
Abstract
The lower Miocene marine units of NW Iran (Hamedan–Miandoab area) were studied to establish a high-resolution biostratigraphy framework in the context of European standard biozonation (SBZ zonal scheme). The units are dominated by larger and small benthic foraminifera together with coralline algae and corals. Due to its position in the Tethyan Seaway between the Mediterranean and Indo-Pacific, the investigated area has high significance in palaeobiogeography. Seven species of benthic foraminifera were described, including Miogypsina globulina, Amphistegina bohdanowiczi, Elphidium crispum, Elphidium sp. 1, Borelis curdica, Austrotrillina asmariensis, and Peneroplis thomasi. The occurrence of Miogypsina globulina in the shallow-marine carbonates of the studied sections documents the Burdigalian SBZ 25 Zone. Austrotrillina asmariensis and Peneroplis thomasi occur with M. globulina. Simultaneously, their stratigraphical range is extended to the Burdigalian in the Middle East. Based on the discovery of lower Miocene deposits in the Hamedan–Miandoab area (previously mapped as Oligocene–Miocene units), the stratigraphy of northwest Central Iran is revised. The biostratigraphic results also present a more precise chronology for the marine transgression in the study area, initiating in the late early Miocene. The coralline algal assemblages and the abundance and diversity of larger benthic foraminifera indicate that shallow-marine Qom Formation got deposited in tropical to subtropical warm waters.
Similar content being viewed by others

Introduction
Northwestern Iran is characterized by well-exposed Mesozoic and Cenozoic marine strata (e.g., Sarfi and Yazdi-Moghadam 2016; Yazdi-Moghadam et al. 2017; Omidvar et al. 2018; Yazdi-Moghadam et al. 2018a, b). Oligo-Miocene shallow-marine carbonates often contain diverse assemblages of larger benthic foraminifera (LBF) that have been extensively used for biostratigraphic dating, palaeoenvironmental interpretation, and palaeobiogeography of shallow-marine sequences of the Western Tethys, the Middle East, and the Indo-Pacific region (e.g., Cushman 1939; Henson 1948; Drooger 1952; Bozorgnia 1966; Hansen and Lykke-Andersen 1976; Schaub 1981; Less 1991; Cahuzac and Poignant 1997; Vaziri-Moghaddam et al. 2006; Hottinger 2007; Özcan and Less 2009; Boukhary et al. 2010; Özcan et al. 2010; Sirel et al. 2013; Hakyemez et al. 2016; Saraswati et al. 2018). Associated with the other shallow-marine biota (e.g., echinoderms, pelecypods, gastropods, coralline algae), LBF characterize widespread platform carbonates of the Oligo-Miocene age which are known under different formational names from central and southern Iran, e.g., Asmari, Mishan, and Gachsaran formations (Zagros Mountains), or Qom Formation (Central Iran). Attempts for detailed stratigraphy, and palaeoenvironmental studies of these platform carbonates in Iran, have been made by several authors and are still in progress (e.g., Ehrenberg et al. 2007; Van Buchem et al. 2010; Shabafrooz et al. 2015; Allahkarampour Dill et al. 2018; Yazdi-Moghadam et al. 2018a, b; Basso et al. 2019; Akbar-Baskalayeh et al. 2020). The present study makes an important contribution to this study process and reports the occurrence of hyaline and porcelaneous LBF recovered from three outcrop sections in the Hamedan–Miandoab area, NW Iran. The stratigraphic and taxonomic implications of the studied associations are of importance for (1) establishment of a biostratigraphic framework within the context of European standard biozonal scheme (SBZ) and (2) dating the initiation of shallow-marine conditions in the study area.
Geological and stratigraphic setting
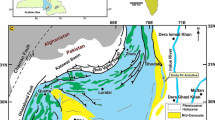
The Cenozoic geology of Iran is characterized by an array of structural units including Zagros fold and thrust belt (ZFTB), Sanandaj–Sirjan Zone (SSZ), Uromia–Dokhtar Magmatic Arc (UDMA), Central Iran, Alborz, Koppeh-Dagh, and eastern Iran (e.g., Stöcklin 1968; Stöcklin and Nabavi 1973; Berberian 1974; Berberian and King 1981) (Fig. 1a). The Lower Red, the Qom and the Upper Red Formations are superbly exposed and usually very thick in Central Iran. In contrast, the strata of the Qom Formation have locally deposited, with less thickness, in UDMA and SSZ, and the Lower Red and Upper Red Formations are absent in many areas of these two structural units. As a part of the Alpine–Himalayan orogenic belt, Central Iran, UDMA, and SSZ exhibit complex structural features that are the result of long structural history from Palaeozoic time up to the present (Letouzey and Rudkiewicz 2005; Morley et al. 2009). Before Permian rifting, Central Iran and Zagros formed a single structural unit. During the Permian–Triassic time boundary, Central Iran was separated from the Zagros Mountains in the south, by an oceanic basin (Neo-Tethys). Following the Late Cretaceous closure of the Neo-Tethys, the northward migration of the Arabian plate continued until the continental collision with the Eurasian/Iranian plate at the Oligocene–Miocene boundary. The occurrence of the Oligo-Miocene marine deposits in Central Iran, UDMA, and SSZ is the result of the subduction of Neo-Tethys oceanic crust below the southern margin of Central Iran, which led to opening a back-arc basin (Central Iran) and a fore-arc basin (SSZ) separated by a magmatic arc (UDMA). In fact, the Qom Formation can be considered as part of a large depositional system of shallow-marine platform carbonates and deep-marine marlstones that characterized large parts of the present-day Iran during the Oligo-Miocene times (e.g., Qom, Asmari, Pabdeh, and Mishan formations). The Oligo-Miocene marine strata of Central Iran, UDMA, SSZ, and ZFTB form important components of the late Paleogene–early Neogene Tethyan warm to temperate marine depositional belts. From the aforementioned data, it becomes clear that the Oligo-Miocene sedimentary sequences of central and southern Iran bear important information for the geodynamic history of Iran and the Middle East.

