
A Single Plasmid of Nisin-Controlled Bovine and Human Lactoferrin Expressing Elevated Antibacterial Activity of Lactoferrin-Resistant Probiotic Strains

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. Cloning of BLF and HLF Genes into Shuttle Plasmids
2.3. Bacterial Transformation and Electroporation
2.4. Analysis of LF Expression in Various Probiotic Transformants
2.5. Determination of LF Concentration in Probiotic Transformants
2.6. Antibacterial Activities of rLFs
2.7. Recombinant Lactobacilli Growth Assay
2.8. Statistical Analysis
3. Results and Discussion
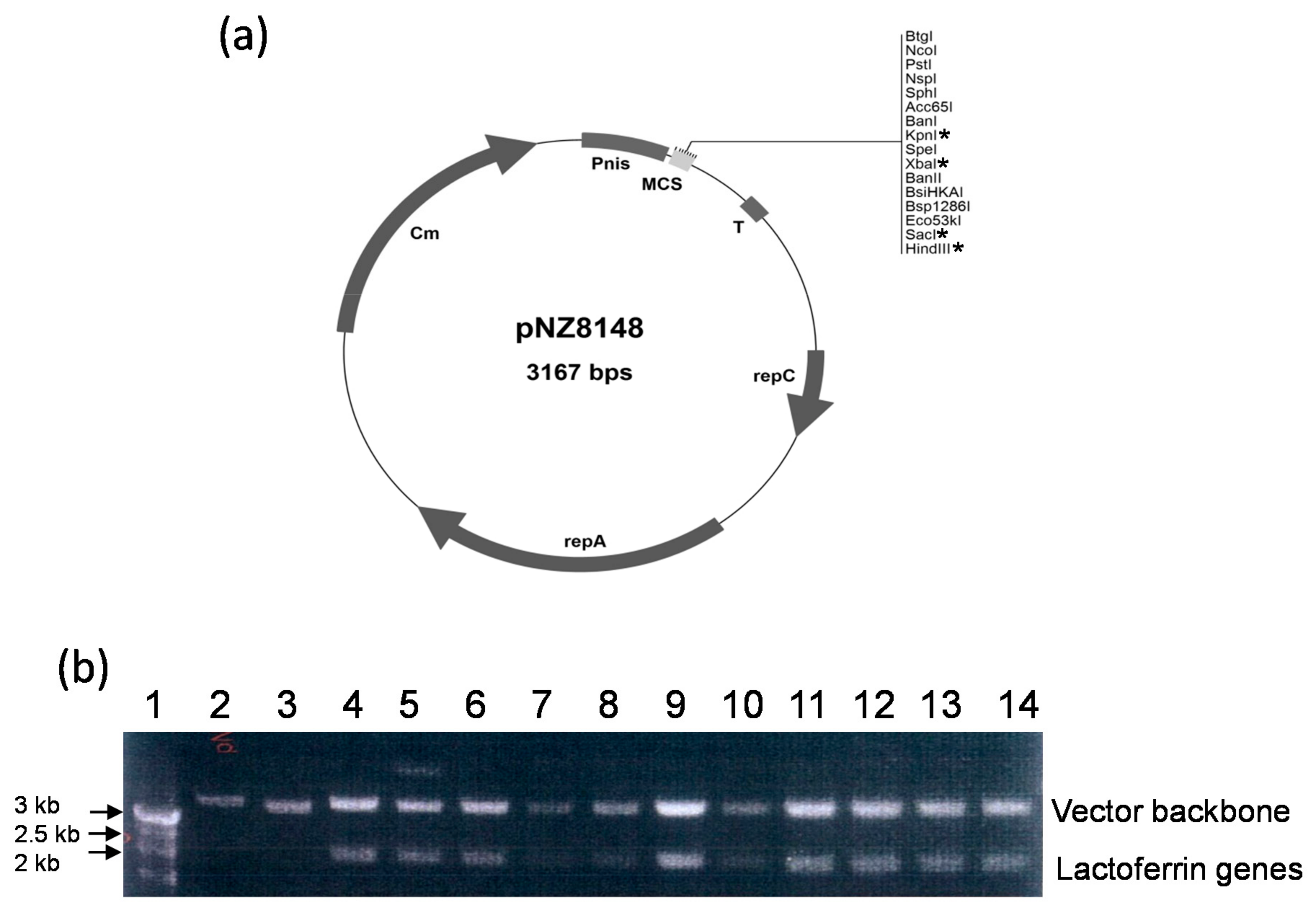
3.1. Construction of Recombinant Vectors
3.2. Transfection of the Constructed Plasmids into Various Hosts Using Electroporation
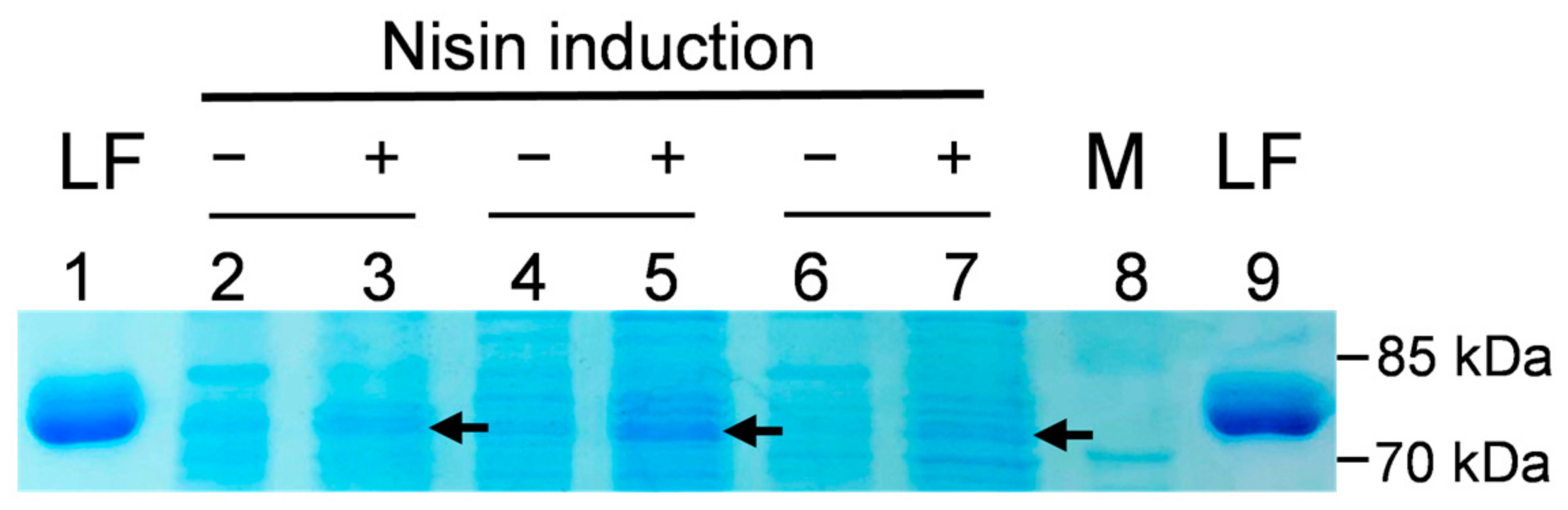
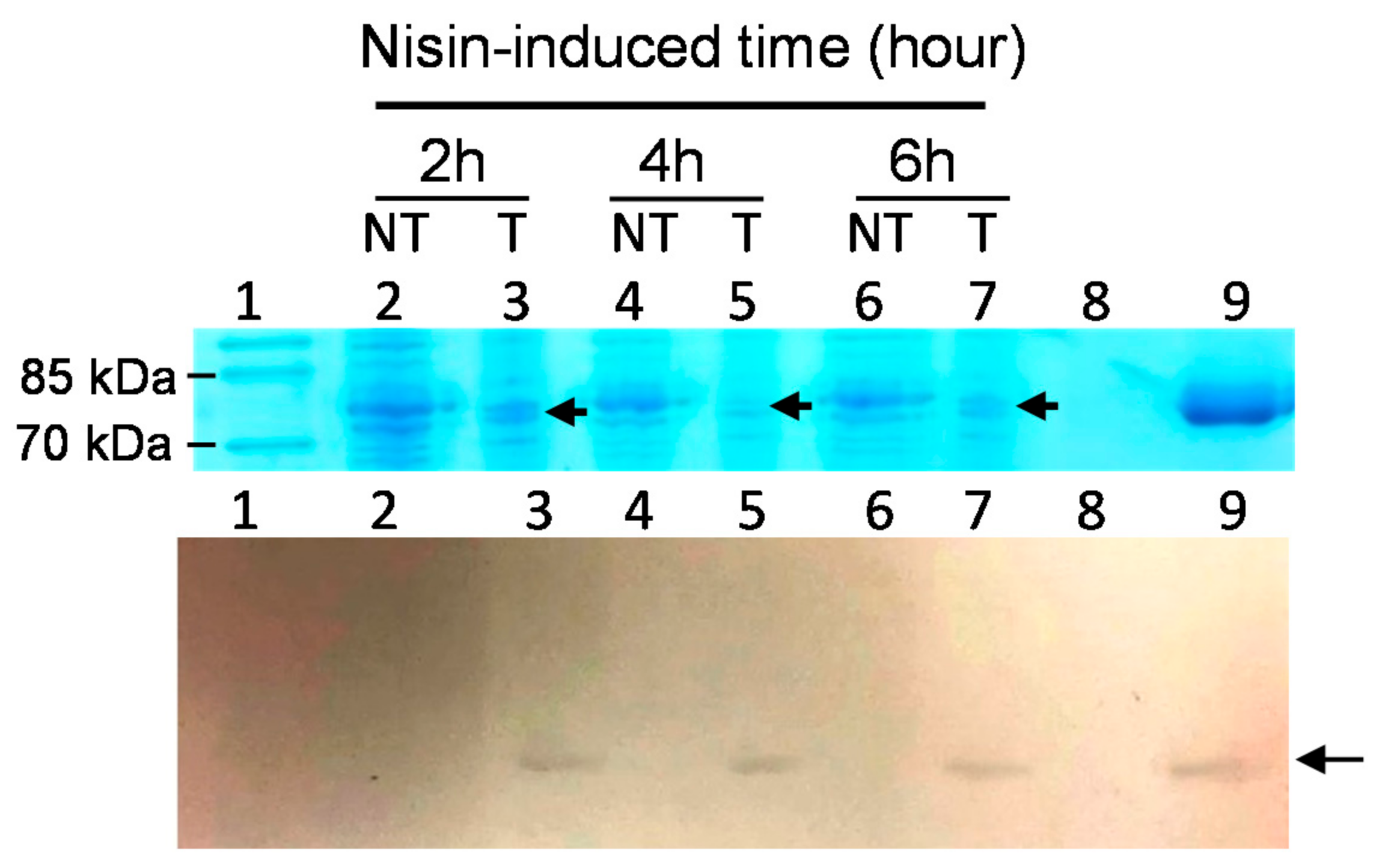
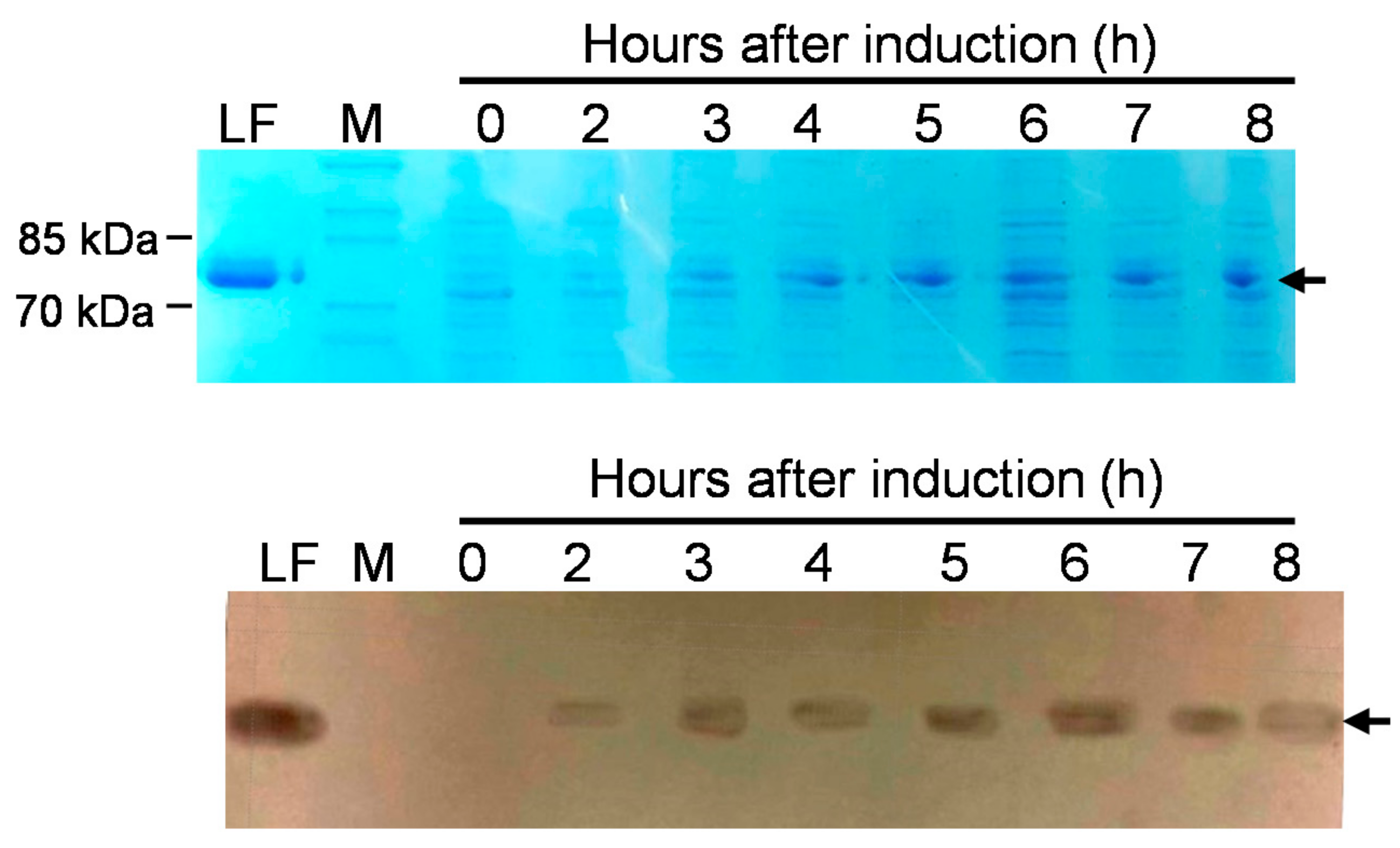
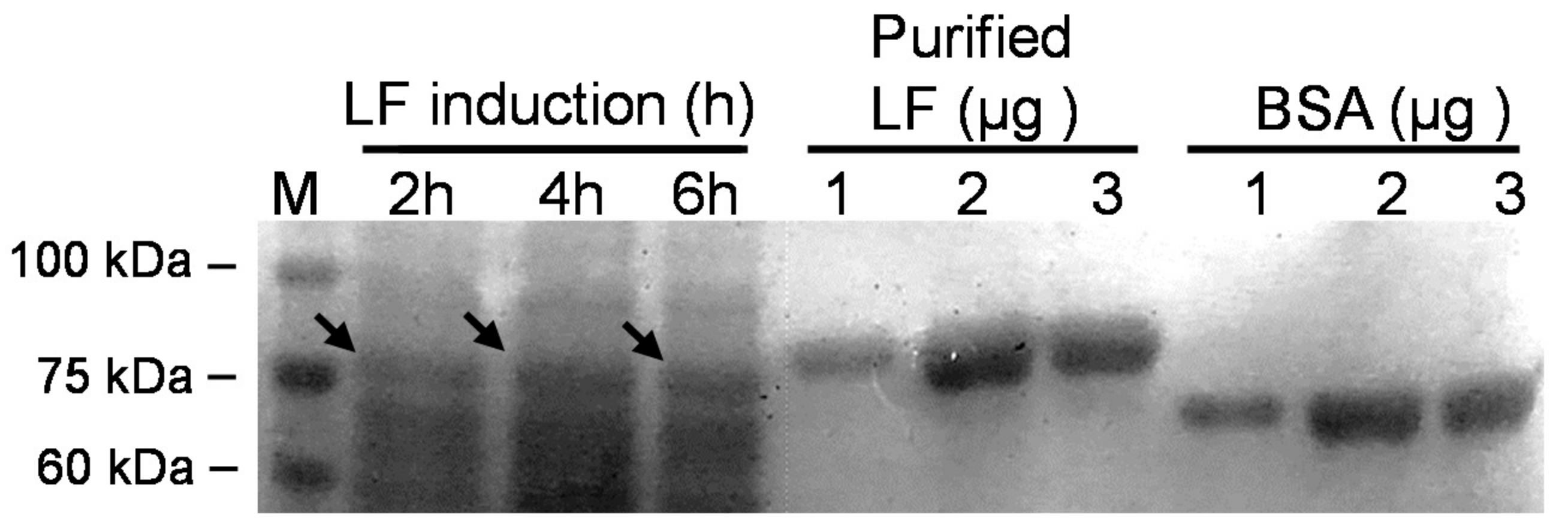
3.3. Expression of rLF Protein in Selected Probiotic Transformants
3.4. In Vitro Antibacterial Activities of rLFs from Two Probiotic Hosts
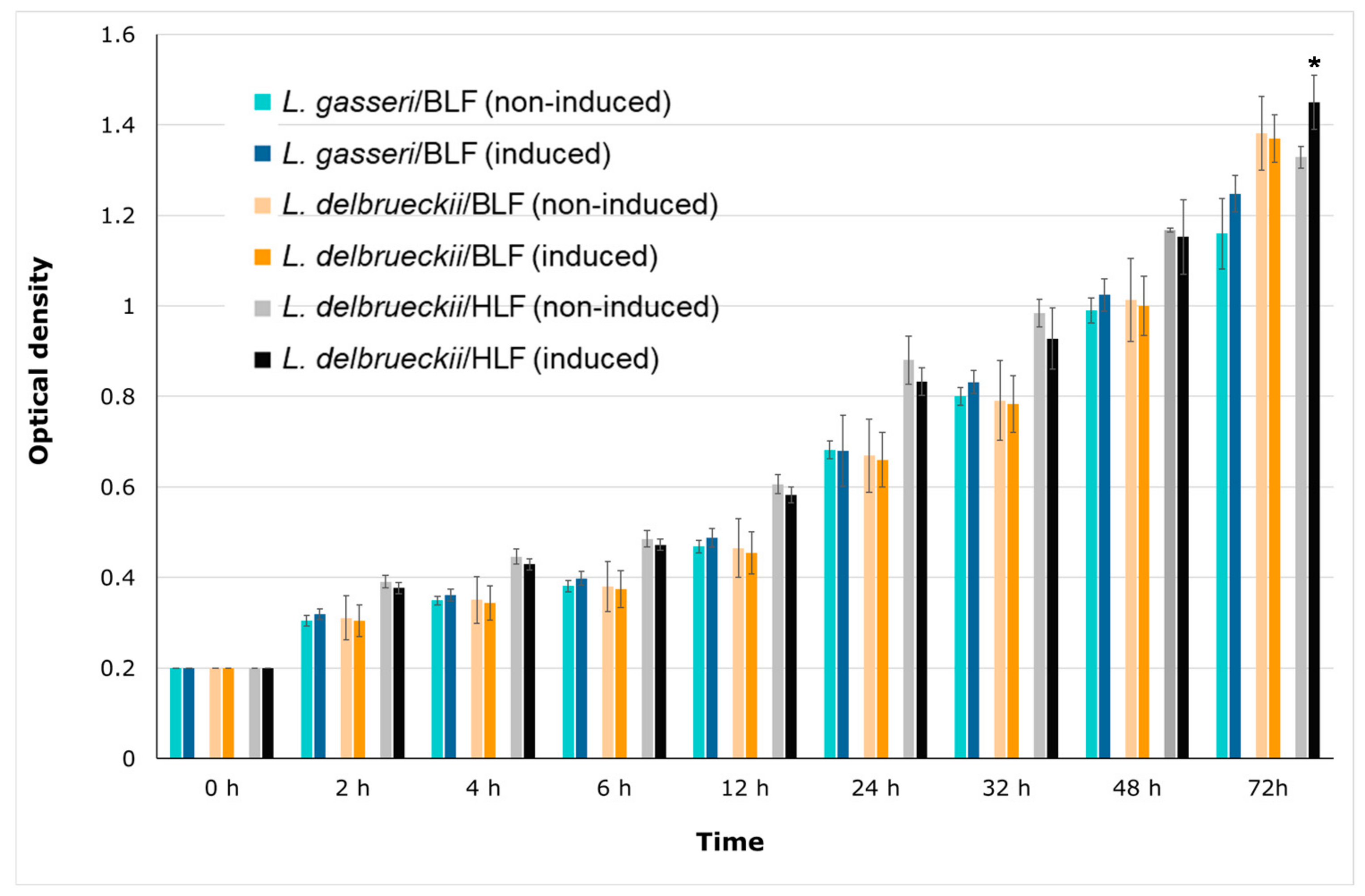
3.5. Effects of Recombinant Human or Bovine Lactoferrin on the Growth of Recombinant L. gasseri/BLF, L. delbrueckii/HLF, and L. delbrueckii/BLF
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Valenti, P.; Antonini, G. Lactoferrin: An important host defence against microbial and viral attack. Cell Mol. Life Sci. 2005, 62, 2576–2587. [Google Scholar] [CrossRef] [PubMed]
- Frioni, A.; Conte, M.P.; Cutone, A.; Longhi, C.; Musci, G.; di Patti, M.C.; di Patti, B.; Natalizi, T.; Marazzato, M.; Lepanto, M.S.; et al. Lactoferrin differently modulates the inflammatory response in epithelial models mimicking human inflammatory and infectious diseases. Biometals 2014, 27, 843–856. [Google Scholar] [CrossRef] [PubMed]
