
Biofunctional Peptide-Modified Extracellular Vesicles Enable Effective Intracellular Delivery via the Induction of Macropinocytosis

1
Department of Biological Science, Graduate School of Science, Osaka Prefecture University, 1-1, Gakuen-cho, Naka-ku, Sakai, Osaka 599-8531, Japan
2
NanoSquare Research Institute, Osaka Prefecture University, 1-2, Gakuen-cho, Naka-ku, Sakai, Osaka 599-8570, Japan
Processes 2021, 9(2), 224; https://doi.org/10.3390/pr9020224
Submission received: 20 November 2020
/
Revised: 17 January 2021
/
Accepted: 20 January 2021
/
Published: 25 January 2021
(This article belongs to the Special Issue Advances of Peptide Engineering)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:We previously reported that macropinocytosis (accompanied by actin reorganization, ruffling of the plasma membrane, and engulfment of large volumes of extracellular fluid) is an important process for the cellular uptake of extracellular vesicles, exosomes. Accordingly, we developed techniques to induce macropinocytosis by the modification of biofunctional peptides on exosomal membranes, thereby enhancing their cellular uptake. Arginine-rich cell-penetrating peptides have been shown to induce macropinocytosis via proteoglycans; accordingly, we developed peptide-modified exosomes that could actively induce macropinocytotic uptake by cells. In addition, the activation of EGFR induces macropinocytosis; based on this knowledge, we developed artificial leucine-zipper peptide (K4)-modified exosomes. These exosomes can recognize E3 sequence-fused EGFR (E3-EGFR), leading to the clustering and activation of E3-EGFR by coiled-coil formation (E3/K4), which induces cellular exosome uptake by macropinocytosis. In addition, modification of pH-sensitive fusogenic peptides (e.g., GALA) also enhances the cytosolic release of exosomal contents. These experimental techniques and findings using biofunctional peptides have contributed to the development of exosome-based intracellular delivery systems.
1. Introduction
Extracellular vesicles (EVs), including exosomes and microvesicles, are secreted from most cells in the human body. EVs can carry biofunctional molecules, such as microRNAs and enzymes, and play crucial roles in cell-to-cell communication and disease progression (e.g., cancer); further, recent EV research has received significant attention for its role in maintaining healthy homeostasis and in disease progression [1,2,3]. For example, exosome-mediated delivery of miR-181c triggers the breakdown of the blood-brain barrier (BBB), leading to brain metastasis [4], and EVs encapsulating matrix metalloproteinase 1 mRNA have been observed in ovarian cancer metastasis [5]. Therefore, experimental techniques, such as molecular weight measurement, antibody-based detection, real-time polymerase chain reaction, and polymer-based reverse-phase glycoprotein array, have been used to detect EV-encapsulated miRNA and protein in cancer diagnosis [3]. Exosomal cargo molecules have been observed to be cancer biomarkers as follows: pancreatic cancer, glycan-1, and zinc transporter 4; colorectal cancer, copine III and miR-6803-5p; non-small cell lung cancer, alpha-2-HS-glycoprotein, and extracellular matrix protein 1; and prostate cancer, miR-1246 [3]. Further, microvesicles released from blood cells can potentially be used to diagnose acute coronary syndrome, ST-elevation myocardial infarction, cardiac remodeling, type2 diabetes mellitus, diabetic retinopathy, and other cardiometabolic diseases [6]. In addition, EV-mediated intracellular delivery of therapeutic molecules is expected to be a next-generation cell-derived tool with pharmaceutical advantages such as immunological control, expression of functional sequences in/on EVs, the encapsulation of artificially designed and synthesized molecules, low cytotoxicity, BBB penetration, and brain targeting, and the effective use of cell-to-cell communication routes. Further interdisciplinary research in medical, pharmaceutical, engineering, and physicochemical fields to develop EV-delivery techniques will enable the intravital detection of disease progression and the manipulation of cellular functions.
2. Characteristics of EVs
EVs include exosomes, microvesicles, and apoptotic bodies, and each vesicle can be secreted from cells via different mechanisms and pathways [1,2,7,8]. Apoptotic bodies can be generated during the process of apoptotic cell death, and they contain organelles and cellular components such as chromatin [9]. The diameter of the vesicles is approximately 800–5000 nm [1,6,8,9]. Apoptotic bodies are eventually engulfed by phagocytes in a process known as efferocytosis [9]. During apoptosis, membrane blebbing occurs in the majority of the cells; however, some cells exhibit other types of membrane protrusions such as microtubule spikes, apoptopodia, and beaded apoptopodia. These membrane protrusions eventually lead to the formation and release of apoptotic bodies [9]. Biofunctional molecules encapsulated in apoptotic bodies can be acquired by recipient cells, leading to the transfer of macromolecules and a biological response [9].
Microvesicles can be generated by plasma membrane budding [1,7,8] and can encapsulate not only membrane proteins and lipids but also nucleic acids including mRNAs, microRNAs, and long non-coding RNAs [6]. The vesicles (0.1–1 μm diameter) bud as small membrane protrusions surrounding a small portion of the cytoplasm and are released to the extracellular milieu by calpain activation, calcium influx, and cytoskeleton reorganization [6,10]. Microvesicles play various roles in intercellular communication, signal transduction, and the immune regulation of (a) coagulation, by coordinating platelet, macrophage, and neutrophil functions; (b) inflammatory diseases, by the release of cytokines; and (c) tumor progression, by facilitating the migration and invasion of cancer cells to trigger metastasis [6,10].
On the other hand, exosomes can be formed within cells; multivesicular endosomes (MVEs), formed by internal budding, encapsulate cytosolic molecules, which is followed by fusion of the MVE membrane with the plasma membrane to extracellularly secrete the formed vesicles (30–200 nm) [1,2]. Inhibitory G protein (Gi)-coupled sphingosine 1-phosphate (S1P) receptors have been shown to regulate MVE maturation to exosomes, and the receptors are essential for cargo sorting and maturation of MVEs via the endosomal sorting complex required for transport (ESCRT)-independent pathway [11]. Inhibitors of sphingosine kinase and reduced expression of S1P receptors affect cargo loading into exosomes [11]. siRNA-mediated knockdown of sphingosine kinase 2, but not sphingosine kinase 1, have been shown to result in a reduction of cargo content in exosomes [12]. Continuous activation of the S1P receptor and subsequent downstream G protein signaling to Gβγ subunits/Rho family GTPases have been observed to regulate F-actin formation on MVEs for cargo sorting into exosomal intralumenal vesicles [13]. Encapsulation of the “message” such as cytosolic microRNAs and proteins into exosomes is essential for transporting biofunctional molecules and intercellular communication, and detailed mechanisms should be elucidated to further understand their role in human homeostasis, mechanisms of disease progression, and their regulation.
3. Cellular Exosomes Uptake and the Importance of the Macropinocytosis Pathway
Cellular uptake of exosomes mediated by receptor-ligand complexes has been observed in the following cases: the interaction between C-type lectin (lectin family) receptor in dendritic cells and C-type lectin on exosomes [14], the interaction between sialoadhesin (CD169; Siglec-1) expressed on macrophages and B-cell-derived exosomal α-2,3-linked sialic acids [15], the interaction between siglec-3 expressed on HeLa cells or antigen-presenting cells and sialic acids on mesenchymal stem cell exosomes [16], the interaction between CD44, CD11, CD54, and CD49d (adhesion molecules) expressed on leukocytes and CD81 and CD9 of pancreatic adenocarcinoma exosomes [17], cellular uptake of breast cancer exosomes expressing integrins α6β4 and pancreatic cancer exosomes expressing αvβ5 into lung fibroblast and liver macrophage [18], the interaction between macrophage exosomes expressing LFA-1 and brain endothelial cells expressing ICAM-1 [19], interaction between heparin sulfate proteoglycans expressed on glioblastoma multiforme and myeloma cells and myeloma exosomes expressing fibronectin [20,21], and between exosomes expressing phosphatidylserine and dendritic cells expressing TIM receptors [22,23]. In exosome-mediated cell-to-cell communication, the receptor-ligand interaction is essential to determine the efficacy of exosome accumulation in targeted cells. Clathrin-mediated endocytosis is a typical endocytosis pathway that occurs by the activation of receptors on recipient cells. However, endosomal vesicles coated with clathrin triskelion and AP2 adaptors are limited by size (approximately 120 nm, because of the triskelion coating), and the cellular uptake efficacy of the exosomes might be affected by such endosomal vesicular formation [24].
Therefore, we focused on the role of macropinocytosis in the cellular uptake of exosomes. Macropinocytosis is an endocytosis process that involves actin polymerization in the cell periphery, increased actin-mediated membrane ruffling, and the cellular uptake by macropinosomes, which is distinct from the clathrin-mediated endocytosis pathway [25,26]. Our research team found that macropinocytosis significantly enhanced the cellular uptake of exosomes [27]. It has been reported that increased macropinocytotic activity in tumors is an essential mechanism for the uptake of nutrients [28]; therefore, our findings could contribute toward the development of targeted therapeutics in cancers. For example, in pancreatic adenocarcinoma, the cellular uptake of exosomes through macropinocytosis is ~10-fold higher in MIA PaCa-2 cells, which are homozygous for the KRASG12C allele (oncogenic Ras-expressing), compared to the BxPC-3 cells, which express wild-type KRAS [27]. Saporin is a ribosome-inactivating toxin protein with potential utility as a cancer therapeutic [29]. Saporin-encapsulated exosomes showed enhanced cytotoxicity against epidermal growth factor receptor (EGFR)-activated cancer cells (the A431 cell line) by the induction of macropinocytosis, suggesting that exosomes may have potential applications in drug delivery and the treatment of malignant tumors.
4. Cell Targeting and Intracellular Delivery Systems Using Biofunctional Peptide-Modified Exosomes
Cell-to-cell communication is an active research topic, and exosomes are an important component of this phenomenon. As mentioned above, exosomes are potential next-generation biological tools for the targeted delivery of therapeutic molecules. They possess multiple advantages, including (i) infinite secretion, (ii) the artificial encapsulation of biofunctional molecules, (iii) the controlled expression of artificial proteins in exosomal membranes, (iv) low cytotoxicity, (v) regulated immunogenicity, (vi) effective usage of cell-to-cell communication routes, and (vii) the possibility of targeting the brain owing to BBB permeability. However, there are certain disadvantages associated with exosome-based delivery systems, which include poor cell targeting, low cellular uptake efficacy, and insufficient cytosolic release of exosomal contents. Therefore, further improvements are required in the design of sophisticated exosome-based delivery systems.
We have previously developed and successfully demonstrated a novel drug delivery system based on biofunctional peptide-modified exosomes (Figure 1); this includes arginine-rich cell-penetrating peptide-modified exosomes for the active induction of macropinocytosis and effective intracellular delivery of therapeutic molecules, a pH-sensitive fusogenic peptide for enhanced cytosolic release of exosomal contents, and a receptor target system using artificial coiled-coil peptide-modified on exosomal membranes.
4.1. Arginine-Rich Peptide-Modified Exosomes for the Active Induction of Macropinocytosis and Effective Intracellular Delivery of Therapeutic Molecules
As described above, we found that active macropinocytosis is essential for the efficient cellular uptake of exosomes. Therefore, we enhanced the cellular uptake of exosomes by exploiting the biological functionality of arginine-rich cell-penetrating peptides (CPPs); these peptides can efficiently induce macropinocytosis by modifying the peptides on exosomal membranes [30] (Figure 2).
