The first reason is quite trivial: Why should there be a need for an independent theory of heredity? The second reason is that the available theories of heredity integrate relatively easily with an appropriate development theory in which different resources (genetics, epigenetics, behavioral, cultural) are involved in development processes [140,141]. Once again, what is needed seems to be greater theoretical integration between the already available perspectives.
As for a theory that takes into account population dynamics, we believe that this is a serious challenge for the EvoDevo. The core issue is that the concept of genes used in population genetics and in MS differs substantially from the concept of genes used in Developmental Synthesis [142]. The ontologies to which the two frameworks refer are profoundly different. Lenny Moss distinguishes between Gene-P and Gene-D, which captures well the problematic nature of this situation [143]. Gene-P is a preformationist conception of genes, in which starting from the genetic level, it is possible to predict the phenotypic outcomes—according to the tradition of genetic transmission [143]. While Gene-D is the developmental gene characterized by a surprising complexity both in its molecular structure and in its interactions with other genes (genetic networks). Overcoming the contrast between these ontologies is the main challenge for the future of EvoDevo [18].
In conclusion, we would like to summarize some issues that have emerged during this discussion and suggest some tasks for the future. The typology of theoretical perspectives is a useful starting point that allows us to consider various approaches to the study of phenomena affecting Development and Evolution. One task for the future is to integrate these perspectives into each other so that we can lay the foundations for subsequent theoretical synthesis. This theoretical synthesis, whose ultimate aim is to explain how Development, Evolution, and Inheritance are linked, requires that a theory of development be developed in the light of the variety of phenomena and approaches presented by the various theoretical perspectives. We want to argue that an EvoDevo resulting from a theoretical integration between the various theoretical perspectives we have presented and strongly focused on the formulation of a theory of development may represent a source of surprises for evolutionary theory.
Funding
This research received no external funding.
Acknowledgments
I would like to thank Alessandro Minelli for inviting me to contribute to the Special Issue “Renegotiating Disciplinary Fields in the Life Sciences” and for his suggestions on a draft version of this paper. I would also like to thank Daphne Liao and Teodora Todorovic for their editorial support. Finally, I would like to thank the three anonymous reviewers for their useful criticism and suggestions.
Conflicts of Interest
The author declares no conflict of interest.
References
- Toulmin, S. Human Understanding, Vol. I, General Introduction and Part I; Clarendon Press: Oxford, UK, 1972; ISBN 9780691071855. [Google Scholar]
- Darwin, C. The Origin of Species by Means of Natural Selection, or the Preservation of Favored Race in the Struggle of Life; Cambridge University Press: New York, NY, USA, 2009; ISBN 9781108005487. [Google Scholar]
- Robert, S.J. Embryology, Epigenesis, and Evolution: Taking Development Seriously; Cambridge University Press: Cambridge, UK, 2004; ISBN 9780521030861. [Google Scholar]
