
Secretion-Based Modes of Action of Biocontrol Agents with a Focus on Pseudozyma aphidis

Plant Pathology and Microbiology Department, Robert H. Smith Faculty of Agriculture, Food and Environment, the Hebrew University of Jerusalem, Rehovot 76100, Israel
*
Author to whom correspondence should be addressed.
Plants 2021, 10(2), 210; https://doi.org/10.3390/plants10020210
Submission received: 6 December 2020
/
Revised: 19 January 2021
/
Accepted: 19 January 2021
/
Published: 22 January 2021
(This article belongs to the Special Issue Biological Control of Plant Diseases)
Abstract
:Plant pathogens challenge our efforts to maximize crop production due to their ability to rapidly develop resistance to pesticides. Fungal biocontrol agents have become an important alternative to chemical fungicides, due to environmental concerns related to the latter. Here we review the complex modes of action of biocontrol agents in general and epiphytic yeasts belonging to the genus Pseudozyma specifically and P. aphidis in particular. Biocontrol agents act through multiple direct and indirect mechanisms, which are mainly based on their secretions. We discuss the direct modes of action, such as antibiosis, reactive oxygen species-producing, and cell wall-degrading enzyme secretions which can also play a role in mycoparasitism. In addition, we discuss indirect modes of action, such as hyperbiotrophy, induced resistance and growth promotion based on the secretion of effectors and elicitors from the biocontrol agent. Due to their unique characteristics, epiphytic yeasts hold great potential for use as biocontrol agents, which may be more environmentally friendly than conventional pesticides and provide a way to reduce our dependency on fungicides based on increasingly expensive fossil fuels. No less important, the complex mode of action of Pseudozyma-based biocontrol agents can also reduce the frequency of resistance developed by pathogens to these agents.
1. Introduction
The world population is rapidly growing and is expected to reach over 9 billion people by the year 2050. Feeding such a huge population will require a 70–100% increase in total food production [1]. Since most soils with high productivity potential are already under cultivation, and there is constant shrinkage of cultivated land area due to urban and industrial development [2], it is necessary to maximize agricultural yield. Among the most important contributors to low agricultural yield are pests and pathogens, which are responsible for destroying up to 20% of the world’s harvest, with a further 10% loss post-harvest [3,4]. Fungal pathogens cause most of the diseases in agricultural ecosystems and are responsible for the most costly damage to many crops, including some of the most devastating plant disease epidemics in history [5]. Bacteria can also cause diseases in plants [6]. While relatively less agriculturally and economically damaging than fungi [7], bacteria cause many serious diseases of plants throughout the world, especially in moist and warm environments [6,8]. There are a few ways to reduce these devastating losses. One is to develop resistant crops; however, these need 10 to 15 years to reach the field and can lose their resistance within a decade, with the emergence of new pathogen mutations. Although this method is widely used, it is time-consuming and in general, resistance can only be developed for one crop at a time. The second alternative is the use of fungicides or biomolecules to reduce the losses. Continual use of chemical-based pesticides has allowed farmers to control the spread of pathogens and pests, and has helped ensure stable and prosperous agricultural systems [9,10]. However, continuous pesticide use also results in the widespread problem of pathogen resistance because pathogens tend to find ways to suppress the pesticides’ effects through detoxification methods, such as degradation, or by active efflux of the toxic compound out of their cells using specific transporters [11,12]. In fact, multidrug resistance (MDR) is a major challenge to agriculture the world over [13,14,15,16]. One solution is the use of multiple pesticides with different modes of action in combination or in rotation [17]. However, excessive use of fungicides is deleterious to the ecosystem and human health, and concerns over the potential impact of pesticides on the environment are becoming more pressing. Thus, more stringent pesticide registration procedures and regulations have reduced the number of synthetic pesticides available for agricultural practice. Biological control agents (BCAs) and natural product-based pesticides (biopesticides) are good alternatives to the use of traditional synthetic chemicals as they are less harmful to both the environment and human health [18,19,20,21]. Furthermore, since BCAs can hinder pathogen growth and development, thereby reducing diseases, via a complex combination of direct and indirect modes of action—including antibiosis, mycoparasitism, competition and induced resistance in the host plant—the development of pathogen resistance to BCAs is more complicated [22,23]. As a result, there is growing interest in the exploration and exploitation of naturally occurring microorganisms, such as fungi, for the control of crop diseases [24]. The complex modes of action of BCAs are mainly directed by secreted molecules and proteins that can act as antibiotics, effectors, elicitors and degrading enzymes (Figure 1). This review summarizes the known modes of action of the epiphytic yeast BCAs in general, and Pseudozyma aphidis in particular, with a focus on our ability to define the functions of their secretions and possible uses for fungal pathogen control.
2. Antibiosis: Fungal Metabolites as a Source for New Pesticides
Many microorganisms, including a large portion of the known BCAs, produce and secrete secondary metabolites with antimicrobial properties, as part of their constant competition with other microorganisms in their natural environment [25] (Figure 1; Table 1). Those metabolites can be exploited as microbial fungicides that may be more environmentally friendly than chemical pesticides, and at the same time lack some of the disadvantages of using living organisms. Due to their natural origins, microbial pesticides are more biodegradable than synthetic chemicals [26]. Microorganisms can synthesize secondary metabolites with versatile chemical structures and diverse biological activities that exceed the scope of synthetic organic chemistry [27]. Therefore, a newly discovered antifungal compound is more likely to have a new and more specific mode of action than the commercial fungicides, and lack cross-resistance to them [28]. The specificity of microbial pesticides is both an advantage and a disadvantage. While it reduces the risk of toxicity to humans, the environment and other beneficial organisms in the field, it also makes it harder for those pesticides to be effectively applied by farmers. Thus, an additional possible use for microbial pesticides could be as lead compounds for the development of novel synthetic fungicides.
Several strains of Trichoderma spp. have been found to produce antifungal compounds that are toxic to phytopathogenic fungi such as Botrytis cinerea, Rhizoctonia solani and Pythium spp. [29,30,31,32]. For example, Trichoderma virens, Trichoderma harziarum and Trichoderma pseudokoningii produce unique peptaibols, which are linear peptide antibiotics which can form pores in the membrane and induce programmed cell death (PCD) [31,33,34,35,36]. In addition, the yeast BCAs, such as Pseudozyma spp., have been found to secrete antimicrobial compounds. Pseudozyma rugulosa, Pseudozyma flocculosa and P. aphidis all exhibit biological activity against the different powdery mildews with which they are associated [37,38,39,40,41,42,43,44]. While the mechanism of this biological activity has not yet been fully elucidated, P. rugulosa and P. flocculosa were found to produce unusual extracellular fatty acids with antifungal properties, which cause the release of intracellular ions and proteins when in contact with sensitive fungi [45,46,47,48,49,50]. Furthermore, Pseudozyma tsukubaensis and Pseudozyma prolifica were shown to produce a very large mycocin with both fungistatic and fungicidal activities. This toxin has very narrow, taxonomically specific activity against some representatives of the orders Microstromatales and Ustilaginales [51,52]. P. flocculosa was also found to produce an unusual and rare cellobiose lipid with antifungal activity [53]. This glycolipid, named flocculosin, inhibited several pathogenic fungi, including Candida albicans and Trichosporon asahii [54]. In a later work, this group reported that flocculosin also has antibacterial properties, directed rather specifically against both aerobic and anaerobic Gram-positive bacteria. In both cases, the antimicrobial activity was achieved by irreversibly damaging the cell membrane of the microorganisms [55]. Another unusual glycolipid, which has a highly similar chemical structure to ustilagic acid, was produced by two other Pseudozyma species: Pseudozyma fusiformata [56] and Pseudozyma graminicola [57]. While the discovery of flocculosin is relatively new, ustilagic acid was discovered and established as an antimicrobial compound more than 60 years ago. Interestingly, the main producer of ustilagic acid is the phytopathogenic fungus Ustilago maydis [58]. Recent work has shown that P. flocculosa is a close relative of the plant pathogen U. maydis whose pathogenicity trait was lost during evolution [59]. However, new evidence indicates that flocculosin plays a secondary, if any, role in the antagonistic activity of P. flocculosa against powdery mildews. Instead, the biocontrol process seems to be mediated by a more complex interaction [60].
P. aphidis extracts have also been found to inhibit spore germination and linear growth of fungal phytopathogens such as B. cinerea and Alternaria brassicicola [43], as well as important bacterial phytopathogens such as Clavibacter michiganensis subsp. michiganensis, Pseudomonas syringae pv. tomato and Xanthomonas campestris pv. campestris [61].Those metabolites have not yet been identified but they are largely lipophilic and found to induce reactive oxygen species (ROS) accumulation and PCD in B. cinerea hyphal cells [62].
Several commercially used pesticides have been developed from microbial metabolites through chemical modifications, such as fludioxonil, fenpiclonil and strobilurins [26,63,64]. Strobilurins are natural substances isolated mainly from basidiomycetes. The first strobilurin was isolated from liquid culture of Strobilurus tenacellus. Natural strobilurins are highly active against yeasts and filamentous fungi, by inhibiting their mitochondrial respiration. Since they break down rapidly in the light, they are not reliable for disease control. Instead, they were used as lead compounds to develop synthetic fungicides that are more stable and more powerful when used commercially in agricultural practice [65]. Several commercial strobilurin-based products are on the market or in development. Since these products are derived from natural products, they are environmentally safe due to their rapid degradation in the environment [64,66]. Since the discovery of the strobilurins, many other microbial metabolites with promising antimicrobial properties have been discovered through activity-based screening against important phytopathogens. Chaetoviridins, which were purified from the broth of Chaetomium globosum, exhibit high levels of in vitro and in vivo antifungal activity against Magnaporthe grisea and inhibit the development of rice blast disease [67,68]. Fusapyrone and deoxyfusapyrone, isolated from Fusarium semitectum, exhibit a broad range of antifungal activity [69]. Several peptides with control efficacy against plant diseases have also been identified. For example, verlamelin is a peptide produced by the fungus Acremonium strictum with strong inhibitory activity against powdery mildew [70,71].