Receptor-mediated and clathrin-mediated endocytosis have been reported to be major pathways for the cellular uptake of exosomes; however, it has a limited size (maximum ~120 nm) for vesicle formation during endocytotic cellular uptake. Moreover, cellular membrane proteins and glycosaminoglycans further reduce the cargo size. This size limitation reduces the efficacy of endocytosis-mediated cellular uptake of exosomes, and negatively charged exosomes (zeta-potential is minus several tens of millivolts) are difficult to accumulate on the negatively charged cellular membranes. Macropinocytosis induces the engulfment of large volumes of extracellular fluid, and this pathway enables the cellular uptake of large-sized cargo (>1 μm). Our research team reported that increased macropinocytosis via activation of cancer-related receptors and oncogenic K-Ras expression enhances the cellular uptake efficacy of exosomes in tumor cells, as described above [27].
Arginine-rich CPPs can induce macropinocytosis, leading to the efficient cellular uptake of the peptides [31]. We found that membrane-associated proteoglycans play a crucial role in the induction of macropinocytosis by CPPs, and the activation of small G-protein Rac1 and reorganization of actin (lamellipodia and membrane ruffling) were observed within a few minutes after treatment of the peptides; this leads to the efficient cellular uptake of the peptides and their cytosolic translocation [32,33,34]. We also discovered that arginine-rich CPPs induce macropinocytosis by clustering syndecan-4 and PKCα binding to the syndecan-4 V domain [35].
The D-form of the octaarginine peptide (R8) is arginine-rich and can induce efficient macropinocytosis [31,32,33,34,35]. To modify the R8 peptides on exosomal membranes, we stearylated the N-terminus of the peptides and thereby inserted the hydrophobic moiety into the exosomal membrane (Figure 2a) [30]. The functionality of exosomal membrane proteins might be conserved even after anchoring the stearyl peptides on the membranes. In that study, exosomes expressing the tetraspanin CD63-green fluorescent protein (GFP) fusion (CD63-GFP) marker were used to assess cellular uptake. Modifying the membrane surface of exosomes with stearyl-R8 peptides significantly enhanced their uptake via macropinocytosis, with syndecan-4 clustering on the recipient cell membranes [30]. The dependency of cellular uptake efficacy on the peptide concentration on exosomal membranes was also shown (e.g., treatment of 16 μM stearyl-R8 peptides increased 33-fold compared to the cellular uptake of the unmodified exosomes) [30]. Treatment with the macropinocytosis inhibitor, 5-(N-ethyl-N-isopropyl)amirolide (EIPA), significantly reduced the cellular uptake of stearyl-R8-modified CD63-GFP exosomes. On the other hand, the inhibitor did not affect the cellular uptake of exosomes without peptide modifications [30]. The use of the stearyl moiety for anchoring functional peptides on exosomal membranes is promising, and this methodology could foster the development of exosome-based intracellular delivery systems with advanced exosomal functionality.
4.2. The Number of Arginine Residues in CPPs of Modified Exosomal Membrane Affects the Cytosolic Release of Exosomal Contents and Biological Activity
As described above, oligoarginine CPPs have been shown to induce macropinocytosis via proteoglycans on plasma membranes [32]. In addition, the number of arginine residues in the peptide sequence has been shown to affect the efficacy of macropinocytosis induction and cellular uptake of exosomes [32,36]. In this study, we examined whether the number of arginine residues in the peptide sequence affected macropinocytosis induction and the cellular uptake of the peptide-modified exosomes [37]. We prepared oligoarginine peptide (Rn; n = 4–16)-modified exosomes with a cross-linker, (N-ε-maleimidocaproyl-oxysulfosuccinimide ester, sulfo-EMCS), by simple mixing of the peptides and targeted exosomes (Figure 2b). The chemical linker covalently binds to the amino group of the exosomal membrane proteins. We then evaluated the relative cellular uptake of Rn-conjugated CD63-GFP-exosomes (20 μg/mL) for 24 h at 37 °C in CHO-K1 cells (derived from Chinese hamster ovary). We observed that the number of arginine residues in the Rn-conjugated CD63-GFP-exosomes had an effect on macropinocytosis induction and improved the efficacy of cellular uptake by 14-fold (R4), 29-fold (R8), or 18-fold (R16) compared to the cellular uptake of the unmodified exosomes [37]. Enhanced lamellipodia formation was also observed on exosomal membranes after arginine-peptide modification [37]. On the other hand, oligoarginine peptides without the chemical linker did not increase the efficiency of cellular uptake of the exosomes. In addition, we artificially encapsulated the ribosome-inactivating protein, saporin, having anti-cancer bioactivity, in exosomes (saporin-exosome) using an electroporation system [37]. Saporin (molecular weight: ~28,000) belongs to the N-glycosidase family of toxins that can specifically and irreversibly inactivate large ribosomal subunits by depurinating specific adenine bases; once it enters the cytosol, saporin can cause cell death by blocking protein synthesis [29]. Therefore, saporin is widely used as a toxin-based anti-cancer drug, and its activity can be further enhanced by antibody-conjugation for cancer-receptor targeting [29]. Interestingly, the saporin-exosome with modified R16 peptides showed higher biological activity (83% cells were dead (48 h treatment)) than that of the R8 peptide (only 17% cells were dead), even though the cellular uptake efficacy of R16-modified exosomes was lower than that of the R8-modified exosomes [37]. This suggests that the cytosolic release efficacy differs depending on the specific modification to CPP arginine number. These results suggest that a balance in efficiency should be struck between cellular exosome uptake and the cytosolic release of exosomal contents to attain sophisticated intracellular delivery using biofunctional peptide-modified exosomes. Further, these can be adapted for the delivery of other therapeutic molecules.
4.3. Targeted Receptor Clustering and Activation by Coiled-Coil Peptide-Modified Exosomes for Achieving Effective Cellular Uptake
The clustering of cellular receptors on the plasma membrane often induces receptor activation. The ability to control this clustering is potentially valuable for manipulating cellular functions and studying signal transduction mechanisms. Our research team has already proposed an approach for controlling receptor activation using coiled-coil recognition elements to stimulate the dimerization of cell-surface receptors [38]. Here, we utilized an artificial receptor system controlled by heterodimeric coiled-coil peptides to achieve targeted receptor assembly on the exosomes to act as scaffolds that control exosome-based intracellular delivery of therapeutic molecules (Figure 3) [39].
In that particular study, we developed a technique for effectively targeting receptors using artificial leucine zipper peptides (E3/K4), which were developed by Hodges and coworkers [40,41], to form heterodimeric-coiled coils. We demonstrated this using an EGFR model; the successful clustering of receptor molecules on cell surfaces led to actin reorganization and stimulated the cellular uptake of exosomes via macropinocytosis. Modification of the exosomal surface using an artificial leucine-zipper K4 peptide (amino acid sequence: KIAALKEKIAALKEKIAALKEKIAALKE) allowed for the recognition of E3 (amino acid sequence: EIAALEKEIAALEKEIAALEK)-fused epidermal growth factor receptor (E3-EGFR) on the cell membrane. As mentioned previously, in order to modify the K4 peptides on exosomal membranes, we stearylated the N-terminus of the peptides to insert the hydrophobic moiety into the exosomal membrane [39]. Clustering of the E3-EGFR molecules on cell surfaces by treatment with K4 peptide-modified exosomes led to the activation of the receptors (autophosphorylation reaction), signal transduction, and actin reorganization (lamellipodia formation), thereby stimulating the cellular uptake of exosomes through a specific form of endocytosis (i.e., macropinocytosis) in human breast adenocarcinoma-derived MDA-MB-231 cells (e.g., treatment of 5 μM stearyl-K4 peptides increased 2-fold compared to the cellular uptake of the unmodified exosomes) [39]. On the other hand, EGFR without an E3 sequence was not activated by treatment with K4 peptide-modified exosomes. The macropinocytosis inhibitor, EIPA, also significantly reduced the cellular uptake of K4 peptide-modified exosomes via E3-EGFR [39]. In this study, we adopted EGFR as a model; however, a similar approach may be possible using other receptors (e.g., chemokine receptors and proteoglycans). Our approach has a high potential for tumor targeting, where the induction of macropinocytosis by receptor clustering (e.g., proteoglycans (syndecan-4) and CXCR4) could be attained by displayed peptides on exosomal surfaces that could then stimulate exosome uptake into the tumor cells. Macropinocytosis is an effective endocytic approach for engulfing exosomes. Therefore, this approach could enhance tumor therapy by using exosomes with specific receptor targeting.
