
Oregano (Origanum vulgare) Extract Enhances Zebrafish (Danio rerio) Growth Performance, Serum and Mucus Innate Immune Responses and Resistance against Aeromonas hydrophila Challenge

 ,
,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Extraction
2.2. Diets Preparation and Feeding Protocol
2.3. Fish Supply and Rearing Conditions
2.4. Growth Performance
2.5. Serum and Skin Mucus Immunity
2.5.1. Sample Collection
2.5.2. Lysozyme Activity
2.5.3. Total Immunoglobulin (Total Ig)
2.5.4. Complement Activity
2.5.5. Enzymes Activity
2.5.6. Total Protein
2.5.7. Malondialdehyde (MDA) Content
2.6. Bacterial Challenge
2.7. Statistical Analysis
3. Results
3.1. Growth Performance
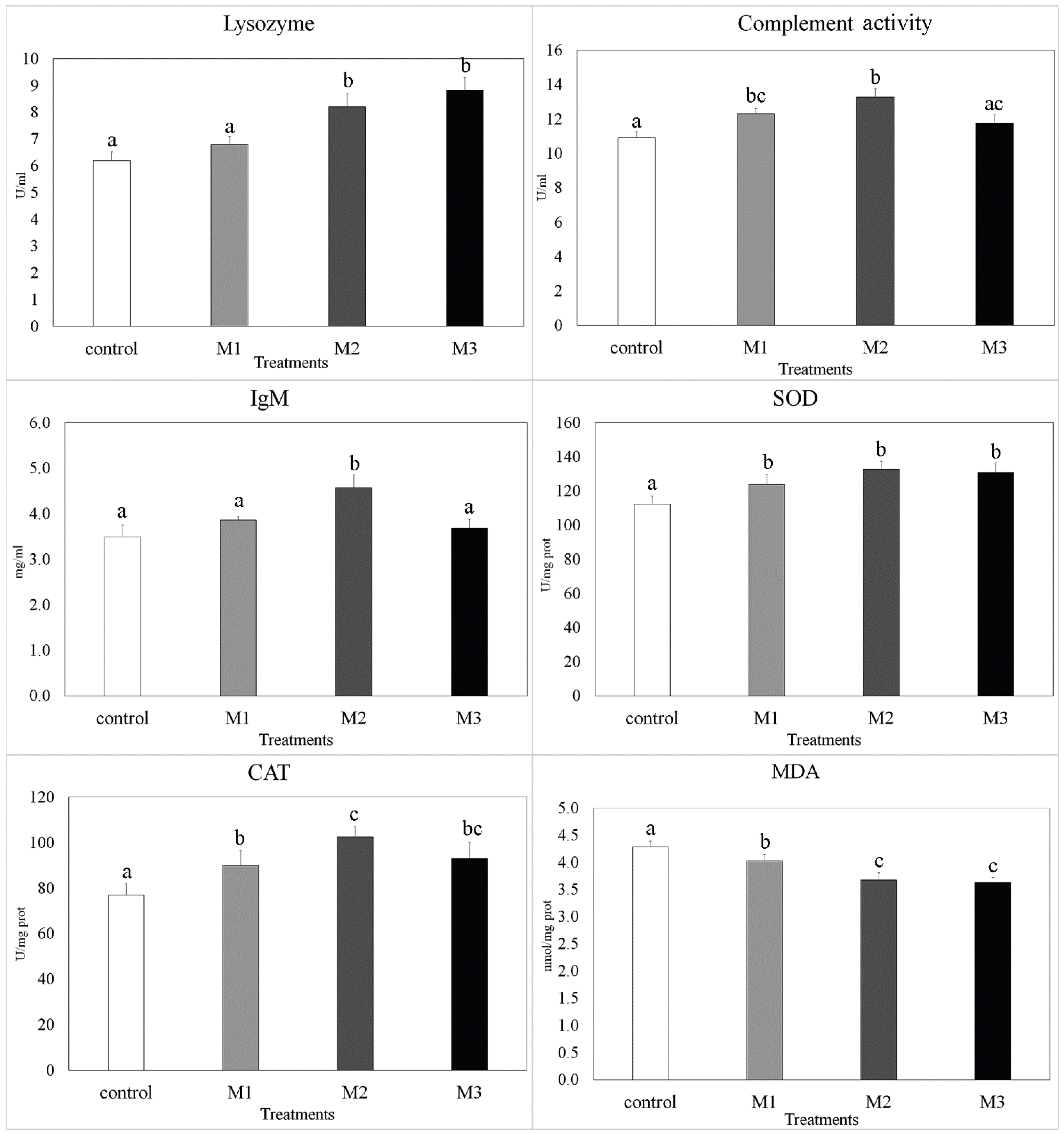
3.2. Immunological and Antioxidant Response Parameters
3.3. Mucus Immune Responses
3.4. Survival Rate
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Immanuel, G.; Vincybai, V.C.; Sivaram, V.; Palavesam, A.; Marian, M. Effect of butanolic extracts from terrestrial herbs and seaweeds on the survival, growth and pathogen (Vibrio parahaemolyticus) load on shrimp Penaeus indicus juveniles. Aquaculture 2004, 236, 53–65. [Google Scholar] [CrossRef]
- Pandey, G.; Sharma, M. Immunostimulant effect of medicinal plants on fish. Int. Res. J. Pharm. 2012, 3, 112–114. [Google Scholar]
- Reverter, M.; Tapissier-Bontemps, N.; Sarter, S.; Sasal, P.; Caruso, D. Moving towards more sustainable aquaculture practices: A meta-analysis on the potential of plant-enriched diets to improve fish growth, immunity and disease resistance. Rev. Aquac. 2020, 1–19. [Google Scholar] [CrossRef]
- Van Hai, N. The use of medicinal plants as immunostimulants in aquaculture: A review. Aquaculture 2015, 446, 88–96. [Google Scholar] [CrossRef]
- Elumalai, P.; Kurian, A.; Lakshmi, S.; Faggio, C.; Esteban, M.A.; Ringø, E. Herbal Immunomodulators in Aquaculture. Rev. Fish. Sci. Aquac. 2020, 1–25. [Google Scholar] [CrossRef]
- Raman, P.R. Applicability, feasibility and efficacy of phytotherapy in aquatic animal health management. Am. J. Plant Sci. 2017. [Google Scholar] [CrossRef] [Green Version]
- Stratev, D.; Zhylyazkov, G.I.; Noundu, X.S.; Krause, R. Beneficial effects of medicinal plants in fish diseases. Aquac. Int. 2008. [Google Scholar] [CrossRef]
- Harikrishnan, R.; Moon, Y.-G.; Kim, M.-C.; Kim, J.-S.; Heo, M.-S.; Balasundaram, C.; Dharaneedharan, S. Phytotherapy of ae romonas hydrophila-infected goldfish, carassius auratus. J. World Aquac. Soc. 2010, 41, 391–401. [Google Scholar] [CrossRef]
- Chakraborty, S.B.; Hancz, C. Application of phytochemicals as immunostimulant, antipathogenic and antistress agents in finfish culture. Rev. Aquac. 2011, 3, 103–119. [Google Scholar] [CrossRef]