3. Effectors and Hyperbiotrophy-Dependent Inhibition
Effectors are molecules that are secreted from pathogens or beneficial microorganisms and affect the outcome of their interaction with their host, such as participating in successful colonization or pathogenicity processes (Figure 1 and Figure 2; Table 1). Most of the plant pathogen-secreted effectors function inside the host plant cell and are mostly virulence-promoting factors that inhibit or interfere with the plant’s defense system [72,73,74,75]. A few recent studies have suggested that effectors may also contribute to the BCAs’ ability to control plant pathogens [76,77,78,79]. In Trichoderma spp., some effectors have been shown to be involved in antagonistic effects, such as the cerato-platanin protein Sm1 and its orthologue EPl1 that induce ROS production and pathogenesis-related (PR) gene expression in the host plant [80,81,82]; (For review see Ramírez-Valdespino et al., 2019 [79]). Two other Trichoderma proteins, Sm2 and Epl2, were found to be more relevant to defense activation in the plant host, but their functions are unknown [83,84]. A recent study on Trichoderma spp. identified 16 effector-like proteins during its interaction with Arabidopsis thaliana and the plant pathogen R. solani. The transcription levels of those putative effectors were modified in response to either the host plant or the fungal pathogen, and some had predicted functions, such as proteases, LysM and thioredoxins. One of those genes, Hydii1 from T. virens, was identified as a hydrophobin participating in root colonization and its expression was correlated with the enhanced antagonistic effect of T. virens against R. solani [78]. Other hydrophobins from Trichoderma, such as HBF2-6, Hytlo1 and Hyd1, were found to be involved in colonization and the host plant’s defense response. A study on the beneficial F. oxysporum CS-20 demonstrated recently that CS20EP a small cysteine-rich protein, predicted as secreted effector, elicits ion exchange and defense response against pathogenic F. oxysporum in tomato plants [85]. Studies comparing the genomes and secretomes of the pathogen U. maydis and other smut pathogens with the BCA P. flocculosa demonstrated that these fungi, which have very different lifestyles, share genome features, including hallmarks of pathogenicity [59,86]. This comprehensive work demonstrated that P. flocculosa had lost a specific subset of candidate secreted effectors connected to virulence but had acquired genes encoding secreted proteins believed to have direct relevance to its biocontrol ability against plant pathogens [59]. Another comparative study of four genomes of Pseudozyma species with that of U. maydis found 113 putative secreted effectors in common: some were validated as effectors, such as the Pep1 protein which suppresses microbe-associated molecular pattern (MAMP)-triggered ROS production [87]. An orthologue of Pep1 from Pseudozyma could also complement a Pep1-deficient U. maydis mutant, restoring its virulence [64]. These new subsets of secreted effectors may function as virulence factors against plant pathogens, as we recently demonstrated that a P. aphidis-secreted fraction can activate ROS production and PCD in B. cinerea hyphal cells [44]. A later study demonstrated hyperbiotrophy as a new mode of action, when a unique effector of P. flocculosa directed inside Blumeria graminis (powdery mildew) caused a flow of nutrients into P. flocculosa [77]. This disruption in the nutrient flux caused collapse of the powdery mildew haustoria, and the plant eventually recovered from the pathogen [65].
4. Induced Resistance by Fungal Metabolites
To cope with constant attacks by invading pathogens, plants have evolved a wide range of defense mechanisms, including the use of preexisting physical (e.g., cuticle and cell wall) and chemical (e.g., antimicrobial compounds) barriers [88]. Pathogens and beneficial microbes can be detected by their MAMPs, which are recognized by specific pattern-recognition receptors (PRRs). MAMP–PRR interactions can lead to a weak response termed pattern or MAMP-triggered immunity (Figure 2) [89]. Pathogens that pass these first layers of defense may encounter the hypersensitive response—the rapid induction of localized cell death—which prevents their spread beyond local infection [90,91]. Those pathogens that persist and overcome the hypersensitive response still need to challenge the well-orchestrated plant defense responses, which involve effector-triggered immunity that includes systemic acquired resistance (SAR)(Figure 2) [90,92,93,94]. SAR is a form of induced resistance in plants with a specific defense-signaling pathway that occurs systemically after localized exposure to a pathogen, and is especially useful against biotrophic pathogens. It is characterized by the accumulation of salicylic acid (SA) and the expression of SAR-marker PR genes [95], but it can also be SA-independent [96]. Some microorganisms are associated with induced systemic resistance (ISR), which does not involve the accumulation of SA. Instead, it is mediated by jasmonic acid (JA) and ethylene (ET), which co-regulate a set of defense-related genes, different from those regulated by SA [97,98]. The JA/ET defense-signaling pathway plays an important role in plant defense responses against necrotrophic pathogens and herbivorous insects [97,98,99].
Some beneficial microbes, especially growth-promoting bacteria and fungi in the rhizosphere, can enhance the induced resistance responses in plants, mainly via secreted elicitors (Figure 2; Table 1). While beneficial microbes are generally associated with ISR, they can also activate SAR. The specifically activated pathways may involve JA/ET, SA, or both [43,100,101]. Prolonged activation of inducible defense involves major costs that affect plant growth and reproduction. Therefore, for the SAR/ISR to be affordable and beneficial, it should mainly be activated when the plant is exposed to attack [94,102,103]. To achieve this, beneficial microbes induce a unique physiological state called priming, which causes the plant to respond with faster and stronger activation of the induced defense responses, but only following an attack by pathogens [104].
Several fungal BCAs, mainly of the genus Trichoderma, have been reported to elicit ISR and priming in colonized plants. For example, T. harzianum T39 primed grapevines against downy mildew [105]; and T. asperellum SKT-1 induced resistance in Arabidopsis against the bacterial pathogen P. syringae pv. tomato [106] and modulated the expression of genes involved in the JA/ET-signaling pathways of ISR [107]. The endophytic basidiomycete fungus Piriformospora indica primes various plant species against powdery mildew [108], and the protective Fusarium oxysporum strain Fo47 primes tomato plants against pathogenic F. oxysporum strains [109]. In addition to the direct control of phytopathogens, P. aphidis and P. churashimaensis were found to indirectly protect plants by local and systemic induction of SAR and ISR, and priming the plant’s defense machinery for stronger induced defense activation after pathogen infection [43,110]. However, P. aphidis suppressed MAMP-triggered callose formation on the plant leaf surface, suggesting that it bypasses the MAMP-triggered immunity to become established on the leaf surface, and only later induces plant resistance and priming (Figure 2; unpublished data).
Various chemicals have been discovered that seem to act at various points in the defense-activating networks and mimic all or part of the biological activation of resistance [111]. Two prominent examples of such chemicals are acibenzolar-s-methyl (ASM) and β-aminobutyric acid (BABA). ASM is a functional analogue of SA that acts downstream of SA accumulation through the activation of SA-response mechanisms. Application of this compound induces similar responses in plants to those induced by biotrophic pathogens or SA, including overexpression of PR genes [112], and it has been widely reported to induce resistance against a broad spectrum of pathogens in many plant species. BABA is a non-protein amino acid known to induce resistance against many plant pathogens in a range of crop plants [113] by both SA-dependent and SA-independent defense mechanisms [114].
Many natural compounds have also been claimed to elicit ISR and priming, including oligosaccharides, glycosides, amides, vitamins, carboxylic acids, and aromatic compounds. Most mimic pathogen interactions and are able to induce or prime defense in a concentration-dependent fashion. However, the mode of action of these elicitors is eventually determined by the hosts and the stress that they are facing [115,116,117,118]. Extracts of plants and microorganisms are a potential source for the discovery and isolation of new natural elicitors, such as hexanoic acid which is a natural compound produced by strawberry. This potent natural priming inducer can activate broad-spectrum defenses by inducing callose deposition and the SA and JA pathways [115,119]. While natural elicitors can occasionally be found anywhere in the natural world, as evidenced by the case of hexanoic acid, a more likely source of such compounds is fungal BCAs. Since the elicitation of ISR and priming by fungal BCAs naturally involves some kind of chemical mechanism, extracts of these fungi and their secretions are more likely to contain the natural elicitors, which can be isolated and identified. Hossain et al. [120] reported that root treatment with culture filtrate of the growth-promoting fungus Penicillium simplicissimum induced resistance to P. syringae pv. tomato in Arabidopsis plants [120]. ISR activation against the same pathogen in Arabidopsis plants was also induced by blends of volatile compounds extracted from the growth-promoting fungi Ampelomyces spp. and Cladosporium spp. m-cresol and methyl benzoate were identified as the major active compounds in these blends [121].
As in many cases with fungal BCAs, most of the extensive studies have been performed with Trichoderma spp. Djonović et al. [80] identified and purified a small protein (designated Sm1) from T. virens secretions that triggered production of ROS and induced the expression of defense-related genes, both locally and systemically, in cotton. Pretreatment of cotton cotyledons with Sm1 also provided high levels of protection against the pathogen Colletotrichum sp. [80] (see also Section 3). T. virens also produces unique peptaibols with antibiotic properties (see also Section 2) [31], but it also has the ability to induce resistance. Cucumber plants co-cultivated with T. virens strains disrupted in their ability to produce these peptaibols showed a significantly reduced systemic resistance response to the leaf pathogen P. syringae pv. lachrymans, and reduced ability to produce phenolic compounds with inhibitory activity. On the other hand, two synthetic peptaibol isoforms (TvBI and TvBII) from T. virens induced systemic protection to leaf-pathogenic bacteria, induced antimicrobial compounds in cucumber cotyledons, and upregulated the expression of several defense-related genes [122].
