
Use of Network Analysis and Spread Models to Target Control Actions for Bovine Tuberculosis in a State from Brazil

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Datasets Description
2.2. Characterization of the Study Population and Movement Network
2.3. Network Analysis
2.3.1. Static Network Description
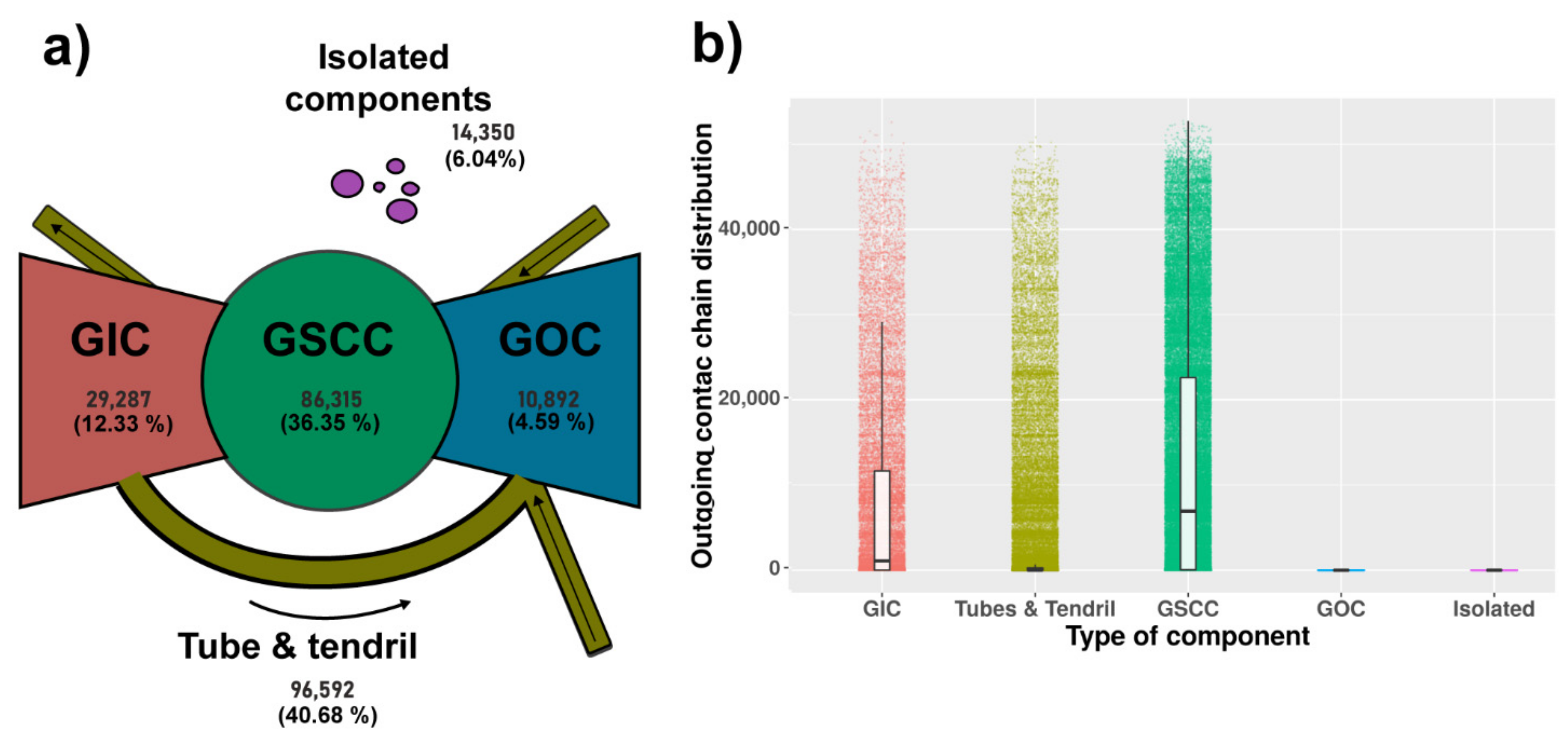
Components Description
2.3.2. Time Series Network Analysis
2.4. Relationship between the Observed Network and the Distribution of TB-Like Lesion Cases
2.5. Disease Spread Model
2.5.1. Parameters of the Model
2.5.2. Disease Spread and Targeted Control Action Modeling