Geological and geographical setting. a Simplified structural map of Iran showing the main tectonic subdivisions (modified from Agard et al. 2011). The white stars indicate the location of the sections studied. a Goli section, b Qaraie section, c Qabakh Tappeh section. b–d Road maps showing the position of the studied sections. (MZT Main Zagros Thrust, SSZ Sanandaj–Sirjan Zone, ZFTB Zagros Fold Thrust Belt, CEIM Central East Iran Microplate, UDMA Uromia–Dokhtar Magmatic Arc)
Furrer and Soder (1955) divided the Qom Formation into six members, including: basal limestones (a-Member), sandy marlstones (b-Member), alternating marlstones and limestones (c-Member), evaporites (d-Member), green marlstones (e-Member), and top limestones (f-Member). Interareal tracing of the Qom Fm. members, mostly developed at the type locality, is complicated by rapid lateral facies changes and strongly varying thicknesses. In other words, except for the f-Member, the other members can hardly be traced in places far from the type area (e.g., NW Iran). One of the key areas for stratigraphical studies in the SSZ and UDMA is NW Iran (including the Hamedan–Miandoab area), where the deposits of the Qom Formation are well exposed and widespread. Early mapping surveys of NW Iran have only been supplemented with gross stratigraphy in this area. Only recently, some modern micropaleontological studies have been published (Yazdi-Moghadam 2011; Daneshian and Ghanbari 2017; Yazdi-Moghadam et al. 2018a, b). During the Miocene, carbonate-dominated strata (Qom Formation) of variable shallow to moderately deep-marine environments have been deposited in the Hamedan–Miandoab area which are the subject of this study for micropaleontology and biostratigraphy. Therefore, this study is intended to describe LBF of northwest of Iran and to build the global SB system of biochronozones as proposed by Pignatti and Papazzoni (2017).
Materials and methods
This study is based on three outcrop sections: Goli (36° 59′ 21.8′′ N, 46° 51′ 59′′ E), Qabakh Tappeh (35° 27′ 1.98′′ N, 48° 33′ 46.33′′ E), and Qaraie (36° 32′ 19.7′′ N, 47° 59′ 13.0′′ E), all located in the northwest Iran in an area extending from Hamedan to Miandoab (Fig. 1). A total of 104 samples (including 165 thin sections) were studied from the three sections, covering the whole marine strata of the Qom Formation. Larger benthic foraminifera occurred in cemented hard rocks, and matrix-free specimens were not available. Therefore, multiple thin sections were prepared from the limestone samples to obtain oriented sections of the specimens. The generic classification of foraminifera follows Loeblich and Tappan (1987). All the samples and thin sections presented in this paper are deposited in the collection of the National Iranian Oil Company Exploration Directorate (NIOCEXP), Tehran, Iran, under the acronyms of RCH (Goli section), ASK (Qaraie), and ANM (Qabakh Tappeh).
Results
Goli section
The sedimentary succession of the Qom Formation at Goli is 214-m-thick and starts with conglomerates covering the palaeo-relief of Eocene igneous rocks. It characterizes the unconformable contact between the overlying Qom strata and the underlying Eocene volcanics. The onlap of the Qom Formation onto the Eocene igneous paleo-relief continued with the deposition of shallow-marine strata composed of very thick to thick bedded limestones and sandy limestones together with thin beds of marlstones. Detailed lithostratigraphic description of the Qom Formation in the Goli section is shown in Fig. 2. The shallow-marine strata are characterized by the presence of LBF, including Miogypsina globulina, Amphistegina bohdanowiczi, Borelis curdica, and Peneroplis thomasi (Fig. 3). Significant occurrences of smaller benthic foraminifera include Elphidium crispum, Elphidium sp. 1, miliolids, and textulariids. Coralline algae are mainly represented by lithophylloids [e.g., Titanoderma pustulatum (Lamouroux)], mastophoroids (Lithoporella melobesioides Foslie), Sporolithales (Sporolithon sp.), and neogoniolithoids (Spongites sp.). Lithophylloids and neogoniolithoids are common. Bryozoans, corals (Porites-Faviidae assemblage and solitary corals), and Kuphus-tubes are subordinate. The Qom Formation at Goli is unconformably overlain by continental deposits of the Upper Red Formation.

Lithostratigraphic log of the Goli section. (URF Upper Red Formation)

Log of the Goli section showing the distribution of benthic foraminifera and non-foraminiferal skeletal elements
Qabakh Tappeh section
The Qom Formation at Qabakh Tappeh is a competent-weathering, cliff-forming unit. The lower contact between the Qom Fm. and the underlying strata is not exposed due to alluvial cover (Fig. 4). The marine succession of the Qom Formation, 169.5-m-thick, is a carbonate sequence consisting of very thick, thick, and medium bedded shallow-marine limestones. As shown in Fig. 5, limestones contain LBF, including Miogypsina globulina, Amphistegina bohdanowiczi, and Borelis curdica. Acervulinids are common encrusting foraminifers. Miliolids and textulariids chiefly represent the association of small benthic foraminifera. Coralline algae include lithophylloids, neogoniolithoids (Spongites sp), melobesioids, and Sporolithales (Sporolithon sp.). Lithophylloids and neogoniolithoids are more abundant. Bryozoans, bivalves, echinoids, and corals (Porites-Faviidae assemblage) are associated. The upper contact of the Qom Formation is erosional, and the Upper Red Formation is not present here.

Lithostratigraphic log of the Qabakh Tappeh section

Log of the Qabakh Tappeh section showing the distribution of benthic foraminifera and non-foraminiferal skeletal elements
Qaraie section
The Qaraie section is characterized by a disconformable contact between the red polymictic conglomerates, marlstones, and siltstones of the underlying Lower Red Formation and the overlying carbonates of the Qom Formation (Fig. 6). The Qom Fm. is composed of two lithotypes in this area, i.e., the shallow-marine carbonates and deep-marine marlstones and argillaceous limestones. It permits a further subdivision into two lithostratigraphic units. Unit I, with a thickness of 32 m, is composed mainly of very thick bedded to thick bedded shallow-marine limestones. Compared to the type locality, this unit can be correlated with the f-Member. The assemblage of LBF shown in Fig. 7, includes Miogypsina globulina, Amphistegina bohdanowiczi, and Austrotrillina asmariensis. An associated assemblage of small benthic foraminifera is mainly represented by Lobatula sp., Textularia spp., and Miliolidae. Coralline algae are chiefly represented by mastophoroids (Lithoporella melobesioides), Sporolithales (Sporolithon sp.), and a few occurrences of sterile melobesioids. Corals (branching Porites) dominate most of the unit, while bryozoans rarely occur. The first marine marlstones appear 32 m above the base of the Qom Formation, marking the base of Unit II. This unit is characterized by the occurrence of planktonic foraminifera, including Globigerinoides altiaperturus, Streptochilus pristinum, and Globigerina praebulloides, documenting the platform drowning. Finally, the marine strata of the Qom Fm. are unconformably overlain by the continental conglomerates of the Upper Red Formation.