- Wang, C.; Liu, H.; Mu, G.; Lu, S.; Wang, D.; Jiang, H.; Sun, X.; Han, S.; Liu, Y. Effects of traditional Chinese medicines on immunity and culturable gut microflora to Oncorhynchus masou. Fish Shellfish. Immunol. 2019, 93, 322–327. [Google Scholar] [CrossRef]
- Doan, H.V.; Soltani, E.; Ingelbrecht, J.; Soltani, M. Medicinal herbs and plants: Potential treatment of monogenean infections in fish. Rev. Fish. Sci. Aquac. 2020, 28, 260–282. [Google Scholar] [CrossRef]
- Ardó, L.; Guojun, Y.; Jeney, Z.; Pao, X.; Jeney, G. Effect of fish feed containing two Chinese herbal extracts (Ganodema Iucidum and Lonicera japonica) on the non-specific immune system of Nile tilapia, Oreochromis niloticus (preliminary results). Acta Agrar. Debr. 2007, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Yin, G.; Jeney, G.; Racz, T.; Xu, P.; Jun, X.; Jeney, Z. Effect of two Chinese herbs (Astragalus radix and Scutellaria radix) on non-specific immune response of tilapia, Oreochromis niloticus. Aquaculture 2006, 253, 39–47. [Google Scholar] [CrossRef]
- Plhalova, L.; Sehonova, P.; Blahová, J.; Doubkova, V.; Tichy, F.; Faggio, C.; Berankova, P.; Svobodova, Z. Evaluation of tramadol hydrochloride toxicity to juvenile zebrafish—morphological, antioxidant and histological responses. Appl. Sci. 2020, 10, 2349. [Google Scholar] [CrossRef] [Green Version]
- Blahova, J.; Cocilovo, C.; Plhalova, L.; Svobodova, Z.; Faggio, C. Embryotoxicity of atrazine and its degradation products to early life stages of zebrafish (Danio rerio). Environ. Toxicol. Pharmacol. 2020, 77, 103370. [Google Scholar] [CrossRef]
- Petrovici, A.; Strungaru, S.-A.; Nicoara, M.; Robea, M.A.; Solcan, C.; Faggio, C. Toxicity of deltamethrin to zebrafish gonads revealed by cellular biomarkers. J. Mar. Sci. Eng. 2020, 8, 73. [Google Scholar] [CrossRef] [Green Version]
- Vokou, D.; Kokkini, S.; Bessiere, J.-M. Geographic variation of Greek oregano (Origanum vulgare ssp. hirtum) essential oils. Biochem. Syst. Ecol. 1993, 21, 287–295. [Google Scholar] [CrossRef]
- Teixeira, B.; Marques, A.; Ramos, C.; Serrano, C.; Matos, O.; Neng, N.R.; Nogueira, J.M.F.; Saraiva, J.A.; Nunes, M.L. Chemical composition and bioactivity of different oregano (Origanum vulgare) extracts and essential oil. J. Sci. Food Agric. 2013, 93, 2707–2714. [Google Scholar] [CrossRef]
- Morshedloo, M.R.; Pirali Hamedani, M.; Yazdani, D. An over review to Origanum vulgare L. and its pharmacological properties. J. Med. Plant. 2018, 4, 15–31. [Google Scholar]
- Bulfon, C.; Volpatti, D.; Galeotti, M. In vitro antibacterial activity of plant ethanolic extracts against fish pathogens. J. World Aquac. Soc. 2014, 45, 545–557. [Google Scholar] [CrossRef]