2.5.3. Sensitivity Analyses
2.5.4. Software
3. Results
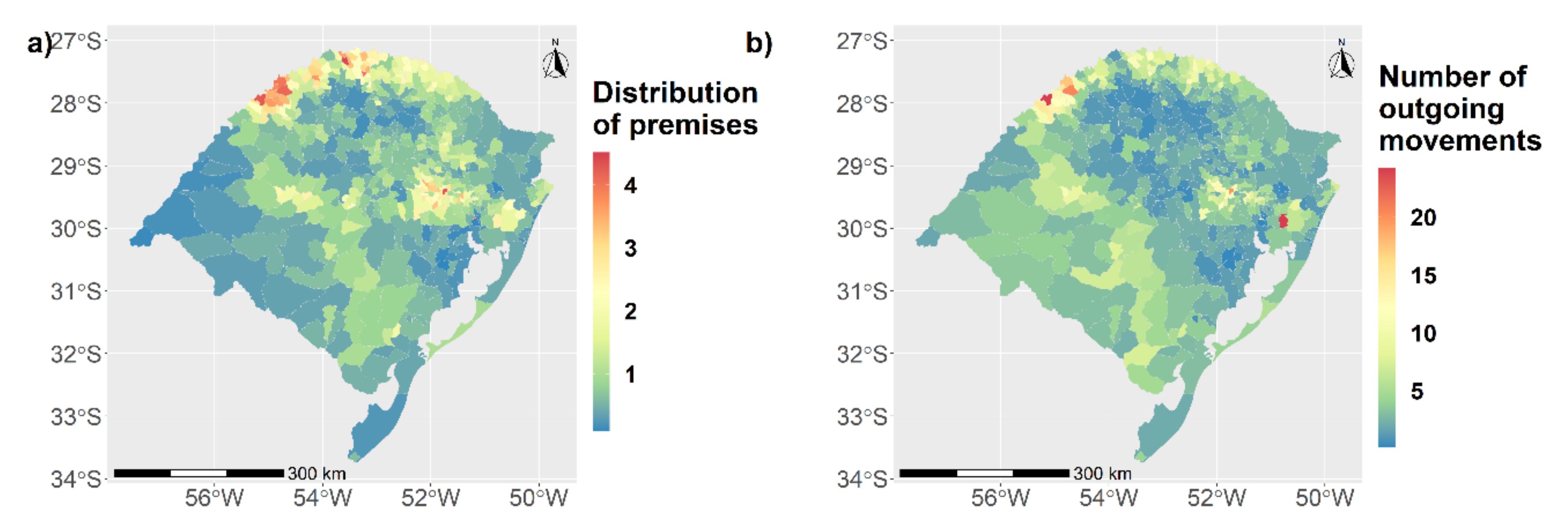
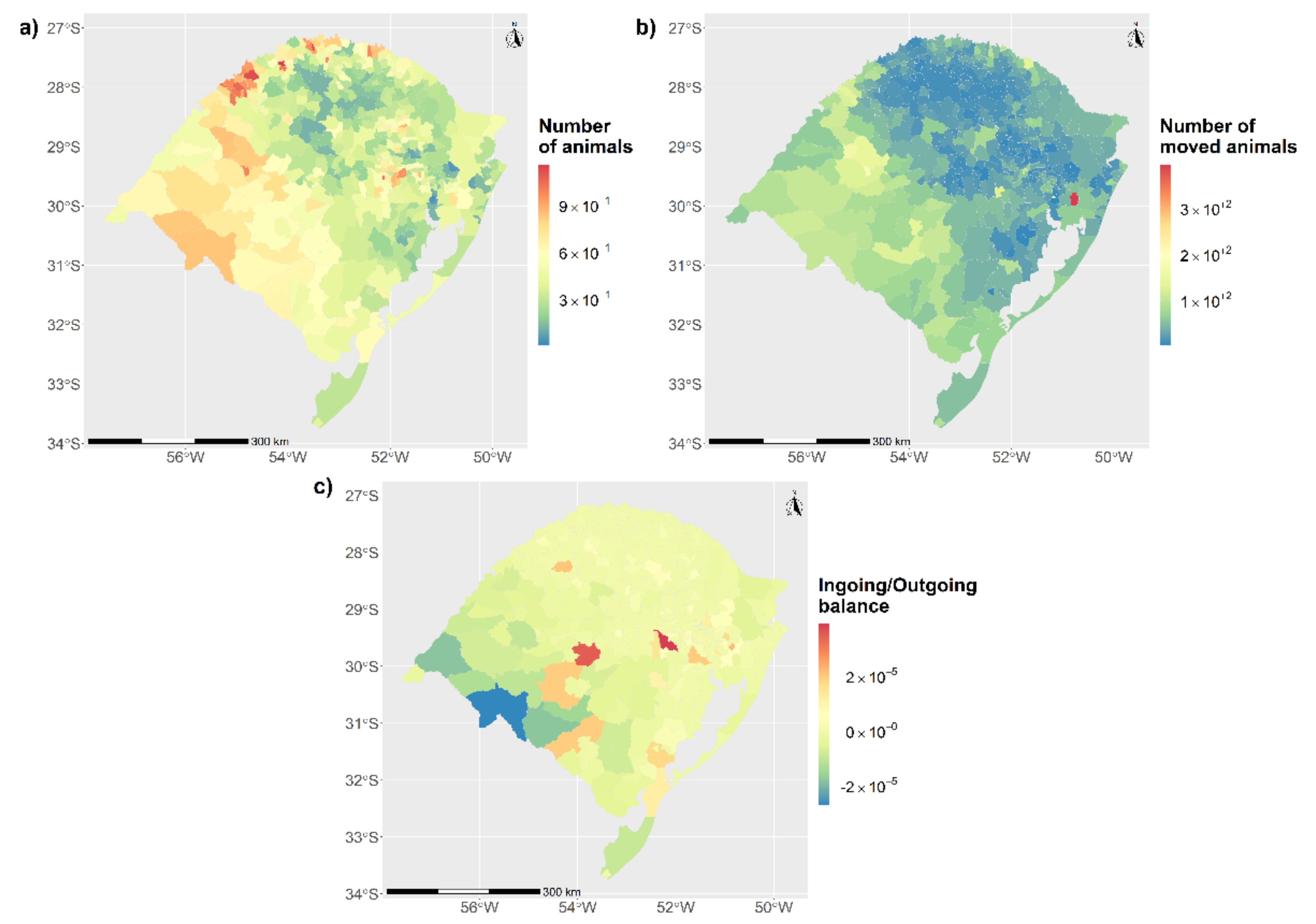
3.1. Characterization of the Study Population and Movement Network
3.1.1. Static Network Description
3.1.2. Components Analysis
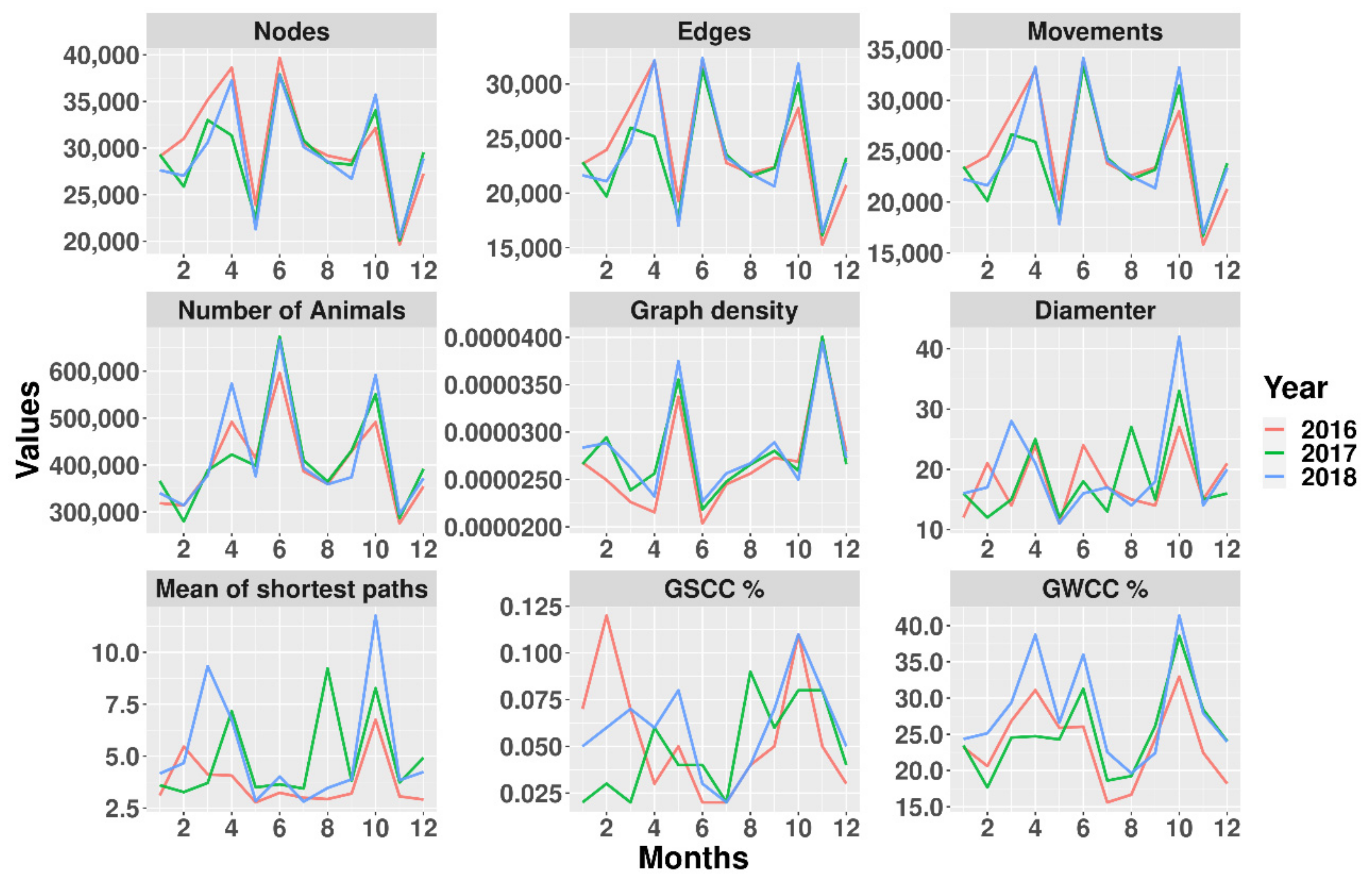
3.2. Time Series Network Description
3.2.1. Relation between the Observed Network and the Distribution of Cases across the Network
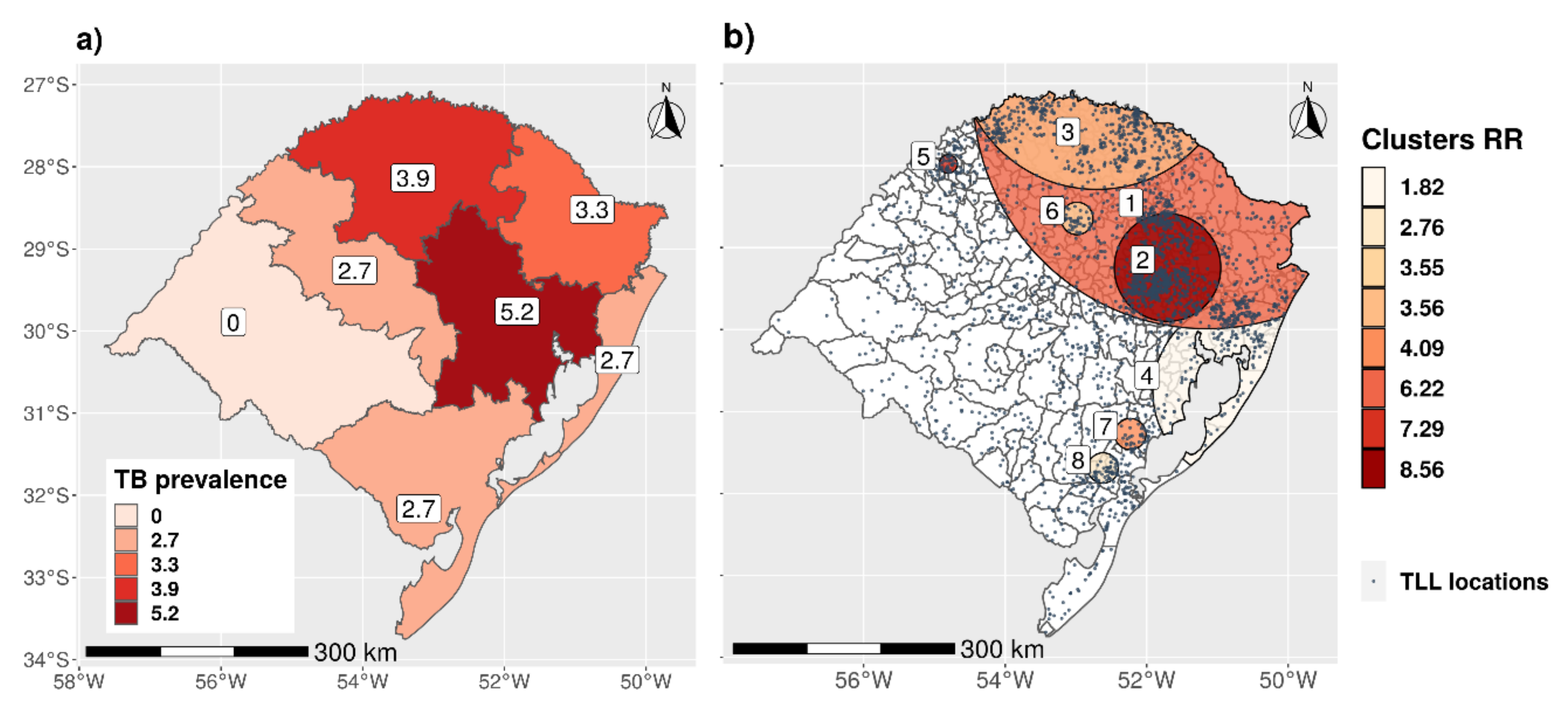
3.2.2. Spatial Distribution and Cluster Analysis
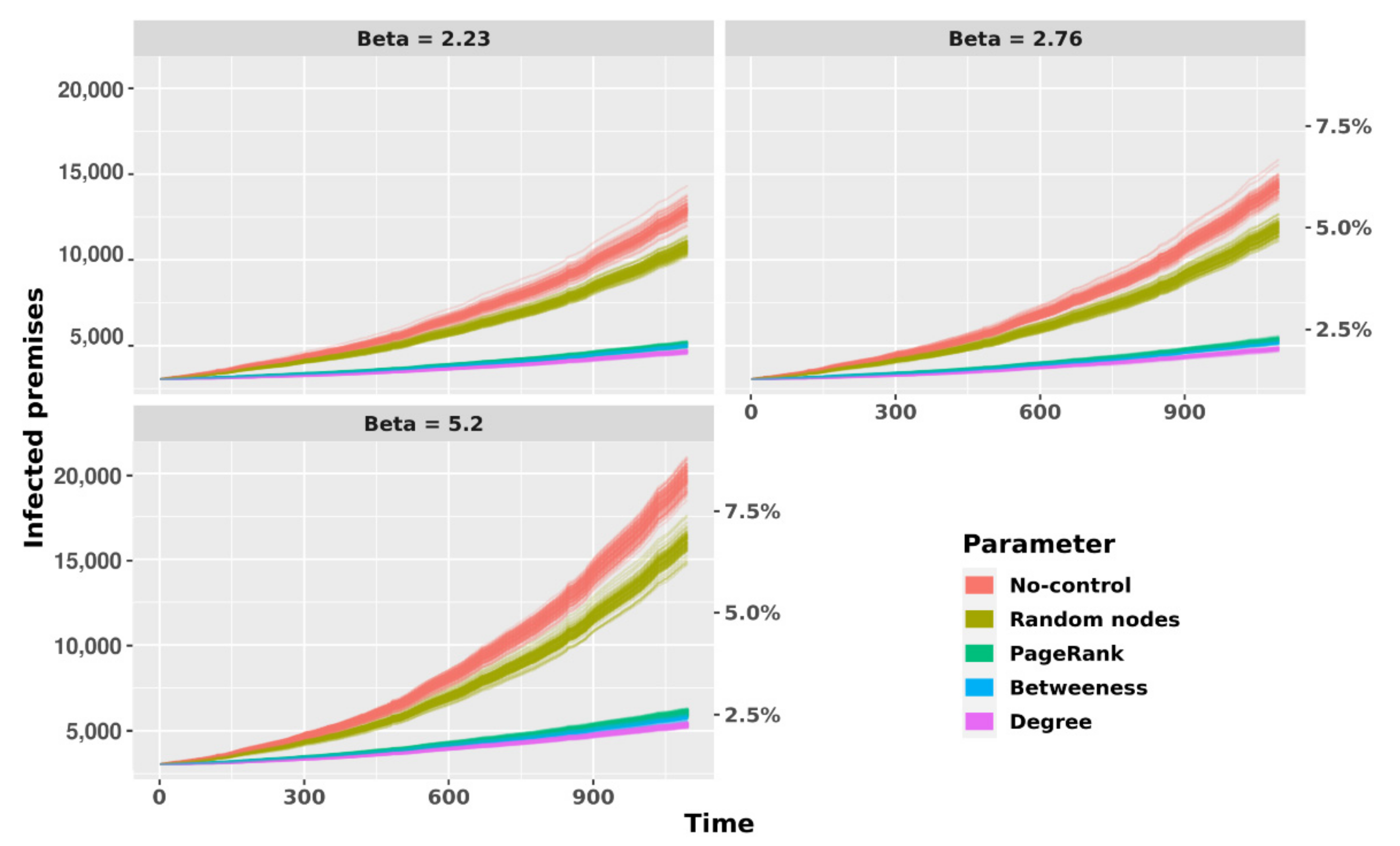
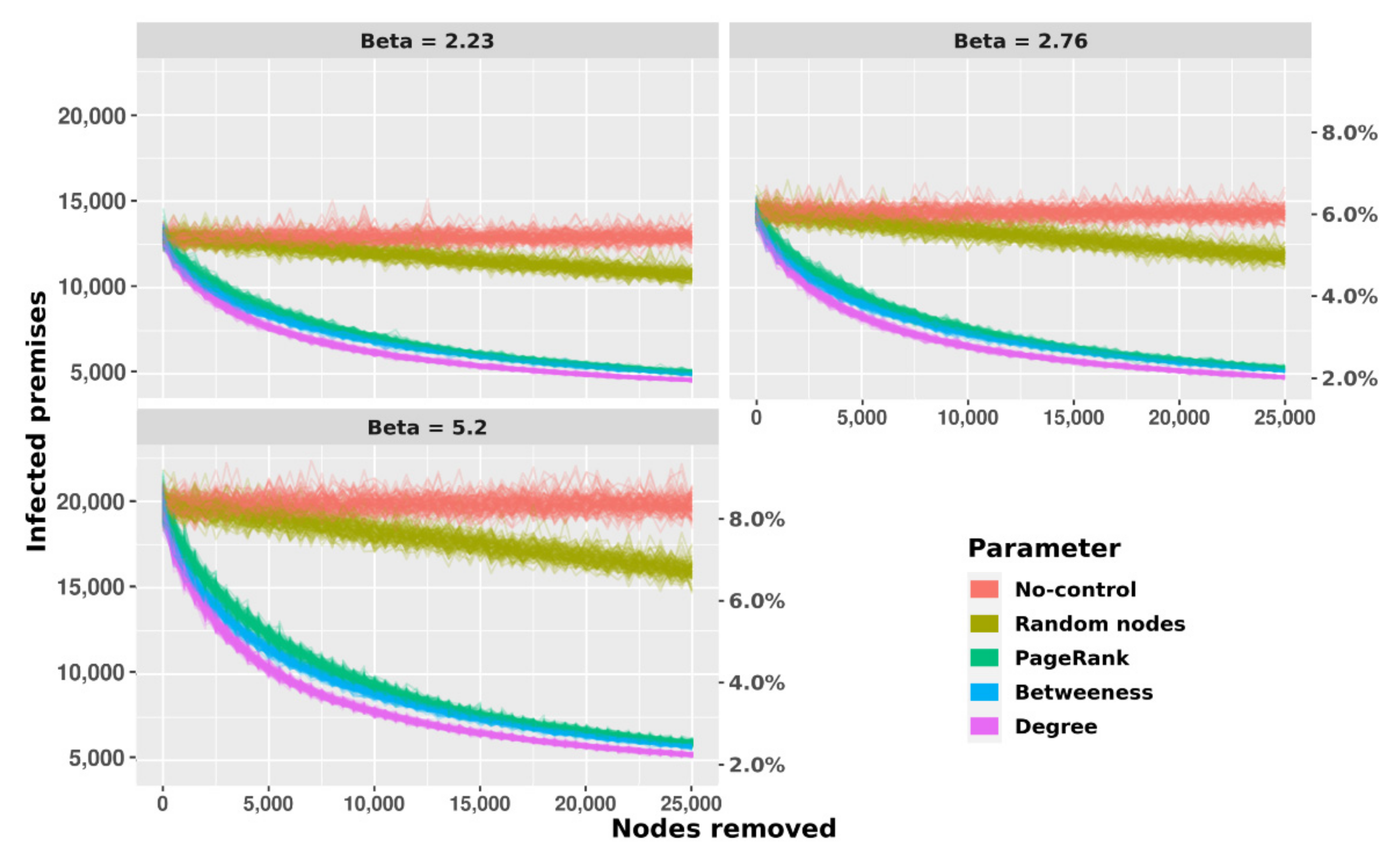