- Legrand, D. Overview of Lactoferrin as a Natural Immune Modulator. J. Pediatr. 2016, 173, S10–S15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, P.W.; Liu, Z.S.; Kuo, T.C.; Hsieh, M.C.; Li, Z.W. Prebiotic effects of bovine lactoferrin on specific probiotic bacteria. Biometals 2017, 30, 237–248. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.S.; Ohashi, M.; Tanaka, T.; Kumura, H.; Kim, G.Y.; Kwon, I.K.; Goh, J.S.; Shimazaki, K. Growth-promoting effects of lactoferrin on L. acidophilus and Bifidobacterium spp. Biometals 2004, 17, 279–283. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.M.; Kim, W.S.; Ito, T.; Kumura, H.; Shimazaki, K. Growth promotion and cell binding ability of bovine lactoferrin to Bifidobacterium longum. Anaerobe 2009, 15, 133–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pammi, M.; Suresh, G. Enteral lactoferrin supplementation for prevention of sepsis and necrotizing enterocolitis in preterm infants. Cochrane Database Syst. Rev. 2020, 3, CD007137. [Google Scholar] [CrossRef]
- Otsuki, K.; Yakuwa, K.; Sawada, M.; Hasegawa, A.; Sasaki, Y.; Mitsukawa, K.; Chiba, H.; Nagatsuka, M.; Saito, H.; Okai, T. Recombinant human lactoferrin has preventive effects on lipopolysaccharide-induced preterm delivery and production of inflammatory cytokines in mice. J. Perinat Med. 2005, 33, 320–323. [Google Scholar] [CrossRef]
- Yakuwa, K.; Otsuki, K.; Nakayama, K.; Hasegawa, A.; Sawada, M.; Mitsukawa, K.; Chiba, H.; Nagatsuka, M.; Okai, T. Recombinant human lactoferrin has a potential to suppresses uterine cervical ripening in preterm delivery in animal model. Arch. Gynecol. Obs. 2007, 275, 331–334. [Google Scholar] [CrossRef]
- Hu, P.; Zhao, F.; Zhu, W.; Wang, J. Effects of early-life lactoferrin intervention on growth performance, small intestinal function and gut microbiota in suckling piglets. Food Funct. 2019, 10, 5361–5373. [Google Scholar] [CrossRef]
- Superti, F.; De Seta, F. Warding Off Recurrent Yeast and Bacterial Vaginal Infections: Lactoferrin and Lactobacilli. Microorganisms 2020, 8, 130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.L.; Lai, Y.W.; Chen, C.S.; Chu, T.W.; Lin, W.; Yen, C.C.; Lin, M.F.; Tu, M.Y.; Chen, C.M. Probiotic Lactobacillus casei expressing human lactoferrin elevates antibacterial activity in the gastrointestinal tract. Biometals 2010, 23, 543–554. [Google Scholar] [CrossRef] [PubMed]