Lithostratigraphic log of the Qaraie section. (LRF Lower Red Formation, URF Upper Red Formation)

Log of the Qaraie section showing the distribution of benthic foraminifera and non-foraminiferal skeletal elements
Systematic paleontology
Miogypsina globulina (Michelotti, 1841) Figs. 8 and 9

Miogypsina globulina (Michelotti). a–i and j–k are equatorial and axial sections of the A-forms, respectively. (a ANM 4594; b RCH 1326; c ASK 3132; d, f RCH 1279; e ASK 3129; g, i: RCH 1313; h RCH 1279; j ANM 4596; k ANM 4592)

1841 Nummulites globulina Michelotti, p. 297, pl. 3, Fig. 6.
1952 Miogypsina irregularis (Michelotti), Drooger, p. 54, pl. 2, Figs. 25, 26, 27, 28 and 29.
1959 Miogypsina globulina (Michelotti), Drooger and Socin, pl. 1, Fig. 5.
1974 Miogypsina globulina (Michelotti), Raju, pp. 82–83, pl. 2, Figs. 1, 2, 3 and 4; pl. 5, Figs. 6, 7; pl. 6, Fig. 1.
1979 Miogypsina globulina (Michelotti), Schiavinotto, pp. 267–268, pl. 3, Figs. 3, 4.
2009 Miogypsina globulina (Michelotti), Özcan and Less, p. 33, pl. 1, Figs. 24, 25; pl. 2, Fig. 6, text-Fig. 8.
2009 Miogypsina globulina (Michelotti), Özcan et al., pp. 568–569, pl. 2, Figs. 20, 21, 22, 23, 24, 25, 26, 27, 28 and 29, text-Fig. 11.
For measurements, see Table 1.
Remarks: According to Drooger (1993), representatives of Miogypsina globulina are defined by having the mean V values between 0 and 45. Our specimens exhibit V values ranging from 29.47 to 39.69 (Table 1), falling within the range of M. globulina.
Amphistegina bohdanowiczi (Bieda 1936) Fig. 10

a–f Amphistegina bohdanowiczi Bieda. (a RCH 1291; b RCH 1321; c–e ASK 1329; f ANM 4610)
1936 Amphistegina bohdanowiczi, Bieda, p. 266, pl. 8, Fig. 4a–c.
1993 Amphistegina bohdanowiczi, Bieda, Rögl and Brandstätter, pl. 3, Figs. 20, 21, 22, 23, 24,25, 26, 27, 28, 29, 30, 31, 32, 33 and 34; pl. 4, Figs. 1, 2, 3, 4, 5 and 6; pl. 6, Figs. 5, 6.
2017 Amphistegina bohdanowiczi, Bieda, Ferràndez-Cañadell and Bover-Arnal, p. 103, Figs. 3E, 15A–J, P. (cum syn.).
Like many parts of the western and central Tethys, this species is widespread in the Oligocene and early Miocene of central and southern Iran. In our studied material, it is characterized by having a small and asymmetrical test reaching to 1 mm in diameter and 0.6 mm in thickness. The triangular test is more or less plano-convex to unequally biconvex with a more inflated ventral side. The tangential section of the test represents a large triangular postulate area in front of the foramen.
Elphidium sp. 1. (Adams and Bourgeois 1967) Fig. 11a–d

a–d Elphidium sp. 1. Adams and Bourgeois. e, f Elphidium crispum (Linnaeus). (a, d, f RCH 1314; b RCH 1289; c RCH 1315; e RCH 1299). (f foramen, rp retral process, s septum, sp spiral canal, up umbilical plug, vc vertical canal)
1967 Elphidium sp. 1. Adams and Bourgeois, p. 709, pl. 19, Fig. 1d.
This species is characterized by having a planispiral, lenticular, and biumbonate test. The umbilical plug is generally large, identical to that in Elphidium craticulatum. Periphery is subacute. The axial section of the test represents the retral process and spiral canal.
Remarks: This species is not adequately described by Adams and Bourgeois (1967). These authors did not point to any features of the internal structure, such as the retral process and spiral canal. According to Adams and Bourgeois (1967), Elphidium sp. 1 has an overall range from the base of Rupelian to Burdigalian. More investigations based on well-preserved material are to be conducted to determine whether or not the Oligocene representatives assigned to this species are true.
Elphidium crispum (Linnaeus 1758) Fig. 11e, f
1758 Nautilus crispus Linnaeus, p. 709, pl. 19, Fig. 1d.
1939 Elphidium crispum (Linnaeus), Cushman p. 50, pl. 13, Figs. 17, 18, 19, 20 and 21.
1958 Elphidium crispum (Linnaeus), Reiss and Merling, pl. 5, Fig. 2.
1976 Elphidium crispum (Linnaeus), Hansen and Lykke-Andersen, p. 6, pl. 1, Figs. 10, 11 and 12; pl. 2, Figs. 1, 2.
2001 Elphidium crispum (Linnaeus), Hottinger, Reiss and Langer, p. 28, pl. 10, Figs. 4, 5, 6 and 7.
Remarks: Elphidium crispum is a typical species of Elphidium. Vertical canals arising from the spiral canal are present in the umbilical plug. Sutures are curved, especially towards the periphery of the test. It is shallow water, normal marine, symbiontic chloroplast-bearing, epibenthic species (Leutenegger 1984; Jorissen 1988; Langer 1989; Hottinger et al. 2001).
Borelis curdica (Reichel 1937) Fig. 12

a–f Borelis curdica (Reichel). (chl chamberlet, ichl intercalary chamberlet, psp preseptal passage, Y-spt Y-shaped septulum). (a RCH 1333; b, e RCH 1299; c, d RCH 1295; f ANM 4592)
1937 Neoalveolina melo curdica n. sp. Reichel, p. 108, pl. 10, Figs. 4, 5, 6 and 7.
2019 Borelis curdica (Reichel), Bassi et al., pp. 11–14, Figs. 9, 10 and 11. (cum syn.).
Only A-forms are present in our material characterized by subspherical to spherical shape of the test. Chamberlets become alternately large and small, resulting in the development of intercalary chamberlets separated by Y-shaped septula. This kind of septula occurs throughout the ontogeny. The elongation index is around 1 and the spherical proloculus varies in diameter from 39 to 60 µm.
Remarks: Borelis curdica is a widespread species in the Burdigalian–Tortonian shallow-marine carbonates of the Mediterranean area. So far, the reliable records of this species are only from the Burdigalian of southern and central Iran (e.g., Hottinger 1974; Seyrafian et al. 2011; Yazdi-Moghadam et al. 2018a).
Peneroplis thomasi (Henson 1950) Fig. 13

a–f Peneroplis thomasi Henson. (a RCH 1337; b–f RCH 1318). (f foramen, fl flexostyle, lw lateral wall, s septum, p protoconch). Scale bars represents 250 µm
1950 Peneroplis thomasi sp. nov. Henson, pp. 36–37, pl. 5, Figs. 7, 8, 9, 10 and 11.
1981 Peneroplis thomasi Henson, Radoiĉić, pl. 1, Figs. 3, 4.
2011 Peneroplis thomasi Henson, Yazdi-Moghadam, pl. 2, Figs. 8.
2013 Peneroplis thomasi Henson, Amirshahkarami, pl. 5, Figs. 9, 10 and 11.
2016 Peneroplis thomasi Henson, Habibi, pl. 2, Fig. 15.
2014 Peneroplis thomasi, Karevan et al., Fig. 5C.
The planispiral, compressed test of Peneroplis thomasi is characterized by initial involute stage with 2–2.5 whorls, followed by a flabelliform evolute final stage of growth. Specimens of the Goli section show up to 14 chambers in the neanic growth stage. In axial and subaxial sections, the test appears as a “knotted rope”. The epidermis is thick. This species is distinct from the other species of the genus Peneroplis by its particular growth morphology and thick epidermis.
Austrotrillina asmariensis (Adams 1968) Fig. 14