- Kirschner, M.W.; Gerhart, J.C. Facilitated Variation. In Evolution the Extended Synthesis; Müller, G.B., Pigliucci, M., Eds.; The MIT Press: Cambridge, MA, USA, 2010; pp. 253–280. ISBN 9780262315142. [Google Scholar]
- Dobzhansky, T. Genetics and the Origin of Species; Columbia University Press: New York, NY, USA, 1951; ISBN 1120819306. [Google Scholar]
- Dobzhansky, T.; Ayala, J.F.; Stebbins, G.L.; Valentine, J.W. Evoluciòn; Edicion Omega: Barcelona, Spain, 1983. [Google Scholar]
- Withfield, J. Postmodern Evolution? Nature 2008, 455, 281–284. [Google Scholar] [CrossRef] [Green Version]
- Hall, B.K. Unlocking the Black Box between Genotype and Phenotype: Cell condensation as Morphogenetic (modular) Units. Biol. Philos. 2003, 18, 219–247. [Google Scholar] [CrossRef]
- Waddington, C.H. The Evolution of an Evolutionist; Edinburgh University Press: Edinburgh, UK, 1975. [Google Scholar]
- Hall, B.K. Evo-devo or devo-evo—Does it matter? Evol. Dev. 2000, 2, 177–178. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, S.F. Evo-Devo, Devo-Evo, and Devgen-Popgen. Biol. Philos. 2003, 18, 347–352. [Google Scholar] [CrossRef]
- Müller, G.B. Evo-Devo: Extending the evolutionary synthesis. Nat. Rev. Genet. 2007, 8, 943–949. [Google Scholar] [CrossRef]
- Müller, G.B.; Wagner, G.P. Novelty in Evolution: Restructuring the concept. Annu. Rev. Ecol. Syst. 1991, 22, 229–256. [Google Scholar] [CrossRef]
- Müller, G.B.; Newman, S.A. The innovation triad: An EvoDevo agenda. J. Exp. Zool. 2005, 304, 487–503. [Google Scholar] [CrossRef]
- Müller, G.B. Homology: The evolution of morphological organization. In Origination of Organismal Form: Beyond the Gene in Developmental and Evolutionary Biology; Müller, G.B., Newman, S.A., Eds.; MIT Press: Cambridge, MA, USA, 2003; pp. 51–69. [Google Scholar]
- Minelli, A.; Fusco, G. Homology. In The Philosophy of Biology: A Companion for Educators; Kampourakis, K., Ed.; Springer: Dordrecht, The Netherlands, 2013; pp. 289–322. ISBN 9789400765375. [Google Scholar]
- Carroll, S.B. Endless Forms Most Beautiful: The New Science of EvoDEvo and the Making of the Animal Kingdom; Norton: New York, NY, USA, 2005; ISBN 9780393327793. [Google Scholar]
- Wagner, G. Homology, Genes, and Evolutionary Innovation; Princeton University Press: Princeton, NJ, USA, 2014; ISBN 9780691180670. [Google Scholar]
- Kirschner, M.; Gerhart, J. Evolvability. Proc. Natl. Acad. Sci. USA 1998, 95, 8420–8427. [Google Scholar] [CrossRef] [Green Version]
- Minelli, A. Evolvability and Its Evolvability. In Challenging the Modern Synthesis: Adapatation, Development, and Inheritance; Huneman, P., Walsh, D., Eds.; Oxford University Press: Oxford, UK, 2017; pp. 211–238. ISBN 978-0199377176. [Google Scholar]
- Arthur, W. The interaction between developmental bias and natural selection: From centipede segments to a general hypothesis. Heredity 2002, 89, 239–246. [Google Scholar] [CrossRef] [PubMed]
- Kuratani, S. Modularity, comparative embryology and evo-devo: Developmental dissection of evolving body plans. Dev. Biol. 2009, 332, 61–69. [Google Scholar] [CrossRef]
- Robert, J.S. Evo-Devo. In The Oxford Handbook of Philosophy of Biology; Ruse, M., Ed.; Oxford University Press: Oxford, UK, 2014; pp. 291–309. ISBN 9780199737260. [Google Scholar]
- Laubichler, M.D. Evolutionary Developmental Biology. In The Cambridge Companion to the Philosophy of Biology; Hull, D., Ruse, M., Eds.; Cambridge University Press: Cambirdge, UK, 2008; pp. 342–360. ISBN 9781139001588. [Google Scholar]
- Love, A.C. Evolutionary Developmental Biology: Philosophical Issues. In Handbook of Evolutionary Thinking in the Sciences; Heams, T., Ed.; Springer: Berlin/Heidelberg, Germany, 2015; pp. 265–283. [Google Scholar]