- Liao, H.; Liu, S.; Wang, H.; Su, H.; Liu, Z. Enhanced antifungal activity of bovine lactoferrin-producing probiotic Lactobacillus casei in the murine model of vulvovaginal candidiasis. BMC Microbiol. 2019, 19, 7. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Zong, X.; Han, B.; Li, Y.; Tang, L. Lactobacillus pentosus expressing porcine lactoferrin elevates antibacterial activity and improves the efficacy of vaccination against Aujeszky’s disease. Acta Vet. Hung. 2016, 64, 289–300. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.-G.; Yu, H.; Zhang, L.; Liu, M.; Qiao, X.-Y.; Cui, W.; Jiang, Y.P.; Wang, L.; Li, Y.J.; Tang, L.J. Probiotic properties of genetically engineered Lactobacillus plantarum producing porcine lactoferrin used as feed additive for piglets. Process Biochem. 2016, 51, 719–724. [Google Scholar] [CrossRef]
- De Vos, W.M.; Kuipers, O.P.; van der Meer, J.R.; Siezen, R.J. Maturation pathway of nisin and other lantibiotics: Post-translationally modified antimicrobial peptides exported by gram-positive bacteria. Mol. Microbiol. 1995, 17, 427–437. [Google Scholar] [CrossRef] [Green Version]
- Field, D.; Cotter, P.D.; Ross, R.P.; Hill, C. Bioengineering of the model lantibiotic nisin. Bioengineered 2015, 6, 187–192. [Google Scholar] [CrossRef] [Green Version]
- Shin, J.M.; Gwak, J.W.; Kamarajan, P.; Fenno, J.C.; Rickard, A.H.; Kapila, Y.L. Biomedical applications of nisin. J. Appl. Microbiol. 2016, 120, 1449–1465. [Google Scholar] [CrossRef] [Green Version]
- De Ruyter, P.G.; Kuipers, O.P.; de Vos, W.M. Controlled gene expression systems for Lactococcus lactis with the food-grade inducer nisin. Appl. Environ. Microbiol. 1996, 62, 3662–3667. [Google Scholar] [CrossRef] [Green Version]
- Kuipers, O.P.; Beerthuyzen, M.M.; de Ruyter, P.G.; Luesink, E.J.; de Vos, W.M. Autoregulation of nisin biosynthesis in Lactococcus lactis by signal transduction. J. Biol. Chem. 1995, 270, 27299–27304. [Google Scholar] [CrossRef] [Green Version]
- Chandrapati, S.; O’Sullivan, D.J. Nisin independent induction of the nisA promoter in Lactococcus lactis during growth in lactose or galactose. FEMS Microbiol. Lett. 1999, 170, 191–198. [Google Scholar] [CrossRef] [Green Version]