- Beltrán, J.M.G.; Espinosa, C.; Guardiola, F.A.; Esteban, M.Á. In vitro effects of Origanum vulgare leaf extracts on gilthead seabream (Sparus aurata L.) leucocytes, cytotoxic, bactericidal and antioxidant activities. Fish Shellfish. Immunol. 2018, 79, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Pourmoghim, H.; Haghighi, M.; dan Rohani, M.S. Effect of dietary inclusion of Origanum vulgare extract on nonspecific immune response and hematological parameters of rainbow trout (Oncorhynchusmykiss). Bull. Environ. Pharmacol. Life Sci. 2015, 4, 33–39. [Google Scholar]
- Beltrán, J.M.G.; Silvera, D.G.; Ruiz, C.E.; Campo, V.; Chupani, L.; Faggio, C.; Esteban, M.Á. Effects of dietary Origanum vulgare on gilthead seabream (Sparus aurata L.) immune and antioxidant status. Fish Shellfish. Immunol. 2020, 99, 452–461. [Google Scholar] [CrossRef] [PubMed]
- Oroian, M.; Dranca, F.; Ursachi, F. Comparative evaluation of maceration, microwave and ultrasonic-assisted extraction of phenolic compounds from propolis. J. Food Sci. Technol. 2020, 57, 70–78. [Google Scholar] [CrossRef] [PubMed]
- McDonald, S.; Prenzler, P.D.; Antolovich, M.; Robards, K. Phenolic content and antioxidant activity of olive extracts. Food Chem. 2001, 73, 73–84. [Google Scholar] [CrossRef]
- Zhishen, J.; Mengcheng, T.; Jianming, W. The determination of flavonoid contents in mulberry and their scavenging effects on superoxide radicals. Food Chem. 1999, 64, 555–559. [Google Scholar] [CrossRef]
- Du, R.-Y.; Chen, J.-X.; Zhu, J.; Feng, J.-Y.; Luo, L.; Lin, S.-M.; Chen, Y.-J. Glucose homeostasis and glucose tolerance were impaired with elevated lipid to starch ratios in practical diets for the omnivorous genetically improved farmed tilapia Oreochromis niloticus. Aquaculture 2020, 523, 735221. [Google Scholar] [CrossRef]
- Sun, S.; Ye, J.; Chen, J.; Wang, Y.; Chen, L. Effect of dietary fish oil replacement by rapeseed oil on the growth, fatty acid composition and serum non-specific immunity response of fingerling black carp, Mylopharyngodon piceus. Aquac. Nutr. 2010, 17, 441–450. [Google Scholar] [CrossRef]
- Pedroso, G.L.; Hammes, T.O.; Escobar, T.D.; Fracasso, L.B.; Forgiarini, L.F.; da Silveira, T.R. Blood collection for biochemical analysis in adult zebrafish. J. Vis. Exp. 2012, e3865. [Google Scholar] [CrossRef] [Green Version]
- Ellis, A.E. Lysozyme assays. In Techniques in Fish Immunology; Stolen, J.S., Fletcher, T.C., Anderson, D.P., Roberson, B.S., van Muiswinkel, W.B., Eds.; SOS Publications: Fair Heaven, NJ, USA, 1990; pp. 101–103. [Google Scholar]