- Müller, G.B. Evolutionary Developmental Biology. In Handbook of Evolution, Vol. 2: The Evolution of Living Systems (Including Hominids); Wuketis, F.M., Ayala, F.J., Eds.; Wiley-VCH: Hoboken, NJ, USA, 2005; pp. 87–115. ISBN 9783527308385. [Google Scholar]
- Minelli, A. Evolutionary Developmental Biology does not Offer a Significant Challenge to the Neo-Darwinian Paradigm. In Contemporary Debates in Philosophy of Biology; Ayala, F.J., Arp, R., Eds.; Wiley-Blackwell: Hoboken, NJ, USA, 2010; pp. 213–226. ISBN 9781405159982. [Google Scholar]
- Laubichler, M.D. Evolutionary Developmental Biology Offers a Significant Challenge to the Neo-Darwinian Paradigm. In Contemporary Debates in Philosophy of Biology; Ayala, F.J., Arp, R., Eds.; Wiley-Blackwell: Hoboken, NJ, USA, 2010; pp. 198–212. ISBN 9781405159982. [Google Scholar]
- Carroll, R.L. Towards a new evolutionary synthesis. Tree 2000, 15, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Raff, R.A. Evo-Devo: The evolution of a new discipline. Nat. Rev. Gen. 2000, 1, 74–79. [Google Scholar] [CrossRef]
- Minelli, A. Grand challenges in evolutionary developmental biology. Front. Ecol. Evol. 2015, 2, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Müller, G.B. Evo-devo as a discipline. In Evolving Pathways: Key themes in Evolutionary Developmental Biology; Minelli, A., Fusco, G., Eds.; Cambridge University Press: Cambridge, UK, 2008; pp. 5–30. ISBN 9781849722698. [Google Scholar]
- Hall, B.K.; Olson, W.M. Keywords and Concepts in Evolutionary Developmental Biology; Harvard University Press: Cambridge, MA, USA, 2003; ISBN 9781849722698. [Google Scholar]
- Müller, G.B. Six Memos for Evo-Devo. In From Embryology to EvoDevo: A History of Developmental Evolution; Laubichler, M.D., Maienschein, J., Eds.; The MIT Press: Cambridge, UK, 2007; pp. 499–524. [Google Scholar]
- Minelli, A. Fusco, Evolving Pathways: Key themes in Evolutionary Developmental Biology; Cambridge University Press: Cambridge, UK, 2008; ISBN 9781107405455. [Google Scholar]
- Burian, R.M. On conflicts between genetic and developmental viewpoints—And their attempted resolution in molecular biology. In Structures and Norms in Science; Dalla Chiara, M.L., Doets, K., Mundici, D., van Benthem, J., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1997; pp. 243–264. ISBN 978-0-7923-4384-4. [Google Scholar]
- Laubichler, M.D.; Wagner, G.P. 2004—Introduction to the papers of the 2001 Kowalevsky Medal Winner Symposium. J. Exp. Zool. Mol. Dev. Evol. 2004, 302, 1–4. [Google Scholar] [CrossRef]
- McGinnis, W.; Levine, M.S.; Hafen, E.; Kuroiwa, A.; Gehring, W.J. A Conserved DNA sequence in homeotic genes of the Drosophila Antennapedia and bithorax complexes. Nature 1984, 308, 428–433. [Google Scholar] [CrossRef]
- Wakimoto, B.T.; Kaufman, T.C. Analysis of Larval Segmentation in Lethal Genotypes Associated with the Antennapedia Gene Complex in Drosophila Melanogaster. Dev. Biol. 1981, 81, 51–64. [Google Scholar] [CrossRef]
- Carroll, S.B. Endless Forms: The Evolution of Gene Regulation and Morphological Diversity. Cell 2000, 101, 577–580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slack, J. A Rosetta Stone for pattern formation in animals? Nature 1984, 310, 364–365. [Google Scholar] [CrossRef]
- Gould, S.J. The Structure of Evolutionary Theory; The Belknap Press of Harvard University Press: Cambridge, MA, USA, 2002; ISBN 9780674006133. [Google Scholar]
- Davidson, E.H. The Regulatory Genome: Gene Regulatory Networks in Development and Evolution; Academic Press: New York, NY, USA, 2006; ISBN 9781280641329. [Google Scholar]