- Kleerebezem, M.; Beerthuyzen, M.M.; Vaughan, E.E.; de Vos, W.M.; Kuipers, O.P. Controlled gene expression systems for lactic acid bacteria: Transferable nisin-inducible expression cassettes for Lactococcus, Leuconostoc, and Lactobacillus spp. Appl. Environ. Microbiol. 1997, 63, 4581–4584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, P.W.; Jheng, T.T.; Shyu, C.L.; Mao, F.C. Antimicrobial potential for the combination of bovine lactoferrin or its hydrolysate with lactoferrin-resistant probiotics against foodborne pathogens. J. Dairy Sci. 2013, 96, 1438–1446. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.W.; Jheng, T.T.; Shyu, C.L.; Mao, F.C. Synergistic antibacterial efficacies of the combination of bovine lactoferrin or its hydrolysate with probiotic secretion in curbing the growth of meticillin-resistant Staphylococcus aureus. J. Med. Microbiol. 2013, 62, 1845–1851. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Wittouck, S.; Salvetti, E.; Franz, C.; Harris, H.M.B.; Mattarelli, P.; O’Toole, P.W.; Pot, B.; Vandamme, P.; Walter, J.; et al. A taxonomic note on the genus Lactobacillus: Description of 23 novel genera, emended description of the genus Lactobacillus Beijerinck 1901, and union of Lactobacillaceae and Leuconostocaceae. Int. J. Syst. Evol. Microbiol. 2020, 70, 2782–2858. [Google Scholar] [CrossRef] [PubMed]
- Sambrook, J.; Fritsch, E.; Maniatis, T.J.N.Y. Molecular Cloning: A Laboratory Manual, 2nd ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1989. [Google Scholar]
- Shim, Y.H.; Lee, S.J.; Lee, J.W. Antimicrobial activity of lactobacillus strains against uropathogens. Pediatr. Int. 2016, 58, 1009–1013. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.C.; Lai, T.M.; Hsieh, Y.M. Evaluation of Lactobacilli for Antagonistic Activity Against the Growth, Adhesion and Invasion of Klebsiella pneumoniae and Gardnerella vaginalis. Indian J. Microbiol. 2019, 59, 81–89. [Google Scholar] [CrossRef]