- Yano, T. The Nonspecific Immune System: Humoral Defense. In Fish Physiology; Iwama, G., Nakahishi, T., Eds.; Elsevier BV: Amsterdam, The Netherlands, 1996; pp. 105–157. [Google Scholar]
- Lovrić, J.; Mesić, M.; Macan, M.; Koprivanac, M.; Kelava, M.; Bradamante, V. Measurement of malondialdehyde (MDA) level in rat plasma after simvastatin treatment using two different analytical methods. Period. Biol. 2008, 110, 63–68. [Google Scholar]
- Strähle, U.; Scholz, S.; Geisler, R.; Greiner, P.; Hollert, H.; Rastegar, S.; Schumacher, A.; Selderslaghs, I.; Weiss, C.; Witters, H.; et al. Zebrafish embryos as an alternative to animal experiments—A commentary on the definition of the onset of protected life stages in animal welfare regulations. Reprod. Toxicol. 2012, 33, 128–132. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, C.; Kim, C.H. Zebrafish as a model for infectious disease and immune function. Fish Shellfish. Immunol. 2008, 25, 341–350. [Google Scholar] [CrossRef] [PubMed]
- Safari, R.; Hoseinifar, S.H.; Van Doan, H.; Dadar, M. The effects of dietary Myrtle (Myrtus communis) on skin mucus immune parameters and mRNA levels of growth, antioxidant and immune related genes in zebrafish (Danio rerio). Fish Shellfish. Immunol. 2017, 66, 264–269. [Google Scholar] [CrossRef] [PubMed]
- Yanishlieva, N.V.; Marinova, E.M.; Gordon, M.H.; Raneva, V.G. Antioxidant activity and mechanism of action of thymol and carvacrol in two lipid systems. Food Chem. 1999, 64, 59–66. [Google Scholar] [CrossRef]
- Lagouri, V.; Boskou, D. Nutrient antioxidants in oregano. Int. J. Food Sci. Nutr. 1996, 47, 493–497. [Google Scholar] [CrossRef]
- Kulisic, T.; Radonic, A.; Katalinic, V.; Milos, M. Use of different methods for testing antioxidative activity of oregano essential oil. Food Chem. 2004, 85, 633–640. [Google Scholar] [CrossRef]
- Zheng, Z.; Tan, J.Y.W.; Liu, H.; Zhou, X.H.; Xiang, X.; Wang, K.Y. Evaluation of oregano essential oil (Origanum heracleoticum L.) on growth, antioxidant effect and resistance against Aeromonas hydrophila in channel catfish (Ictalurus punctatus). Aquaculture 2009, 292, 214–218. [Google Scholar] [CrossRef]
- Abdel-Latif, H.M.R.; Khalil, R.H. Evaluation of two Phytobiotics, Spirulina platensis and Origanum vulgare extract on Growth, Serum antioxidant activities and Resistance of Nile tilapia (Oreochromis niloticus) to pathogenic Vibrio alginolyticus. Int. J. Fish. Aquat. Stud. 2014, 250, 250–255. [Google Scholar]
- Diler, O.; Gormez, O.; Diler, I.; Metin, S. Effect of oregano (Origanum onites L.) essential oil on growth, lysozyme and antioxidant activity and resistance against Lactococcus garvieaein rainbow trout, Oncorhynchus mykiss (Walbaum). Aquac. Nutr. 2017, 23, 844–851. [Google Scholar] [CrossRef]
