Genetic Diversities and Historical Dynamics of Native Ethiopian Horse Populations (Equus caballus) Inferred from Mitochondrial DNA Polymorphisms


Abstract
:1. Introduction
2. Materials and Methods
Data Analysis
3. Results
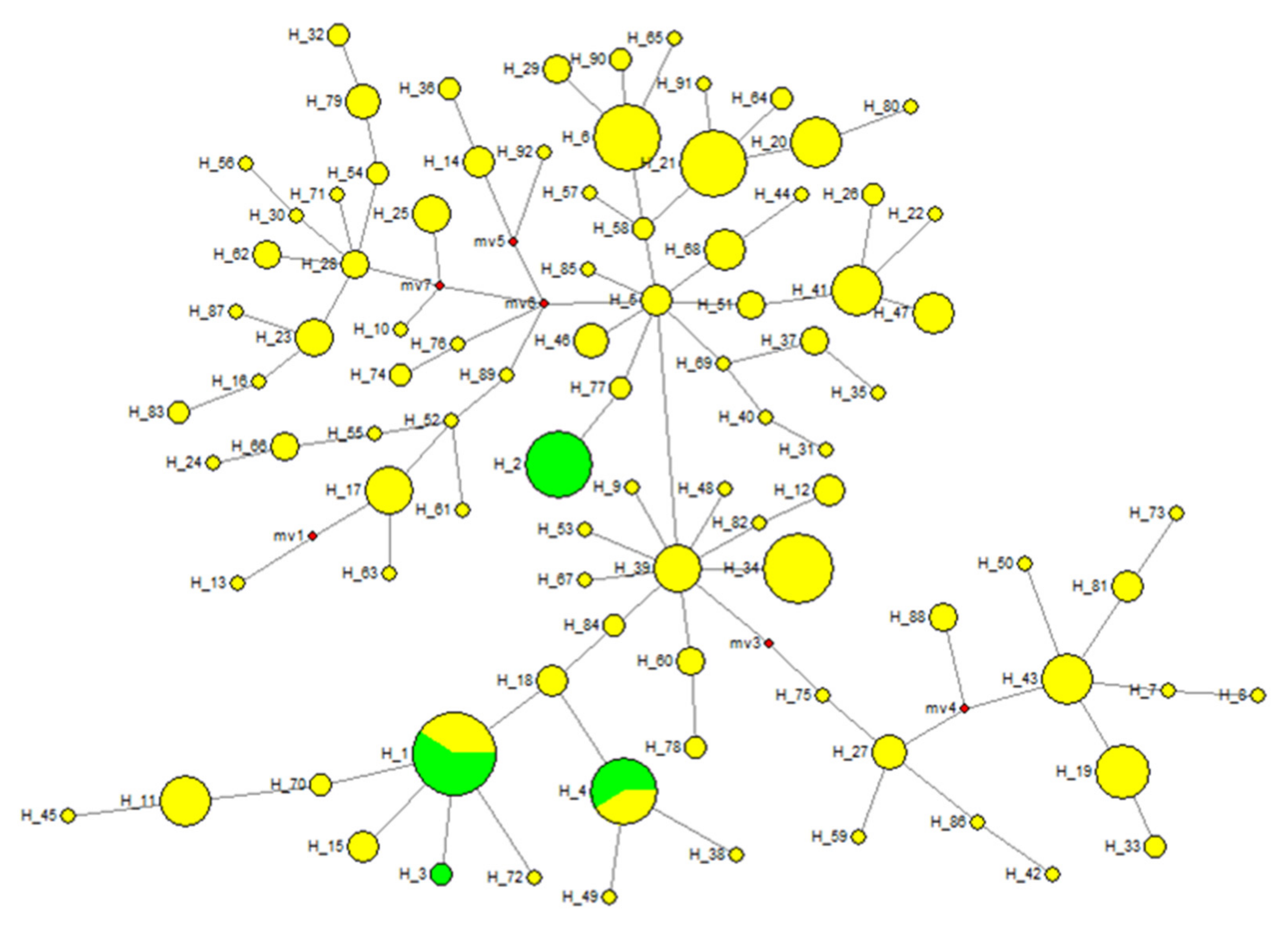
3.1. Haplotype Diversities in Native Ethiopian Horse Populations
3.2. Nucleotide Diversities in Native Ethiopian Horse Populations
3.3. Genetic Distances between Native Ethiopian Horse Populations
3.4. Nucleotide Sequence Divergence and Genetic Differentiation
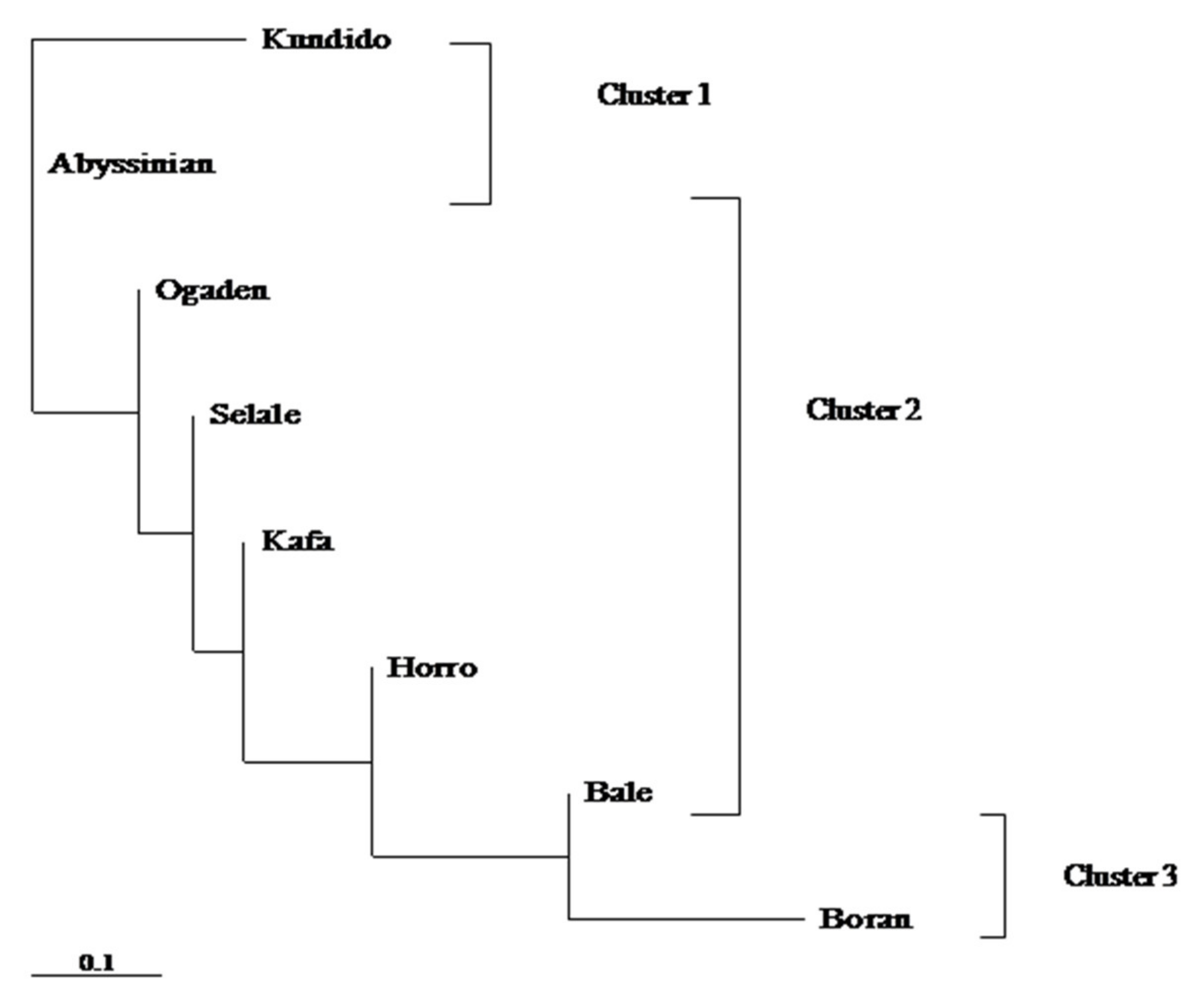
3.5. Matrilineal Genetic Lineage of Native Ethiopian Horse Populations
4. Discussion and Conclusions
4.1. Haplotype Diversities in Native Ethiopian Horse Populations
4.2. Genetic Diversities and Historical Connections in Native Ethiopian Horse Populations
4.3. Matrilineal Genetic Root of Native Ethiopian Horse Populations
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Diamond, J. Guns, Germs and Still: The Fates of Human Societies; W. W. Norton: New York, NY, USA, 2005. [Google Scholar]
- Olsen, S.L. Early Horse Domestication on the Eurasian Steppe. In Documenting Domestication New Genetic and Archaeological Paradigms; Zeder, M.A., Bradley, D.G., Emshwiller, E., Smith, B.D., Eds.; University of California Press: Berkeley, CA, USA, 2006; pp. 245–269. [Google Scholar]
- Beja-Pereira, A.; England, P.R.; Ferrand, N.; Jordan, S.; Bakhiet, A.O.; Abdalla, M.A.; Mashkour, M.; Jordana, J.; Taberlet, P.; Luikart, G. African origin of domestic donkey. Science 2004, 304, 1781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vila, C.; Leonard, A.J.; Gotherstrom, A.; Marklund, S.; Sandberg, K.; Lide, K.; Wayne, R.K.; Ellegren, H. Widespread origins