a–i Austrotrillina asmariensis Adams. (a–d ASK 3129; e–h ASK 3130; i ASK 3133). (al alveole, fl flexostyle, p protoconch)
1968 Austrotrillina asmariensis Adams, pp. 82–85, pl. 1, Figs. 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11 and 12.
2017 Austrotrillina asmariensis Adams, Ferràndez-Cañadell and Bover-Arnal, p. 88, Figs. 4c–g. (cum syn.).
Only A-forms are recorded. This species is characterized by triloculine fashion of growth and exoskeleton as simple, closely spaced, narrow, and deep alveols. Alveols first appear at the initial stages of the growth. The globular to subglobular proloculus of 76–117 µm has a reniform flexostyle that attains a length of half the circumference of the proloculus (Fig. 14i). The specimens of the Qaraie section reach the length of 0.8–0.106 mm which are somewhat shorter than the original material (1–1.5 mm) described by Adams (1968). The width of the specimens varies between 0.62 and 0.9 mm. Alveoli range in diameter from 13 to 26 µm. The thickness of the wall in the final chambers is between 70 and 82 µm and the number of alveoli per 100 µm in cross-section varies from 3 to 4. Chamber walls are rounded. Our specimens show a few bifurcations in the exoskeleton of the final chambers, a feature that can be observed in B-forms by Adams (1968) and is also reported by Ferràndez-Cañadell and Bover-Arnal (2017).
Discussion
Biostratigraphy
Larger benthic foraminiferal associations of the Goli, Qabakh Tappeh, and Qaraie sections correspond to SBZ 25 based on the occurrence of key taxon Miogypsina globulina. This miogypsinid species is well known from the early Burdigalian of the Tethys realm (Drooger and Socin 1959; Raju 1974; Schiavinotto 1979; Drooger 1993; Cahuzac and Poignant 1997; Özcan and Less 2009; Özcan et al. 2009; Bassi et al. 2019). It marks the SBZ 25 Zone (e.g., Cahuzac and Poignant 1997). Borelis curdica has an overall range of Burdigalian–Tortonian (Adams 1970; Cahuzac and Poignant 1997; Jones et al. 2006; Bassi et al. 2019). Peneroplis thomasi is common in the lower to upper Oligocene shallow-marine deposits of the Middle East (e.g., Henson 1950; Radoiĉić 1981; Yazdi-Moghadam 2011; Karevan et al. 2014; Habibi 2016). The occurrence of this species in the upper Oligocene of Europe (Western Tethys) is reported by Ferràndez-Cañadell and Bover-Arnal (2017). A Rupelian–Aquitanian stratigraphic range is suggested by Eames et al. (1962), and a Chattian–Aquitanian range has also been inferred by Amirshahkarami (2013). The occurrence of this species from the Burdigalian (SBZ 25 Zone) shallow-marine limestones of the Qom Fm. in the Goli section, represents its youngest known record. Therefore, the overall range of Peneroplis thomasi has to be considered as Rupelian–Burdigalian (SBZ 21-SBZ 25). Austrotrillina asmariensis has been mostly reported from the Rupelian to Aquitanian in Europe and the Middle East (e.g., Adams 1968; Accordi et al. 1998; Gallardo et al. 2001; Sirel 2003; Bassi et al. 2007; Gedik 2015; Serra-Kiel et al. 2016; Ferràndez-Cañadell and Bover-Arnal 2017). The only occurrence of this species from the Burdigalian of the Far East (Upper Te, Borneo) is by BouDagher-Fadel et al. (2000). The occurrence of Austrotrillina asmariensis from the Burdigalian (SBZ 25) shallow-marine limestones of the Qom Fm. in the Qaraie section represents its youngest record in Iran and the Middle East. Hence, the total range of this species has to be revised to Rupelian–Burdigalian (SBZ 21–SBZ 25). So far, Elphidium crispum is known from the Miocene to recent (e.g., Reiss and Merling 1958; Hottinger 2010). Our observations based on previously published data (Yazdi-Moghadam et al. 2018a), together with the results of the current study, suggest that the earliest occurrence of Elphidium crispum characterizes the Burdigalian age (SBZ 25) in Central Iran.
Palaeoenvironmental and palaeoclimatic implications
Larger benthic foraminifera and coralline algae occur abundantly in many platform deposits (e.g., Beavington-Penney and Racey 2004; Bassi and Nebelsick 2010; Ghosh and Sarkar 2013; Sarkar 2016; Pomar et al. 2017). The groups are distributed in a wide range of platform environments. Therefore, LBF and coralline algae are excellent palaeoenvironmental indicators which may be used as valuable tools to discern environmental changes in lithologically monotonous platforms (e.g., Nebelsick et al. 2013; Sarkar and Ghosh 2015; Basso et al. 2019). According to Drooger (1993), Miogypsina thrives in shallow waters (up to 50–80 m). The high abundance of miogypsinids indicates middle ramp to proximal part of the outer ramp environment. Amphistegina is an inhabitant of the tropical-subtropical belt. It is dependent on symbionts and lives in shallow waters down to 70–80 m, occasionally somewhat deeper (Rögl and Branstӓtter 1993). Borelis is said to thrive in shallow, warm, protected zones of reef complexes (Cole 1954, 1957; Chevalier 1973; Ghose 1977). A detailed ecological study in the Gulf of Aqaba revealed that Borelis hosts B-3 diatom symbionts and is restricted to depths of 5–65 m (Reiss and Hottinger 1984). For the Miocene, these authors postulate a shallow environment where the alveolinids (e.g., Borelis) lived down to a depth between 20 and 40 m. A warm, clear, shallow water up to 33 m deep environment with a temperature around 25 is inferred for Borelis