- Carroll, S.B. Evo-Devo and an Expanding Evolutionary Synthesis: A Genetic Theory of Morphological Evolution. Cell 2008, 134, 25–36. [Google Scholar] [CrossRef] [Green Version]
- Carroll, S.B. 2001 The Big Picture. Nature 2001, 409, 669. [Google Scholar] [CrossRef] [PubMed]
- Sterelny, K. Development, Evolution, and Adaptation. Philos. Sci. 2000, 67, 369–387. [Google Scholar] [CrossRef]
- Futuyma, D.J. Evolutionary biology today and the call for an extended synthesis. Interface Focus 2017, 7, 20160145. [Google Scholar] [CrossRef]
- Hoekstra, H.E.; Coyne, J.A. The Locus of Evolution: EvoDevo and the genetics of adaptations. Evolution 2007, 61, 995–1016. [Google Scholar] [CrossRef] [PubMed]
- Nikla, K.J. Thinking outside the Hox. Biol. Theory 2006, 1, 128–129. [Google Scholar] [CrossRef]
- Medina, M.L. Two “EvoDevos”. Biol. Theory 2010, 5, 7–11. [Google Scholar] [CrossRef]
- Amundson, R. The Changing Role of the Embryo in Evolutionary Thought; Cambridge University Press: Cambridge, UK, 2005; ISBN 9780521703970. [Google Scholar]
- Gehring, W.J.; Hiromi, Y. Homeotic Genes and the Homeobox. Annu. Rev. Genet. 1986, 20, 147–173. [Google Scholar] [CrossRef]
- Gehring, W.J. Master Control Genes in Development and Evolution: The Homeobox Story; Yale University Press: New Haven, CT, USA, 1998; ISBN 9780300074093. [Google Scholar]
- Forgacs, G.; Newman, S.A. Biological Physics of the Developing Embryo; Cambridge University Press: Cambridge, UK, 2005; ISBN 9780521783378. [Google Scholar]
- Kauffman, S. The Origins of Order: Self-Organizating and Selection in Evolution; Oxford University Press: Oxford, UK, 1993; ISBN 9780195058116. [Google Scholar]
- Newman, S.A.; Muller, G.B. Epigenetic mechanisms of character origination. J. Exp. Zool. B Mol. Dev. Evol. 2000, 288, 304–317. [Google Scholar] [CrossRef]
- Callebaut, W.; Muller, G.B.; Newman, S.A. The Organismic Systems Approach: Evo-Devo and the Streamlining of the Naturalistic Agenda. In Integrating Evolution and Development: From Theory to Practice; Samson, R., Brandon, R., Eds.; The MIT Press: Cambridge, MA, USA, 2007; ISBN 9780262195607. [Google Scholar]
- Müller, G.B.; Newman, S. Origination of Organismal Form: The Forgotten Cause in Evolutionary Theory. In Origination of Organismal Form: Beyond the Gene in Developmental and Evolutionary Biology; Müller, G.B., Newman, S., Eds.; MIT Press: Cambridge, MA, USA, 2003; pp. 3–10. ISBN 9780262280327. [Google Scholar]
- Müller, G.B.; Newman, S. Generation, Integration, Autonomy: Three Steps in the Evolution of Homology. Homology (Novartis Foundation Symposium 222); Wiley: Chichester, UK, 1999; pp. 65–79. ISBN 9780470515662. [Google Scholar]
- Goodwin, B. Evolution and the generative order. In Theoretical Biology: Epigenetic and Evolutionary Order from Complex Systems; Goodwin, B., Saunders, P.T., Eds.; Edinburgh University Press: Edinburgh, UK, 1989; pp. 89–100. ISBN 9780801845192. [Google Scholar]
- Webster, G.; Goodwin, B. Form and Transformation: Generative and Relational Principles in Biology; Cambridge University Press: Cambridge, UK, 1996; ISBN 9780521207430. [Google Scholar]
- Ho, M.W.; Saunders, P.T. Beyond Neo-darwinism: And epigenetic approach to evolution. J. Theor. Biol. 1979, 78, 573–591. [Google Scholar] [CrossRef]
- Callebaut, W. Self-organization and optimization: Conflicting or complementary approaches? In Evolutionary Systems; Van de Vijver, G., Salthe, S.N., Delpos, M., Eds.; Kluwer: Dordrecht, The Netherlands, 1998; pp. 79–100. [Google Scholar]
- Weber, B.H.; Depew, P.J. Natural selection and self-organization. Biol. Philos. 1996, 11, 33–65. [Google Scholar] [CrossRef]