- Woodman, T.; Strunk, T.; Patole, S.; Hartmann, B.; Simmer, K.; Currie, A. Effects of lactoferrin on neonatal pathogens and Bifidobacterium breve in human breast milk. PLoS ONE 2018, 13, e0201819. [Google Scholar] [CrossRef]
- Mathipa, M.G.; Thantsha, M.S.J. Probiotic engineering: Towards development of robust probiotic strains with enhanced functional properties and for targeted control of enteric pathogens. Gut Pathog. 2017, 9, 28. [Google Scholar] [CrossRef]
- Amalaradjou, M.A.R.; Bhunia, A.K.J.B. Bioengineered probiotics, a strategic approach to control enteric infections. Bioengineered 2013, 4, 379–387. [Google Scholar] [CrossRef] [Green Version]
- Bober, J.R.; Beisel, C.L.; Nair, N.U. Synthetic biology approaches to engineer probiotics and members of the human microbiota for biomedical applications. Annu. Rev. Biomed. Eng. 2018, 20, 277–300. [Google Scholar] [CrossRef] [PubMed]
- Alkhatib, Z.; Abts, A.; Mavaro, A.; Schmitt, L.; Smits, S.H. Lantibiotics: How do producers become self-protected? J. Biotechnol. 2012, 159, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Stein, T.; Heinzmann, S.; Solovieva, I.; Entian, K.D. Function of Lactococcus lactis nisin immunity genes nisI and nisFEG after coordinated expression in the surrogate host Bacillus subtilis. J. Biol. Chem. 2003, 278, 89–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuipers, O.P.; de Ruyter, P.G.; Kleerebezem, M.; de Vos, W.M.J. Quorum sensing-controlled gene expression in lactic acid bacteria. J. Biotechnol. 1998, 64, 15–21. [Google Scholar] [CrossRef] [Green Version]
- Abu-Qarn, M.; Eichler, J.; Sharon, N. Not just for Eukarya anymore: Protein glycosylation in Bacteria and Archaea. Curr. Opin. Struct. Biol. 2008, 18, 544–550. [Google Scholar] [CrossRef]
- Szymanski, C.M.; Wren, B.W. Protein glycosylation in bacterial mucosal pathogens. Nat. Rev. Microbiol. 2005, 3, 225–237. [Google Scholar] [CrossRef]