melo, an early-middle Miocene marker in the Mediterranean area and the Middle East, by Bandy (1960) and Murray (1991). The genus Peneroplis, very common on the reef flat, clearly prefers hardgrounds of the shallowest slope parts down to 30 m (Hohenegger et al. 1999). Schuster and Wielandt (1999) state that the association of LBF, including Borelis curdica, Peneroplis evolutus, Archaias sp., and miliolids occurring within the patch reefs in the Qom Fm., are typical for very shallow and well-illuminated habitats in sheltered, back-reef environments. According to these authors, the late Oligocene and early Miocene coral faunas of Central Iran, represent a mixture of Indo-pacific and Mediterranean elements characterizing the normal marine sedimentation. Coralline algae are also common in taxonomic composition of the Qom Formation. The association of coralline algae and corals together with LBF are usually considered to live in the oligophotic zone of the middle ramp environment (e.g., Pomar 2001a; b; Brandano and Corda 2002; Corda and Brandano 2003; Bassi 2005). Similarly, limestones of the Qom Fm. composed of corallinaceans and LBF have been interpreted to be typical for the middle ramp (e.g., Reuter et al. 2009). The abundance of red algae is compatible with a water depth of about 30–50 m for the oligophotic zones (Hallock 1987, 1988; Brandano et al. 2010). Coralline algae are common both in tropical as well as in temperate waters (Braga and Aguirre 2001). According to Aguirre et al. (2000), in modern tropical environments, mastophoroids are the most abundant coralline algae in shallow water, whereas melobesioids become more abundant below 10–20 m, and Sporolithales are abundant at greater depth (40–50 m). Neogoniolithoids are common in shallow subtidal depths below 40 m (Adey et al. 1982; Perrin et al. 1995; Aguirre et al. 2000). The presence of tropical genera (Lithoporella and Sporolithon) in the red algal assemblages of the Qom Fm. suggests that carbonate sedimentation took place in warm waters. LBF are generally restricted to warm tropical and subtropical waters, with means of 18–20 °C for the warmest months of the year (Adams et al. 1990). Oligocene and early Miocene LBF, such as miogypsinids are thought to be restricted to tropical to subtropical regions in oligo- to slightly mesotrophic conditions (Betzler et al. 1997). Therefore, also the larger benthic foraminiferal assemblages of the Qom Fm. support deposition in warm waters. The presence of planktonic foraminifera is indicative of open marine environment and their relative abundance increases basinward (e.g., Geel 2000). Summing up, the biotic assemblages of the shallow-marine part of the Qom Fm. (f-Member) in the study area suggest that the deposition took place in shallow-marine environment (middle ramp) under oligotrophic conditions and in warm waters of the tropical to subtropical zones. The common occurrences of neogoniolithoids and lithophylloids together with the abundance of certain LBF (Borelis and Peneroplis) in Goli and Qabakh Tappeh sections suggest a water depth less than 40 m (5–40 m). Instead, a greater water depth (30–50 m) can be inferred for the shallow-marine limestones of the Qaraie section due to the dominance of Sporolithales and mastophoroids with respect to melobesioids. The presence of planktonic foraminifera in the upper part of this section (Unit II) is indicative of outer ramp deposition with a water depth possibly more than 50 m. The absence of deep-marine representatives of coralline algae in this part of the Qaraie section may point to fine-grained sedimentation which severely reduced water transparency.
Conclusions
In the lower Miocene shallow-marine platforms of NW Iran, Peneroplis thomasi and Austrotrillina asmariensis coexist with key Burdigalian species Miogypsina globulina (Fig. 15). Therefore, the upper range of these porcelaneous foraminifera has to be extended into the Burdigalian (SBZ 25). The larger foraminiferal and red algal assemblages of the Qom Fm. (Hamedan–Miandoab area) confirm sedimentation in warm waters of the tropical–subtropical zones (middle ramp setting). Deposition in the oligotrophic conditions is also compatible with the composition of red algal assemblages and the abundance and diversity of larger benthic foraminifera. The results of this study together with the previously published data suggest that the LBF composition of Central Iran show faunal elements from both western and eastern sides of the Tethys during the Burdigalian. This demonstrates the persisting connection between the Qom Basin with both Mediterranean and Indo-Pacific Oceans until the early to mid-Burdigalian. However, the presence of porcelaneous larger benthic foraminifera, including A. asmariensis and P. thomasi, so far completely unknown from the Burdigalian strata in Western Tethys, points to the restriction of the Tethyan Seaway before its final closure. This restriction of seaway connection is in agreement with the idea of Reuter et al. (2009), who stated that the differentiation into an Atlantic–Mediterranean and an Indo-Pacific bioprovince took place before the final paleogeographic disjunction of the Tethys (Terminal Tethyan Event). Our biostratigraphic results suggest that the marine transgression of the Hamedan–Miandoab platform (Qom Fm.) occurred during the early Burdigalian.