The resistance induced by fungal BCAs can also be elicited by small secreted molecules such as secondary metabolites. Vinale et al. [123] demonstrated that pre-application of secondary metabolites isolated from the culture filtrate of several biocontrol strains of Trichoderma reduces the disease symptoms of B. cinerea on tomato plants, and induces overexpression of PR proteins [123]. We recently found that application of P. aphidis extract (a fraction that does not contain antibiotic activity) can induce PR genes in tomato plants and activate the plant defense system (Harris R and Levy M unpublished data).
5. Mycoparasitisim and Cell Wall-Degrading Enzymes
The secreted fraction of BCAs contains factors other than antibiotics and elicitors/effectors that might play a part in their ability to control pathogens. Some of these are cell wall-degrading enzymes and proteases which have been well documented as being part of the mycoparasitic mode of action (Figure 1; Table 1). As early as 1932, Weindling [124] described mycoparasitism of R. solani hyphae by the hyphae of Trichoderma lignorum, including coiling around the pathogen hyphae, and penetration and subsequent dissolution of the R. solani cytoplasm [30]. Mycoparasitism was demonstrated as one of the BCA’s direct modes of action that relies on the production of fungal cell wall-degrading enzymes such as chitinases, the latter having been shown to affect pathogenicity of germinating conidia of B. cinerea [125,126]. However, it has also been shown that some Trichoderma spp. isolates that have the potential to produce cell wall-degrading enzymes do not necessarily provide good biocontrol activity [127].
The antagonistic activity of P. aphidis against powdery mildew seems to be mainly parasitic. Scanning electron microscopy analysis revealed that P. aphidis dimorph from yeast-like to hypha-like and ecto-parasitizes the powdery mildew hypha by coiling around it [44]. Similar mycoparasitism occurs with P. flocculosa on conidia of cucumber powdery mildew [59]. In contrast, we have shown that to control gray mold disease caused by B. cinerea, P. aphidis remains as a yeast-like structure and thus does not coil around B. cinerea hyphae [44]. However, the P. aphidis cells could adhere to B. cinerea hyphae and secrete lytic enzymes and proteases when exposed to B. cinerea cell wall extract, suggesting that close proximity enhances the antibiotic activity and cell wall-degrading enzyme efficiency, thereby also supporting the parasitism dogma [62]. The dimorphic switch from hypha-like to yeast-like is a well-known phenomenon in fungal pathogens [128]. In C. albicans, this dimorphism is essential for pathogenicity [129,130]. This morphological transition is mainly determined by environmental cues and conditions, such as pH, temperature and nutrient availability [128,131]. For example, C. albicans and Penicillium marneffei are yeast-like at low pH, while at higher pH they transform into hypha-like [132,133]. Pseudozyma spp. also demonstrate dimorphism in response to different cues [43,44], and we also detected P. aphidis with a hypha-like structure at high pH, whereas it was yeast-like at low pH (unpublished data).
6. Competition for Space and Nutrients
Competition can occur between the BCA and the pathogen on plants for either space or nutrients; competition for both can hinder plant pathogens and control their ability to proliferate and infect plants. Similar to other BCAs, different Trichoderma strains use divergent modes of action to control phytopathogens, including competition for space and for micro- or macronutrients [118]. Low availability or limited sources of these nutrients challenge BCAs to develop different strategies to utilize them. One strategy might be competition for space, ultimately excluding the pathogen from the host plant. For example, T. harzianum is able to control B. cinerea on grapes by colonizing the blossom tissue to compete for space and eventually excluding the pathogen from its infection site [134]. Other strategies rely on the ability of BCAs to mobilize nutrients using low pH [123], specialized enzymes such as invertase [135], sugar transporters [136,137] or siderophores [138]. The ability to use diverse carbon or nitrogen sources from the plant and the pathogen can also contribute to the competitive ability. For example, Kloeckera apiculate competing with Penicillium italicum for nutrients and vitamins has a lower spectrum of carbon sources but a wide range of nitrogen sources as compared to the pathogen [139]. There are some examples of combined competition for both space and nutrients, such as the BCA Pichia guilliermondii which controls Rhizopus nigricans on tomato fruit during storage [140]. Aureobasidium pullulans also controls the decay of apple fruit caused by B. cinerea and Penicillium expansum by competing for both space and nutrients [141]. Competition for space and nutrients was also demonstrated after application of P. aphidis on tomato plants. P. aphidis accumulated and proliferated significantly more in the lesion area of B. cinerea than in the uninfected area, suggesting competition for space [43]. Furthermore, using excess sugar on tomato fruit reduces the competition for nutrients between the fungi and thus decreases the efficacy of P. aphidis against B. cinerea [62]. The ability to utilize more sources of carbon or nitrogen will give the BCA the advantage over the pathogen, and this will eventually reduce the infection.
7. Conclusions
Health and environmental concerns, along with the rapid development of pathogen resistance to widely used pesticides, call for enhancing the use of BCAs and biopesticides developed from their secretions in the long term, for the benefit of the consumer and the growers. Most biocontrol modes of action are mediated by their secretion-based fraction, which can include metabolites and proteins with different functions—from antibiotics through effectors—and enzymes that can affect or interact with the pathogen and/or the plant host. Much attention is being given to the characterization of such BCA-secreted molecules and proteins, but the development of robust molecular research tools is needed. Recent work shows that Pseudozyma can be modified using CRISPR-Cas9 system [142] for gene editing, and being yeast-like, it is comparatively easier to transform than filamentous fungi [44]. Large-scale studies, such as tritrophic interactions and omics [77,143], albeit challenging, will also help decipher pathways and reveal novel targets in pathogens and plants for directed control mechanisms. These tools will help further elucidate the molecular mechanism underlying the BCAs’ secretion components and will advance our ability to improve BCAs’ modes of action to suppress plant pathogens.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflict of interest.
References
- The State of Food Insecurity in the World 2009: Economic Crisis–Impact and Lesson Learned; Food and Agriculture Organization of the United Nations: Rome, Italy, 2009.
- Oerke, E.C.; Dehne, H.W. Safeguarding production—Losses in major crops and the role of crop protection. Crop Prot. 2004, 23, 275–285. [Google Scholar] [CrossRef]
- Bebber, D.P.; Gurr, S.J. Crop-destroying fungal and oomycete pathogens challenge food security. Fungal Genet. Biol. 2015, 74, 62–64. [Google Scholar] [CrossRef] [PubMed]
- Oerke, E.C. Crop losses to pests. J. Agric. Sci. 2006, 144, 31–43. [Google Scholar] [CrossRef]
- Lee, N.; D’Souza, C.A.; Kronstad, J.W. Of smuts, blasts, mildews, and blights: cAMP Signaling in Phytopathogenic Fungi. Annu. Rev. Phytopathol. 2003, 41, 399–427. [Google Scholar] [CrossRef] [PubMed]
- Agrios, G. (Ed.) Plant Pathology, 5th ed.; Elsevier Academic Press: Amsterdam, The Netherlands, 2005. [Google Scholar]
- Kennedy, B.W.; Alcorn, S.M. Estimates of U.S. Crop Losses to Procaryote Plant Pathogens. Plant Dis. 1980, 64, 674. [Google Scholar] [CrossRef]
- Vidhyasekaran, P. Bacterial Disease Resistance in Plants: Molecular Biology and Biotechnological Applications; CRC Press: Boca, Florida, USA, 2002; ISBN 9781560229254. [Google Scholar]
- Denholm, I.; Rowland, M.W. Tactics for Managing Pesticide Resistance in Arthropods: Theory and Practice. Annu. Rev. Entomol. 1992, 37, 91–112. [Google Scholar] [CrossRef] [PubMed]
- Leroux, P.; Fritz, R.; Debieu, D.; Albertini, C.; Lanen, C.; Bach, J.; Gredt, M.; Chapeland, F. Mechanisms of resistance to fungicides in field strains of Botrytis cinerea. Pest Manag. Sci. 2002, 58, 876–888. [Google Scholar] [CrossRef] [PubMed]
- Vela-Corcía, D.; Aditya Srivastava, D.; Dafa-Berger, A.; Rotem, N.; Barda, O.; Levy, M. MFS transporter from Botrytis cinerea provides tolerance to glucosinolate-breakdown products and is required for pathogenicity. Nat. Commun. 2019, 10, 2886. [Google Scholar] [CrossRef] [Green Version]