4.4. Enhanced Cytosolic Release of Exosomal Contents Using a pH-Sensitive Fusogenic Peptide
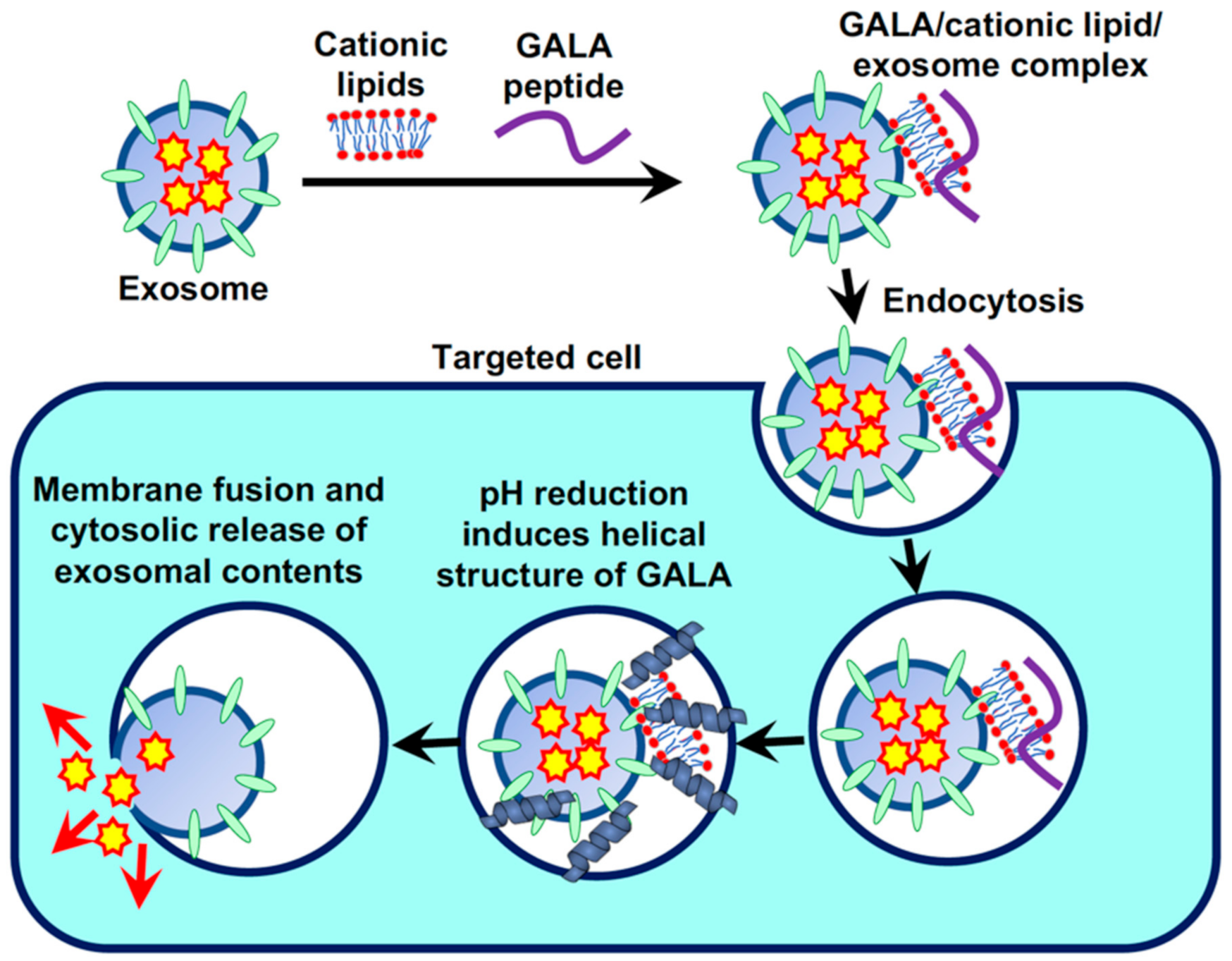
To improve the efficiency of cytosolic release of exosomal contents following the cellular uptake of exosomes, we studied the effects of the pH-sensitive fusogenic peptide GALA on endosomal and exosomal membranes and its role in the intracellular trafficking of exosomes via endocytosis [42]. In this study, we used a 30-residue amphipathic peptide, GALA, with a repeating sequence of glutamic acid-alanine-leucine-alanine (the amino acid sequence of GALA is WEAALAEALAEALAEHLAEALAEALEALAA); this sequence mimics the viral fusion protein sequences that mediate the escape of the viral gene from the acidic endosomes to the cytosol [43,44,45]. The secondary structure of the GALA peptide changes from random to helical in reduced pH (ranging from 7 to 5), leading to membrane destabilization and fusion [43,44,45]. We have previously reported the cytosolic delivery of avidin by using the GALA peptide in combination with cationic lipids to enhance the accumulation of negatively charged GALA peptides on the plasma membrane and, thereby, cellular uptake [45,46]. Cationic lipids can act similar to glue for pasting exosomes to targeted cells (as mentioned earlier, exosomal membranes are also negatively charged), and we examined the cellular uptake of exosomes using a commercially available transfection reagent, Lipofectamine LTX, which contains cationic lipids. The combination treatment of exosomes with the GALA peptide and cationic lipids shows efficient cellular uptake of both peptides and the exosomes. This leads to the enhanced cytosolic release of exosomal contents since the low pH of endosomes results in increased membrane disruption and fusion by the GALA peptides (Figure 4) [42]. In this experimental technique, a high concentration of cationic lipids induces cytotoxicity (for instance, 8.0% (v/v) Lipofectamine LTX treatment resulted in ~90% cell death); thus, optimization of the combinational treatment is required to prevent cytotoxic effects (for instance, 2.0% (v/v) Lipofectamine LTX treatment resulted in approximately 11-fold enhanced cellular uptake without any cytotoxicity) [42]. Dextran (molecular weight: 70,000) was used as a model macromolecular cargo in exosomes (encapsulated by electroporation), and the effective cytosolic release of fluorescently labeled dextran originally encapsulated in exosomes could be observed in the combinational treatment with the GALA peptides and cationic lipids [42]. In the absence of cationic lipids, cellular uptake efficacy of the GALA peptides was significantly reduced, leading to low efficacy of exosomal and endosomal cytosolic release in cells [42]. In addition, treatment with NH4Cl to prevent endosomal pH reduction decreased the cytosolic release of fluorescently labeled dextran from the delivered exosomes, even in the combinational treatment with GALA peptides and cationic lipids [42]. Saporin encapsulated in exosomes also showed significant cytotoxic effects against cancer cells in the combinational treatment with GALA peptides and cationic lipids (98% cells were dead by treatment of 2 μM GALA peptides 2% (v/v) Lipofectamine LTX modified saporin-encapsulated exosomes, on the other hand, only 6% cells were dead by the unmodified exosomes (24 h treatment on HeLa cells)) [42], suggesting a broad spectrum of the utility of exosome-based biofunctional protein delivery into the cytosol. Thus, we successfully developed a straightforward methodology for enhancing the cellular uptake of exosomes, followed by the efficient cytosolic release of the encapsulated cargos from the exosomes via the formation of a complex with cationic lipids and the pH-sensitive fusogenic peptide.
5. Other Active Target Systems Based on Exosomes Recently Developed
5.1. Expressing System: LAMP-2B
For delivery of microRNA (miR)-140 into chondrocytes across the dense nonvascular extracellular matrix of cartilage, a chondrocyte affinity peptide (CAP) attached to exosomes expressing the lysosome-associated membrane glycoprotein 2b (Lamp2b) protein has been developed. CAP has been shown to successfully deliver miR-140 to deep cartilage regions through the dense mesochondrium in a rat model [47]. Exosomal expression of interleukin 3 fused to Lamp2b was also attempted for targeting chronic myeloid leukemia [48].
Rabies virus glycoprotein (RVG) was also fused with Lamp2b by genetic engineering, leading to expression of the fused protein on secreted exosomes for delivering high-mobility group box 1 (HMGB1) small interfering RNA (siRNA) against ischemic strokes [49] and alpha-synuclein functionalized short hairpin RNA (shRNA) for specific delivery into the brain in an alpha-synuclein preformed fibril model of parkinsonism [50]. In addition, exosomes genetically engineered to express Lamp2B fused with RVG and glycosylation-stabilized peptides on the exosomal membrane could be coated onto the surfaces of gold nanoparticles using mechanical extrusion methods, leading to efficient brain targeting in mice after administration by intravenous injection [51].
Exosomes from cardiosphere-derived cells (CDCs) have been shown to modulate cardiomyocyte apoptosis and hypertrophy. Cardiomyocyte-specific peptide (CMP, amino acid sequence: WLSEAGPVVTVRALRGTGSW) fused with the extra-exosomal N-terminus of Lamp2b was expressed in CDCs to achieve the secretion of CMP-expressing exosomes [52]. These exosomes resulted in improved specific uptake by cardiomyocytes, decreased cardiomyocyte apoptosis, and enhanced cardiac retention following intramyocardial injection in vivo [52]. Another cardiac-targeting peptide (CTP, amino acid sequence: APWHLSSQYSRT) was also adapted for Lamp2b-based expression on the exosomal membrane for targeting heart tissues in vivo [53].
In addition, the possibility of degradation of the peptides fused to the N-terminus of Lamp2b has been pointed out. In this context, Dr. Hung and Leonard have developed a method to introduce a glycosylation motif that protects the peptide from degradation and leads to an increase in overall expression of the Lamp2b-fused protein in both cells and exosomes [54].
5.2. Expressing System: CD63
Curley et al. reported the engineering of exosomal marker protein CD63 expression as a molecular scaffold to impart cellular targeting properties to the exosomal surface [55]. Sequential deletions of the transmembrane helix of CD63 were carried out to generate a series of truncated CD63 analogs, and the transmembrane helix 3 (TM3) was targeted and anchored to the exosome membrane with n-, N-, Ω-, or I-shaped membrane topologies [55].
In addition, Duong et al. have developed a CD63 surface-display system to produce decoy exosomes bearing genetically transformed TNFα receptors. The authors designed and constructed a chimeric gene composed of three geometrically defined protein domains, namely the transmembrane protein CD63, the decoy TNF receptor 1 on the outer surface (second loop on the outer exosomal surface), and the fluorescent protein on the inner surface of the exosomes (C-terminus inside lumen), in HEK293 cells [56]. The decoy exosomes were found to exhibit a specific anti-TNFα activity on an inflammatory cell reporter system [56].
5.3. Expressing System: VSVG
The exosome-anchoring protein vesicular stomatitis virus glycoprotein (VSVG) could be fused with lysosomal β-glucocerebrosidase (GBA) by genetic engineering, leading to efficient exosomal loading of the fused protein without alterations in exosomal surface markers, particle size, or distribution [57]. The developed system was aimed at treating a large family of lysosomal storage diseases and was validated by the significant increase in GBA biological activity observed upon endocytosis of GBA-exosomes [57].
5.4. Membrane Protein-Ligand System
Exosomes have been reported to contain various membrane proteins such as transferrin receptors (TfR), known to bind its natural ligand transferrin (Tf), which could be employed for binding to multiple functional molecules on exosomal membranes [58,59]. Carboxylated chitosan interacts with the amino group of Tf in the presence of carbodiimide and N-hydroxysulfosuccinimide to yield Tf-modified supermagnetic iron oxide nanoparticles (SPIONs), which could be used for surface modification of exosomes with high TfR expression level [58,59]. The authors successfully achieved TNF-α-based therapeutic delivery using this exosomal system, leading to enhanced therapeutic effects on various types of cancers including breast cancer [59]. In addition, the BAY 55-9837 peptide delivery system was employed to target the pancreas and improve glucose-dependent insulin secretion [58]. Yang et al. used a superparamagnetic system to achieve precise and mild separation of TfR-expressing exosomes in blood, along with a pH-dependent dissociation of Tf and TfR [60].
Streptavidin-lactadherin modified exosomes designed by gene engineering have been developed by Dr. Takakura group, and those functionalized with biotin-tagged CpG DNA on the exosomal surface (CpG-sEV) have also been reported [61]. CpG-sEV could be mixed with cross-linker DNA to induce assembly of the CpG-sEVs [61]. The assembled CpG-sEV was selectively taken up by antigen-presenting cells, leading to an effective tumor antigen-specific immune response in vivo [61].
5.5. DSPE or Other Hydrophobic Moiety Insertion System
Feng et al. developed human hyaluronidase expressing exosomes using genetic engineering by introducing folic acid (FA) modification of 1,2-distearoyl-sn-glycero-3-phosphoethanolamine-N-[folate(polyethylene glycol)] (DSPE-PEG-FA) (collectively termed Exo-PH20-FA). These can degrade high molecular weight hyaluronan to low molecular weight analogs in the tumor stroma, leading to reorientation of pro-tumorigenic M2-like tumor-associated macrophages (TAMs) to anti-tumorigenic M1-like TAMs [62]. Due to its effective use of therapeutic molecules (e.g., anticancer drugs such as doxorubicin) encapsulated in exosomes, the Exo-PH20-FA system could emerge as a strong candidate to attain combined chemotherapy and immune therapy strategies [62].
Similar systems, including FA-modified exosomes with encapsulated erastin (ferroptosis inducer) for targeting triple-negative breast cancer cells, have been reported [63].
The DSPE system is particularly well-suited for PEG functionalization of exosomal surfaces to improve the circulation lifetime in vivo. Eman et al. have shown that PEGylated exosomes (PEG2000) accumulated in murine colorectal cancer cell (C26)-tissue, leading to enhanced exosome uptake by the cancer cells and immune cells [64].
In addition, mannose-conjugated PEG-DSPE (DSPE-PEG-mannose) was prepared using the reaction of amino-PEG-DSPE with α-D-mannopyranosylphenyl isothiocyanate. Mannose-PEG-DSPE-modified exosomes exhibited excellent intracellular uptake into dendritic cells, and enhanced the immune response on account of the adjuvant monophosphoryl lipid A encapsulated within the exosomes [65].