- Ahmadifar, E.; Sheikhzadeh, N.; Roshanaei, K.; Dargahi, N.; Faggio, C. Can dietary ginger (Zingiber officinale) alter biochemical and immunological parameters and gene expression related to growth, immunity and antioxidant system in zebrafish (Danio rerio)? Aquaculture 2019, 507, 341–348. [Google Scholar] [CrossRef]
- Jian, J.; Wu, Z. Effects of traditional Chinese medicine on nonspecific immunity and disease resistance of large yellow croaker, Pseudosciaena crocea (Richardson). Aquaculture 2003, 218, 1–9. [Google Scholar] [CrossRef]
- Citarasu, T. Herbal biomedicines: A new opportunity for aquaculture industry. Aquac. Int. 2010, 18, 403–414. [Google Scholar] [CrossRef]
- Harikrishnan, R.; Balasundaram, C.; Heo, M.-S. Impact of plant products on innate and adaptive immune system of cultured finfish and shellfish. Aquaculture 2011, 317, 1–15. [Google Scholar] [CrossRef]
- Ahmad, M.H.; El Mesallamy, A.M.D.; Samir, F.; Zahran, F. Effect of Cinnamon (Cinnamomum zeylanicum) on growth performance, feed utilization, whole-body composition, and resistance to Aeromonas hydrophilain Nile Tilapia. J. Appl. Aquac. 2011, 23, 289–298. [Google Scholar] [CrossRef]
- Ahmad, M.H.; Abdel-Tawwab, M. The use of caraway seed meal as a feed additive in fish diets: Growth performance, feed utilization, and whole-body composition of Nile tilapia, Oreochromis niloticus (L.) fingerlings. Aquaculture 2011, 314, 110–114. [Google Scholar] [CrossRef]
- Alexander, J.B.; Ingram, G.A. Noncellular nonspecific defence mechanisms of fish. Annu. Rev. Fish Dis. 1992, 2, 249–279. [Google Scholar] [CrossRef]
- Tukmechi, A.; Bandboni, M. Effects of Saccharomyces cerevisiae supplementation on immune response, hematological parameters, body composition and disease resistance in rainbow trout, Oncorhynchus mykiss (Walbaum, 1792). J. Appl. Ichthyol. 2014, 30, 55–61. [Google Scholar] [CrossRef]
- Hoseinifar, S.H.; Yousefi, S.; Capillo, G.; Paknejad, H.; Khalili, M.; Tabarraei, A.; Van Doan, H.; Spanò, N.; Faggio, C. Mucosal immune parameters, immune and antioxidant defence related genes expression and growth performance of zebrafish (Danio rerio) fed on Gracilaria gracilis powder. Fish Shellfish. Immunol. 2018, 83, 232–237. [Google Scholar] [CrossRef]
- Farsani, M.N.; Hoseinifar, S.H.; Rashidian, G.; Ghafarifarsani, H.; Ashouri, G.; Van Doan, H. Dietary effects of Coriandrum sativum extract on growth performance, physiological and innate immune responses and resistance of rainbow trout (Oncorhynchus mykiss) against Yersinia ruckeri. Fish Shellfish. Immunol. 2019, 91, 233–240. [Google Scholar] [CrossRef]