of domestic horse lineages. Science 2001, 291, 474–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jansen, T.; Forster, P.; Levine, M.A.; Oelke, H.; Hurles, M.; Renfrew, C.; Weber, J.; Olek, K. Mitochondrial DNA and the origins of the domestic horse. Proc. Natl. Acad. Sci. USA 2002, 99, 10905–10910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Epstein, H. The Origin of the Domestic Animals of Africa; African Publishing Corporation (APC): New York, NY, USA, 1971. [Google Scholar]
- Goodall, D.M. Horses of the World; Macmillan: New York, NY, USA, 1973. [Google Scholar]
- Bowling, A.T.; Ruvinsky, A. Genetic Aspects of Domestication, Breeds and Their Origins. In The Genetics of the Horse; CAB International: Wallingford, UK, 2000; pp. 25–48. [Google Scholar]
- Khadka, R. Global Horse Population with Respect to Breeds and Risk Status. Master’s Thesis, Swedish University of Agricultural Sciences, Uppsala, Sweden, 2010. [Google Scholar]
- Pankhurst, R. The early history of Ethiopian horse-names. Paideuma 1989, 35, 197–206. [Google Scholar]
- Kefena, E.; Dessie, T.; Han, J.; Kurtu, M.; Rosenbom, S.; Beja-Pereira, A. Morphological diversities and ecozones of Ethiopian horse populations. Anim. Genet. Resour./Ressour. Génét. Anim./Recur. Genét. Anim. 2012, 50, 1–12. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL-W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acid Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bandelt, H.J.; Forster, P.; Rohl, A. Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Koban, E.; Denizci, M.; Aslan, O.; Aktoprakligil, D.; Aksu, S.; Bower, M.; Balcioğlu, B.K.; Bahadir, A.O.; Bilgin, R.; Erdag, B.; et al. High microsatellite and mitochondrial diversity in Anatolian native horse breeds shows Anatolia as a genetic conduit between Europe and Asia. Anim. Genet. 2011, 43, 401–409. [Google Scholar] [CrossRef] [PubMed]