- Lebeer, S.; Claes, I.J.; Balog, C.I.; Schoofs, G.; Verhoeven, T.L.; Nys, K.; von Ossowski, I.; de Vos, W.M.; Tytgat, H.L.; Agostinis, P.; et al. The major secreted protein Msp1/p75 is O-glycosylated in Lactobacillus rhamnosus GG. Microb Cell Fact. 2012, 11, 15. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; O’Sullivan, D.J. Heterologous expression of the Lactococcus lactis bacteriocin, nisin, in a dairy Enterococcus strain. Appl. Environ. Microbiol. 2002, 68, 3392–3400. [Google Scholar] [CrossRef] [Green Version]
- Fang, C.-Y.; Lu, J.-R.; Chen, B.-J.; Wu, C.; Chen, Y.-P.; Chen, M.-J.J. Selection of uremic toxin-reducing probiotics in vitro and in vivo. J. Funct. Foods 2014, 7, 407–415. [Google Scholar] [CrossRef]
- Jang, S.E.; Jeong, J.J.; Choi, S.Y.; Kim, H.; Han, M.J.; Kim, D.H. Lactobacillus rhamnosus HN001 and Lactobacillus acidophilus La-14 Attenuate Gardnerella vaginalis-Infected Bacterial Vaginosis in Mice. Nutrients 2017, 9, 531. [Google Scholar] [CrossRef] [Green Version]
- Bertuccini, L.; Russo, R.; Iosi, F.; Superti, F.J. Lactobacilli and lactoferrin: Biotherapeutic effects for vaginal health. J. Funct. Foods 2018, 45, 86–94. [Google Scholar] [CrossRef]
- Jimenez, E.; Fernandez, L.; Maldonado, A.; Martin, R.; Olivares, M.; Xaus, J.; Rodríguez, J.M. Oral administration of Lactobacillus strains isolated from breast milk as an alternative for the treatment of infectious mastitis during lactation. Appl. Environ. Microbiol. 2008, 74, 4650–4655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arroyo, R.; Martín, V.; Maldonado, A.; Jiménez, E.; Fernández, L.; Rodríguez, J.M. Treatment of infectious mastitis during lactation: Antibiotics versus oral administration of Lactobacilli isolated from breast milk. Clin. Infect. Dis. 2010, 50, 1551–1558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandez, L.; Arroyo, R.; Espinosa, I.; Marin, M.; Jimenez, E.; Rodriguez, J.M. Probiotics for human lactational mastitis. Benef. Microbes 2014, 5, 169–183. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Upadhyay, V.; Upadhyay, A.K.; Singh, S.M.; Panda, A.K. Protein recovery from inclusion bodies of Escherichia coli using mild solubilization process. Microb Cell Fact. 2015, 14, 41. [Google Scholar] [CrossRef] [Green Version]