3.3. Disease Spread Modeling
3.4. Sensitivity Analyses
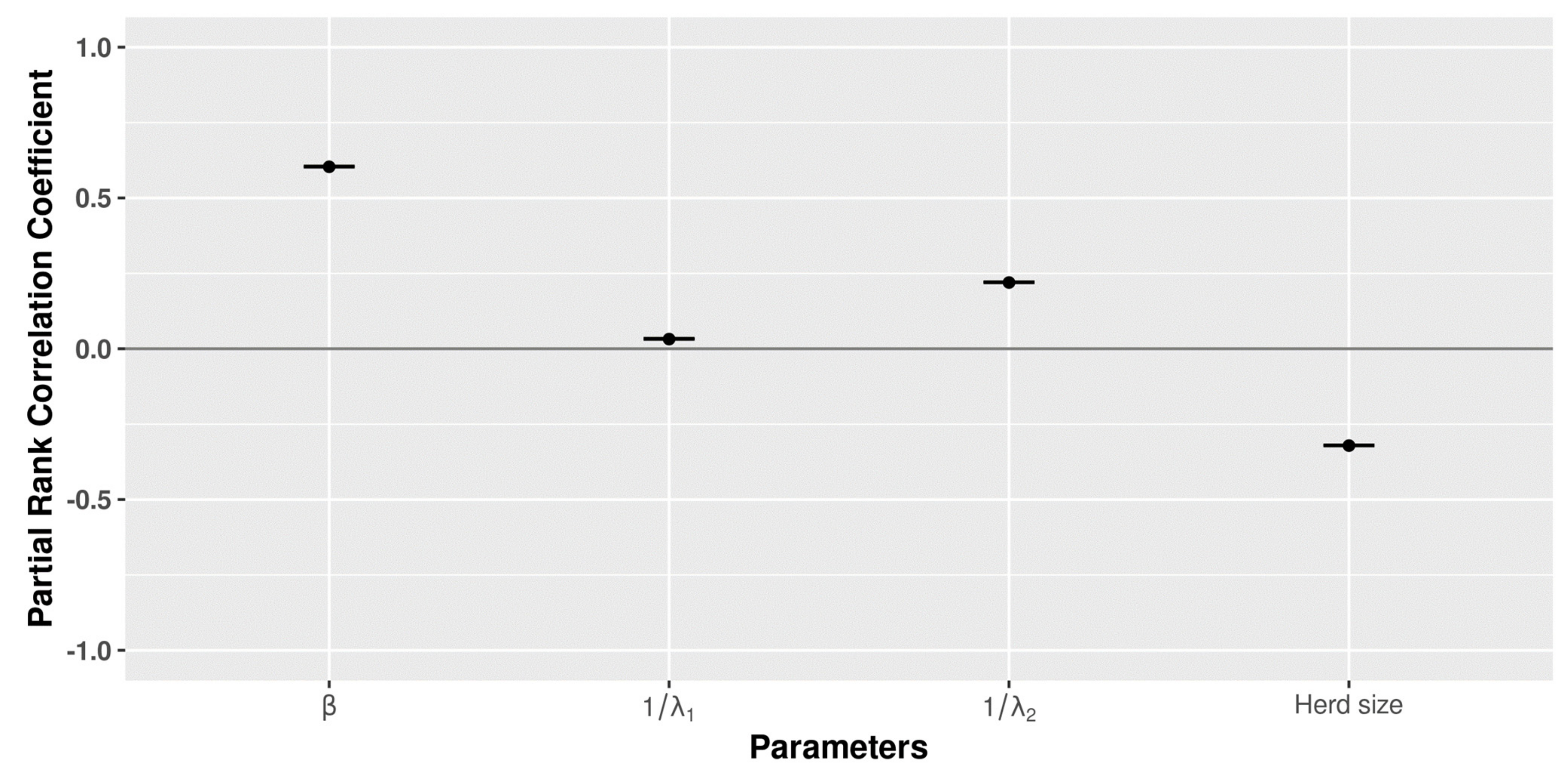
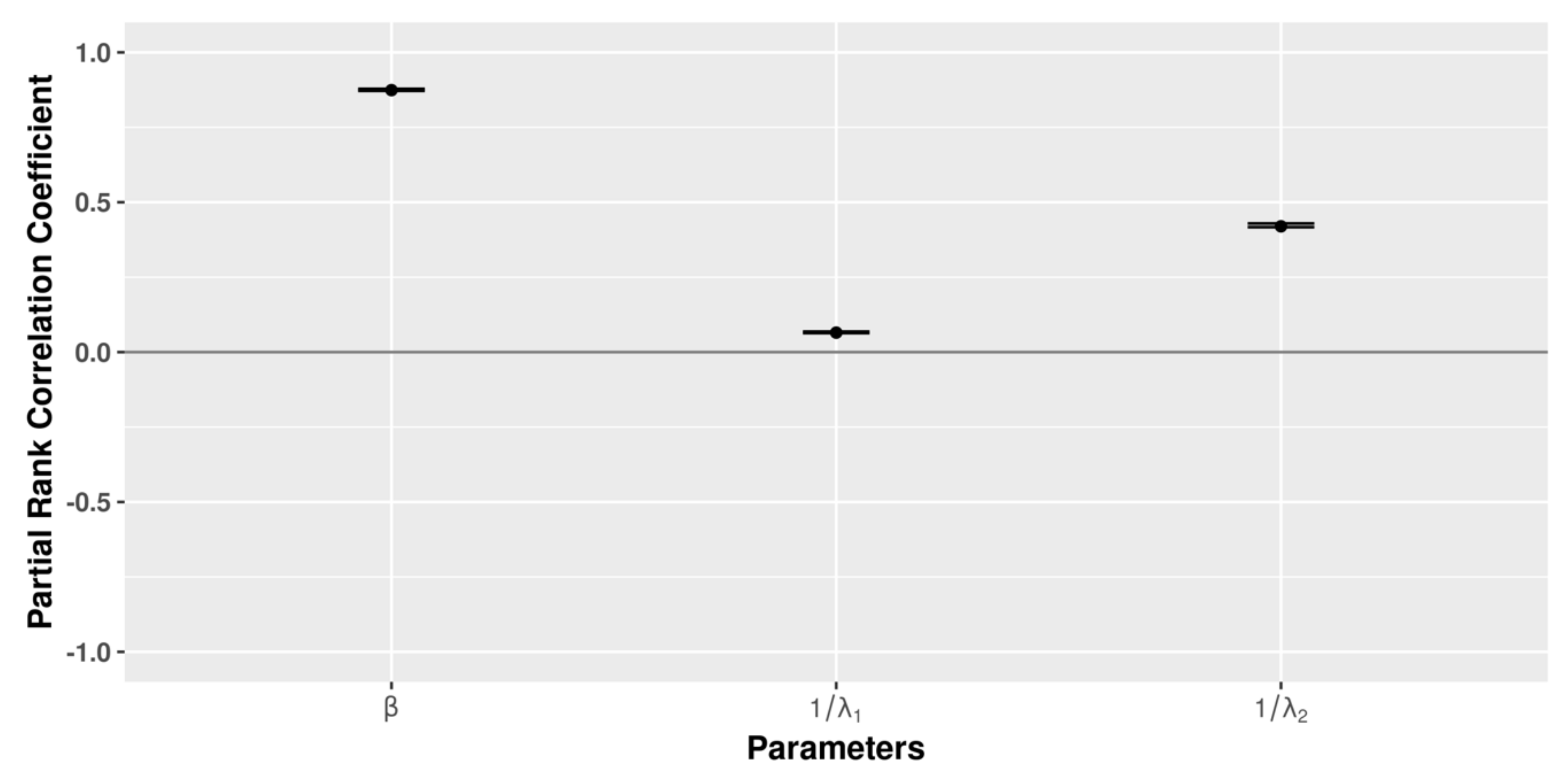
3.5. Global Sensitivity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bajardi, P.; Barrat, A.; Savini, L.; Colizza, V. Optimizing surveillance for livestock disease spreading through animal movements. J. R. Soc. Interface 2012, 9, 2814–2825. [Google Scholar] [CrossRef] [PubMed]
- Bajardi, P.; Barrat, A.; Natale, F.; Savini, L.; Colizza, V. Dynamical patterns of cattle trade movements. PLoS ONE 2011, 6, e19869. [Google Scholar] [CrossRef] [PubMed]