Stratigraphic distribution of selected hyaline and porcelaneous benthic foraminifera based on Cahuzac and Poignant (1997) and results of this study
References
Accordi G, Carbone F, Pignatti J (1998) Depositional history of a Paleogene carbonate ramp (western Cephalonia, Ionian Islands, Greece). GeolRomana 34:131–205
Adams CG (1968) A revision of the foraminiferal genus Austrotrillina PARR. Bull Br Mus Nat HistGeol 16:71–97
Adams CG (1970) A reconsideration of the East Indian Letter Classification of the Tertiary. Bull Br Mus Nat HistGeol 19:87–137
Adams T, Bourgeois F (1967) Asmari biostratigraphy: Iranian Oil Operating Companies. Geolog Explor Division, Rep No. 1074
Adams CG, Lee DE, Rosen BR (1990) Conflicting isotopic and biotic evidence for tropical sea-surface temperatures during the Tertiary. PalaeogeogrPalaeoclimatolPalaeoecol 77:289–313
Adey WH, Townsend RA, Boykins WT (1982) The crustose coralline algae (Rhodophyta: Corallinaceae) of the Hawaiian Islands. Smithson Contrib Mar Sci 15:1–74
Agard P, Omrani J, Jolivet, Whitechurch, Vrielynck B, Spakman W, Monie P, Meyer B, Wortel R (2011) Zagros orogeny: a subduction-dominated process. Geol Mag 148(5–6):692–725
Aguirre J, Riding R, Braga JC (2000) Diversity of coralline red algae: origination and extinction patterns from the Early Cretaceous to the Pleistocene. Paleobiology 26:651–667
Akbar-Baskalayeh N, Less G, Ghasemi-Nejad E, Yazdi-Moghadam M, Pignatti J (2020) Biometric study of late Oligocene larger benthic Foraminifera (Lepidocyclinidae and Nummulitidae) from the Qom Formation, Central Iran (Tajar-Kuh section). J Paleontol 94:593–615
Allahkarampour Dill M, Vaziri-Moghaddam H, Seyrafian A, Behdad A (2018) Oligo-Miocene carbonate platform evolution in the northern margin of the Asmari intra-shelf basin, SW Iran. Mar Pet Geol 92:437–461
Amato V, Drooger CW (1969) How to measure the angle γ in the Miogypsinidae. Rev EspMicropaleontol 1:19–24
Amirshahkarami M (2013) Revision in the paleontology and distribution of the larger benthic foraminifera in the Oligocene-Miocene deposits of the Zagros Basin, southwest Iran. Hist Biol 25:339–361
Bandy OL (1960) General correlation of foraminiferal structure with environment. In: Proc. 21st int. geol. cong., Norden, vol 22, pp 7–19
Bassi D (2005) Larger foraminiferal and coralline algal facies in an Upper Eocene storm-influenced, shallow-water carbonate platform (Colli Berici, north-eastern Italy). PalaeogeogrPalaeoclimatolPalaeoecol 226:17–35
Bassi D, Nebelsick JH (2010) Components, facies and ramps: redefining Upper Oligocene shallow water carbonates using coralline red algae and larger foraminifera (Venetian area, northeast Italy). PalaeogeogrPalaeoclimatolPalaeoecol 295:258–280
Bassi D, Hottinger L, Nebelsick J (2007) Larger foraminifera from the Upper Oligocene of the Venetian area, North-East Italy. Palaeontology 50:845–868
Bassi D, Braga JC, Di Domenico G, Pignatti J, Abramovich S, Hallock P, Könen J, Kovács Z, Langer MR, Pavia G, Iryu Y (2019) Palaeobiogeography and evolutionary patterns of the larger foraminifer Borelis de Montfort (Borelidae). Pap Palaeontol. https://doi.org/10.5061/dryad.st65n12
Basso D, Coletti G, Bracchi VA, Yazdi-Moghadam M (2019) Lower oligocene coralline algae of the Uromieh section (Qom Formation, NW Iran) and the oldest record of Titanoderma pustulatum (Corallinophycidae, Rhodophyta). RivItalPaleontol S 125:197–218
Beavington-Penney SJ, Racey A (2004) Ecology of extant nummulitids and other larger benthic foraminifera: applications in palaeoenvironmental analysis. Earth Sci Rev 67:219–265
Berberian M (1974) A brief geological description of north-central Iran. Mater Study Seismotectonics Iran North Central Iran GeolSurv Iran Rep 29:127–138
Berberian M, King G (1981) Towards a paleogeography and tectonic evolution of Iran. Can J Earth Sci 18:210–265
Betzler C, Brachert TC, Nebelsick JH (1997) The warm temperate carbonate province. A review of facies, zonations, and delimitations. CourForschInstSenckenberg 201:83–99
Bieda F (1936) MiocenBrzozowej I Gromnika I jego fauna otwornicowa (Le Miocéne de Brzozowaet de Gromniketsa fauna des Foraminiferes). Polskiego Tow Geol 12:255–275
BouDagher-Fadel MK, Noad JJ, Lord AR (2000) Larger foraminifera from late Oligocene-earliest Miocene reefal limestones of north east Borneo. Rev EspMicropaleontol 32:341–362
Boukhary M, Abdelghany O, Hussein-Kamel Y, Bahr S, Alsayigh AR, Abdelraouf M (2010) Oligocene larger foraminifera from United Arab Emirates, Oman and Western Desert of Egypt. Hist Biol 22:348–366
Bozorgnia F (1966) Qom formation stratigraphy of the Central Basin of Iran and its intercontinental position. Iran Pet Inst Bull 24:69–76
Braga JC, Aguirre J (2001) Coralline algal assemblages in Upper Neogene reef and temperate carbonates in Southern Spain. PalaeogeogrPalaeoclimatolPalaeoecol 175:27–41
Brandano M, Corda L (2002) Nutrients, sea level and tectonics: constrains for the facies architecture of a Miocene carbonate ramp in central Italy. Terra Nova 14:257–262
Brandano M, Morsilli M, Vannucci G, Parente M, Bosellini F, Mateu-Vicens G (2010) Rodolith-rich lithofacies of the Porto Badisco Calcarenites (upper Chattian, Salento, southern Italy). Boll Soc Geol It 129:119–131
Cahuzac B, Poignant A (1997) Essai de biozonation de l’Oligo-Miocène dans les bassins européens à l’aide des grands foraminifères néritiques. B Soc Géol Fr 168:155–169
Chevalier JP (1973) Coral reefs of New Caledonia. In: Jones OA, Endean R (eds) Biology and geology of Coral Reefs, 1, Geology 1. Academic Press, New York, pp 143–167
Cole WS (1954) Larger Foraminifera and smaller diagnostic Foraminifera from Bikini drill holes. US GeolSurv Prof Pap 260–0:569–608
Cole WS (1957) Larger Foraminifera from Eniwetok Atoll drill holes. US Geol Surv Prof Pap 260–V:743–784
Corda L, Brandano M (2003) Aphotic zone carbonate production on a Miocene ramp Central Apennines, Italy. Sediment Geol 6:55–70
Cushman JA (1939) A monograph of the foraminiferal family Nonionidae. US Government Printing Office
Daneshian J, Ghanbari M (2017) Stratigraphic distribution of planktonic foraminifera from the Qom Formation: a case study from the Zanjan area (NW Central Iran). NeuesJahrbGeol P A 283:239–254
Drooger CW (1952) Study of American Miogypsinidae. University of Utrecht, Utrecht, p 80
Drooger CW (1993) Radial foraminifera: morphometrics and evolution. Verhandelingen der Koninklijke Nederlandse Akademie van Wetenschappen, Afd. Natuurkunde, Erste Reeks, Amsterdam, deel 41
Drooger C, Socin C (1959) Miocene foraminifera from Rosignano, northern Italy. Micropaleontology 5:415–426
Eames F, Banner F, Blow W, Clarke W, Smout A (1962) Morphology, taxonomy, and stratigraphic occurrence of the Lepidocyclininae. Micropaleontology 8:289–322