- Palmer, I.; Wingfield, P.T. Preparation and extraction of insoluble (inclusion-body) proteins from Escherichia coli. Curr. Protoc. Protein Sci. 2012, 6, 3. [Google Scholar] [CrossRef] [Green Version]
- Berstad, A.; Raa, J.; Midtvedt, T.; Valeur, J. Probiotic lactic acid bacteria—The fledgling cuckoos of the gut? Microb. Ecol. Health Dis. 2016, 27, 31557. [Google Scholar] [CrossRef]
- Besselink, M.G.; van Santvoort, H.C.; Buskens, E.; Boermeester, M.A.; van Goor, H.; Timmerman, H.M.; Nieuwenhuijs, V.B.; Bollen, T.L.; van Ramshorst, B.; Witteman, B.J.M.; et al. Probiotic prophylaxis in predicted severe acute pancreatitis: A randomised, double-blind, placebo-controlled trial. Lancet 2008, 371, 651–659. [Google Scholar] [CrossRef] [Green Version]
- Angelakis, E.; Merhej, V.; Raoult, D. Related actions of probiotics and antibiotics on gut microbiota and weight modification. Lancet Infect. Dis. 2013, 13, 889–899. [Google Scholar] [CrossRef]
- Mathur, S.; Singh, R. Antibiotic resistance in food lactic acid bacteria—A review. Int. J. Food Microbiol. 2005, 105, 281–295. [Google Scholar] [CrossRef]
- Arnold, R.; Brewer, M.; Gauthier, J.J. Bactericidal activity of human lactoferrin: Sensitivity of a variety of microorganisms. Infect. Immun. 1980, 28, 893–898. [Google Scholar]
- Chen, P.W.; Ho, S.P.; Shyu, C.L.; Mao, F.C. Effects of bovine lactoferrin hydrolysate on the in vitro antimicrobial susceptibility of Escherichia coli strains isolated from baby pigs. Am. J. Vet. Res. 2004, 65, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Kanwar, J.R.; Roy, K.; Patel, Y.; Zhou, S.F.; Singh, M.R.; Singh, D.; Nasir, M.; Sehgal, R.; Sehgal, A.; Singh, R.S.; et al. Multifunctional iron bound lactoferrin and nanomedicinal approaches to enhance its bioactive functions. Molecules 2015, 20, 9703–9731. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.W.; Ku, Y.W.; Chu, F.Y. Influence of bovine lactoferrin on the growth of selected probiotic bacteria under aerobic conditions. Biometals 2014, 27, 905–914. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plasmid | |||
|---|---|---|---|
| Probiotic | pNZ8148 /HLF | pNZ8148 /BLF | pNZ8148 /PLF |
| Lactobacillus delbrueckii | + 1 | + | − 2 |