- Slatkin, M. A Measure of Population Subdivision Based on Microsatellite Allele Frequencies. Genetics 1995, 139, 457–462. [Google Scholar] [CrossRef] [PubMed]
- Excoffier, L.; Lischer, H.E.L. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef] [PubMed]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis ver-sion 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nei, M. Molecular Evolutionary Genetics; Columbia University Press: New York, NY, USA, 1987. [Google Scholar]
- Tajima, F. Evolutionary relationship of DNA sequences in finite populations. Genetics 1983, 105, 437–460. [Google Scholar] [CrossRef] [PubMed]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gubitz, T.; Thorpe, R.S.; Malhotra, A. Phylogeographic and natural selection in the Tenerife gecko Tarentoal delalandii: Testing historical and adaptive hypothesis. Mol. Ecol. 2000, 9, 1213–1221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.H.; Kim, K.I.; Cothran, E.G.; Flannery, A.R. Genetic diversity of Cheju horses (Equus caballus) determined by using mitochondrial DNA D-loop polymorphism. Biochem. Genet. 2002, 40, 175–186. [Google Scholar] [CrossRef] [PubMed]
- Lopez, M.S.; Mendonca, D.; Cymbron, T.; Valera, M.; da Costa-Fereira, J.; da Camara Machado, A. The Lusitano horse maternal lineage based on mitochondrial D-loop sequence variation. Animal. Genet. 2005, 36, 196–202. [Google Scholar] [CrossRef] [PubMed]
- Ivankovic, A.; Ramljak, J.; Konjačić, M.; Kelava, N.; Dovč, P.; Mijić, P. Mitochondrial D-loop sequence variation among autochthonous horse breeds in Croatia. Czech J. Anim. Sci. 2009, 54, 101–111. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