- Fontana, W. Modelling EvoDevo with RNA. BioEssays 2002, 24, 1164–1177. [Google Scholar] [CrossRef]
- Fontana, W. The topology of the possible. In Understanding Change: Models, Methodologies and Metaphors; Wimmer, A., Kössler, R., Eds.; Palgrave Macmillan: London, UK, 2006; pp. 67–84. ISBN 9780230524644. [Google Scholar]
- Lewontin, R.C. The Docrtine of DNA: Biology as Ideology; Penguin: New York, NY, USA, 1993; ISBN 9780140232196. [Google Scholar]
- Rose, S.; Lewontin, R.C.; Konin, L. Not in Our Genes: Biology, Ideology and Human Nature; Penguin Books: Harmondsworth, UK, 1990; ISBN 9780394508177. [Google Scholar]
- Lewontin, R.C. The Triple Helix: Gene, Organism, and Environment; Harvard University Press: Cambridge, MA, USA, 2000; ISBN 9780674006775. [Google Scholar]
- Levins, R.; Lewontin, R.C. A program for biology. Biol. Theory 2006, 1, 333–335. [Google Scholar] [CrossRef]
- Kohl, P.; Crampin, E.J.; Quinn, T.A.; Noble, D. Systems Biology: An Approach. Clin. Pharmacol. Ther. 2010, 88, 25–33. [Google Scholar] [CrossRef]
- Kitano, H. Systems Biology: Toward System-Level Understanding of Biological Systems. In Foundations of Systems Biology; Kitano, H., Ed.; The MIT Press: Cambridge, MA, USA, 2001; pp. 1–36. ISBN 9780262277204. [Google Scholar]
- Kirschner, M.W. The Meaning of Systems Biology. Cell 2005, 121, 503–504. [Google Scholar] [CrossRef] [Green Version]
- Krohs, U.; Callebaut, W. Data without models merging with models without data. In Systems Biology: Philosophical Foundations; Boogerd, F.C., Bruggeman, F.J., Hofmeyer, S.J.-H., Westerhoff, H.V., Eds.; Elsevier: Amsterdam, The Netherlands, 2007; pp. 181–213. ISBN 9780444520852. [Google Scholar]
- Noble, D. The Aims of Systems Biology: Between Molecules and Organisms. Pharmacopsychiatry 2011, 44 (Suppl. 1), 9–14. [Google Scholar] [CrossRef]
- Kitano, H. Systems Biology: A Brief Overview. Science 2002, 295. [Google Scholar] [CrossRef] [Green Version]
- Newman, S.A. The Fall and Rise of Systems Biology: Recovering from a Halfcentury Gene Hing; GeneWatch: London, UK, 2003; Volume 16, pp. 8–12. [Google Scholar]
- Noble, D. A theory of biological relativity: No privileged level of causation. Interface Focus 2012, 2, 55–64. [Google Scholar] [CrossRef] [Green Version]
- Noble, D. The Music of Life: Biology beyond the Genome; Oxford University Press: Oxford, UK, 2006; ISBN 9780199295739. [Google Scholar]
- Piaget, J. Behavior and Evolution; Pantheon Books: New York, NY, USA, 1978; ISBN 9780394735887. [Google Scholar]
- Weiss, P. Cellular Dynamics. Rev. Mod. Phys. 1959, 31, 11–20. [Google Scholar] [CrossRef]
- Griesemer, J.R. Reproduction and the reduction of genetics. In The Concept of the Gene in Development and Evolution: Historical and Epistemological Perspectives; Beurton, P.J., Falk, R., Rheinberger, H.J., Eds.; Cambridge University Press: New York, NY, USA, 2000; pp. 240–285. ISBN 9780521060240. [Google Scholar]
- Griesemer, J. The units of evolutionary transitions. Selection 2000, 1, 67–80. [Google Scholar] [CrossRef] [Green Version]
- Gottlieb, G. Probabilistic Epigenesis. Dev. Sci. 2007, 10, 1–11. [Google Scholar] [CrossRef]
- Griesemer, J.R. Reproduction and scaffolded developmental processes. In Towards a Theory of Development; Minelli, A., Pradeu, T., Eds.; Oxford University Press: Oxford, UK, 2014; pp. 182–202. ISBN 9780199671434. [Google Scholar]
- Wimsatt, W.C.; Griesemer, J.R. Reproducing entrenchments to scaffold culture. In Integrating Evolution and Development: From Theory to Practice; Sanson, R., Brandon, R.N., Eds.; The MIT Press: Cambridge, MA, USA, 2007; pp. 227–323. ISBN 9780262195607. [Google Scholar]