To target CD44-expressing cancer cells, hyaluronic acid (HA) grafted with octadecyl lipids was used to functionalize exosomal membranes [66]. HA-functionalized exosomes with encapsulated doxorubicin (DOX) substantially increased intracellular DOX accumulation in drug-resistant breast cancer cells [66].
Multifunctional chimeric peptide engineered exosomes (ChiP-Exo) were developed by Cheng et al. for plasma membrane and nucleus targeted photodynamic therapy (PTD) [67]. The multifunctional chimeric peptide (ChiP) of C16-K(protoporphyrin IX (PpIX))-PKKKRKV consisted of an alkyl chain for exosomal decoration, a photo sensitizer of PpIX for PTD, and a nuclear localization signal (NLS) peptide for nuclear translocation [67]. Plasma membrane-targeted ChiP-Exo could activate ROS upon light irradiation, and subsequently disrupt plasma membrane structure and cause cell death [67]. Lysosomal escape at the first of stage light irradiation improved cellular delivery to the cytosol, and NLS peptide enhanced the intranuclear PTD effect at the second-stage light in vitro and in vivo both investigations [67].
The cholesterol moiety is also suitable for insertion into the exosomal membrane and is, therefore, used to incorporate functional molecules on exosomal surfaces. For targeting non-small cell lung cancer (NSCLC) cells, the octapeptide cNGQGEQc (CC8), which specifically binds to integrin α3β1, was conjugated to the maleimide-PEG2000-cholesterol (CC8-Mal-PEG-CLT) linker [68]. CC8-Mal-PEG-CLT was added to imperialine-loaded exosomes, leading to the development of CC8-modified exosomes for specific delivery to NSCLC [68].
5.6. DNA and RNA Nanotechnology
Dr. Guo’s group developed an RNA-based nanotechnological breakthrough to obtain FA-decorated exosomes for efficient cell targeting, siRNA delivery, and cancer regression, such as those functionalized with survivin siRNA for use in colorectal cancer xenografts [69,70,71]. Cholesterol-modified and folate-harboring RNA nanoparticles were incubated with siRNA-loaded exosomes for FA-based functionalization [69,70,71]. Recently, these authors demonstrated receptor-mediated cytosolic delivery of siRNA payload without endosomal trapping using these FA-studded exosomes, leading to successful realization of biological activities of siRNAs in targeted cells [71].
The DNA tethering technique for exosomal surface functionalization was also developed recently by Yerneni et al. [72]. In this method, the authors used an 18-mer DNA tether with a 5′-tetra(ethylene glycol) (TEG) spacer and a cholesterol moiety (Chol-DNA) [72]. Simple mixing of Chol-DNA and exosomes in buffered solution for 5 min resulted in DNA modification on the exosomal surface [72]. The DNA strand complementary to the Chol-DNA, used for annealing with Chol-DNA inserted into the exosomal membrane, leads to an exosome with a duplex DNA [72]. This system can be utilized to modify functional proteins on exosomal membranes by binding to complementary DNA strands, such as Fas ligands for apoptosis induction in vitro and in vivo [72].
5.7. Coating with Gel
Amphiphilic cationic nanogels can be complexed with exosomes to improve their intracellular delivery [73]. The developed nanogel has amino group-modified cholesterol-bearing pullulan (CHP), which forms a hybrid material with exosomes [73]. By effective usage of this system, the authors have successfully achieved neuron-like differentiation of human adipocyte-derived MSCs using exosomes from PC12 cells [73]. In addition, magnetic nanogels, consisting of CHP and iron oxide nanoparticles, were hybridized to exosomes isolated from PC12 cells [74]. The magnetic field efficiently transferred the exosome-magnetic hybrid into target adipose-derived stem cells, leading to differentiation of the stem cells into neuron-like cells [74].
5.8. Antibody Modification System
Dual-targeted exosomes with anti-somatostatin receptor 2 (SSTR2) monoclonal antibody (mAb) and anti-C-X-C motif chemokine receptor 4 (CXCR4) mAb against neuroendocrine (NE) cancers in vivo have been developed [75]. Both mAbs were tagged to extracellular vesicles including exosomes via a DSPE-PEG succinimidyl ester (DSPE-PEG-NHS) linker. Encapsulation of the naturally occurring cytotoxic marine compound verrucarin A and the microtubule polymerization inhibitor meytansine (DM1) in these exosomes effectively inhibited NE tumor growth in vivo in comparison to treatments based on single targeting and single drugs [75]. Similar Ab-functionalization techniques using the DSPE-PEG-NHS linker have been reported for exosomal target delivery of romidepsin (FK228, a potent histone deacetylase inhibitor that induces apoptosis through the generation of hydrogen peroxide) using SSTR2 Ab against NE cancers [76].
Azide-modified exosomes derived from M1 macrophages have been conjugated with dibenzocyclooctyne-modified Abs such as CD47 and signal regulatory proteins alpha-linked to pH-sensitive benzoic-imine bonds [77]. The Ab-decorated exosomes accumulated on the CD47-overexpressed tumor surface, leading to the selective cleavage of the benzoic-imine bond in the slightly acidic microenvironment and release of the Abs [77]. The released Abs suppressed the “don’t eat me” signaling and improved the phagocytosis of macrophages [77].
By effective usage of lactadherin C1-C2 domains that bind to exosomal surfaces, high-affinity anti-HER2 single-chain variable fragment (scFv; Kd = 10−9 mol/L) could be attached to the exosomal surface for targeting the cancer HER2 receptor in vitro (e.g., HCC1954 and SK-BR-3 cells [78]) and BT474 HER2+ xenografts implanted in mice [79].
Topping et al. reported a sonication technique for the incorporation of anti-ROS-CII Ab in extracellular vesicles derived from human neutrophils using aqueous energy dissemination, in which sonication resulted in the production of liposomes entrapping the Ab while simultaneously integrating within the vesicle membranes [80]. The vesicles and antibody solution were added to the phospholipid film, and the solution was sonicated for 5 min to attain enrichment of the vesicles with Abs for specifically targeting the arthritic joint in vivo [80].
Zou et al. developed expression of scFv of HIV-1-specific mAb (10E8) on the exosomal surface using the pDisplay gene engineering technique [81]. Curcumin (for killing HIV-1- infected cells) or miR-143 (for apoptosis induction) encapsulated exosomes were used to deliver the cargos to Env+ target cells to induce a strong suppression of the tumor growth in vivo [81].
Nanobodies are single variable domains derived from the heavy chains of Camelidae heavy chain Abs, and nanobody-PEG-micelles were mixed with exosomes to transfer nanobody-PEG-lipids to the exosomal membrane targeting EGFR with cell specificity, prolonged circulation time, and enhanced accumulation in targeted tissues [82]. In addition, GPI-linked nanobodies were developed for attachment to the exosomal surface to improve cell association to EGFR-expressing tumor cells under flow conditions [83].
6. Enhancement of Cellular Exosome Uptake by Cotreatment of Anticancer Drug
In addition to functional peptide-modified exosomes for enhanced cellular uptake, we also determined the effects of anticancer drugs on the internalization of exosomes by cancer cells. In this study, we demonstrated the effects of gefitinib, a tyrosine kinase inhibitor of EGFR, on the cellular uptake of exosomes [84]. In EGFR-mutant HCC827 non-small cell lung cancer (NSCLC) cells, which are sensitive to gefitinib, macropinocytosis was suppressed by gefitinib treatment. However, the cellular uptake of exosomes was increased by gefitinib treatment (10 nM gefitinib treatment increased cellular uptake of exosomes (4-fold) in gefitinib-sensitive HCC827 cells but not in A549 cells that are not sensitive to gefitinib), whereas that of the liposomes was reduced (10 nM gefitinib treatment reduced cellular uptake of liposomes (35%) in gefitinib-sensitive HCC827 cells but not in A549 cells that are not sensitive to gefitinib) [84]. In accordance with the results of the cellular uptake studies, the anti-cancer activity of doxorubicin (DOX)-loaded exosomes in HCC827 cells was significantly increased in the presence of gefitinib (10 nM gefitinib treatment increased cell death (~50%) in gefitinib-sensitive HCC827 cells but not in A549 cells that are not sensitive to gefitinib), whereas that of DOX-loaded liposomes was reduced (10 nM gefitinib treatment reduced cell death (~50%) in gefitinib-sensitive HCC827 cells but not in A549 cells that are not sensitive to gefitinib) [84]. The digestion of exosomal membrane proteins by trypsin surprisingly increased uptake, suggesting a complex cellular uptake mechanism. These results suggest that gefitinib can enhance cell-to-cell communication via exosomes within the tumor microenvironment. It is likely that this unidentified mechanism greatly improves cellular uptake by the additive effect of both drugs; however, further elucidation of the mechanisms is needed to confirm this. Regardless, exosomes show potential as drug delivery vehicles in combination with gefitinib for the treatment of patients harboring EGFR-mutant NSCLC tumors.
7. Conclusions
In this review, the importance of macropinocytosis in the cellular uptake of exosomes and the methods for modification of biofunctional peptides on exosomes to induce macropinocytosis in targeted recipient cells have been discussed. Another active target system based on exosomes recently developed was also summarized and discussed, and macropinocytosis induction might possibly contribute to improvement for the cellular uptake efficacy and biological efficiency in the system. In exosome-based cell-to-cell communication, cellular uptake pathways may significantly affect their communication efficacy, and cellular macropinocytosis induction, which is affected by each cellular innate capacity and environmental condition, might also decide exosome-based control of cellular functions even in disease progression, including cancer (e.g., macropinocytosis inducible cancer-related receptors such as epidermal growth factor receptors and chemokine receptors (EGFR and CXCR4)). Further elucidation of exosome-based communication in homeostasis and disease progression is required to develop sophisticated therapeutic technology utilizing cellular uptake. We have developed straightforward techniques to modify biofunctional peptides on exosomal membranes which could be useful in controlling intercellular communications and the development of therapeutic delivery systems. These findings and experimental techniques for “macropinocytosis in exosomes” controlled by biologically functional peptides will hopefully contribute toward the study of exosome-based communication. And other macropinocytosis inducer will be applicable, however, there are few macropinocytosis inducers reported including arginine-rich peptides, and ligands for macropinocytosis inducible cancer-related receptors such as EGFR and CXCR4, therefore, further development of macropinocytosis inducer, which is useful for enhancing cellular uptake in not only exosome-based intracellular delivery but also other systems such as antibody-based delivery and liposomes, should be needed.
Funding
This study was supported in part by JSPS KAKENHI (JP19H05553) and JST CREST (JPMJCRI7H3). This study was also supported by the Leading University as a Base for Human Resource Development in Nanoscience and Nanotechnology, Osaka Prefecture University.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
Kayo Hirano (Osaka Prefecture University) assisted in the preparation of this manuscript.
Conflicts of Interest
The author declares no conflict of interest.