- Rashidian, G.; Kajbaf, K.; Prokić, M.D.; Faggio, C. Extract of common mallow (Malvae sylvestris) enhances growth, immunity, and resistance of rainbow trout (Oncorhynchus mykiss) fingerlings against Yersinia ruckeri infection. Fish Shellfish. Immunol. 2020, 96, 254–261. [Google Scholar] [CrossRef]
- Dügenci, S.K.; Arda, N.; Candan, A. Some medicinal plants as immunostimulant for fish. J. Ethnopharmacol. 2003, 88, 99–106. [Google Scholar] [CrossRef]
- Ahmadifar, E.; Dawood, M.A.O.; Moghadam, M.S.; Sheikhzadeh, N.; Hoseinifar, S.H.; Musthafa, M.S. Modulation of immune parameters and antioxidant defense in zebrafish (Danio rerio) using dietary apple cider vinegar. Aquaculture 2019, 513, 734412. [Google Scholar] [CrossRef]
- Rafieepour, A.; Hajirezaee, S.; Rahimi, R. Dietary oregano extract (Origanum vulgare L.) enhances the antioxidant defence in rainbow trout, Oncorhynchus mykiss against toxicity induced by organophosphorus pesticide, diazinon. Toxin Rev. 2020, 39, 397–407. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, C.; Zhang, J.; Xie, J.; Yang, L.; Xing, Y.; Li, Z. The effects of quercetin on immunity, antioxidant indices, and disease resistance in zebrafish (Danio rerio). Fish Physiol. Biochem. 2020, 46, 759–770. [Google Scholar] [CrossRef] [PubMed]
- Riche, M. Analysis of refractometry for determining total plasma protein in hybrid striped bass (Morone chrysops × M. saxatilis) at various salinities. Aquaculture 2007, 264, 279–284. [Google Scholar] [CrossRef]
- Fazio, F.; Marafioti, S.; Torre, A.; Sanfilippo, M.; Panzera, M.; Faggio, C. Haematological and serum protein profiles of Mugil cephalus: Effect of two different habitats. Ichthyol. Res. 2013, 60, 36–42. [Google Scholar] [CrossRef]
- Suwan, C.; Noimoon, P.; Yawichai, P.; Jitmanowan, S.; Chitmanat, C. Effects of medicinal plants on fish immunity and its growth performances. Burapha Sci. J. 2020, 25, 595–616. [Google Scholar]
- Mohammadi, G.; Rashidian, G.; Hoseinifar, S.H.; Naserabad, S.S.; Van Doan, H. Ginger (Zingiber officinale) extract affects growth performance, body composition, haematology, serum and mucosal immune parameters in common carp (Cyprinus carpio). Fish Shellfish. Immunol. 2020, 99, 267–273. [Google Scholar] [CrossRef]
- Aragona, M.; Lauriano, E.R.; Pergolizzi, S.; Faggio, C. Opuntia ficus-indica (L.) Miller as a source of bioactivity compounds for health and nutrition. Nat. Prod. Res. 2017, 32, 2037–2049. [Google Scholar] [CrossRef]