- Oyama, S.; Griffiths, P.E.; Gray, R.D. Introduction: What is developmental Systems Theory? In Cycles of Contingency: Developmental Systems and Evolution; Oyama, S., Griffiths, P.E., Gray, R.D., Eds.; The MIT Press: Cambridge, MA, USA, 2001; ISBN 97802626506321-12. [Google Scholar]
- Oyama, S. The Ontogeny of Information: Developmental Systems and Evolution; Duke University Press: Durham, NC, USA, 2000; ISBN 9780822324317. [Google Scholar]
- Robert, J.S.; Hall, B.K.; Olson, W.M. Bridging the gap between developmental systems theory and evolutionary developmental biology. BioEssays 2001, 23, 954–962. [Google Scholar] [CrossRef]
- Caporael, L.R. Evolution, Groups, and Scaffolded Mings. In Developing Scaffolds in Evolution, Culture, and Cognition; Caporael, L.R., Griesemer, J.R., Wimsatt, W.C., Eds.; The MIT Press: Cambridge, MA, USA, 2014; pp. 57–76. ISBN 9781461952367. [Google Scholar]
- Caporael, L.R. The Evolution of Truly Social Cognition: The Core Configurations Model. Pers. Soc. Psychol. Rev. 1997, 1, 276–298. [Google Scholar] [CrossRef]
- West-Eberhard, M.-J. Developmental Plasticity and Evolution; Oxford University Press: Oxford, UK, 2003. [Google Scholar]
- West-Eberhard, M.-J. Developmental Plasticity and the Origin of Species Differences. Proc. Natl. Acad. Sci. USA 2005, 102 (Suppl. 1), 6543–6549. [Google Scholar] [CrossRef] [Green Version]
- Kupiec, K.J. The Origin of Individuals; Worlds Scientific Publishing: Singapore, 2009; ISBN 9789812704993. [Google Scholar]
- Kupiec, K.J. L’Ontophylogenese: Evolution des Especes et Developpement de L’individu; Editions Quæ: Versailles, France, 2012; ISBN 9782759217861. [Google Scholar]
- Gottlieb, G. Individual Development and Evolution: The Genesis of Novel Behavior; Oxford University Press: New York, NY, USA, 1992; ISBN 9780415648486. [Google Scholar]
- Abouheif, E.; Favé, M.-J.; Ibarraran-Viniegra, A.S.; Lesoway, M.P.; Rafiqi, A.M.; Rajakumar, R. Eco-Evo-Devo: The times has come. In Ecological Genomics: Ecology and the Evolution of Genes and Genomes; Laundry, C.R., Aubin-North, N., Eds.; Advances in Experimental Medicine and Biology 781; Springer: Dordrecht, The Netherlands, 2014; pp. 107–125. [Google Scholar]
- Gilbert, S.F.; Epel, D. Ecological Developmental Biology: The Environmental Regulation of Development, Health, and Evolution; Sinauer Associates: Sunderland, MA, USA, 2015; ISBN 9781605353449. [Google Scholar]
- Bateson, P.P.G. Behaviour, Development and Evolution; Cambridge University Press: Cambridge, UK, 2017; ISBN 9781783742486. [Google Scholar]
- Stotz, K. Extended evolutionary psychology: The importance of transgenerational developmental plasticity. Front. Psychol. 2014, 5, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Wilkins, A.S. The Evolution of Developmental Pathways; Sinauer Associates: Sunderland, MA, USA, 2002; ISBN 9780878939169. [Google Scholar]
- Griesemer, J. Development, Culture, and the Units of Inheritance. Philos. Sci. 2000, 348–368. [Google Scholar] [CrossRef] [Green Version]
- Giere, R.N. Scientific Perspectivism; The University of Chicago Press: Chicago, IL, USA, 2006; ISBN 9780226292137. [Google Scholar]
- Robert, J.S. How developmental is evolutionary developmental biology? Biol. Philos. 2002, 17, 591–611. [Google Scholar] [CrossRef]
- Maynard-Smith, J. Shaping Life: Genes, Embryos and Evolution; Yale University Press: New Haven, CT, USA, 1998; ISBN 9780300080223. [Google Scholar]