- Moslonka-Lefebvre, M.; Gilligan, C.A.; Monod, H.; Belloc, C.; Ezanno, P.; Filipe, J.A.N.; Vergu, E. Market analyses of livestock trade networks to inform the prevention of joint economic and epidemiological risks. J. R. Soc. Interface 2016, 13, 20151099. [Google Scholar] [CrossRef] [PubMed]
- Ortiz-Pelaez, A.; Pfeiffer, D.U.; Soares-Magalhães, R.J.; Guitian, F.J. Use of social network analysis to characterize the pattern of animal movements in the initial phases of the 2001 foot and mouth disease (FMD) epidemic in the UK. Prev. Vet. Med. 2006, 76, 40–55. [Google Scholar] [CrossRef]
- Payen, A.; Tabourier, L.; Latapy, M. Spreading dynamics in a cattle trade network: Size, speed, typical profile and consequences on epidemic control strategies. PLoS ONE 2019, 14, e0217972. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, M.; Mitchell, A.; Bourn, D.; Mawdsley, J.; Clifton-Hadley, R.; Wint, W. Cattle movements and bovine tuberculosis in Great Britain. Nature 2005, 435, 491–496. [Google Scholar] [CrossRef]
- Lage, A.; Roxo, E.; Müller, E.; Poester, F.; Cavalléro, J. Programa Nacional de Controle e Erradicação da Brucelose e da Tuberculose Animal (PNCEBT); Ministério da Agricultura, Pecuária e Abastecimento: Brasilia, Brazil, 2006.