Ehrenberg S, Pickard A, Laursen G, Monibi S, Mossadegh Z, Svånå T, Aqrawi A, McArthur J, Thirlwall M (2007) Strontium isotope stratigraphy of the Asmari Formation (Oligocene-Lower Miocene), SW Iran. J Pet Geol 30:107–128
Ferràndez-Cañadell C, Bover-Arnal T (2017) Late Chattian Larger Foraminifera from the Prebetic Domain (SE Spain): new data on Shallow Benthic Zone 23. Palaios 32:83–109
Furrer M, Soder P (1955) The Oligo-Miocene marine formation in the Qom region (Central Iran). In: Proceedings of the 4th world petroleum congress, Rome, Section I/A/5, pp 267–277.
Gallardo A, Serra-Kiel J, Ferràndez-Cañadell C, Roger J, Boix C, Caus E (2001) Macroforaminíferos porcelanados del Eoceno Superior-Oligoceno Inferior del Dhofar (Sultanato de Omán) In: Meléndez G, Herrera Z, Delvene G, Azanza B (eds) Los Fósiles y la Paleogeografía. Publicaciones del Seminario de Paleontología de Zaragoza (SEPAZ), vol 5, pp 83–89
Gedik F (2015) Benthic foraminiferal biostratigraphy of Malatya Oligo-Miocene succession (Eastern Taurids, Eastern Turkey). B Mineral Res Expl 150:19–50
Geel T (2000) Recognition of stratigraphic sequences in carbonate platform and slope deposits: empirical models based on microfacies analysis of Palaeogene deposits in southeastern Spain. PalaeogeogrPalaeoclimatolPalaeoecol 155:211–238
Ghos BK (1977) Paleoecology of the Cenozoic reefal foraminifers and algae-a brief review. PalaeogeogrPalaeoclimatolPalaeoecol 22:231–256
Ghosh AK, Sarkar S (2013) Facies analysis and palaeoenvironmental interpretation of Piacenzian carbonate deposits from the Guitar Formation of Car Nicobar Island, India. Geosci Front 4:755–764
Habibi T (2016) Bio- and sequence stratigraphy and microfacies analysis of the Oligocene Asmari Formation at Sepidar Anticline, Interior Fars sub-Basin, SW Iran. Hist Biol 28:519–532
Hakyemez A, Özgen-Erdem N, Kangal Ö (2016) Planktonic and benthic foraminiferal biostratigraphy of the Middle Eocene-Lower Miocene successions from the Sivas Basin (Central Anatolia, Turkey). GeolCarapathica 67:21–40
Hallock P (1987) Fluctuations in the trophic resource continuum: a factor in global diversity cycles? Paleoceanography 2:457–471
Hallock P (1988) The role of nutrient availability in bioerosion: consequences to carbonate buildups. PaleogeogrPaleoclimatolPaleoecol 63:275–291
Hansen HJ, Lykke-Andersen AL (1976) Wall structure and classification of fossil and recent elphidiid and nonionid Foraminifera: Universitetsforlaget. Oslo, p 37
Henson FRS (1948) Larger imperforate Foraminifera of Southwestern Asia: Families Lituolidue, Orbitolinidae and Meandropsinidae. British Museum Natural History, pp 1–127
Henson FRS (1950) Middle Eastern Tertiary Peneroplidae (Foraminifera): with remarks on the phylogeny and taxonomy of the family. The West Yorkshire Printing Co. Limited, Wakefield
Hohenegger J, Yordanova E, Nakano Y, Tatzreiter F (1999) Habitats of larger foraminifera on the upper reef slope of Sesoko Island, Okinawa, Japan. Mar Micropaleontol 36:109–168
Hottinger L (1974) Alveolinids, Cretaceous-Tertiary larger foraminifera. Exxon Production Research Company, Technical Information Services
Hottinger L (2007) Revision of the foraminiferal genus Globoreticulina RAHAGHI, 1978, and of its associated fauna of larger foraminifera from the late Middle Eocene of Iran. Carnets Geol. https://doi.org/10.4267/2042/9213
Hottinger L (2010) The rotaliids of the Lockhartia Sea as indicators of foraminiferal hot spot diversity in the Neotethys during the Eocene in Forams 2010. In: International symposium on foraminifera abstract volume, University of Bonn, Germany, p 110
Hottinger L, Reiss Z, Langer M (2001) Spiral canals of some Elphidiidae. Micropaleontology 47:5–34
Jones RW, Simmons MD, Whittaker JE (2006) On the stratigraphical and palaeobiogeographical significance of Borelis melo melo (Fichtel & Moll, 1798) and B. melo curdica (Reichel, 1937) (Foraminifera, Miliolida, Alveolinidae). J Micropalaeontol 25:175–185
Jorissen FJ (1988) Benthic foraminifera from the Adriatic Sea: principles of phenotypic variation. Utrecht Micropaleont Bul 37:1–174
Karevan M, Vaziri-Moghaddam H, Mahboubi A, Moussavi-Harami R (2014) Biostratigraphy and paleo-ecological reconstruction on Scleractinian reef corals of Rupelian-Chattian succession (Qom Formation) in northeast of Delijan area. Geopersia 4:11–24
Langer MR (1989) Distribution, diversity and functional morphology of benthic foraminifera from Vulcano (Mediterranean Sea), Unpublished Ph.D. Thesis. Universitaet Basel, Switzerland, p 159
Less G (1991) Upper Oligocene larger foraminifers of the Bükk Mountains. Magyar ÁllamiFöldtaniIntézetÉviJelentése 1989:411–465
Letouzey J, Rudkiewicz J (2005) Structural geology in the Central Iranian Basin, Institut Français du Petrole report F0214001
Leutenegger S (1984) Symbiosis in benthic foraminifera; specificity and host adaptations. J Foraminifer Res 14:16–35
Linnaeus C (1758) Systema Naturae per regna tria naturae, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Editio decima, reformata, Laurentius Salvius: Holmiae
Loeblich AR, Tappan H (1987) Foraminiferal genera and their classification. Van Nostrand Reinhold Co., New York
Michelotti G (1841) Saggiostorico dei rizopodicaratteristici dei terrenisopracretacei. MemFisicaSocietaItalScienze 22:253–302
Morley CK, Kongwung B, Julapour AA, Abdolghafourian M, Hajian M, Waples D, Warren J, Otterdoom H, Srisuriyon K, Kazemi H (2009) Structural development of a major late Cenozoic basin and transpressional belt in central Iran: the Central Basin in the Qom-Saveh area. Geosphere 5:325–362
Murray JW (1991) Ecology and paleoecology of benthonic foraminifera. Longman Scientific and Technical, England, p 397
Nebelsick JH, Bassi D, Lempp J (2013) Tracking paleoenvironmental changes in coralline algal-dominated carbonates of the Lower Oligocene Calcareniti di Castelgomberto formation (Monti Berici, Italy). Facies 59:133–148
Omidvar M, Safari A, Vaziri-Moghaddam H, Ghalavand H (2018) Foraminiferal biostratigraphy of Upper Cretaceous (Campanian – Maastrichtian) sequences in the Peri-Tethys basin; Moghan area, NW Iran. J Afr Earth Sci 140:94–113
Özcan E, Less G (2009) First record of the co-occurrence of western Tethyan and Indo-Pacific larger foraminifera in the Burdigalian of the Mediterranean province. J Foraminifer Res 39:23–39
Özcan E, Less G, Báldi-Beke M, Kollányi K, Acar F (2009) Oligo-Miocene foraminiferal record (Miogypsinidae, Lepidocyclinidae and Nummulitidae) from the Western Taurides (SW Turkey): biometry and implications for the regional geology. J Asian Earth Sci 34:740–760
Özcan E, Less G, Báldi-Beke M, Kollányi K (2010) Oligocene hyaline larger foraminifera from Kelereşdere Section (Muş, Eastern Turkey). Micropaleontology 56:465–493