- Nuno de la Rosa, L. Computing the Extended Synthesis: Mapping the Dynamics and Conceptual Structure of the Evolvability Research Front. J. Exp. Zool. Mol. Dev. Evol. 2017, 328, 395–396. [Google Scholar] [CrossRef]
- Winther, R.G. Evo-devo as a Trading zone. In Conceptual Change in Biology, Boston Studies in the Philosophy and History of Science; Love, A.C., Ed.; Springer: Dordrecht, The Netherlands, 2015; pp. 459–482. ISBN 9789401794121. [Google Scholar]
- Pigliucci, M. Do we need an extended evolutionary synthesis? Evolution 2007, 61, 2743–2749. [Google Scholar] [CrossRef] [Green Version]
- Pigliucci, M.; Muller, G.B. Evolution: The Extended Synthesis; The MIT Press: Cambridge, MA, USA, 2010; ISBN 9780262315142. [Google Scholar]
- Hall, B.K. Evolutionary Developmental Biology (Evo-Devo): Past, Present, and Future. Evol. Educ. Outreach 2012, 5, 184–193. [Google Scholar] [CrossRef] [Green Version]
- Laland, K.N.; Uller, T.; Feldman, M.W.; Sterelny, K.; Müller, G.B.; Moczek, A.P.; Jablonka, E.; Odling-Smee, J.; Wray, G.A.; Hoekstra, H.E.; et al. Does evolutionary theory need a rethink? Nat. Cell Biol. 2014, 514, 161–164. [Google Scholar] [CrossRef] [Green Version]
- Wray, G.A.; Hoekstra, H.E.; Futuyma, D.J.; Lensky, R.E.; Mackay, T.F.C.; Schluter, D.; Strassmann, J.E. Does evolutionary theory need a rethink? No, all is well. Nature 2014, 514, 161–164. [Google Scholar] [CrossRef] [Green Version]
- Laland, K.N.; Uller, T.; Feldman, M.W.; Sterelny, K.; Müller, G.B.; Moczek, A.P.; Jablonka, E.; Odling-Smee, F.J. The extended evolutionary synthesis: Its structure, assumptions and predictions. Proc. R. Soc. B Boil. Sci. 2015, 282, 20151019. [Google Scholar] [CrossRef]
- Müller, G.B. EvoDevo Shapes the Extended Synthesis. Biol. Theory 2014, 9, 119–121. [Google Scholar] [CrossRef] [Green Version]
- Futuyma, D.J. Can Modern Evolutionary Theory Explain Macroevolution? In Macroevolution; Serrelli, E., Gontier, N., Eds.; Springer: Berlin/Heidelberg, Germany, 2015; pp. 29–85. ISBN 978-3-319-15044-4. [Google Scholar]
- Fàbregas-Tejeda, A.; Vergara-Silva, F. The emerging structure of the Extended Evolutionary Synthesis: Where does Evo-Devo fit in? Theory Biosci. 2018, 137, 169–184. [Google Scholar] [CrossRef] [Green Version]
- Sarkar, S. Evolutionary Theory in the 1920s: The Nature of the “Synthesis”. Philos. Sci. 2004, 71, 1215–1226. [Google Scholar] [CrossRef] [Green Version]
- Love, A.C. Evolutionary Morphology, Innovation, and the Synthesis of Evolutionary and Developmental Biology. Biol. Philos. 2003, 18, 309–345. [Google Scholar] [CrossRef]
- Griesemer, J. Towards a theory of Extended Developments. In Perspective on Evolutionary and Developmental Biology; Fusco, G., Ed.; Padova University Press: Padova, Italy, 2019; pp. 319–334. ISBN 978-88-6938-140-9. [Google Scholar]
- Craig, L.R. The So-Called Extended Synthesis and Population Genetics. Biol. Theory 2010, 5, 117–123. [Google Scholar] [CrossRef]
- Minelli, A.; Pradeu, T. (Eds.) Towards a Theory of Development; Oxford University Press: Oxford, UK, 2014; ISBN 9780199671434. [Google Scholar]
- Laubichler, M.D.; Prohaska, S.J.; Stadler, P.F. Toward a mechanistic explanation of phenotypic evolution: The need for a theory of theory integration. J. Exp. Zool. Part B Mol. Dev. Evol. 2018, 330, 5–14. [Google Scholar] [CrossRef] [PubMed]
- Jenner, R.A. EvoDevo’s identity: From model organisms to developmental types. In Evolving Pathways: Key Themes in Evolutionary Developmental Biology; Minelli, A., Fusco, G., Eds.; Cambridge University Press: Cambridge, UK, 2009; pp. 100–119. [Google Scholar]
- Minelli, A. Understanding Development; Cambridge University Press: Cambridge, UK, forthcoming.