- Ferreira Neto, J.S.; da Silveira, G.B.; Rosa, B.M.; Gonçalves, V.S.P.; Grisi-Filho, J.H.H.; Amaku, M.; Dias, R.A.; Ferreira, F.; Heinemann, M.B.; Telles, E.O.; et al. Analysis of 15 years of the national program for the control and eradication of animal brucellosis and tuberculosis, Brazil. Semin. Ciências Agrárias 2016, 37, 3385. [Google Scholar] [CrossRef] [Green Version]
- Moustakas, A. Spatio-temporal data mining in ecological and veterinary epidemiology. Stoch. Environ. Res. Risk Assess. 2017, 31, 829–834. [Google Scholar] [CrossRef] [Green Version]
- VanderWaal, K.; Morrison, R.B.; Neuhauser, C.; Vilalta, C.; Perez, A.M. Translating big data into smart data for veterinary epidemiology. Front. Vet. Sci. 2017, 4, 110. [Google Scholar] [CrossRef]
- Mekonnen, G.A.; Ameni, G.; Wood, J.L.N.; Berg, S.; Conlan, A.J.K. Network analysis of dairy cattle movement and associations with bovine tuberculosis spread and control in emerging dairy belts of Ethiopia. BMC Vet. Res. 2019, 15, 262. [Google Scholar] [CrossRef]
- VanderWaal, K.; Enns, E.A.; Picasso, C.; Alvarez, J.; Perez, A.; Fernandez, F.; Gil, A.; Craft, M.; Wells, S. Optimal surveillance strategies for bovine tuberculosis in a low-prevalence country. Sci. Rep. 2017, 7, 4140. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, V.S.P.; de Moraes, G.M. The application of epidemiology in national veterinary services: Challenges and threats in Brazil. Prev. Vet. Med. 2017, 137, 140–146. [Google Scholar] [CrossRef] [PubMed]
- Savini, L.; Candeloro, L.; Conte, A.; De Massis, F.; Giovannini, A. Development of a forecasting model for brucellosis spreading in the Italian cattle trade network aimed to prioritise the field interventions. PLoS ONE 2017, 12, e0177313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- IBGE. Banco de Tabelas Estatisticas—Instituto Brasiliro de Geografia e Estatistica Brazil. Available online: https://sidra.ibge.gov.br/home/pms/brasil (accessed on 17 August 2020).
- Queiroz, M.R.; Groff, A.C.M.; dos Silva, N.; Grisi-Filho, J.H.H.; Amaku, M.; Dias, R.A.; Telles, E.O.; Heinemann, M.B.; Ferreira Neto, J.S.; Gonçalves, V.S.P.; et al. Epidemiological status of bovine tuberculosis in the State of Rio Grande Do Sul, Brazil. Semin. Ciências Agrárias 2016, 37, 3647. [Google Scholar] [CrossRef] [Green Version]
- MAPA Regulamento Tecnico—Ministério da Agricultura, Pecuária e Abastecimento, Brazil. Available online: https://www.gov.br/agricultura/pt-br/assuntos/sanidade-animal-e-vegetal/saude-animal/programas-de-saude-animal/brucelose-e-tuberculose/principais-normas-pncebt/in-10-de-3-de-marco-de-2017-aprova-o-regulamento-tecnico-do-pncebt.pdf/view (accessed on 17 August 2020).
- Palisson, A.; Courcoul, A.; Durand, B. Role of cattle movements in bovine tuberculosis spread in france between 2005 and 2014. PLoS ONE 2016, 11, e0152578. [Google Scholar] [CrossRef] [PubMed]
- Green, D.M.; Kiss, I.Z.; Mitchell, A.P.; Kao, R.R. Estimates for local and movement-based transmission of bovine tuberculosis in British cattle. Proc. R. Soc. B Biol. Sci. 2008, 275, 1001–1005. [Google Scholar] [CrossRef] [PubMed]
- Christen, A.; Maulén-Yañez, M.A.; González-Olivares, E.; Curé, M. Modeling a SI epidemic with stochastic transmission: Hyperbolic incidence rate. J. Math. Biol. 2018, 76, 1005–1026. [Google Scholar] [CrossRef] [PubMed]
- Birch, C.P.D.; Goddard, A.; Tearne, O. A new bovine tuberculosis model for England and Wales (BoTMEW) to simulate epidemiology, surveillance and control. BMC Vet. Res. 2018, 14, 273. [Google Scholar] [CrossRef]
- Picasso-Risso, C.; Alvarez, J.; VanderWaal, K.; Kinsley, A.; Gil, A.; Wells, S.J.; Perez, A. Modeling the effect of test-and-slaughter strategies to control bovine tuberculosis in endemic high prevalence herds. Transbound. Emerg. Dis. 2020, tbed.13774. [Google Scholar] [CrossRef]
- Wasserman, S.; Faust, K. Social Network Analysis: Methods and Applications; Structural Analysis in the Social Sciences; Cambridge University Press: Cambridge, UK, 1994. [Google Scholar]
- Brin, S.; Page, L. The anatomy of a large-scale hypertextual Web search engine. Comput. Netw. ISDN Syst. 1998, 30, 107–117. [Google Scholar] [CrossRef]
- Fielding, H.R.; McKinley, T.J.; Silk, M.J.; Delahay, R.J.; McDonald, R.A. Contact chains of cattle farms in Great Britain. R. Soc. Open Sci. 2019, 6, 180719. [Google Scholar] [CrossRef] [Green Version]
- Broder, A.; Kumar, R.; Maghoul, F.; Raghavan, P.; Rajagopalan, S.; Stata, R.; Tomkins, A.; Wiener, J. Graph structure in the Web. Comput. Netw. 2000, 33, 309–320. [Google Scholar] [CrossRef]
- Knific, T.; Ocepek, M.; Kirbiš, A.; Lentz, H.H.K. Implications of cattle trade for the spread and control of infectious diseases in Slovenia. Front. Vet. Sci. 2020, 6, 454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holme, P.; Saramäki, J. Temporal networks. Phys. Rep. 2012, 519, 97–125. [Google Scholar] [CrossRef] [Green Version]
- VanderWaal, K.; Enns, E.A.; Picasso, C.; Packer, C.; Craft, M.E. Evaluating empirical contact networks as potential transmission pathways for infectious diseases. J. R. Soc. Interface 2016, 13, 20160166. [Google Scholar] [CrossRef]