References
- Tan, A.; Rajadas, J.; Seifalian, A.M. Exosomes as nano-theranostic delivery platforms for gene therapy. Adv. Drug Deliv. Rev. 2013, 65, 357–367. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, T.; Kosaka, N.; Ochiya, T. Latest advances in extracellular vesicles: From bench to bedside. Sci. Technol. Adv. Mater. 2019, 20, 746–757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, L.; Gu, Y.; Du, Y.; Liu, J. Exosomes: Diagnostic Biomarkers and Therapeutic Delivery Vehicles for Cancer. Mol. Pharm. 2019, 16, 3333–3349. [Google Scholar] [CrossRef] [PubMed]
- Tominaga, N.; Kosaka, N.; Ono, M.; Katsuda, T.; Yoshioka, Y.; Tamura, K.; Lötvall, J.; Nakagama, H.; Ochiya, T. Brain metastatic cancer cells release microRNA-181c-containing extracellular vesicles capable of destructing blood-brain barrier. Nat. Commun. 2015, 6, 6716. [Google Scholar] [CrossRef] [Green Version]
- Yokoi, A.; Yoshioka, Y.; Yamamoto, Y.; Ishikawa, M.; Ikeda, S.I.; Kato, T.; Kiyono, T.; Takeshita, F.; Kajiyama, H.; Kikkawa, F.; et al. Malignant extracellular vesicles carrying MMP1 mRNA facilitate peritoneal dissemination in ovarian cancer. Nat. Commun. 2017, 8, 14470. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Li, G.; Liu, M.L. Microvesicles as Emerging Biomarkers and Therapeutic Targets in Cardiometabolic Diseases. Genom. Proteom. Bioinform. 2018, 16, 50–62. [Google Scholar] [CrossRef]
- Raposo, G.; Stoorvogel, W. Extracellular vesicles: Exosomes, microvesicles, and friends. J. Cell Biol. 2013, 200, 373–383. [Google Scholar] [CrossRef] [Green Version]
- Lázaro-Ibáñez, E.; Sanz-Garcia, A.; Visakorpi, T.; Escobedo-Lucea, C.; Siljander, P.; Ayuso-Sacido, A.; Yliperttula, M. Different gDNA content in the subpopulations of prostate cancer extracellular vesicles: Apoptotic bodies, microvesicles, and exosomes. Prostate 2014, 74, 1379–1390. [Google Scholar] [CrossRef]
- Xu, X.; Lai, Y.; Hua, Z.C. Apoptosis and apoptotic body: Disease message and therapeutic target potentials. Biosci. Rep. 2019, 39, BSR20180992. [Google Scholar] [CrossRef] [Green Version]
- Cocucci, E.; Racchetti, G.; Meldolesi, J. Shedding microvesicles: Artefacts no more. Trends Cell Biol. 2009, 19, 43–51. [Google Scholar] [CrossRef]
- Kajimoto, T.; Okada, T.; Miya, S.; Zhang, L.; Nakamura, S. Ongoing activation of sphingosine 1-phosphate receptors mediates maturation of exosomal multivesicular endosomes. Nat. Commun. 2013, 4, 2712. [Google Scholar] [CrossRef] [Green Version]
- Mohamed, N.N.I.; Okada, T.; Kajimoto, T.; Nakamura, S.I. Essential Role of Sphingosine Kinase 2 in the Regulation of Cargo Contents in the Exosomes from K562 Cells. Kobe J. Med. Sci. 2018, 63, E123–E129. [Google Scholar] [PubMed]
- Kajimoto, T.; Mohamed, N.N.I.; Badawy, S.M.M.; Matovelo, S.A.; Hirase, M.; Nakamura, S.; Yoshida, D.; Okada, T.; Ijuin, T.; Nakamura, S.I. Involvement of Gβγ subunits of Gi protein coupled with S1P receptor on multivesicular endosomes in F-actin formation and cargo sorting into exosomes. J. Biol. Chem. 2018, 293, 245–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, S.; Bai, O.; Li, F.; Yuan, J.; Laferte, S.; Xiang, J. Mature dendritic cells pulsed with exosomes stimulate efficient cytotoxic T-lymphocyte responses and antitumour immunity. Immunology 2007, 120, 90–102. [Google Scholar] [CrossRef]
- Saunderson, S.C.; Dunn, A.C.; Crocker, P.R.; McLellan, A.D. CD169 mediates the capture of exosomes in spleen and lymph node. Blood 2014, 123, 208–216. [Google Scholar] [CrossRef] [PubMed]
- Shimoda, A.; Tahara, Y.; Sawada, S.I.; Sasaki, Y.; Akiyoshi, K. Glycan profiling analysis using evanescent-field fluorescence-assisted lectin array: Importance of sugar recognition for cellular uptake of exosomes from mesenchymal stem cells. Biochem. Biophys. Res. Commun. 2017, 491, 701–707. [Google Scholar] [CrossRef]
- Zech, D.; Rana, S.; Büchler, M.W.; Zöller, M. Tumor-exosomes and leukocyte activation: An ambivalent crosstalk. Cell Commun. Signal. 2012, 10, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoshino, A.; Costa-Silva, B.; Shen, T.L.; Rodrigues, G.; Hashimoto, A.; Mark, M.T.; Molina, H.; Kohsaka, S.; Di Giannatale, A.; Ceder, S.; et al. Tumour exosome integrins determine organotropic metastasis. Nature 2015, 527, 329–335. [Google Scholar] [CrossRef] [Green Version]
- Yuan, D.; Zhao, Y.; Banks, W.A.; Bullock, K.M.; Haney, M.; Batrakova, E.; Kabanov, A.V. Macrophage exosomes as natural nanocarriers for protein delivery to inflamed brain. Biomaterials 2017, 142, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Christianson, H.C.; Svensson, K.J.; Van Kuppevelt, T.H.; Li, J.P.; Belting, M. Cancer cell exosomes depend on cell-surface heparan sulfate proteoglycans for their internalization and functional activity. Proc. Natl. Acad. Sci. USA 2013, 110, 17380–17385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Purushothaman, A.; Bandari, S.K.; Liu, J.; Mobley, J.A.; Brown, E.E.; Sanderson, R.D. Fibronectin on the Surface of Myeloma Cell-derived Exosomes Mediates Exosome-Cell Interactions. J. Biol. Chem. 2016, 291, 1652–1663. [Google Scholar] [CrossRef] [Green Version]
- Baglio, S.R.; Van Eijndhoven, M.A.; Koppers-Lalic, D.; Berenguer, J.; Lougheed, S.M.; Gibbs, S.; Léveillé, N.; Rinkel, R.N.; Hopmans, E.S.; Swaminathan, S.; et al. Sensing of latent EBV infection through exosomal transfer of 5′pppRNA. Proc. Natl. Acad. Sci. USA 2016, 113, E587–E596. [Google Scholar] [CrossRef] [Green Version]
- Gonda, A.; Kabagwira, J.; Senthil, G.N.; Wall, N.R. Internalization of Exosomes through Receptor-Mediated Endocytosis. Mol. Cancer Res. 2019, 17, 337–347. [Google Scholar] [CrossRef] [Green Version]
- Conner, S.D.; Schmid, S.L. Regulated portals of entry into the cell. Nature 2003, 422, 37–44. [Google Scholar] [CrossRef]
- Swanson, J.A.; Watts, C. Macropinocytosis. Trends Cell Biol. 1995, 5, 424–428. [Google Scholar] [CrossRef]
- Swanson, J.A. Shaping cups into phagosomes and macropinosomes. Nat. Rev. Mol. Cell Biol. 2008, 9, 639–649. [Google Scholar] [CrossRef] [Green Version]
- Nakase, I.; Kobayashi, N.B.; Takatani-Nakase, T.; Yoshida, T. Active macropinocytosis induction by stimulation of epidermal growth factor receptor and oncogenic Ras expression potentiates cellular uptake efficacy of exosomes. Sci. Rep. 2015, 5, 10300. [Google Scholar] [CrossRef] [Green Version]
- Commisso, C.; Davidson, S.M.; Soydaner-Azeloglu, R.G.; Parker, S.J.; Kamphorst, J.J.; Hackett, S.; Grabocka, E.; Nofal, M.; Drebin, J.A.; Thompson, C.B.; et al. Macropinocytosis of protein is an amino acid supply route in Ras-transformed cells. Nature 2013, 497, 633–637. [Google Scholar] [CrossRef] [Green Version]
- Fabbrini, M.S.; Katayama, M.; Nakase, I.; Vago, R. Plant Ribosome-Inactivating Proteins: Progesses, Challenges and Biotechnological Applications (and a Few Digressions). Toxins 2017, 9, 314. [Google Scholar] [CrossRef] [Green Version]
- Nakase, I.; Noguchi, K.; Fujii, I.; Futaki, S. Vectorization of biomacromolecules into cells using extracellular vesicles with enhanced internalization induced by macropinocytosis. Sci. Rep. 2016, 6, 34937. [Google Scholar] [CrossRef] [Green Version]
- Futaki, S.; Nakase, I. Cell-Surface Interactions on Arginine-Rich Cell-Penetrating Peptides Allow for Multiplex Modes of Internalization. Acc. Chem. Res. 2017, 50, 2449–2456. [Google Scholar] [CrossRef]