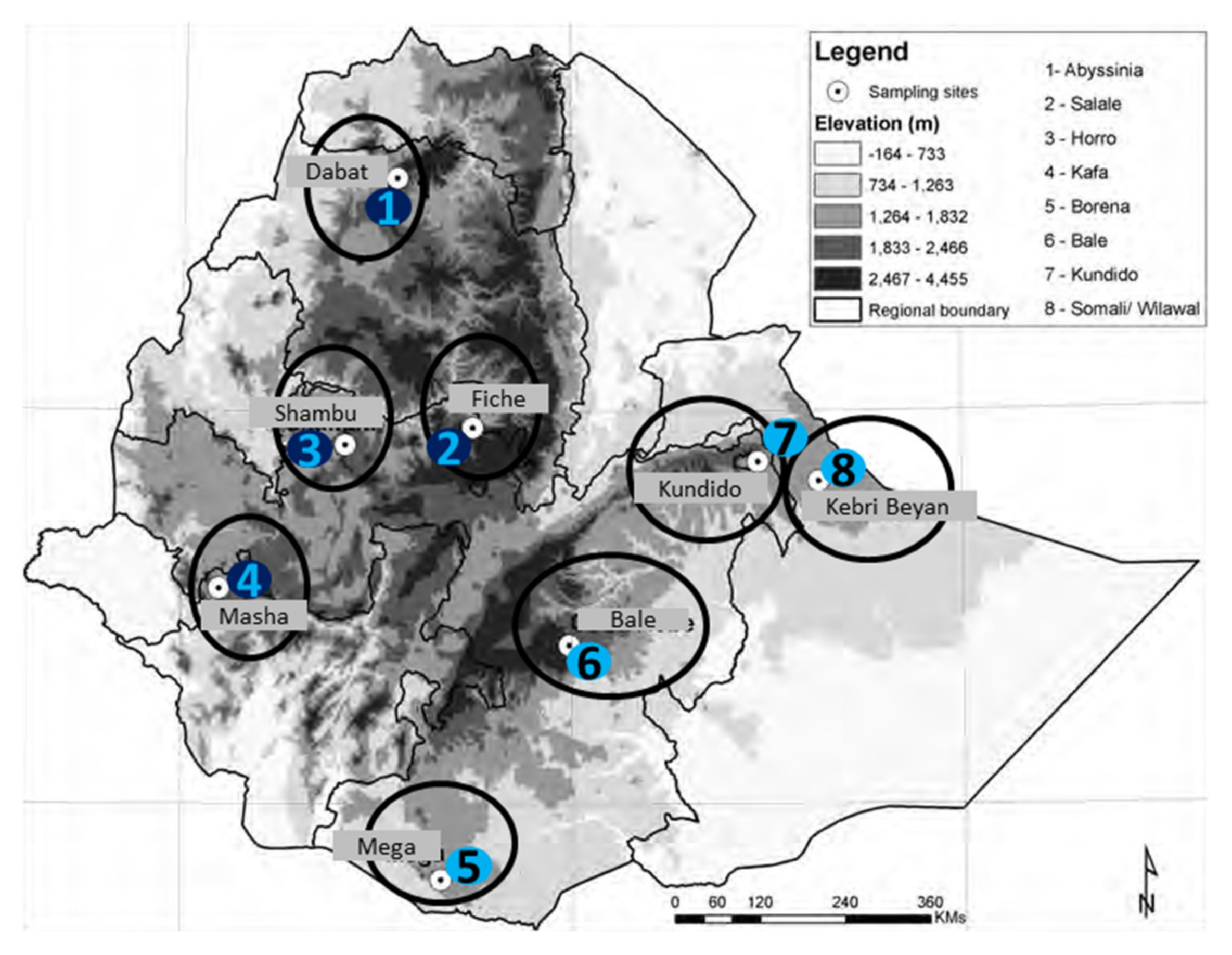
| No | Sampling Site | Horse Population Represented |
|---|---|---|
| 1 | Dabat | Abyssinia |
| 2 | Goba | Bale |
| 3 | Mega | Borana |
| 4 | Shambu | Horro |
| 5 | Masha | Kafa |
| 6 | Kundido | Kundido |
| 7 | Kebri Bayan | Ogaden |
| 8 | Fiche | Selale |
| No | Population Name | No. of Samples | Gender | |
|---|---|---|---|---|
| M | F | |||
| 1 | Abyssinia | 5 | 0 | 5 |
| 2 | Bale | 5 | 3 | 2 |
| 3 | Borana | 6 | 0 | 6 |
| 4 | Horro | 6 | 2 | 4 |
| 5 | Kafa | 6 | 3 | 3 |
| 6 | Kundido | 6 | 3 | 3 |
| 7 | Ogaden | 6 | 6 | 0 |
| 8 | Selale | 6 | 5 | 1 |
| 46 | 22 | 24 | ||
| Population | n | S | Hp | Hd ± S.D. | π ± S.D. | K |
|---|---|---|---|---|---|---|
| Abyssinian | 5 | 13 | 3 | 0.800 ± 0.164 | 0.017 ± 0.005 | 7.600 |
| Bale | 5 | 13 | 3 | 0.700 ± 0.218 | 0.016 ± 0.005 | 7.400 |
| Boran | 6 | 13 | 2 | 0.333 ± 0.215 | 0.010 ± 0.006 | 4.330 |
| Horro | 6 | 13 | 3 | 0.733 ± 0.155 | 0.017 ± 0.003 | 7.467 |
| Kafa | 6 | 13 | 3 | 0.733 ± 0.155 | 0.015 ± 0.004 | 6.733 |
| Kundido feral | 6 | 2 | 2 | 0.600 ± 0.129 | 0.003 ± 0.001 | 1.200 |
| Ogaden | 6 | 13 | 3 | 0.800 ± 0.122 | 0.015 ± 0.004 | 6.933 |
| Selale | 6 | 13 | 3 | 0.800 ± 0.122 | 0.015 ± 0.004 | 6.933 |
| Average | 0.706 ± 0.032 | 0.014 ± 0.001 | 6.320 |
| Population | Abyssinian | Bale | Boran | Horro | Kafa | Kundido Feral | Ogaden | Selale |
|---|---|---|---|---|---|---|---|---|
| Abyssinian | 0 | |||||||
| Bale | 0.149 | 0 | ||||||
| Boran | 0.837 | 0.000 | 0 | |||||
| Horro | 0.055 | 0.000 | 0.064 | 0 | ||||
| Kafa | 0.000 | 0.000 | 0.385 | 0.000 | 0 | |||
| Kundido feral | 0.092 | 0.863 | 2.734 | 0.5000 | 0.218 | 0 | ||