- Pozo, P.; VanderWaal, K.; Grau, A.; de la Cruz, M.L.; Nacar, J.; Bezos, J.; Perez, A.; Minguez, O.; Alvarez, J. Analysis of the cattle movement network and its association with the risk of bovine tuberculosis at the farm level in Castilla y Leon, Spain. Transbound. Emerg. Dis. 2019, 66, 327–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kulldorff, M. A spatial scan statistic. Commun. Stat. Theory Methods 1997, 26, 1481–1496. [Google Scholar] [CrossRef]
- Álvarez, J.; Bezos, J.; de la Cruz, M.L.; Casal, C.; Romero, B.; Domínguez, L.; de Juan, L.; Pérez, A. Bovine tuberculosis: Within-herd transmission models to support and direct the decision-making process. Res. Vet. Sci. 2014, 97, S61–S68. [Google Scholar] [CrossRef]
- Gillespie, D.T. Exact stochastic simulation of coupled chemical reactions. J. Phys. Chem. 1977, 81, 2340–2361. [Google Scholar] [CrossRef]
- Perez, A.M.; Ward, M.P.; Charmandarián, A.; Ritacco, V. Simulation model of within-herd transmission of bovine tuberculosis in Argentine dairy herds. Prev. Vet. Med. 2002, 54, 361–372. [Google Scholar] [CrossRef]
- Verteramo Chiu, L.J.; Tauer, L.W.; Smith, R.L.; Grohn, Y.T. Assessment of the bovine tuberculosis elimination protocol in the United States. J. Dairy Sci. 2019, 102, 2384–2400. [Google Scholar] [CrossRef] [Green Version]
- Fischer, E.A.J.; van Roermund, H.J.W.; Hemerik, L.; van Asseldonk, M.A.P.M.; de Jong, M.C.M. Evaluation of surveillance strategies for bovine tuberculosis (Mycobacterium bovis) using an individual based epidemiological model. Prev. Vet. Med. 2005, 67, 283–301. [Google Scholar] [CrossRef]
- Barlow, N.D.; Kean, J.M.; Hickling, G.; Livingstone, P.G.; Robson, A.B. A simulation model for the spread of bovine tuberculosis within New Zealand cattle herds. Prev. Vet. Med. 1997, 32, 57–75. [Google Scholar] [CrossRef]
- Smith, R.L.; Schukken, Y.H.; Lu, Z.; Mitchell, R.M.; Grohn, Y.T. Development of a model to simulate infection dynamics of Mycobacterium bovis in cattle herds in the United States. J. Am. Vet. Med. Assoc. 2013, 243, 411–423. [Google Scholar] [CrossRef] [Green Version]
- Allepuz, A.; Casal, J.; Napp, S.; Saez, M.; Alba, A.; Vilar, M.; Domingo, M.; González, M.A.; Duran-Ferrer, M.; Vicente, J.; et al. Analysis of the spatial variation of Bovine tuberculosis disease risk in Spain (2006–2009). Prev. Vet. Med. 2011, 100, 44–52. [Google Scholar] [CrossRef] [Green Version]
- O’Hare, A.; Orton, R.J.; Bessell, P.R.; Kao, R.R. Estimating epidemiological parameters for bovine tuberculosis in British cattle using a Bayesian partial-likelihood approach. Proc. R. Soc. B Biol. Sci. 2014, 281, 20140248. [Google Scholar] [CrossRef]
- Rose, H.; Wall, R. Modelling the impact of climate change on spatial patterns of disease risk: Sheep blowfly strike by Lucilia sericata in Great Britain. Int. J. Parasitol. 2011, 41, 739–746. [Google Scholar] [CrossRef]
- Yang, Y.; Wu, J.; Li, J.; Xu, X. Tuberculosis with relapse: A model. Math. Popul. Stud. 2017, 24, 3–20. [Google Scholar] [CrossRef]
- Bidah, S.; Zakary, O.; Rachik, M. Stability and global sensitivity analysis for an agree-disagree model: Partial rank correlation coefficient and Latin hypercube sampling methods. Int. J. Differ. Equ. 2020, 2020, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Nabi, K.N. Forecasting COVID-19 pandemic: A data-driven analysis. Chaos Solitons Fractals 2020, 139, 110046. [Google Scholar] [CrossRef]
- Guo, Z.; Xu, S.; Tong, L.; Dai, B.; Liu, Y.; Xiao, D. An artificially simulated outbreak of a respiratory infectious disease. BMC Public Health 2020, 20, 135. [Google Scholar] [CrossRef] [Green Version]
- Marino, S.; Hogue, I.B.; Ray, C.J.; Kirschner, D.E. A methodology for performing global uncertainty and sensitivity analysis in systems biology. J. Biol. 2008, 254, 178–196. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing. 2017. Available online: https://www.r-project.org/ (accessed on 18 January 2021).
- Csardi, G.; Nepusz, T. The igraph software package for complex network research. InterJ. Complex Syst. 2006, 1695, 1–9. [Google Scholar]
- Wickham, H.; Averick, M.; Bryan, J.; Chang, W.; McGowan, L.; François, R.; Grolemund, G.; Hayes, A.; Henry, L.; Hester, J.; et al. Welcome to the Tidyverse. J. Open Source Softw. 2019, 4, 1686. [Google Scholar] [CrossRef]
- Widgren, S.; Bauer, P.; Eriksson, R.; Engblom, S. Siminf: An R package for data-driven stochastic disease spread simulations. J. Stat. Softw. 2019. [Google Scholar] [CrossRef] [Green Version]
- Pebesma, E. Simple features for R: Standardized support for spatial vector data. R J. 2018, 10, 439. [Google Scholar] [CrossRef] [Green Version]
- Prado Siqueira, R. Brazilmaps: Brazilian Maps from Different Geographic Levels. 2017. Available online: https://rdrr.io/cran/brazilmaps/ (accessed on 18 January 2021).
- Kamvar, Z.N.; Cai, J.; Pulliam, J.R.C.; Schumacher, J.; Jombart, T. Epidemic curves made easy using the R package incidence. F1000Research 2019, 8, 139. [Google Scholar] [CrossRef] [Green Version]
- Stevenson, M.; Nunes, T.; Sanchez, J.; Thornton, R.; Reiczigel, J.; Robison-Cox, J.; Sebastiani, P. EpiR: An R Package for the Analysis of Epidemiological Data. 2013. Available online: http://www2.uaem.mx/r-mirror/web/packages/epiR/epiR.pdf (accessed on 18 January 2021).