- Nakase, I.; Niwa, M.; Takeuchi, T.; Sonomura, K.; Kawabata, N.; Koike, Y.; Takehashi, M.; Tanaka, S.; Ueda, K.; Simpson, J.C.; et al. Cellular uptake of arginine-rich peptides: Roles for macropinocytosis and actin rearrangement. Mol. Ther. 2004, 10, 1011–1022. [Google Scholar] [CrossRef]
- Nakase, I.; Tadokoro, A.; Kawabata, N.; Takeuchi, T.; Katoh, H.; Hiramoto, K.; Negishi, M.; Nomizu, M.; Sugiura, Y.; Futaki, S. Interaction of arginine-rich peptides with membrane-associated proteoglycans is crucial for induction of actin organization and macropinocytosis. Biochemistry 2007, 46, 492–501. [Google Scholar] [CrossRef]
- Nakase, I.; Hirose, H.; Tanaka, G.; Tadokoro, A.; Kobayashi, S.; Takeuchi, T.; Futaki, S. Cell-surface accumulation of flock house virus-derived peptide leads to efficient internalization via macropinocytosis. Mol. Ther. 2009, 17, 1868–1876. [Google Scholar] [CrossRef]
- Nakase, I.; Osaki, K.; Tanaka, G.; Utani, A.; Futaki, S. Molecular interplays involved in the cellular uptake of octaarginine on cell surfaces and the importance of syndecan-4 cytoplasmic V domain for the activation of protein kinase Cα. Biochem. Biophys. Res. Commun. 2014, 446, 857–862. [Google Scholar] [CrossRef]
- Kosuge, M.; Takeuchi, T.; Nakase, I.; Jones, A.T.; Futaki, S. Cellular internalization and distribution of arginine-rich peptides as a function of extracellular peptide concentration, serum, and plasma membrane associated proteoglycans. Bioconjug. Chem. 2008, 19, 656–664. [Google Scholar] [CrossRef]
- Nakase, I.; Noguchi, K.; Aoki, A.; Takatani-Nakase, T.; Fujii, I.; Futaki, S. Arginine-rich cell-penetrating peptide-modified extracellular vesicles for active macropinocytosis induction and efficient intracellular delivery. Sci. Rep. 2017, 7, 1991. [Google Scholar] [CrossRef] [PubMed]
- Nakase, I.; Okumura, S.; Tanaka, G.; Osaki, K.; Imanishi, M.; Futaki, S. Signal transduction using an artificial receptor system that undergoes dimerization upon addition of a bivalent leucine-zipper ligand. Angew. Chem. Int. Ed. Engl. 2012, 51, 7464–7467. [Google Scholar] [CrossRef]
- Nakase, I.; Ueno, N.; Katayama, M.; Noguchi, K.; Takatani-Nakase, T.; Kobayashi, N.B.; Yoshida, T.; Fujii, I.; Futaki, S. Receptor clustering and activation by multivalent interaction through recognition peptides presented on exosomes. Chem. Commun. 2017, 53, 317–320. [Google Scholar] [CrossRef] [PubMed]
- Litowski, J.R.; Hodges, R.S. Designing heterodimeric two-stranded alpha-helical coiled-coils. Effects of hydrophobicity and alpha-helical propensity on protein folding, stability, and specificity. J. Biol. Chem. 2002, 277, 37272–37279. [Google Scholar] [CrossRef] [Green Version]
- Lindhout, D.A.; Litowski, J.R.; Mercier, P.; Hodges, R.S.; Sykes, B.D. NMR solution structure of a highly stable de novo heterodimeric coiled-coil. Biopolymers 2004, 75, 367–375. [Google Scholar] [CrossRef]
- Nakase, I.; Futaki, S. Combined treatment with a pH-sensitive fusogenic peptide and cationic lipids achieves enhanced cytosolic delivery of exosomes. Sci. Rep. 2015, 5, 10112. [Google Scholar] [CrossRef]
- Subbarao, N.K.; Parente, R.A.; Szoka, F.C., Jr.; Nadasdi, L.; Pongracz, K. pH-dependent bilayer destabilization by an amphipathic peptide. Biochemistry 1987, 26, 2964–2972. [Google Scholar] [CrossRef]
- Li, W.; Nicol, F.; Szoka, F.C., Jr. GALA: A designed synthetic pH-responsive amphipathic peptide with applications in drug and gene delivery. Adv. Drug Deliv. Rev. 2004, 56, 967–985. [Google Scholar] [CrossRef]
- Nakase, I.; Kogure, K.; Harashima, H.; Futaki, S. Application of a fusiogenic peptide GALA for intracellular delivery. Methods Mol. Biol. 2011, 683, 525–533. [Google Scholar]
- Kobayashi, S.; Nakase, I.; Kawabata, N.; Yu, H.H.; Pujals, S.; Imanishi, M.; Giralt, E.; Futaki, S. Cytosolic targeting of macromolecules using a pH-dependent fusogenic peptide in combination with cationic liposomes. Bioconjug. Chem. 2009, 20, 953–959. [Google Scholar] [CrossRef]
- Liang, Y.; Xu, X.; Li, X.; Xiong, J.; Li, B.; Duan, L.; Wang, D.; Xia, J. Chondrocyte-Targeted MicroRNA Delivery by Engineered Exosomes toward a Cell-Free Osteoarthritis Therapy. Acs Appl. Mater. Interfaces 2020, 12, 36938–36947. [Google Scholar] [CrossRef]
- Bellavia, D.; Raimondo, S.; Calabrese, G.; Forte, S.; Cristaldi, M.; Patinella, A.; Memeo, L.; Manno, M.; Raccosta, S.; Diana, P.; et al. Interleukin 3-receptor targeted exosomes inhibit in vitro and in vivo Chronic Myelogenous Leukemia cell growth. Theranostics 2017, 7, 1333–1345. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Kim, G.; Hwang, D.W.; Lee, M. Delivery of high mobility group box-1 sirna using brain-targeting exosomes for ischemic stroke therapy. J. Biomed. Nanotechnol. 2019, 15, 2401–2412. [Google Scholar] [CrossRef]
- Izco, M.; Blesa, J.; Schleef, M.; Schmeer, M.; Porcari, R.; Al-Shawi, R.; Ellmerich, S.; De Toro, M.; Gardiner, C.; Seow, Y.; et al. Systemic Exosomal delivery of shRNA minicircles prevents parkinsonian pathology. Mol. Ther. 2019, 27, 2111–2122. [Google Scholar] [CrossRef]
- Khongkow, M.; Yata, T.; Boonrungsiman, S.; Ruktanonchai, U.R.; Graham, D.; Namdee, K. Surface modification of gold nanoparticles with neuron-targeted exosome for enhanced blood-brain barrier penetration. Sci. Rep. 2019, 9, 8278. [Google Scholar] [CrossRef] [PubMed]
- Mentkowski, K.I.; Lang, J.K. Exosomes engineered to express a cardiomyocyte binding peptide demonstrate improved cardiac retention in vivo. Sci. Rep. 2019, 9, 10041. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Yun, N.; Mun, D.; Kang, J.Y.; Lee, S.H.; Park, H.; Park, H.; Joung, B. Cardiac-specific delivery by cardiac tissue-targeting peptide-expressing exosomes. Biochem. Biophys. Res. Commun. 2018, 499, 803–808. [Google Scholar] [CrossRef]
- Hung, M.E.; Leonard, J.N. Stabilization of exosome-targeting peptides via engineered glycosylation. J. Biol. Chem. 2015, 290, 8166–8172. [Google Scholar] [CrossRef] [Green Version]
- Curley, N.; Levy, D.; Do, M.A.; Brown, A.; Stickney, Z.; Marriott, G.; Lu, B. Sequential deletion of CD63 identifies topologically distinct scaffolds for surface engineering of exosomes in living human cells. Nanoscale 2020, 12, 12014–12026. [Google Scholar] [CrossRef] [PubMed]
- Duong, N.; Curley, K.; Brown, A.; Campanelli, A.; Do, M.A.; Levy, D.; Tantry, A.; Marriottm, G.; Lu, B. Decoy exosomes as a novel biologic reagent to antagonize inflammation. Int. J. Nanomed. 2019, 14, 3413–3425. [Google Scholar] [CrossRef] [Green Version]
- Do, M.A.; Levy, D.; Brown, A.; Marriott, G.; Lu, B. Targeted delivery of lysosomal enzymes to the endocytic compartment in human cells using engineered extracellular vesicles. Sci. Rep. 2019, 9, 17274. [Google Scholar] [CrossRef] [Green Version]
- Zhuang, M.; Du, D.; Pu, L.; Song, H.; Deng, M.; Long, Q.; Yin, X.; Wang, Y.; Rao, L. SPION-decorated exosome delivered BAY55-9837 targeting the pancreas through magnetism to improve the blood GLC reesponse. Small 2019, 15, e1903135. [Google Scholar] [CrossRef]
- Zhuang, M.; Chen, X.; Du, D.; Shi, J.; Deng, M.; Long, Q.; Yin, X.; Wang, Y.; Rao, L. SPION decorated exosome delivery of TNF-α to cancer cell membranes through magnetism. Nanoscale 2020, 12, 173–188. [Google Scholar] [CrossRef]
- Yang, L.; Han, D.; Zhan, Q.; Li, X.; Shan, P.; Hu, Y.; Ding, H.; Wang, Y.; Zhang, L.; Zhang, Y.; et al. Blood TfR+ exosomes separated by a pH-responsive method deliver chemotherapeutics for tumor therapy. Theranostics 2019, 9, 7680–7696. [Google Scholar] [CrossRef]