- Leroux, P.; Walker, A.S. Activity of fungicides and modulators of membrane drug transporters in field strains of Botrytis cinerea displaying multidrug resistance. Eur. J. Plant Pathol. 2013, 135, 683–693. [Google Scholar] [CrossRef]
- Arendrup, M.C.; Patterson, T.F. Multidrug-Resistant Candida: Epidemiology, Molecular Mechanisms, and Treatment. J. Infect. Dis. 2017, 216, S445–S451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rupp, S.; Weber, R.W.S.; Rieger, D.; Detzel, P.; Hahn, M. Spread of Botrytis cinerea strains with multiple fungicide resistance in German horticulture. Front. Microbiol. 2017, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leroch, M.; Plesken, C.; Weber, R.W.S.; Kauff, F.; Scalliet, G.; Hahn, M. Gray mold populations in German strawberry fields are resistant to multiple fungicides and dominated by a novel clade closely related to Botrytis cinerea. Appl. Environ. Microbiol. 2013, 79, 159–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mernke, D.; Dahm, S.; Walker, A.S.; Lalève, A.; Fillinger, S.; Leroch, M.; Hahn, M. Two promoter rearrangements in a drug efflux transporter gene are responsible for the appearance and spread of multidrug resistance phenotype MDR2 in Botrytis cinerea isolates in French and German vineyards. Phytopathology 2011, 101, 1176–1183. [Google Scholar] [CrossRef] [Green Version]
- Russ, D.; Kishony, R. Additivity of inhibitory effects in multidrug combinations. Nat. Microbiol. 2018, 3, 1339–1345. [Google Scholar] [CrossRef] [PubMed]
- Dayan, F.; Romagni, J.; Tellez, M.; Romando, A.; Duke, S. Managing weeds with natural products. Pestic. Outlook 1999, 10, 185–188. [Google Scholar]
- Dayan, F.E.; Cantrell, C.L.; Duke, S.O. Natural products in crop protection. Bioorganic Med. Chem. 2009, 17, 4022–4034. [Google Scholar] [CrossRef] [PubMed]
- Copping, L.G.; Duke, S.O. Natural products that have been used commercially as crop protection agents. Pest Manag. Sci. 2007, 63, 524–554. [Google Scholar] [CrossRef]
- Isman, M.B. Repellents in modern agriculture and an increasingly regulated world. Annu. Rev. Entomol. 2006, 51, 45–66. [Google Scholar] [CrossRef] [Green Version]
- Wilson, R.A.; Jenkinson, J.M.; Gibson, R.P.; Littlechild, J.A.; Wang, Z.Y.; Talbot, N.J. Tps1 regulates the pentose phosphate pathway, nitrogen metabolism and fungal virulence. EMBO J. 2007, 26, 3673–3685. [Google Scholar] [CrossRef] [Green Version]
- Elad, Y.; Freeman, S. Biological Control of Fungal Plant Pathogens. In Agricultural Applications; Springer: Berlin/Heidelberg, Germany, 2002; pp. 93–109. [Google Scholar]
- Butt, M.T.; Jackson, C.; Magan, N. (Eds.) Fungi as Biocontrol Agents: Progress, Problems and Potential, 1st ed.; CABI: Wallingford, UK, 2001. [Google Scholar]
- Raaijmakers, J.M.; Vlami, M.; de Souza, J.T. Antibiotic production by bacterial biocontrol agents. Antonie Van Leeuwenhoek 2002, 81, 537–547. [Google Scholar] [CrossRef]
- Kim, B.S.; Hwang, B.K. Microbial Fungicides in the Control of Plant Diseases. J. Phytopathol. 2007, 155, 641–653. [Google Scholar] [CrossRef]
- Porter, N. Physicochemical biophysical panel symposium biologically active secondary metabolites. Pestic. Sci. 1985, 16, 422–427. [Google Scholar]
- Früh, T.; Chemla, P.; Ehrler, J.; Farooq, S. Natural products as pesticides: Two examples of stereoselective synthesis. Pestic. Sci. 1996, 46, 37–47. [Google Scholar] [CrossRef]
- Zhang, X.; Harvey, P.R.; Stummer, B.E.; Warren, R.A.; Zhang, G.; Guo, K.; Li, J.; Yang, H. Antibiosis functions during interactions of Trichoderma afroharzianum and Trichoderma gamsii with plant pathogenic Rhizoctonia and Pythium. Funct. Integr. Genom. 2015, 15, 599–610. [Google Scholar] [CrossRef]
- Howell, C.R. Mechanisms employed by Trichoderma species in the biological control of plant diseases: The history and evolution of current concepts. Plant Dis. 2003, 87, 4–10. [Google Scholar] [CrossRef] [Green Version]
- Reino, J.L.; Guerrero, R.F.; Hernández-Galán, R.; Collado, I.G. Secondary metabolites from species of the biocontrol agent Trichoderma. Phytochem. Rev. 2008, 7, 89–123. [Google Scholar] [CrossRef]
- Sood, M.; Kapoor, D.; Kumar, V.; Sheteiwy, M.S.; Ramakrishnan, M.; Landi, M.; Araniti, F.; Sharma, A. Trichoderma: The “Secrets” of a Multitalented Biocontrol Agent. Plants 2020, 9, 762. [Google Scholar] [CrossRef]
- Krause, C.; Kirschbaum, J.; Jung, G.; Brückner, H. Sequence diversity of the peptaibol antibiotic suzukacillin-A from the mold Trichoderma viride. J. Pept. Sci. 2006, 12, 321–327. [Google Scholar] [CrossRef]
- Szekeres, A.; Leitgeb, B.; Kredics, L.; Antal, Z.; Hatvani, L.; Manczinger, L.; Vágvölgyi, C. Peptaibols and related peptaibiotics of Trichoderma: A review. Acta Microbiol. Immunol. Hung. 2005, 52, 137–168. [Google Scholar] [CrossRef]
- Shi, M.; Chen, L.; Wang, X.W.; Zhang, T.; Zhao, P.B.; Song, X.Y.; Sun, C.Y.; Chen, X.L.; Zhou, B.C.; Zhang, Y.Z. Antimicrobial peptaibols from Trichoderma pseudokoningii induce programmed cell death in plant fungal pathogens. Microbiology 2012, 158, 166–175. [Google Scholar] [CrossRef] [Green Version]
- Schirmbock, M.; Lorito, M.; Wang, Y.L.; Hayes, C.K.; Arisan-Atac, I.; Scala, F.; Harman, G.E.; Kubicek, C.P. Parallel formation and synergism of hydrolytic enzymes and peptaibol antibiotics, molecular mechanisms involved in the antagonistic action of Trichoderma harzianum against phytopathogenic fungi. Appl. Environ. Microbiol. 1994, 60, 4364–4370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hajlaoui, M.R.; Bélanger, R.R. Comparative effects of temperature and humidity on the activity of three potential antagonists of rose powdery mildew. Neth. J. Plant Pathol. 1991, 97, 203–208. [Google Scholar] [CrossRef]
- Hajlaoui, M.R.; Traquair, J.A.; Jarvis, W.R.; Belanger, B. Antifungal Activity of Extracellular Metabolites Produced by Sporothrix flocculosa. Biocontrol Sci. Technol. 1994, 4, 229–237. [Google Scholar] [CrossRef]
- Jarvis, W.R.; Shaw, L.A.; Traquair, J.A. Factors affecting antagonism of cucumber powdery mildew by Stephanoascus flocculosus and S. rugulosus. Mycol. Res. 1989, 92, 162–165. [Google Scholar] [CrossRef]
- Hajlaoui, M.R.; Belanger, R.R. Antagonism of the Yeast-like Phylloplane Fungus Sporothrix flocculosa against Erysiphe graminis var tritici. Biocontrol Sci. Technol. 1993, 3, 427–434. [Google Scholar] [CrossRef]
- Dik, A.J.; Verhaar, M.A.; Bélanger, R.R. Comparison of three biological control agents against cucumber powdery mildew (Sphaerotheca fuliginea) in semi-commercial-scale glasshouse trials. Eur. J. Plant Pathol. 1998, 104, 413–423. [Google Scholar] [CrossRef]
- Hammami, W.; Castro, C.Q.; Rémus-Borel, W.; Labbé, C.; Bélanger, R.R. Ecological basis of the interaction between Pseudozyma flocculosa and powdery mildew fungi. Appl. Environ. Microbiol. 2011, 77, 926–933. [Google Scholar] [CrossRef] [Green Version]
- Buxdorf, K.; Rahat, I.; Gafni, A.; Levy, M. The epiphytic fungus Pseudozyma aphidis induces jasmonic acid- and salicylic acid/nonexpressor of PR1-independent local and systemic resistance. Plant Physiol. 2013, 161, 2014–2022. [Google Scholar] [CrossRef] [Green Version]
- Gafni, A.; Calderon, C.E.; Harris, R.; Buxdorf, K.; Dafa-Berger, A.; Zeilinger-Reichert, E.; Levy, M. Biological control of the cucurbit powdery mildew pathogen Podosphaera xanthii by means of the epiphytic fungus Pseudozyma aphidis and parasitism as a mode of action. Front. Plant Sci. 2015, 6, 132. [Google Scholar] [CrossRef] [Green Version]
- Avis, T.J.; Boulanger, R.R.; Bélanger, R.R. Synthesis and biological characterization of (Z)-9-heptadecenoic and (Z)-6-methyl-9-heptadecenoic acids: Fatty acids with antibiotic activity produced by Pseudozyma flocculosa. J. Chem. Ecol. 2000, 26, 987–1000. [Google Scholar] [CrossRef]
- Benyagoub, M.; Bel Rhlid, R.; Bélanger, R.R. Purification and characterization of new fatty acids with antibiotic activity produced by Sporothrix flocculosa. J. Chem. Ecol. 1996, 22, 405–413. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, S.R.; Traquair, J.A.; Jarvis, W.R. 4-Methyl-7,11-heptadecadienal and 4-methyl-7,11-heptadecadienoic acid: New antibiotics from Sporothrix flocculosa and Sporothrix rugulosa. J. Nat. Prod. 1994, 57, 700–704. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, S.R.; Traquair, J.A.; Jarvis, W.R. New extracellular fatty acids in culture filtrates of Sporothrix flocculosa and S. rugulosa. Can. J. Chem. 1995, 73, 84–87. [Google Scholar] [CrossRef]
- Hajlaoui, M.R. Cytochemical Study of the Antagonistic Activity of Sporothrix flocculosa on Rose Powdery Mildew, Sphaerotheca pannosa var. rosae. Phytopathology 1992, 82, 583. [Google Scholar] [CrossRef]
- Avis, T.J.; Belanger, R.R. Specificity and mode of action of the antifungal fatty acid cis-9-heptadecenoic acid produced by Pseudozyma flocculosa. Appl. Environ. Microbiol. 2001, 67, 956–960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Golubev, W.I.; Pfeiffer, I.; Golubeva, E.W. Mycocin production in Pseudozyma tsukubaensis. Mycopathologia 2006, 162, 313–316. [Google Scholar] [CrossRef]