- Jung, I.; Kulldorff, M.; Richard, O.J. A spatial scan statistic for multinomial data. Stat. Med. 2010, 29, 1910–1918. [Google Scholar] [CrossRef] [Green Version]
- Dos Santos, D.V.; Silva, G.S.E.; Weber, E.J.; Hasenack, H.; Groff, F.H.S.; Todeschini, B.; Borba, M.R.; Medeiros, A.A.R.; Leotti, V.B.; Canal, C.W.; et al. Identification of foot and mouth disease risk areas using a multi-criteria analysis approach. PLoS ONE 2017, 12, e0178464. [Google Scholar] [CrossRef]
- Natale, F.; Giovannini, A.; Savini, L.; Palma, D.; Possenti, L.; Fiore, G.; Calistri, P. Network analysis of Italian cattle trade patterns and evaluation of risks for potential disease spread. Prev. Vet. Med. 2009, 92, 341–350. [Google Scholar] [CrossRef]
- Gates, M.C.; Woolhouse, M.E.J. Controlling infectious disease through the targeted manipulation of contact network structure. Epidemics 2015, 12, 11–19. [Google Scholar] [CrossRef]
- Motta, P.; Porphyre, T.; Handel, I.; Hamman, S.M.; Ngwa, V.N.; Tanya, V.; Morgan, K.; Christley, R.; Barend, M. Implications of the cattle trade network in Cameroon for regional disease prevention and control. Sci. Rep. 2017, 7, 43932. [Google Scholar] [CrossRef] [Green Version]
- Willeberg, P.W.; McAloon, C.G.; Houtsma, E.; Higgins, I.; Clegg, T.A.; More, S.J. The Herd-level sensitivity of abattoir surveillance for bovine tuberculosis: Simulating the effects of current and potentially modified meat inspection procedures in Irish Cattle. Front. Vet. Sci. 2018, 5, 82. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, R.A.; Meneses, I.I.F.S.; Jorge, K.S.G.; Silva, M.R.; Santos, L.R.; Lilenbaum, W.; Etges, R.N.; Araújo, F.R. False-negative reactions to the comparative intradermal tuberculin test for bovine tuberculosis. Pesqui. Veterinária Bras. 2017, 37, 1380–1384. [Google Scholar] [CrossRef]
- Neves, E.D.; Mezalira, T.S.; Dias, E.H.; Dourado, M.R.; de Paula, M.K.; Gusman, C.R.; Da Silva Caetano, I.C.; Beltrami, J.M.; Otutumi, L.K. Lesões de tuberculose bovina em abatedouros-frigoríficos no Brasil: Bibliometria. J. Interdiscip. Biociências 2017, 2, 22–27. [Google Scholar] [CrossRef] [Green Version]
- Grisi Filho, J.H.H.; Rosales, C.A.R.; Ferreira, F.; Amaku, M.; Dias, R.A.; Ferreira Neto, J.S. Análise epidemiológica das condenações de bovinos por tuberculose em abatedouros do estado de São Paulo. Arq. Do Inst. Biológico 2011, 78, 175–181. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Definition | Reference |
|---|---|---|
| Nodes | The unit of interest in network analysis, for example, premises or slaughterhouses. | [23] |
| Edge | Link between two nodes in the network. | |
| Degree (k) | Number of unique contacts to and from a specific premise. When the directionality is considered, the ingoing and outgoing contacts are defined: out-degree is the number of contacts originating from a specific premise, and in-degree is the number of contacts coming into a specific premise. | |
| Movements | The number of animal movements records over a certain period of time. | |
| PageRank | Google PageRank measure, a link analysis algorithm that produces a ranking based on the importance for all nodes in a network with a range of values between zero and one. The PageRank calculation considers the indegree of a given premise and the indegree of its neighbors. | [24] |
| Betweenness | The extent to which a node lies on paths connecting other pairs of nodes, defined by the number of geodesics (shortest paths) going through a node. | |
| Clustering coefficient | Measures the degree to which nodes in a network tend to cluster together (i.e., if A → B and B → C, what is the probability that A → C), with a range of values between zero and one. | |
| Giant weakly connected component (GWCC) | Proportion of nodes that are connected in the largest component when directionality of movement is ignored. | [23] |
| Giant strongly connected component (GSCC). | Proportion of the nodes that are connected in the largest component when directionality of movement is considered. | [23] |
| Out-Going Contact chain (OCC) | The outgoing contact chain (OCC) quantifies the number of ‘downstream’ premises that could potentially acquire infection from the index premises through outgoing animal movements, adhering to the chronological order of the movements. | [5,25] |
| Year | Nodes | Edges | Animals | Graph Density | Diameter | GSCC (%) | GWCC (%) | Clustering Coefficient | Mean Shortest Paths |
|---|---|---|---|---|---|---|---|---|---|
| 2016 | 158,469 | 316,455 | 4,809,197 | 1.26 × 10−5 | 40 | 29,148 (18.39) | 133,069 (83.97) | 0.021 | 10.71 |
| 2017 | 151,498 | 307,943 | 4,960,304 | 1.34 × 10−5 | 45 | 27,326 (18.04) | 125,685 (82.96) | 0.020 | 10.29 |
| 2018 | 149,754 | 314,282 | 5,030,184 | 1.40 × 10−5 | 35 | 26,797 (17.89) | 126,709 (84.61) | 0.017 | 9.86 |
| All years | 237,436 | 938,680 | 14,799,685 | 1.67 × 10−5 | 32 | 86,315 (36.35) | 223,086 (93.96) | 0.025 | 8.13 |
| No. of Cases in Cluster | ||||||
|---|---|---|---|---|---|---|
| Cluster | Radius (km) | No. of Herds in Cluster | Observed | Expected | Relative Risk | p-Value |
| 4 | 123.55 | 3629 | 257 | 146.91 | 1.82 | 3.18 × 10−11 |
| 8 | 20.47 | 418 | 52 | 19.1 | 2.76 | 0.000152 |
| 6 | 21.95 | 1406 | 43 | 12.23 | 3.55 | 3.44 × 10−6 |
| 3 | 197.9 | 21,766 | 517 | 167.34 | 3.56 | 1.00 × 10−17 |
| 7 | 21.04 | 1180 | 30 | 7.39 | 4.09 | 0.000134 |
| 1 | 328.17 | 55,109 | 1854 | 662.12 | 6.22 | 1.00 × 10−17 |
| 5 | 12.5 | 1134 | 32 | 4.43 | 7.29 | 3.52 × 10−11 |
| 2 | 73 | 15,744 | 801 | 124.82 | 8.56 | 1.00 × 10−17 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cardenas, N.C.; Pozo, P.; Lopes, F.P.N.; Grisi-Filho, J.H.H.; Alvarez, J. Use of Network Analysis and Spread Models to Target Control Actions for Bovine Tuberculosis in a State from Brazil. Microorganisms 2021, 9, 227. https://doi.org/10.3390/microorganisms9020227
Cardenas NC, Pozo P, Lopes FPN, Grisi-Filho JHH, Alvarez J. Use of Network Analysis and Spread Models to Target Control Actions for Bovine Tuberculosis in a State from Brazil. Microorganisms. 2021; 9(2):227. https://doi.org/10.3390/microorganisms9020227
Chicago/Turabian StyleCardenas, Nicolas Cespedes, Pilar Pozo, Francisco Paulo Nunes Lopes, José H. H. Grisi-Filho, and Julio Alvarez. 2021. "Use of Network Analysis and Spread Models to Target Control Actions for Bovine Tuberculosis in a State from Brazil" Microorganisms 9, no. 2: 227. https://doi.org/10.3390/microorganisms9020227