Perrin C, Bosence DWJ, Rosen B (1995) Quantitative approaches to palaeozonation and palaeobathymetry of corals and coralline algae in Cenozoic reefs. In: Bosence DWJ, Allison PA (eds) Marine palaeoenvironmental analysis from fossils. Geol Soc Spec Pub, vol 83, pp 181–229
Pignatti J, Papazzoni CA (2017) Oppelzones and their heritage in current larger foraminiferal biostratigraphy. Lethaia 50:369–380
Pomar L (2001a) Ecological control of sedimentary accommodation: evolution from carbonate ramp to rimmed shelf, Upper Miocene, Balearic Islands. PalaeogeogrPalaeoclimatolPalaeoecol 175:249–272
Pomar L (2001b) Types of carbonate platforms: a genetic approach. Basin Res 13:313–334
Pomar L, Baceta JI, Hallock P, Mateu-Vicens G, Basso D (2017) Reef building and carbonate production modes in the west-central Tethys during the Cenozoic. Mar Pet Geol 83:261–304
Radoiĉić R (1981) Some new data about subsurface biostratigraphy of the Western Iraqi desert (Block 7). Academ Serb Scidences Arts Bull 75:115–137
Raju DS (1974) Study of Indian Miogypsinidae. Utrecht Micropaleontol Bull 9:1–148
Reichel M (1937) Étude sur les Alveolines. Mém Société Paléontol Suisse 59:95–147
Reiss Z, Hottinger L (1984) The Gulf of Aqaba: ecological micropalaeontology. Springer Verlag, Berlin, pp 1–354
Reiss Z, Merling P (1958) Structure of some Rotaliidea. B GeologSurv Israel 21:1–19
Reuter M, Piller W, Harzhauser M, Mandic O, Berning B, Rögl F, Kroh A, Aubry M-P, Wielandt-Schuster U, Hamedani A (2009) The Oligo-/Miocene Qom Formation (Iran): evidence for an early Burdigalian restriction of the Tethyan Seaway and closure of its Iranian gateways. Int J Earth Sci 98:627–650
Rögl F, Brandstätter F (1993) The foraminifera genus Amphistegina in the Korytnica Clays (Holy Cross Mts, Central Poland) and its significance in the Miocene of the Paratethys. Acta Geol Pol 43:121–146
Saraswati PK, Khanolkar S, Banerjee S (2018) Paleogene stratigraphy of Kutch, India: an update about progress in foraminiferal biostratigraphy. Geodin Acta 30:100–118
Sarfi M, Yazdi-Moghadam M (2016) Stratigraphy of the Upper Jurassic shallow marine carbonates of the Moghan area (NW Iran), with paleobiogeography implication on Alveosepta jaccardi (Schrodt, 1894). Geopersia 6:187–196
Sarkar S (2016) Upper Pliocene heterozoan assemblage from the Guitar Formation of Car Nicobar Island, India: palaeoecological implications and taphonomic signatures. PalaeobiodiversityPalaeoenvironments 96:221–237
Sarkar S, Ghosh AK (2015) Evaluation of coralline algal diversity from the Late Middle Miocene sediments of Little Andaman Island (Hut Bay). Carbonates Evaporites 30:13–24
Schaub H (1981) Nummulites et Assilina de la Te’thyspale’og’ene. Taxinomie Phylogen’ese et biostratigraphieMem Suisses Paleontol 104–106:1–236
Schiavinotto F (1979) Miogypsina e Lepidocyclinadel Miocene di Monte La Serra (L’Aquila-Appennino centrale). GeolRomana 18:253–293
Schuster F, Wielandt U (1999) Oligocene and early Miocene coral faunas from Iran: paleoecology and paleobiogeography. Int J Earth Sci 88:571–581
Serra-Kiel J, Gallardo-Garcia A, Razin P, Robinet J, Roger J, Grelaud C, Leroy S, Robin C (2016) Middle Eocene-Early Miocene larger foraminifera from Dhofar (Oman) and Socotra Island (Yemen). Arab J Geosci 9:344
Seyrafian A, Vaziri-Moghaddam H, Arzani N, Taheri A (2011) Facies analysis of the Asmari Formation in central and north-central Zagros basin, southwest Iran: biostratigraphy, paeleoecology and diagenesis. Rev MexCiencGeol 28:439–458
Shabafrooz R, Mahboubi A, Vaziri-Moghaddam H, Ghabeishavi A, Moussavi-Harami R (2015) Depositional architecture and sequence stratigraphy of the Oligo-Miocene Asmari platform; Southeastern Izeh Zone, Zagros Basin, Iran. Facies 61:1–32
Sirel E (2003) Foraminiferal description and biostratigraphy of the Bartonian, Priabonian and Oligocene shallow-water sediments of the southern and eastern Turkey. Rev Paleobiol 22:269–339
Sirel E, Özgen-Erdem N, Kangal Ö (2013) Systematics and biostratigraphy of Oligocene (Rupelian-Early Chattian) foraminifera from lagoonal-very shallow water limestone in the eastern Sivas Basin (central Turkey). Geol Croat 66:83–109
Stöcklin J (1968) Structural history and tectonics of Iran: a review. AAPG Bull 52:1229–1258
Stöcklin J, Nabavi M (1973) Tectonic map of Iran (1:2,500,000). GeolSurv Iran, Tehran
Van Buchem F, Allan T, Laursen G, Lotfpour M, Moallemi A, Monibi S, Motiei H, Pickard N, Tahmasbi A, Vedrenne V (2010) Regional stratigraphic architecture and reservoir types of the Oligo-Miocene deposits in the Dezful Embayment (Asmari and Pabdeh Formations) SW Iran. J Geol Soc Lond Spec Publ 329:219–263
Vaziri-Moghaddam H, Kimiagari M, Taheri A (2006) Depositional environment and sequence stratigraphy of the Oligo-Miocene Asmari Formation in SW Iran. Facies 52:41–51
Yazdi-Moghadam M (2011) Early Oligocene larger foraminiferal biostratigraphy of the Qom Formation, south of Uromieh (NW Iran). Turk J Earth Sci 20:847–856
Yazdi-Moghadam M, Sarfi M, Sharifi M, Ariafar B, Sajjadi F, Abbasi P (2017) Early Barremianorbitolinid record from the Moghan area, NW Iran: Northern margin of the Neotethys. Cretac Res 77:133–142
Yazdi-Moghadam M, Sadeghi A, Adabi MH, Tahmasbi A (2018a) Foraminiferal biostratigraphy of the lower Miocene Hamzian and Arashtanab sections (NW Iran), northern margin of the Tethyan Seaway. Geobios 51:231–246
Yazdi-Moghadam M, Sadeghi A, Adabi MH, Tahmasbi A (2018b) Stratigraphy of the Lower Oligocene nummulitic limestones, north of Sonqor (NW Iran). RivItalPaleontol S 124:407–419
Acknowledgements
The first author is grateful to NIOC Exploration Directorate for permission of publication and all the authors thank Damghan, Tehran, and Shahid Beheshti universities to support this study. We acknowledge constructive comments by three anonymous reviewers that remarkably improved an earlier version of this article. We extend our sincere gratitude for editorial handling to prof. Wolf-Christian Dullo, Editor in Chief, International Journal of Earth Sciences. Last but not least, we dedicate the paper to the late prof. Ebrahim Ghasemi-Nejad (Tehran University) who sadly passed away in December 2020 because of Covid-19.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Rights and permissions
About this article

Cite this article
Yazdi-Moghadam, M., Sarfi, M., Ghasemi-Nejad, E. et al. Early Miocene larger benthic foraminifera from the northwestern Tethyan Seaway (NW Iran): new findings on Shallow Benthic Zone 25. Int J Earth Sci (Geol Rundsch) 110, 719–740 (2021). https://doi.org/10.1007/s00531-021-01986-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00531-021-01986-1