- Golubev, W.I. Mycocinogeny in smut yeast-like fungi of the genus Pseudozyma. Microbiology 2007, 76, 719–722. [Google Scholar] [CrossRef]
- Cheng, Y.; McNally, D.J.; Labbé, C.; Voyer, N.; Belzile, F.; Bélanger, R.R. Insertional mutagenesis of a fungal biocontrol agent led to discovery of a rare cellobiose lipid with antifungal activity. Appl. Environ. Microbiol. 2003, 69, 2595–2602. [Google Scholar] [CrossRef] [Green Version]
- Mimee, B.; Labbé, C.; Pelletier, R.; Bélanger, R.R. Antifungal activity of flocculosin, a novel glycolipid isolated from Pseudozyma flocculosa. Antimicrob. Agents Chemother. 2005, 49, 1597–1599. [Google Scholar] [CrossRef] [Green Version]
- Mimee, B.; Pelletier, R.; Bélanger, R.R. In vitro antibacterial activity and antifungal mode of action of flocculosin, a membrane-active cellobiose lipid. J. Appl. Microbiol. 2009, 107, 989–996. [Google Scholar] [CrossRef]
- Kulakovskaya, T.V.; Shashkov, A.S.; Kulakovskaya, E.V.; Golubev, W.I. Ustilagic acid secretion by Pseudozyma fusiformata strains. FEMS Yeast Res. 2005, 5, 919–923. [Google Scholar] [CrossRef] [Green Version]
- Golubev, W.I.; Kulakovskaya, T.V.; Shashkov, A.S.; Kulakovskaya, E.V.; Golubev, N. V Antifungal cellobiose lipid secreted by the epiphytic yeast Pseudozyma graminicola. Microbiology 2008, 77, 171–175. [Google Scholar] [CrossRef]
- Haskins, R.H.; Thorn, J.A. Biochemistry of the Ustilaginales: VII. Antibiotic activity of Ustilagic acid. Can. J. Bot. 1951, 29, 585–592. [Google Scholar] [CrossRef]
- Lefebvre, F.; Joly, D.L.; Labbé, C.; Teichmann, B.; Linning, R.; Belzile, F.; Bakkeren, G.; Bélanger, R.R. The transition from a phytopathogenic smut ancestor to an anamorphic biocontrol agent deciphered by comparative whole-genome analysis. Plant Cell 2013, 25, 1946–1959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bélanger, R.R.; Labbé, C.; Lefebvre, F.; Teichmann, B. Mode of action of biocontrol agents: All that glitters is not gold. Can. J. Plant Pathol. 2012, 34, 469–478. [Google Scholar] [CrossRef]
- Barda, O.; Shalev, O.; Alster, S.; Buxdorf, K.; Gafni, A.; Levy, M. Pseudozyma aphidis induces salicylic-acid-independent resistance to Clavibacter michiganensis in tomato plants. Plant Dis. 2015, 99, 621–626. [Google Scholar] [CrossRef]
- Calderón, C.E.; Rotem, N.; Harris, R.; Vela-Corcía, D.; Levy, M. Pseudozyma aphidis activates reactive oxygen species production, programmed cell death and morphological alterations in the necrotrophic fungus Botrytis cinerea. Mol. Plant Pathol. 2019, 20, 562–574. [Google Scholar] [CrossRef] [Green Version]
- Nyfeler, R.; Ackermann, P. Phenylpyrroles, a New Class of Agricultural Fungicides Related to the Natural Antibiotic Pyrrolnitrin. In Synthesis and Chemistry of Agrochemicals, 3rd ed.; ACS Publications: Washington, DC, USA, 1992; pp. 395–404. [Google Scholar]
- Anke, T.; Oberwinkler, F.; Steglich, W.; Schramm, G. The strobilurins—New antifungal antibiotics from the basidiomycete Strobilurus tenacellus. J. Antibiot. 1977, 30, 806–810. [Google Scholar] [CrossRef]
- Bartlett, D.W.; Clough, J.M.; Godwin, J.R.; Hall, A.A.; Hamer, M.; Parr-Dobrzanski, B. The strobilurin fungicides. Pest Manag. Sci. 2002, 58, 649–662. [Google Scholar] [CrossRef]
- Balba, H. Review of strobilurin fungicide chemicals. J. Environ. Sci. Health B 2007, 42, 441–451. [Google Scholar] [CrossRef]
- Takahashi, M.; Koyama, K.; Natori, S. Four New Azaphilones from Chaetomium globosum var. flavo-viridae. Chem. Pharm. Bull. 1990, 38, 625–628. [Google Scholar] [CrossRef] [Green Version]
- Park, J.H.; Gyung, J.C.; Kyoung, S.J.; He, K.L.; Heung, T.K.; Kwang, Y.C.; Kim, J.C. Antifungal activity against plant pathogenic fungi of chaetoviridins isolated from Chaetomium globosum. FEMS Microbiol. Lett. 2005, 252, 309–313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altomare, C.; Pengue, R.; Favilla, M.; Evidente, A.; Visconti, A. structure-activity relationships of derivatives of Fusapyrone, an antifungal metabolite of Fusarlum semitectum. J. Agric. Food Chem. 2004, 52, 2997–3001. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.C.; Choi, G.J.; Kim, H.J.; Kim, H.T.; Ahn, J.W.; Cho, K.Y. Verlamelin, an antifungal compound produced by a mycoparasite, Acremonium strictum. Plant Pathol. J. 2002, 18, 102–105. [Google Scholar] [CrossRef]
- Lee, D.W.; Kim, B.S. Antimicrobial cyclic peptides for plant disease control. Plant Pathol. J. 2015, 31, 1–11. [Google Scholar] [CrossRef]
- Kämper, J.; Kahmann, R.; Bölker, M.; Ma, L.J.; Brefort, T.; Saville, B.J.; Banuett, F.; Kronstad, J.W.; Gold, S.E.; Müller, O.; et al. Insights from the genome of the biotrophic fungal plant pathogen Ustilago maydis. Nature 2006, 444, 97–101. [Google Scholar] [CrossRef]
- Win, J.; Chaparro-Garcia, A.; Belhaj, K.; Saunders, D.G.O.; Yoshida, K.; Dong, S.; Schornack, S.; Zipfel, C.; Robatzek, S.; Hogenhout, S.A.; et al. Effector biology of plant-associated organisms: Concepts and perspectives. Cold Spring Harb. Symp. Quant. Biol. 2012, 77, 235–247. [Google Scholar] [CrossRef]
- Doehlemann, G.; van der Linde, K.; Assmann, D.; Schwammbach, D.; Hof, A.; Mohanty, A.; Jackson, D.; Kahmann, R. Pep1, a secreted effector protein of Ustilago maydis, is required for successful invasion of plant cells. PLoS Pathog. 2009, 5, e1000290. [Google Scholar] [CrossRef] [Green Version]
- Doehlemann, G.; Reissmann, S.; Aßmann, D.; Fleckenstein, M.; Kahmann, R. Two linked genes encoding a secreted effector and a membrane protein are essential for Ustilago maydis-induced tumour formation. Mol. Microbiol. 2011, 81, 751–766. [Google Scholar] [CrossRef]
- Sharma, R.; Ökmen, B.; Doehlemann, G.; Thines, M. Saprotrophic yeasts formerly classified as Pseudozyma have retained a large effector arsenal, including functional Pep1 orthologs. Mycol. Prog. 2019, 18, 763–768. [Google Scholar] [CrossRef]
- Laur, J.; Ramakrishnan, G.B.; Labbé, C.; Lefebvre, F.; Spanu, P.D.; Bélanger, R.R. Effectors involved in fungal-fungal interaction lead to a rare phenomenon of hyperbiotrophy in the tritrophic system biocontrol agent-powdery mildew-plant. New Phytol. 2018, 217, 713–725. [Google Scholar] [CrossRef] [Green Version]
- Guzmán-Guzmán, P.; Alemán-Duarte, M.I.; Delaye, L.; Herrera-Estrella, A.; Olmedo-Monfil, V. Identification of effector-like proteins in Trichoderma spp. and role of a hydrophobin in the plant-fungus interaction and mycoparasitism. BMC Genet. 2017, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramírez-Valdespino, C.A.; Casas-Flores, S.; Olmedo-Monfil, V. Trichoderma as a model to study effector-like molecules. Front. Microbiol. 2019, 10, 1030. [Google Scholar] [CrossRef] [PubMed]
- Djonović, S.; Pozo, M.J.; Dangott, L.J.; Howell, C.R.; Kenerley, C.M. Sm1, a proteinaceous elicitor secreted by the biocontrol fungus Trichoderma virens induces plant defense responses and systemic resistance. Mol. Plant Microbe Interact. 2006, 19, 838–853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seidl, V.; Marchetti, M.; Schandl, R.; Allmaier, G.; Kubicek, C.P. Epl1, the major secreted protein of Hypocrea atroviridis on glucose, is a member of a strongly conserved protein family comprising plant defense response elicitors. FEBS J. 2006, 273, 4346–4359. [Google Scholar] [CrossRef] [PubMed]
- Salas-Marina, M.A.; Isordia-Jasso, M.I.; Islas-Osuna, M.A.; Delgado-Sánchez, P.; Jiménez-Bremont, J.F.; Rodríguez-Kessler, M.; Rosales-Saavedra, M.T.; Herrera-Estrella, A.; Casas-Flores, S. The Epl1 and Sm1 proteins from Trichoderma atroviride and Trichoderma virens differentially modulate systemic disease resistance against different life style pathogens in Solanum lycopersicum. Front. Plant Sci. 2015, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crutcher, F.K.; Moran-Diez, M.E.; Ding, S.; Liu, J.; Horwitz, B.A.; Mukherjee, P.K.; Kenerley, C.M. A paralog of the proteinaceous elicitor SM1 is involved in colonization of maize roots by Trichoderma virens. Fungal Biol. 2015, 119, 476–486. [Google Scholar] [CrossRef] [PubMed]
- Gaderer, R.; Lamdan, N.L.; Frischmann, A.; Sulyok, M.; Krska, R.; Horwitz, B.A.; Seidl-Seiboth, V. Sm2, a paralog of the Trichoderma cerato-platanin elicitor Sm1, is also highly important for plant protection conferred by the fungal-root interaction of Trichoderma with maize. BMC Microbiol. 2015, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shcherbakova, L.A.; Odintsova, T.I.; Stakheev, A.A.; Fravel, D.R.; Zavriev, S.K. Identification of a novel small cysteine-rich protein in the fraction from the biocontrol Fusarium oxysporum strain CS-20 that mitigates fusarium wilt symptoms and triggers defense responses in tomato. Front. Plant Sci. 2016, 6. [Google Scholar] [CrossRef] [Green Version]