- Pradeu, T.; Laplane, L.; Prévot, K.; Hoquet, T.; Reynaud, V.; Fusco, G.; Minelli, A.; Orgogozo, V.; Vervoort, M. Defining “Development”. Curr. Top. Dev. Biol. 2016, 117, 171–183. [Google Scholar] [CrossRef] [Green Version]
- Minelli, A. The Development of Animal Form; Cambridge University Press: Cambridge, UK, 2003; ISBN 9780521808514. [Google Scholar]
- Minelli, A. Forms of Becoming: The Evolutionary Biology of Development; Princeton University Press: Princeton, NJ, USA, 2009; ISBN 9780691135687. [Google Scholar]
- Minelli, A. Animal Development, an Open-Ended Segment of Life. Biol. Theory 2011, 6, 4–15. [Google Scholar] [CrossRef]
- Badyaev, A.V.; Uller, T. Parental effects in ecology and evolution: Mechanisms, processes and implications. Philos. Trans. R. Soc. B Biol. Sci. 2009, 364, 1169–1177. [Google Scholar] [CrossRef] [Green Version]
- Chiu, L.; Gilbert, S.F. The Birth of the Holobiont: Multi-species Birthing Through Mutual Scaffolding and Niche Construction. Biosemiotics 2015, 8, 191–210. [Google Scholar] [CrossRef]
- Caporael, L.R.; Griesemer, J.R.; Wimsatt, W.C. Developing Scaffold in Evolution, Culture and Cognition; The MIT Press: Cambridge, MA, USA, 2014; ISBN 9781461952367. [Google Scholar]
- Griffiths, P. Stotz, Genetics and Philosophy: An Introduction; Cambridge University Press: Cambridge, UK, 2013. [Google Scholar]
- Griffiths, P.E.; Stotz, K. Genes in the Postgenomic Era. Theor. Med. Bioeth. 2006, 27, 499–521. [Google Scholar] [CrossRef]
- Gerstein, M.B.; Bruce, C.; Rozowsky, J.S.; Zheng, D.; Du, J.; Korbel, J.O.; Emanuelsson, O.; Zhang, Z.D.; Weissman, S.; Snyder, M. What is a gene, post-ENCODE? History and updated definition. Genome Res. 2007, 17, 669–681. [Google Scholar] [CrossRef] [Green Version]
- Shapiro, J.A. How life changes itself: The Read–Write (RW) genome. Phys. Life Rev. 2013, 10, 287–323. [Google Scholar] [CrossRef]
- Gilbert, S.F. The Reactive Genome. In Origination of Organismal Form: Beyond the Gene in Developmental and Evolutionary Biology; Muller, G.B., Newman, S.A., Eds.; The MIT Press: Cambridge, MA, USA, 2003; pp. 87–101. ISBN 9780262280327. [Google Scholar]
- Lamm, E. The genome as a developmental organ. J. Physiol. 2014, 592, 2283–2293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muller, G.B.; Pigliucci, M. Extended Synthesis: Theory Expansion or Alternative? Biol. Theory 2010, 5, 275–276. [Google Scholar] [CrossRef]
- Jablonka, E.; Lamb, M.J. Evolution in Four Dimensions: Genetic, Epigenetic, Behavioral, and Symbolic Variation in the History of Life; The MIT Press: Cambridge, MA, USA, 2005; ISBN 9780262525848 0262525844. [Google Scholar]
- Bonduriansky, R.; Day, T. Extended Heredity: A New Understanding of Inheritance and Evolution; Princeton University Press: Princeton, NJ, USA, 2018; ISBN 9781400890156. [Google Scholar]
- Gilbert, S.F. Genes Classical and Genes Developmental: The different use genes in Evolutionary Synthesis. In The Concept of the Gene in Development and Evolution: Historical and Epistemological Perspectives; Falk, R., Rheinberger, H.-J., Eds.; Cambridge University Press: New York, NY, USA, 2000; pp. 178–192. ISBN 0521060249 9780521060240. [Google Scholar]
- Moss, L. What Genes Can’t Do; The MIT Press: Cambridge, MA, USA, 2003; ISBN 9780262632973. [Google Scholar]
- Falk, R. The Gene: A concept in tension. In The Concept of the Gene in Development and Evolution: Historical and Epistemological Perspectives; Falk, R., Rheinberger, H.-J., Eds.; Cambridge University Press: New York, NY, USA, 2000; pp. 178–192. ISBN 0521060249 9780521060240. [Google Scholar]
Figure 1.
Theoretical Continuum of the various EvoDevo perspectives.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Amato, S.I. EvoDevo: An Ongoing Revolution? Philosophies 2020, 5, 35. https://doi.org/10.3390/philosophies5040035
AMA Style
Amato SI. EvoDevo: An Ongoing Revolution? Philosophies. 2020; 5(4):35. https://doi.org/10.3390/philosophies5040035
Chicago/Turabian StyleAmato, Salvatore Ivan. 2020. "EvoDevo: An Ongoing Revolution?" Philosophies 5, no. 4: 35. https://doi.org/10.3390/philosophies5040035