- Van Doan, H.; Hoseinifar, S.H.; Sringarm, K.; Jaturasitha, S.; Yuangsoi, B.; Dawood, M.A.O.; Esteban, M.A.; Ringø, E.; Faggio, C. Effects of assam tea extract on growth, skin mucus, serum immunity and disease resistance of Nile tilapia (Oreochromis niloticus) against Streptococcus agalactiae. Fish Shellfish. Immunol. 2019, 93, 428–435. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Proximate Composition (%) | Control | M1 (0.5%) | M2 (1%) | M3 (2%) |
|---|---|---|---|---|
| Dry matter | 92.44 | 92.14 | 92.48 | 92.66 |
| Ash | 13.66 | 12.99 | 13.42 | 13.84 |
| Crude protein | 51.69 | 50.97 | 51.48 | 52.19 |
| Crude lipids | 11.94 | 11.92 | 11.53 | 11.25 |
| Growth Parameters | Control | M1 (0.5%) | M2 (1%) | M3 (2%) |
|---|---|---|---|---|
| Initial weight (mg) | 81.13 ± 0.87 a | 82.58 ± 1.17 a | 81.62 ± 1.05 a | 81.03 ± 1.27 a |
| Final weight (mg) | 190.46 ± 1.87 a | 194.52 ± 1.97 a | 199.19 ± 2.9 b | 194.33 ± 1.77 a |
| WG (mg) | 109.32 ± 1.15 a | 111.93 ± 2.23 a | 117.57 ± 3.94 b | 113.30 ± 1.07 ab |
| WG (%) | 134.74 ± 1.12 a | 135.57 ± 4.00 a | 144.10 ± 6.67 b | 139.83 ± 2.41 ab |
| FCR | 1.92 ± 0.06 a | 1.87 ± 0.03 ab | 1.82 ± 0.03 b | 1.85 ± 0.02 ab |
| SGR (%/d) | 1.42 ± 0.01 a | 1.43 ± 0.03 a | 1.49 ± 0.05 b | 1.46 ± 0.02 ab |
| SR (%) | 100 ± 0 | 100 ± 0 | 100 ± 0 | 100 ± 0 |
| Treatment | Control | M1 (0.5%) | M2 (1%) | M3 (2%) |
|---|---|---|---|---|
| Lysozyme (U mL−1) | 31.23 ± 0.67 a | 35.71 ± 1.34 b | 41.20 ± 1.93 c | 35.11 ± 1.45 b |
| ALP (U mL−1) | 12.83 ± 1.68 a | 17.92 ± 1.54 b | 23.66 ± 2.23 c | 20.37 ± 0.87 b |
| Protease | 24.88 ± 1.77 a | 27.88 ± 1.19 ab | 31.04 ± 2.06 b | 25.79 ± 1.63 a |
| Total Ig (mg mL−1) | 9.28 ± 0.85 a | 12.13 ± 0.80 b | 16.81 ± 1.03 c | 15.06 ± 0.72 d |
| Total protein (mg mL−1) | 4.26 ± 0.33 a | 4.61 ± 0.30 ab | 5.70 ± 0.35 c | 5.10 ± 0.24 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rashidian, G.; Boldaji, J.T.; Rainis, S.; Prokić, M.D.; Faggio, C. Oregano (Origanum vulgare) Extract Enhances Zebrafish (Danio rerio) Growth Performance, Serum and Mucus Innate Immune Responses and Resistance against Aeromonas hydrophila Challenge. Animals 2021, 11, 299. https://doi.org/10.3390/ani11020299
Rashidian G, Boldaji JT, Rainis S, Prokić MD, Faggio C. Oregano (Origanum vulgare) Extract Enhances Zebrafish (Danio rerio) Growth Performance, Serum and Mucus Innate Immune Responses and Resistance against Aeromonas hydrophila Challenge. Animals. 2021; 11(2):299. https://doi.org/10.3390/ani11020299
Chicago/Turabian StyleRashidian, Ghasem, Javad Tahmasebi Boldaji, Simona Rainis, Marko D. Prokić, and Caterina Faggio. 2021. "Oregano (Origanum vulgare) Extract Enhances Zebrafish (Danio rerio) Growth Performance, Serum and Mucus Innate Immune Responses and Resistance against Aeromonas hydrophila Challenge" Animals 11, no. 2: 299. https://doi.org/10.3390/ani11020299