- Schuster, M.; Schweizer, G.; Kahmann, R. Comparative analyses of secreted proteins in plant pathogenic smut fungi and related basidiomycetes. Fungal Genet. Biol. 2018, 112, 21–30. [Google Scholar] [CrossRef]
- Hemetsberger, C.; Mueller, A.N.; Matei, A.; Herrberger, C.; Hensel, G.; Kumlehn, J.; Mishra, B.; Sharma, R.; Thines, M.; Hückelhoven, R.; et al. The fungal core effector Pep1 is conserved across smuts of dicots and monocots. New Phytol. 2015, 206, 1116–1126. [Google Scholar] [CrossRef] [PubMed]
- Malinovsky, F.G.; Fangel, J.U.; Willats, W.G.T. The role of the cell wall in plant immunity. Front. Plant Sci. 2014, 5, 178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saijo, Y.; Loo, E.P.I.; Yasuda, S. Pattern recognition receptors and signaling in plant–microbe interactions. Plant J. 2018, 93, 592–613. [Google Scholar] [CrossRef] [PubMed]
- Grant, M.; Lamb, C. Systemic immunity. Curr. Opin. Plant Biol. 2006, 9, 414–420. [Google Scholar] [CrossRef] [PubMed]
- Glazebrook, J. Contrasting Mechanisms of Defense against Biotrophic and Necrotrophic Pathogens. Annu. Rev. Phytopathol. 2005, 43, 205–227. [Google Scholar] [CrossRef] [PubMed]
- Somssich, I.E. Closing another gap in the plant SAR puzzle. Cell 2003, 113, 815–816. [Google Scholar] [CrossRef] [Green Version]
- Haas, D.; Défago, G. Biological control of soil-borne pathogens by fluorescent pseudomonads. Nat. Rev. Microbiol. 2005, 3, 307–319. [Google Scholar] [CrossRef]
- Heil, M. Ecological costs of induced resistance. Curr. Opin. Plant Biol. 2002, 5, 345–350. [Google Scholar] [CrossRef]
- Llorens, E.; García-Agustín, P.; Lapeña, L. Advances in induced resistance by natural compounds: Towards new options for woody crop protection. Sci. Agric. 2017, 74, 90–100. [Google Scholar] [CrossRef]
- Pieterse, C.M.J.; Van Loon, L.C. Salicylic acid-independent plant defence pathways. Trends Plant Sci. 1999, 4, 52–58. [Google Scholar] [CrossRef] [Green Version]
- Penninckx, I.A.M.A.; Thomma, B.P.H.J.; Buchala, A.; Métraux, J.P.; Broekaert, W.F. Concomitant activation of jasmonate and ethylene response pathways is required for induction of a plant defensin gene in Arabidopsis. Plant Cell 1998, 10, 2103–2113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomma, B.P.H.J.; Eggermont, K.; Penninckx, I.A.M.A.; Mauch-Mani, B.; Vogelsang, R.; Cammue, B.P.A.; Broekaert, W.F. Separate jasmonate-dependent and salicylate-dependent defense-response pathways in Arabidopsis are essential for resistance to distinct microbial pathogens. Proc. Natl. Acad. Sci. USA 1998, 95, 15107–15111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Penninckx, I.A.; Eggermont, K.; Terras, F.R.; Thomma, B.P.; De Samblanx, G.W.; Buchala, A.; Métraux, J.P.; Manners, J.M.; Broekaert, W.F. Pathogen-induced systemic activation of a plant defensin gene in Arabidopsis follows a salicylic acid-independent pathway. Plant Cell 1996, 8, 2309–2323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pieterse, C.M.J.; Zamioudis, C.; Berendsen, R.L.; Weller, D.M.; Van Wees, S.C.M.; Bakker, P.A.H.M. Induced systemic resistance by beneficial microbes. Annu. Rev. Phytopathol. 2014, 52, 347–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buxdorf, K.; Rahat, I.; Levy, M. Pseudozyma aphidis induces ethylene-independent resistance in plants. Plant Signal. Behav. 2013, 8, e26273. [Google Scholar] [CrossRef]
- Agrawal, A.A. Induced responses to herbivory and increased plant performance. Science 1998, 279, 1201–1202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baldwin, I.T. Jasmonate-induced responses are costly but benefit plants under attack in native populations. Proc. Natl. Acad. Sci. USA 1998, 95, 8113–8118. [Google Scholar] [CrossRef] [Green Version]
- Conrath, U.; Beckers, G.J.M.; Flors, V.; García-Agustín, P.; Jakab, G.; Mauch, F.; Newman, M.A.; Pieterse, C.M.J.; Poinssot, B.; Pozo, M.J.; et al. Priming: Getting ready for battle. Mol. Plant Microbe Interact. 2006, 19, 1062–1071. [Google Scholar] [CrossRef] [Green Version]
- Perazzolli, M.; Roatti, B.; Bozza, E.; Pertot, I. Trichoderma harzianum T39 induces resistance against downy mildew by priming for defense without costs for grapevine. Biol. Control 2011, 58, 74–82. [Google Scholar] [CrossRef]
- Yoshioka, Y.; Ichikawa, H.; Naznin, H.A.; Kogure, A.; Hyakumachi, M. Systemic resistance induced in Arabidopsis thaliana by Trichoderma asperellum SKT-1, a microbial pesticide of seedborne diseases of rice. Pest Manag. Sci. 2012, 68, 60–66. [Google Scholar] [CrossRef]
- Shoresh, M.; Yedidia, I.; Chet, I. Involvement of jasmonic acid/ethylene signaling pathway in the systemic resistance induced in cucumber by Trichoderma asperellum T203. Phytopathology 2005, 95, 76–84. [Google Scholar] [CrossRef] [Green Version]
- Molitor, A.; Zajic, D.; Voll, L.M.; Pons-Kühnemann, J.; Samans, B.; Kogel, K.H.; Waller, F. Barley leaf transcriptome and metabolite analysis reveals new aspects of compatibility and Piriformospora indica-mediated systemic induced resistance to powdery mildew. Mol. Plant Microbe Interact. 2011, 24, 1427–1439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aime, S.; Alabouvette, C.; Steinberg, C.; Olivain, C. The endophytic strain Fusarium oxysporum Fo47: A good candidate for priming the defense responses in tomato roots. Mol. Plant Microbe Interact. 2013, 26, 918–926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, G.; Lee, S.-H.; Kim, K.M.; Ryu, C.-M. Foliar application of the leaf-colonizing yeast Pseudozyma churashimaensis elicits systemic defense of pepper against bacterial and viral pathogens. Sci. Rep. 2017, 7, 39432. [Google Scholar] [CrossRef] [PubMed]
- Oostendorp, M.; Kunz, W.; Dietrich, B.; Staub, T. Induced disease resistance in plants by chemicals. Eur. J. Plant Pathol. 2001, 107, 19–28. [Google Scholar] [CrossRef]
- Friedrich, L.; Lawton, K.; Ruess, W.; Masner, P.; Specker, N.; Rella, M.G.; Meier, B.; Dincher, S.; Staub, T.; Uknes, S.; et al. A benzothiadiazole derivative induces systemic acquired resistance in tobacco. Plant J. 1996, 10, 61–70. [Google Scholar] [CrossRef]
- Walters, D.R.; Ratsep, J.; Havis, N.D. Controlling crop diseases using induced resistance: Challenges for the future. J. Exp. Bot. 2013, 64, 1263–1280. [Google Scholar] [CrossRef]
- Ton, J.; Jakab, G.; Toquin, V.; Flors, V.; Iavicoli, A.; Maeder, M.N.; Métraux, J.P.; Mauch-Mani, B. Dissecting the β-aminobutyric acid-induced priming phenomenon in Arabidopsis. Plant Cell 2005, 17, 987–999. [Google Scholar] [CrossRef] [Green Version]
- Aranega-Bou, P.; de la O Leyva, M.; Finiti, I.; Garcfa-Agustfn, P.; Gonzalez-Bosch, C. Priming of plant resistance by natural compounds. Hexanoic acid as a model. Front. Plant Sci. 2014, 5, 488. [Google Scholar] [CrossRef] [Green Version]
- Iriti, M.; Castorina, G.; Vitalini, S.; Mignani, I.; Soave, C.; Fico, G.; Faoro, F. Chitosan-induced ethylene-independent resistance does not reduce crop yield in bean. Biol. Control 2010, 54, 241–247. [Google Scholar] [CrossRef]
- Ahn, I.P.; Kim, S.; Lee, Y.H.; Suh, S.C. Vitamin B1-induced priming is dependent on hydrogen peroxide and the NPR1 gene in arabidopsis. Plant Physiol. 2007, 143, 838–848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noutoshi, Y.; Okazaki, M.; Kida, T.; Nishina, Y.; Morishita, Y.; Ogawa, T.; Suzuki, H.; Shibata, D.; Jikumaru, Y.; Hanada, A.; et al. Novel plant immune-priming compounds identified via high-throughput chemical screening target salicylic acid glucosyltransferases in Arabidopsis. Plant Cell 2012, 24, 3795–3804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soufleros, E.H.; Mygdalia, S.A.; Natskoulis, P. Production process and characterization of the traditional Greek fruit distillate “Koumaro” by aromatic and mineral composition. J. Food Compos. Anal. 2005, 18, 699–716. [Google Scholar] [CrossRef]
- Hossain, M.M.; Sultana, F.; Kubota, M.; Koyama, H.; Hyakumachi, M. The Plant Growth-Promoting Fungus Penicillium simplicissimum GP17-2 Induces Resistance in Arabidopsis thaliana by Activation of Multiple Defense Signals. Plant Cell Physiol. 2007, 48, 1724–1736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naznin, H.A.; Kiyohara, D.; Kimura, M.; Miyazawa, M.; Shimizu, M.; Hyakumachi, M. Systemic resistance induced by volatile organic compounds emitted by plant growth-promoting fungi in Arabidopsis thaliana. PLoS ONE 2014, 9, e86882. [Google Scholar] [CrossRef] [Green Version]
- Viterbo, A.; Wiest, A.; Brotman, Y.; Chet, I.; Kenerley, C. The 18mer peptaibols from Trichoderma virens elicit plant defence responses. Mol. Plant Pathol. 2007, 8, 737–746. [Google Scholar] [CrossRef]
- Vinale, F.; Sivasithamparam, K.; Ghisalberti, E.L.; Marra, R.; Barbetti, M.J.; Li, H.; Woo, S.L.; Lorito, M. A novel role for Trichoderma secondary metabolites in the interactions with plants. Physiol. Mol. Plant Pathol. 2008, 72, 80–86. [Google Scholar] [CrossRef]
- Weindling R Trichoderma lignorum as a parasite of other soil fungi. Phytopathology 1932, 22, 837–845.