- Matsumoto, A.; Takahashi, Y.; Ariizumi, R.; Nishikawa, M.; Takakura, Y. Development of DNA-anchored assembly of small extracellular vesicle for efficient antigen delivery to antigen presenting cells. Biomaterials 2019, 225, 119518. [Google Scholar] [CrossRef] [PubMed]
- Feng, C.; Xiong, Z.; Wang, C.; Xiao, W.; Xiao, H.; Xie, K.; Chen, K.; Liang, H.; Zhang, X.; Yang, H. Folic acid-modified Exosome-PH20 enhances the efficiency of therapy via modulation of the tumor microenvironment and directly inhibits tumor cell metastasis. Bioact. Mater. 2020, 6, 963–974. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Gai, C.; Li, Z.; Ding, D.; Zheng, J.; Zhang, W.; Lv, S.; Li, W. Targeted exosome-encapsulated erastin induced ferroptosis in triple negative breast cancer cells. Cancer Sci. 2019, 110, 3173–3182. [Google Scholar] [CrossRef] [Green Version]
- Emam, S.E.; Abu Lila, A.S.; Elsadek, N.E.; Ando, H.; Shimizu, T.; Okuhira, K.; Ishima, Y.; Mahdy, M.A.; Ghazy, F.S.; Ishida, T. Cancer cell-type tropism is one of crucial determinants for the efficient systemic delivery of cancer cell-derived exosomes to tumor tissues. Eur. J. Pharm. Biopharm. 2019, 145, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Choi, E.S.; Song, J.; Kang, Y.Y.; Mok, H. Mannose-modified serum exosomes for the elevated uptake to murine dendritic cells and lymphatic accumulation. Macromol. Biosci. 2019, 19, e1900042. [Google Scholar] [CrossRef]
- Liu, J.; Ye, Z.; Xiang, M.; Chang, B.; Cui, J.; Ji, T.; Zhao, L.; Li, Q.; Deng, Y.; Xu, L.; et al. Functional extracellular vesicles engineered with lipid-grafted hyaluronic acid effectively reverse cancer drug resistance. Biomaterials 2019, 223, 119475. [Google Scholar] [CrossRef]
- Cheng, H.; Fan, J.H.; Zhao, L.P.; Fan, G.L.; Zheng, R.R.; Qiu, X.Z.; Yu, X.Y.; Li, S.Y.; Zhang, X.Z. Chimeric peptide engineered exosomes for dual-stage light guided plasma membrane and nucleus targeted photodynamic therapy. Biomaterials 2019, 211, 14–24. [Google Scholar] [CrossRef]
- Lin, Q.; Qu, M.; Zhou, B.; Patra, H.K.; Sun, Z.; Luo, Q.; Yang, W.; Wu, Y.; Zhang, Y.; Li, L.; et al. Exosome-like nanoplatform modified with targeting ligand improves anti-cancer and anti-inflammation effects of imperialine. J. Control. Release 2019, 311–312, 104–116. [Google Scholar] [CrossRef]
- Pi, F.; Binzel, D.W.; Lee, T.J.; Li, Z.; Sun, M.; Rychahou, P.; Li, H.; Haque, F.; Wang, S.; Croce, C.M.; et al. Nanoparticle orientation to control RNA loading and ligand display on extracellular vesicles for cancer regression. Nat. Nanotechnol. 2018, 13, 82–89. [Google Scholar] [CrossRef]
- Li, Z.; Wang, H.; Yin, H.; Bennett, C.; Zhang, H.G.; Guo, P. Arrowtail RNA for ligand display on ginger exosome-like nanovesicles to systemic deliver siRNA for cancer suppression. Sci. Rep. 2018, 8, 14644. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Z.; Li, Z.; Xu, C.; Guo, B.; Guo, P. Folate-displaying exosome mediated cytosolic delivery of siRNA avoiding endosome trapping. J. Control. Release 2019, 311–312, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Yerneni, S.S.; Lathwal, S.; Shrestha, P.; Shirwan, H.; Matyjaszewski, K.; Weiss, L.; Yolcu, E.S.; Campbell, P.G.; Das, S.R. Rapid on-demand extracellular vesicle augmentation with versatile oligonucleotide tethers. ACS Nano 2019, 13, 10555–10565. [Google Scholar] [CrossRef] [PubMed]
- Sawada, S.I.; Sato, Y.T.; Kawasaki, R.; Yasuoka, J.I.; Mizuta, R.; Sasaki, Y.; Akiyoshi, K. Nanogel hybrid assembly for exosome intracellular delivery: Effects on endocytosis and fusion by exosome surface polymer engineering. Biomater. Sci. 2020, 8, 619–630. [Google Scholar] [CrossRef] [PubMed]
- Mizuta, R.; Sasaki, Y.; Kawasaki, R.; Katagiri, K.; Sawada, S.I.; Mukai, S.A.; Akiyoshi, K. Magnetically navigated intracellular delivery of extracellular vesicles using amphiphilic nanogels. Bioconjug. Chem. 2019, 30, 2150–2155. [Google Scholar] [CrossRef] [PubMed]
- Si, Y.; Guan, J.; Xu, Y.; Chen, K.; Kim, S.; Zhou, L.; Jaskula-Sztul, R.; Liu, X.M. Dual-targeted extracellular vesicles to facilitate combined therapies for neuroendocrine cancer treatment. Pharmaceutics 2020, 12, 1079. [Google Scholar] [CrossRef]
- Si, Y.; Kim, S.; Zhang, E.; Tang, Y.; Jaskula-Sztul, R.; Markert, J.M.; Chen, H.; Zhou, L.; Liu, X.M. Targeted exosomes for drug delivery: Biomanufacturing, surface tagging, and validation. Biotechnol. J. 2020, 15, e1900163. [Google Scholar] [CrossRef]
- Nie, W.; Wu, G.; Zhang, J.; Huang, L.L.; Ding, J.; Jiang, A.; Zhang, Y.; Liu, Y.; Li, J.; Pu, K.; et al. Responsive Exosome nano-bioconjugates for synergistic cancer therapy. Angew. Chem. Int. Ed. Engl. 2020, 59, 2018–2022. [Google Scholar] [CrossRef]
- Longatti, A.; Schindler, C.; Collinson, A.; Jenkinson, L.; Matthews, C.; Fitzpatrick, L.; Blundy, M.; Minter, R.; Vaughan, T.; Shaw, M.; et al. High affinity single-chain variable fragments are specific and versatile targeting motifs for extracellular vesicles. Nanoscale 2018, 10, 14230–14244. [Google Scholar] [CrossRef] [Green Version]
- Forterre, A.V.; Wang, J.H.; Delcayre, A.; Kim, K.; Green, C.; Pegram, M.D.; Jeffrey, S.S.; Matin, A.C. Extracellular vesicle-mediated in vitro transcribed mRNA delivery for treatment of HER2+ breast cancer xenografts in mice by prodrug CB1954 without general toxicity. Mol. Cancer Ther. 2020, 19, 858–867. [Google Scholar] [CrossRef] [Green Version]
- Topping, L.M.; Thomas, B.L.; Rhys, H.I.; Tremoleda, J.L.; Foster, M.; Seed, M.; Voisin, M.B.; Vinci, C.; Law, H.L.; Perretti, M.; et al. Targeting extracellular vesicles to the arthritic joint using a damaged cartilage-specific antibody. Front. Immunol. 2020, 11, 10. [Google Scholar] [CrossRef]
- Zou, X.; Yuan, M.; Zhang, T.; Wei, H.; Xu, S.; Jiang, N.; Zheng, N.; Wu, Z. Extracellular vesicles expressing a single-chain variable fragment of an HIV-1 specific antibody selectively target Env+ tissues. Theranostics 2019, 9, 5657–5671. [Google Scholar] [CrossRef] [PubMed]
- Kooijmans, S.A.A.; Fliervoet, L.A.L.; Van der Meel, R.; Fens, M.H.A.M.; Heijnen, H.F.G.; Van Bergen En Henegouwen, P.M.P.; Vader, P.; Schiffelers, R.M. PEGylated and targeted extracellular vesicles display enhanced cell specificity and circulation time. J. Control. Release 2016, 224, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Kooijmans, S.A.; Aleza, C.G.; Roffler, S.R.; Van Solinge, W.W.; Vader, P.; Schiffelers, R.M. Display of GPI-anchored anti-EGFR nanobodies on extracellular vesicles promotes tumour cell targeting. J. Extracell. Vesicles 2016, 5, 31053. [Google Scholar] [CrossRef] [PubMed]
- Takenaka, T.; Nakai, S.; Katayama, M.; Hirano, M.; Ueno, N.; Noguchi, K.; Takatani-Nakase, T.; Fujii, I.; Kobayashi, S.S.; Nakase, I. Effects of gefitinib treatment on cellular uptake of extracellular vesicles in EGFR-mutant non-small cell lung cancer cells. Int. J. Pharm. 2019, 572, 118762. [Google Scholar] [CrossRef] [PubMed]
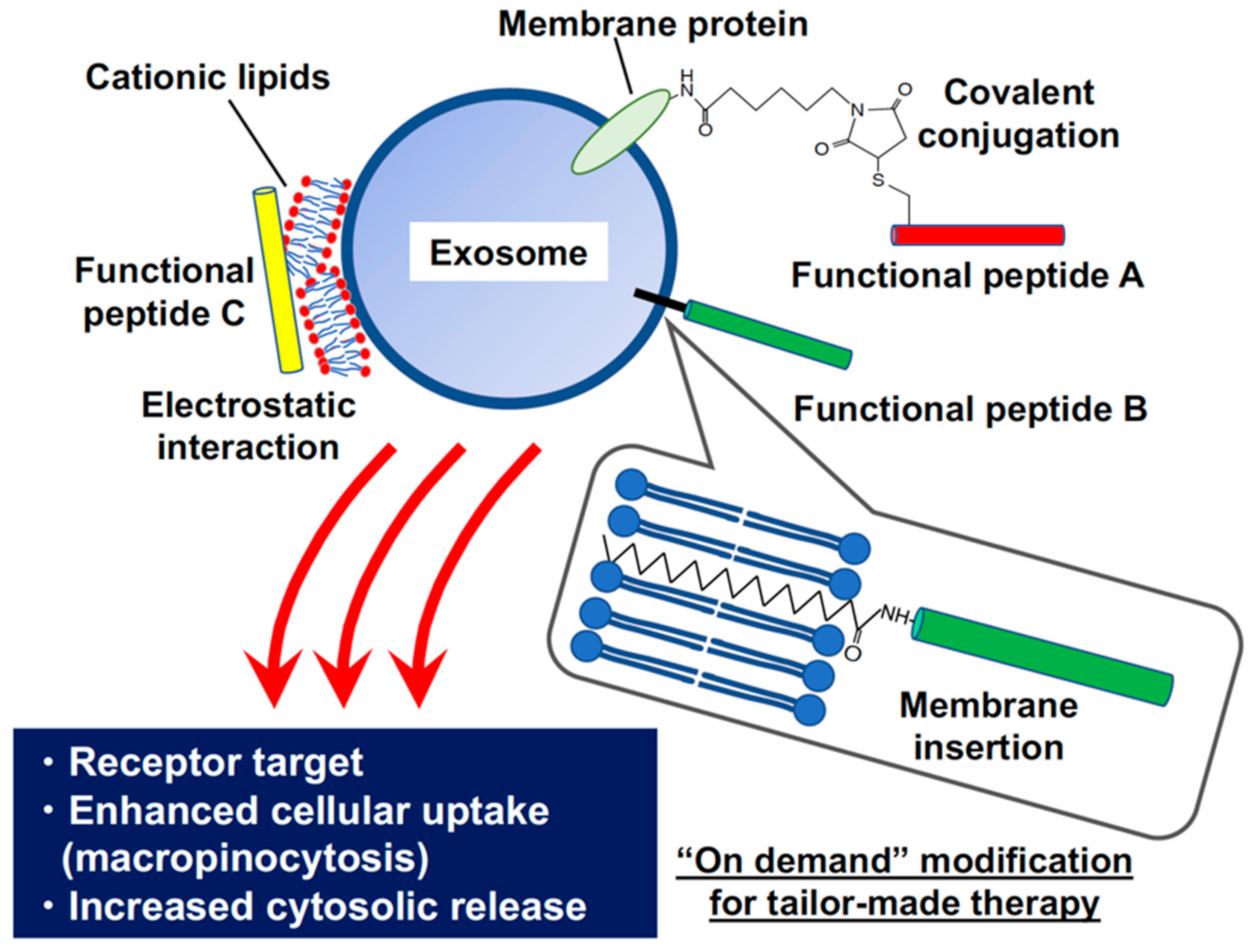
Figure 1.
Modification of biofunctional peptides on exosomal membranes to target recipient cellular membrane proteins and enhance the cytosolic release of exosome contents.
Figure 1.
Modification of biofunctional peptides on exosomal membranes to target recipient cellular membrane proteins and enhance the cytosolic release of exosome contents.

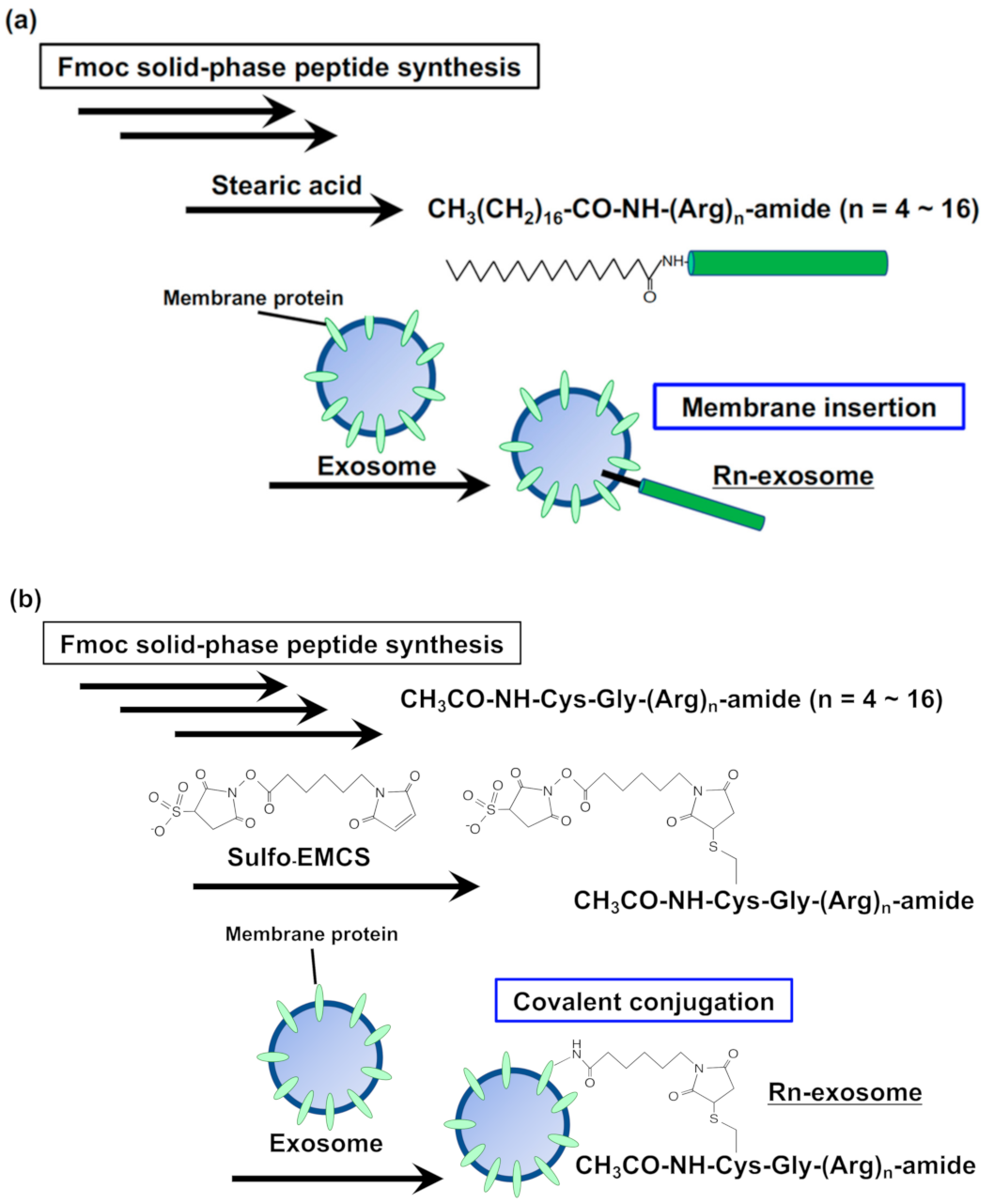
Figure 2.