| Pediococcus pentosaceus | − | + | − |
| Bifidobacterium angulatum | − | + | + |
| Lactobacillus paraplantarum | + | + | − |
| Bifidobacterium breve | − | + | − |
| Bifidobacterium catenulatum | − | − | + |
| Lactobacillus rhamnosus | + | − | − |
| Lactobacillus gasseri, strain A (Laboratory stock) | − | + | − |
| Inhibitory Zone Diameter (mm) 1 | ||||
|---|---|---|---|---|
| Lactobacillus Strains | S. aureus | En. faecalis | S. enterica | E. coli |
| L. delbrueckii/HLF (induced) 2 | 14.8 ± 0.7 *** | 14.8 ± 0.5 *** | 15.8 ± 1.4 *** | 15.0 ± 2.0 ** |
| L. delbrueckii/HLF (non-induced) 2 | 12.7 ± 0.9 | 12.5 ± 1.0 | 12.9 ± 0.7 | 12.4 ± 0.9 |
| L. delbrueckii/BLF (induced) 2 | 14.5 ± 1.3 *** | 15.3 ± 1.1 *** | 15.3 ± 1.7 ** | 14.9 ± 1.3 ** |
| L. delbrueckii/BLF (non-induced) 2 | 12.5 ± 0.9 | 12.8 ± 1.1 | 12.7 ± 0.9 | 13.0 ± 0.9 |
| L. gasseri/BLF (induced) 2 | 14.8 ± 0.3 *** | 15 ±0.9 *** | 14.9 ± 0.9 *** | 15.1 ± 0.9 *** |
| L. gasseri/BLF (non-induced) 2 | 12.4 ± 0.3 | 13.1 ± 0.4 | 13.1 ± 0.4 | 13.4 ± 0.8 |
| Blank control | 10 ± 0 | 10 ± 0 | 10 ± 0 | 10 ± 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Z.-S.; Lin, C.-F.; Lee, C.-P.; Hsieh, M.-C.; Lu, H.-F.; Chen, Y.-F.; Ku, Y.-W.; Chen, P.-W. A Single Plasmid of Nisin-Controlled Bovine and Human Lactoferrin Expressing Elevated Antibacterial Activity of Lactoferrin-Resistant Probiotic Strains. Antibiotics 2021, 10, 120. https://doi.org/10.3390/antibiotics10020120
Liu Z-S, Lin C-F, Lee C-P, Hsieh M-C, Lu H-F, Chen Y-F, Ku Y-W, Chen P-W. A Single Plasmid of Nisin-Controlled Bovine and Human Lactoferrin Expressing Elevated Antibacterial Activity of Lactoferrin-Resistant Probiotic Strains. Antibiotics. 2021; 10(2):120. https://doi.org/10.3390/antibiotics10020120
Chicago/Turabian StyleLiu, Zhen-Shu, Chuen-Fu Lin, Chung-Pei Lee, Min-Chi Hsieh, Hung-Fu Lu, Ying-Fang Chen, Yu-We Ku, and Po-Wen Chen. 2021. "A Single Plasmid of Nisin-Controlled Bovine and Human Lactoferrin Expressing Elevated Antibacterial Activity of Lactoferrin-Resistant Probiotic Strains" Antibiotics 10, no. 2: 120. https://doi.org/10.3390/antibiotics10020120