- Lorito, M.; Harman, G.; Hayes, C.; Broadway, R.; Tronsmo, A.; Woo, S.; Di Pietro, A. Chitinolytic enzymes produced by Trichoderma harzianum: Antifungal activity of purified Endochitinase and Chitobiosidase. Phytopathology 1993, 83, 302. [Google Scholar] [CrossRef]
- Viterbo, A.; Haran, S.; Friesem, D.; Ramot, O.; Chet, I. Antifungal activity of a novel endochitinase gene (chit36) from Trichoderma harzianum Rifai TM. FEMS Microbiol. Lett. 2001, 200, 169–174. [Google Scholar] [CrossRef]
- Elad, Y. Biological control of foliar pathogens by means of Trichoderma harzianum and potential modes of action. Crop Prot. 2000, 19, 709–714. [Google Scholar] [CrossRef]
- Boyce, K.J.; Andrianopoulos, A. Fungal dimorphism: The switch from hyphae to yeast is a specialized morphogenetic adaptation allowing colonization of a host. FEMS Microbiol. Rev. 2015, 39, 797–811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murad, A.M.A.; D’Enfert, C.; Gaillardin, C.; Tournu, H.; Tekaia, F.; Talibi, D.; Marechal, D.; Marchais, V.; Cottin, J.; Brown, A.J.P. Transcript profiling in Candida albicans reveals new cellular functions for the transcriptional repressors CaTup1, CaMig1 and CaNrg1. Mol. Microbiol. 2001, 42, 981–993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lo, H.J.; Köhler, J.R.; Didomenico, B.; Loebenberg, D.; Cacciapuoti, A.; Fink, G.R. Nonfilamentous C. albicans mutants are avirulent. Cell 1997, 90, 939–949. [Google Scholar] [CrossRef] [Green Version]
- Berrocal, A.; Oviedo, C.; Nickerson, K.W.; Navarrete, J. Quorum sensing activity and control of yeast-mycelium dimorphism in Ophiostoma floccosum. Biotechnol. Lett. 2014, 36, 1503–1513. [Google Scholar] [CrossRef]
- Jacobsen, I.D.; Wilson, D.; Wächtler, B.; Brunke, S.; Naglik, J.R.; Hube, B. Candida albicans dimorphism as a therapeutic target. Expert Rev. Anti. Infect. Ther. 2012, 10, 85–93. [Google Scholar] [CrossRef] [Green Version]
- Vanittanakom, N.; Cooper, C.R.; Fisher, M.C.; Sirisanthana, T. Penicillium marneffei infection and recent advances in the epidemiology and molecular biology aspects. Clin. Microbiol. Rev. 2006, 19, 95–110. [Google Scholar] [CrossRef] [Green Version]
- Gullino, M.L. Control of Botrytis Rot of Grapes and Vegetables with Trichoderma Spp. In Biological Control of Plant Diseases; Springer: New York, NY, USA, 1992; pp. 125–132. [Google Scholar]
- Vargas, W.A.; Mandawe, J.C.; Kenerley, C.M. Plant-derived sucrose is a key element in the symbiotic association between Trichoderma virens and maize plants. Plant Physiol. 2009, 151, 792–808. [Google Scholar] [CrossRef] [Green Version]
- Delgado-Jarana, J.; Moreno-Mateos, M.Á.; Benítez, T. Glucose uptake in Trichoderma harzianum: Role of gtt1. Eukaryot. Cell 2003, 2, 708–717. [Google Scholar] [CrossRef] [Green Version]
- Benítez, T.; Rincón, A.M.; Limón, M.C.; Codón, A.C. Biocontrol mechanisms of Trichoderma strains. Int. Microbiol. 2004, 7, 249–260. [Google Scholar]
- Ghosh, S.K.; Banerjee, S.; Sengupta, C. Siderophore production by antagonistic fungi. J. Biopest. 2017, 10, 105–112. [Google Scholar]
- Liu, P.; Luo, L.; Long, C. An Characterization of competition for nutrients in the biocontrol of Penicillium italicum by Kloeckera apiculata. Biol. Control 2013, 67, 157–162. [Google Scholar] [CrossRef]
- Zhao, Y.; Tu, K.; Shao, X.; Jing, W.; Su, Z. Effects of the yeast Pichia guilliermondii against Rhizopus nigricans on tomato fruit. Postharvest Biol. Technol. 2008, 49, 113–120. [Google Scholar] [CrossRef]
- Ippolito, A.; El Ghaouth, A.; Wilson, C.L.; Wisniewski, M. Control of postharvest decay of apple fruit by Aureobasidium pullulans and induction of defense responses. Postharvest Biol. Technol. 2000, 19, 265–272. [Google Scholar] [CrossRef]
- Kunitake, E.; Tanaka, T.; Ueda, H.; Endo, A.; Yarimizu, T.; Katoh, E.; Kitamoto, H. CRISPR/Cas9-mediated gene replacement in the basidiomycetous yeast Pseudozyma antarctica. Fungal Genet. Biol. 2019, 130, 82–90. [Google Scholar] [CrossRef]
- Massart, S.; Perazzolli, M.; Höfte, M.; Pertot, I.; Jijakli, M.H. Impact of the omic technologies for understanding the modes of action of biological control agents against plant pathogens. BioControl 2015, 60, 725–746. [Google Scholar] [CrossRef]
- Wang, Q.-M.; Begerow, D.; Groenewald, M.; Liu, X.-Z.; Theelen, B.; Bai, F.-Y.; Boekhout, T. Multigene phylogeny and taxonomic revision of yeasts and related fungi in the Ustilaginomycotina. Stud. Mycol. 2015, 81, 55–83. [Google Scholar] [CrossRef] [Green Version]
- Piątek, M.; Lutz, M.; Yorou, N.S. A molecular phylogenetic framework for Anthracocystis (Ustilaginales), including five new combinations (inter alia for the asexual Pseudozyma flocculosa), and description of Anthracocystis grodzinskae sp. nov. Mycol. Prog. 2015, 14, 1–15. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Biocontrol modes of action. Illustration of biocontrol secretion-based direct and indirect modes of action against phytopathogens with a focus on Pseudozyma aphidis. ROS, reactive oxygen species; CWDE, cell wall-degrading enzymes.
Figure 1.
Biocontrol modes of action. Illustration of biocontrol secretion-based direct and indirect modes of action against phytopathogens with a focus on Pseudozyma aphidis. ROS, reactive oxygen species; CWDE, cell wall-degrading enzymes.

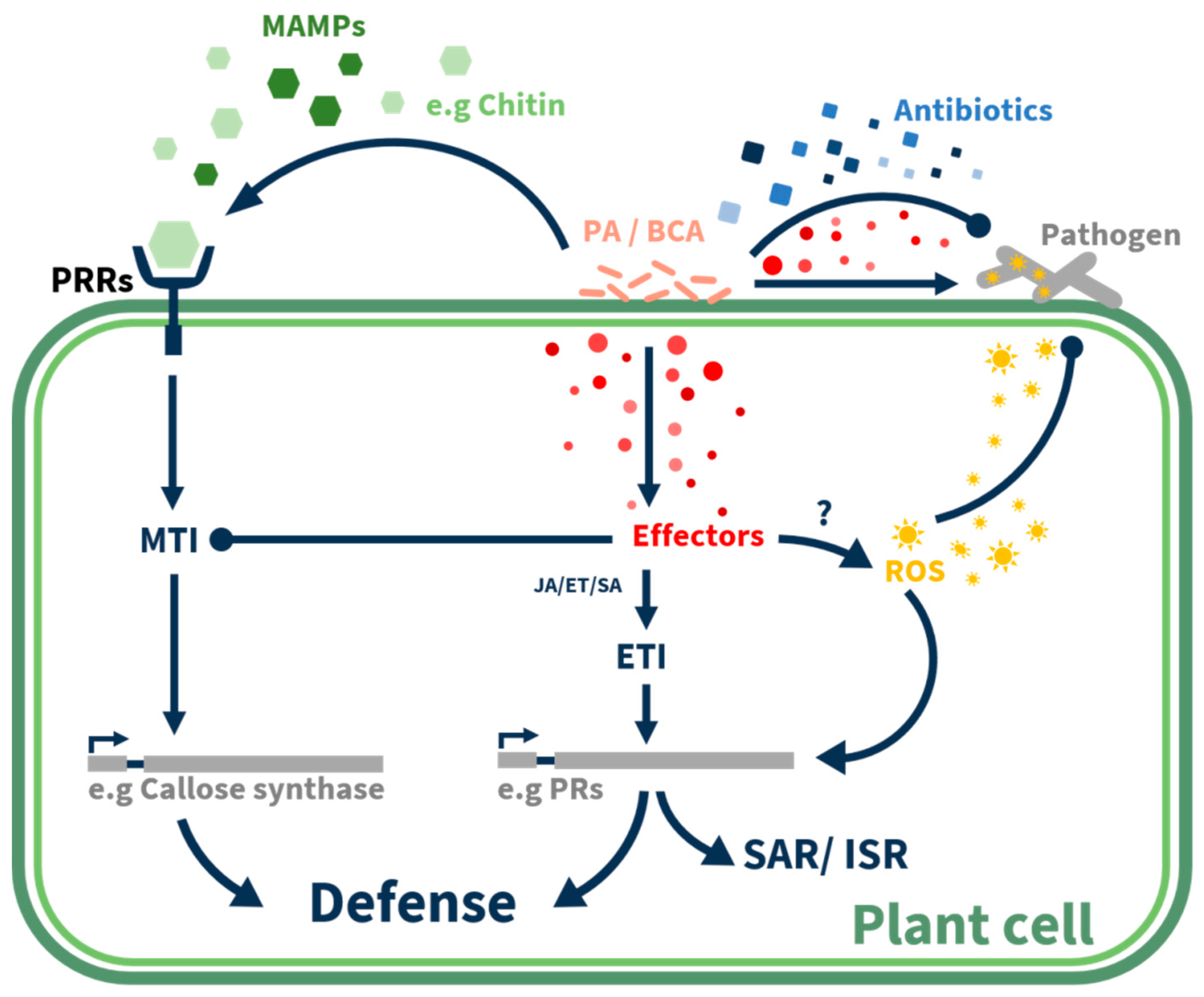
Figure 2.