(a,b) Arginine-rich cell-penetrating peptide-modified (membrane insertion (a) and covalently conjugation (b) methods) exosomes for macropinocytosis induction and efficient recipient cellular uptake.
Figure 2.
(a,b) Arginine-rich cell-penetrating peptide-modified (membrane insertion (a) and covalently conjugation (b) methods) exosomes for macropinocytosis induction and efficient recipient cellular uptake.

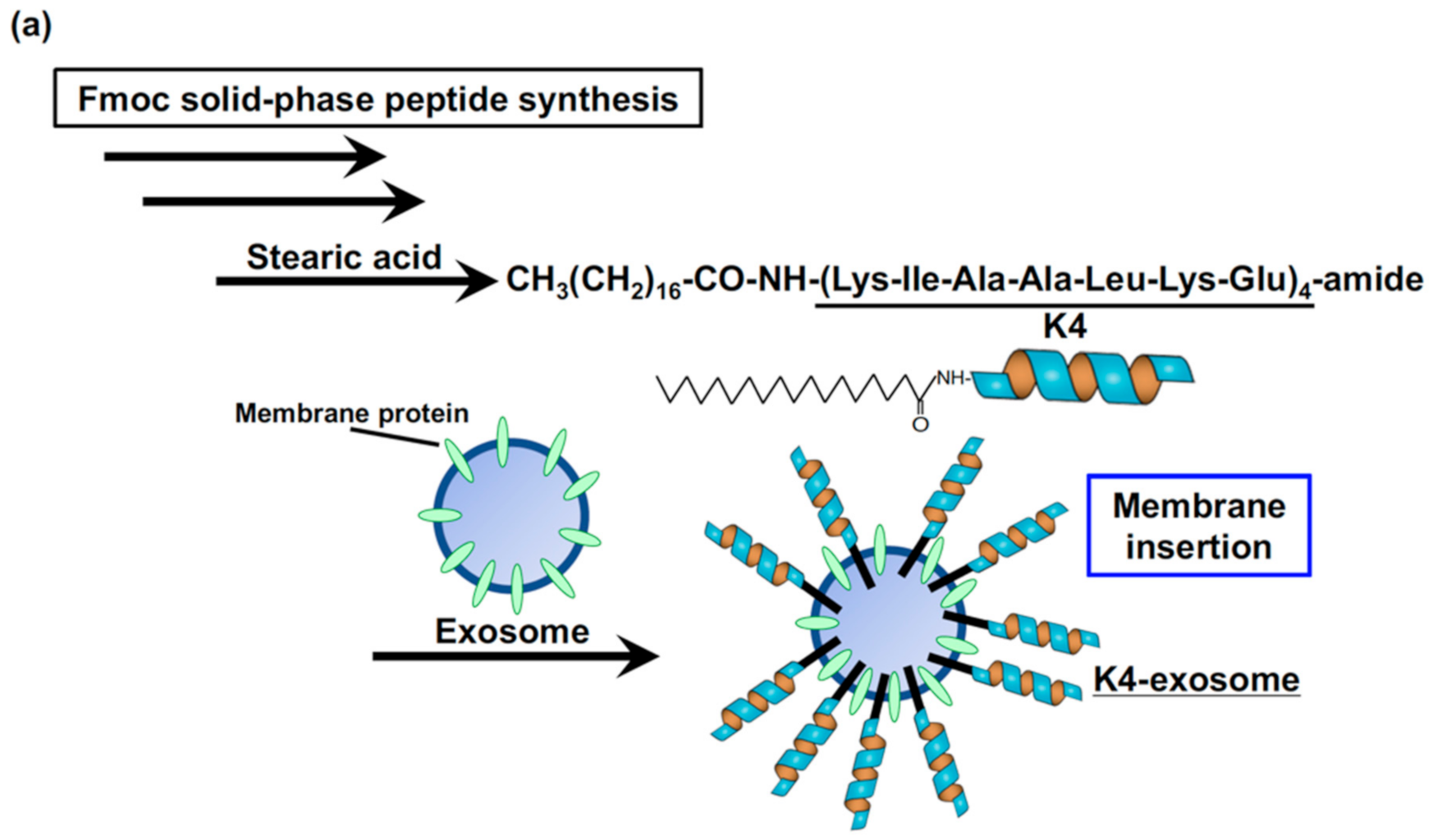
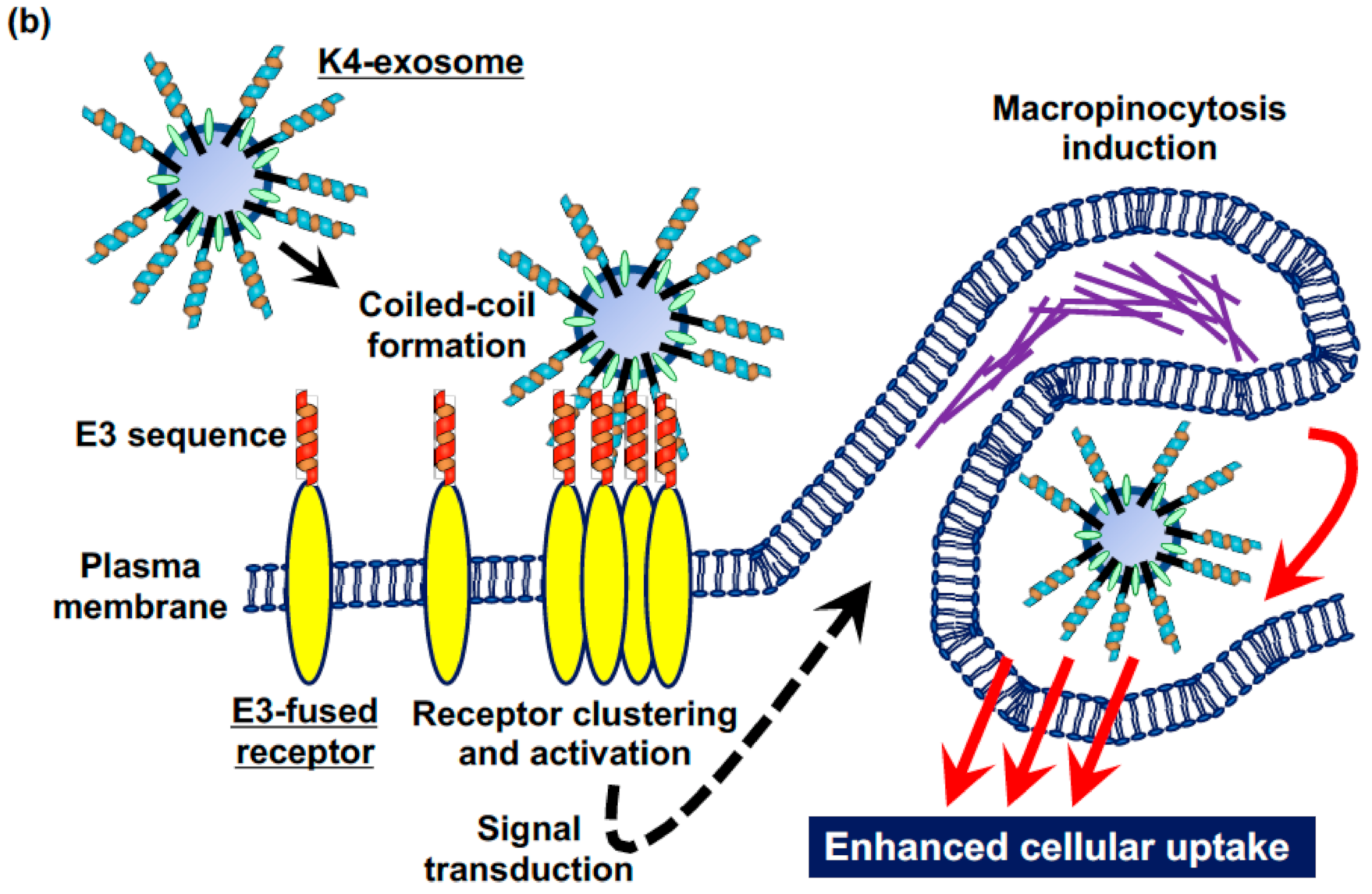
Figure 3.
(a,b) Coiled-coil peptide-modified exosomes for recognition of tagged receptors, clustering, receptor activation, and cellular uptake via macropinocytosis. (a) Synthesis of stearyl-K4 peptide. (b) K4-exosomes recognize E3-receptors, leading to the receptor clustering, activation, signal transduction, macropinocytosis induction, and enhanced cellular uptake of the exosomes.
Figure 3.
(a,b) Coiled-coil peptide-modified exosomes for recognition of tagged receptors, clustering, receptor activation, and cellular uptake via macropinocytosis. (a) Synthesis of stearyl-K4 peptide. (b) K4-exosomes recognize E3-receptors, leading to the receptor clustering, activation, signal transduction, macropinocytosis induction, and enhanced cellular uptake of the exosomes.


Figure 4.
Enhanced cellular uptake and cytosolic release of exosomal contents by combinational treatment of pH-sensitive fusogenic peptide, GALA, and cationic lipids.
Figure 4.
Enhanced cellular uptake and cytosolic release of exosomal contents by combinational treatment of pH-sensitive fusogenic peptide, GALA, and cationic lipids.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Nakase, I. Biofunctional Peptide-Modified Extracellular Vesicles Enable Effective Intracellular Delivery via the Induction of Macropinocytosis. Processes 2021, 9, 224. https://doi.org/10.3390/pr9020224
AMA Style
Nakase I. Biofunctional Peptide-Modified Extracellular Vesicles Enable Effective Intracellular Delivery via the Induction of Macropinocytosis. Processes. 2021; 9(2):224. https://doi.org/10.3390/pr9020224
Chicago/Turabian StyleNakase, Ikuhiko. 2021. "Biofunctional Peptide-Modified Extracellular Vesicles Enable Effective Intracellular Delivery via the Induction of Macropinocytosis" Processes 9, no. 2: 224. https://doi.org/10.3390/pr9020224
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.