| Ogaden | 0.000 | 0.000 | 0.351 | 0.000 | 0.000 | 0.148 | 0 | |
| Selale | 0.000 | 0.000 | 0.351 | 0.000 | 0.000 | 0.148 | 0.000 | 0 |
| Within population | 0.011 | 0.016 | 0.010 | 0.016 | 0.015 | 0.003 | 0.015 | 0.015 |
| Population | Abyssinian | Bale | Boran | Horro | Kafa | Kundido Feral | Ogaden | Selale |
|---|---|---|---|---|---|---|---|---|
| Abyssinian | - | 0.002 | 0.009 | 0.001 | 0 | 0.000 | 0 | 0 |
| Bale | 0.130 | - | 0 | 0 | 0 | 0.007 | 0 | 0 |
| Boran | 0.497 | 0 | - | 0.001 | 0.006 | 0.018 | 0.005 | 0.005 |
| Horro | 0.059 | 0 | 0.104 | - | 0 | 0.005 | 0 | 0 |
| Kafa | 0 | 0 | 0.325 | 0 | - | 0.002 | 0 | 0 |
| Kundido feral | 0.059 | 0.434 | 0.768 | 0.333 | 0.179 | - | 0.001 | 0.001 |
| Ogaden | 0 | 0 | 0.306 | 0 | 0 | 0.129 | - | 0 |
| Selale | 0 | 0 | 0.306 | 0 | 0 | 0.129 | 0 | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Effa, K.; Rosenbom, S.; Han, J.; Dessie, T.; Beja-Pereira, A. Genetic Diversities and Historical Dynamics of Native Ethiopian Horse Populations (Equus caballus) Inferred from Mitochondrial DNA Polymorphisms. Genes 2021, 12, 155. https://doi.org/10.3390/genes12020155
Effa K, Rosenbom S, Han J, Dessie T, Beja-Pereira A. Genetic Diversities and Historical Dynamics of Native Ethiopian Horse Populations (Equus caballus) Inferred from Mitochondrial DNA Polymorphisms. Genes. 2021; 12(2):155. https://doi.org/10.3390/genes12020155
Chicago/Turabian StyleEffa, Kefena, Sonia Rosenbom, Jianlin Han, Tadelle Dessie, and Albano Beja-Pereira. 2021. "Genetic Diversities and Historical Dynamics of Native Ethiopian Horse Populations (Equus caballus) Inferred from Mitochondrial DNA Polymorphisms" Genes 12, no. 2: 155. https://doi.org/10.3390/genes12020155