Biocontrol agents’ involvement in plant defense pathways with a focus on Pseudozyma aphidis. Recognition of biocontrol agents by microbe associated molecular patterns (MAMPs) and suppression of MAMP-triggered immunity (MTI) by biocontrol agents, while activation the effector-triggered immunity (ETI) and expression of pathogenesis relate genes (PRs), and later on activation of the systemic acquired resistance (SAR) and induced-systemic resistance (ISR) in distal tissue.
Figure 2.
Biocontrol agents’ involvement in plant defense pathways with a focus on Pseudozyma aphidis. Recognition of biocontrol agents by microbe associated molecular patterns (MAMPs) and suppression of MAMP-triggered immunity (MTI) by biocontrol agents, while activation the effector-triggered immunity (ETI) and expression of pathogenesis relate genes (PRs), and later on activation of the systemic acquired resistance (SAR) and induced-systemic resistance (ISR) in distal tissue.


{kind=link}
{kind=link}
Table 1.
Overview of biological control agents (BCAs) modes of action mechanisms.
| Biocontrol Agent | Compound/Protein/Gene | Mechanism/Activity | Pathogen | Ref. |
|---|---|---|---|---|
| Antibiosis/ ROS/ PCD | ||||
| Acremonium strictum | Verlamelin | Unknown | Erysiphe graminis f. sp. hordei Puccnia recondita Botrytis cinerea | [5,70] |
| Trichoderma virens Trichoderma spp. | Peptaibols | Formation of pores in bilayer lipid membrane. | B. cinerea Mucor mucedo | [3,6,31,33,122] |
| Trichoderma pseudokoningii | Peptaibols | Induces metacaspase-independent apoptotic cell death. | Fusarium oxysporum | [35] |
| T. virens | Peptaibols | Induced resistance. | Pseudomonas syringae | [3,12,122] |
| Pseudozyma aphidis | Mostly lipophilic compounds. | ROS/PCD | Podosphaera xanthii B. cinerea Clavibacter michiganensis | [44,61,62,101] |
| Biocontrol Agent | Compound/Protein/Gene | Mechanism/Activity | Pathogen | Ref. |
| Psudozyma rugulosa Pseudozyma flocculosa (syn.Sporothrix flocculosa) | (Z)-9-heptadecenoic Z)-6-methyl-9-hepta decenoic acids Cis-9-Heptadecenoic acid (CHDA) | Disturbance in fluidity of the cell membrane, leakage of electrolytes and proteins. | E. graminis var. tritici, powdery mildews | [45,46,47,48,49,50] |
| P. flocculosa | Flocculosin | Leakage of cell membrane. | Candida albicans, Trichosporon asahii, powdery mildews | [53,54,55] |
| Pseudozyma tsukubaensis Pseudozyma prolifica | Mycocins | Membrane disruption. | Ustilaginomycetes | [51,52] |
| Pseudozyma graminicola Pseudozyma fusiformata | Ustilagic acid | Disruption of the cytoplasmic membrane permeability. | ~300 tested species of yeastlike and mycelial fungi | [56,57] |
| Chaetomium globosum | Chaetoviridins A and B | Antibiosis | Puccinia recondita, Magnaporthe grisea | [67,68] |
| Fusarium semitectum | Fusapyrone and deoxyfusapyrone | Antibiosis | B.cinerea, Aspergillus parasiticus, and Penicillium brevi | [69] |
| Gliocladium virens p | Gliovirin and heptelidic acid | Suppressing TNF-alpha synthesis. | Pythium ultimum | [30] |
| G. irens q | Gliotoxin and dimethylgliotoxin | Oxidative stress | Rhizoctonia solani | [30] |
| Effectors | ||||
| P. flocculosa | pf02826, pf00303 and pf02382 | Dissemination and sequestration of nutrient. | Blumeria graminis | [77] |
| Pseudozyma sp. | Pep1, Cmu1, Cwh41 and Hum3 | BCA -Plant interactions | [76,87] | |
| T. virens | tvlysm1 | Colonization of BCA and defense against the pathogen. | Rhizoctonia solani | [78] |
| T. virens | Cerato-platanin protein Sm1 | Induces ROS and PR genes expression in cotton. | Cochliobolus heterostrophus | [80] |
| T. virens | Sm2 and Epl2 | Induced resistance | C. heterostrophus | [80] |
| Fusarium oxysporum strain CS-20 | CS20EP | Elicits defense responses and ion exchange in tomato plants. | F. oxysporum | [85] |
| Hydrolytic Enzymes | ||||
| Trichoderma harzianum | Endo-chitinase, chitobiosidase | B.cinerea; F. oxysporum Sclerotium rolfsii | [125,126,127] | |
| P. aphidis | Chitinase, protease lipase, cellulase | P. xanthii, B. cinerea | [44,62] | |
| Parasitism | ||||
| Trichoderma lignorum | Mycoparasitism, haustoria formation. | R. solani | [30,124] | |
| P. flocculosa | Mycoparasitism, Hyperbiotrophy. | P. xanthii B. graminis | [55,77] | |
| P. aphidis | Ecto-parasitism | P. xanthii | [44] | |
| Biocontrol Agent | Compound/Protein/Gene | Mechanism/Activity | Pathogen | Ref. |
| Induced resistance (IR) | ||||
| T. harzianum T39 | Through jasmonic acid(JA) and ethylene(ET) signals. | Induced resistance in grapevine. | Plasmopara viticola | [105] |
| Trichoderma asperellum SKT-1 | Through salicylic acid (SA), JA and ET signaling pathways. | Induced resistance in Arabidopsis thaliana. | P. syringae | [106] |
| T. asperellum T203 | Through JA and ET signals. | Induced resistance in cucumber. | P. syringae | [107] |
| T. virens | Sm1 and 18 mer peptaibols. | Induced resistance in cucumber. | P. syringae | [80,122] |
| Piriformospora indica | ISR, upregulation of PR genes and heat-shock proteins. | Induced resistance in barley. | B. graminis | [108] |
| F. oxysporum Fo47 | Root colonization upregulation of GLUA and PR-1a. | Induced resistance in tomato. | Fusarium wilt | [109] |
| Penicillium simplicissimum GP17-2 | Through SA, JA and ET signaling pathways. | Induced resistance in A. thaliana. | P. syringae | [111] |
| Ampelomyces sp. and Cladosporium sp | Volatiles; m-cresol and methyl benzoate induce ISR. | Induced resistance in A. thaliana. | P.syringae pv. tomato DC3000 | [121] |
| P. aphidis | Induce ISR and SAR through JA/ET and SA pathways. | Induce resistance in A. thaliana, tomato and cucumber. | B. cinerea C. michiganensis | [43,61,101] |
| Competition | ||||
| T. harzianum | Competition for space. | B. cinerea | [134] | |
| P. aphidis | Competition for space and nutrients. | P. xanthii, B. cinerea | [43,44,62] | |
| Pichia guilliermondii | Competition for space and nutrients. | Rhizopus nigricans | [140] | |
| Kloeckera apiculate | Competition for nutrients | Penicillium italicum | [139] | |
Programmed cell Death, PCD; Reactive Oxygen Species, ROS; Tumor Necrosis Factor, TNF; Induced Systemic Resistance, ISR; Systemic Acquired Resistance, SAR.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Srivastava, D.A.; Harris, R.; Breuer, G.; Levy, M. Secretion-Based Modes of Action of Biocontrol Agents with a Focus on Pseudozyma aphidis. Plants 2021, 10, 210. https://doi.org/10.3390/plants10020210
AMA Style
Srivastava DA, Harris R, Breuer G, Levy M. Secretion-Based Modes of Action of Biocontrol Agents with a Focus on Pseudozyma aphidis. Plants. 2021; 10(2):210. https://doi.org/10.3390/plants10020210
Chicago/Turabian StyleSrivastava, Dhruv Aditya, Raviv Harris, Gilli Breuer, and Maggie Levy. 2021. "Secretion-Based Modes of Action of Biocontrol Agents with a Focus on Pseudozyma aphidis" Plants 10, no. 2: 210. https://doi.org/10.3390/plants10020210
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.