
Screening of Emerging Pollutants (EPs) in Estuarine Water and Phytoremediation Capacity of Tripolium pannonicum under Controlled Conditions

 , ,
, ,  , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
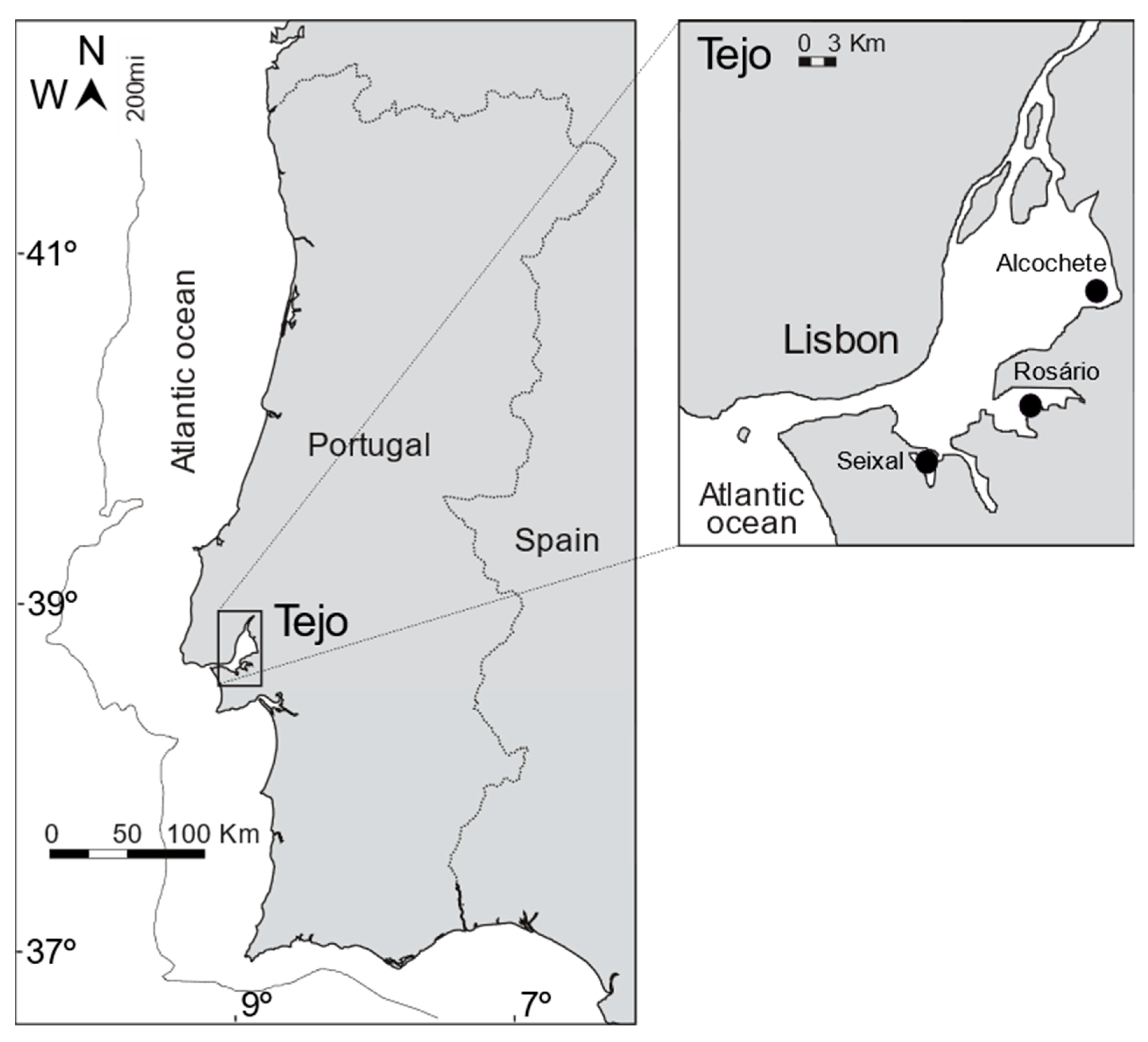
2.1. Sample Collection and Study Site Description
2.2. Water Sample Processing
2.3. Plant Cultivation under Controlled Conditions
2.4. Xenobiotic Degradation Using Crude Plant Extracts
2.5. Extraction of Xenobiotics from Plant Samples
2.6. LC/MS Conditions, Quantification and Linearity
2.7. Statistical Analysis
3. Results and Discussion
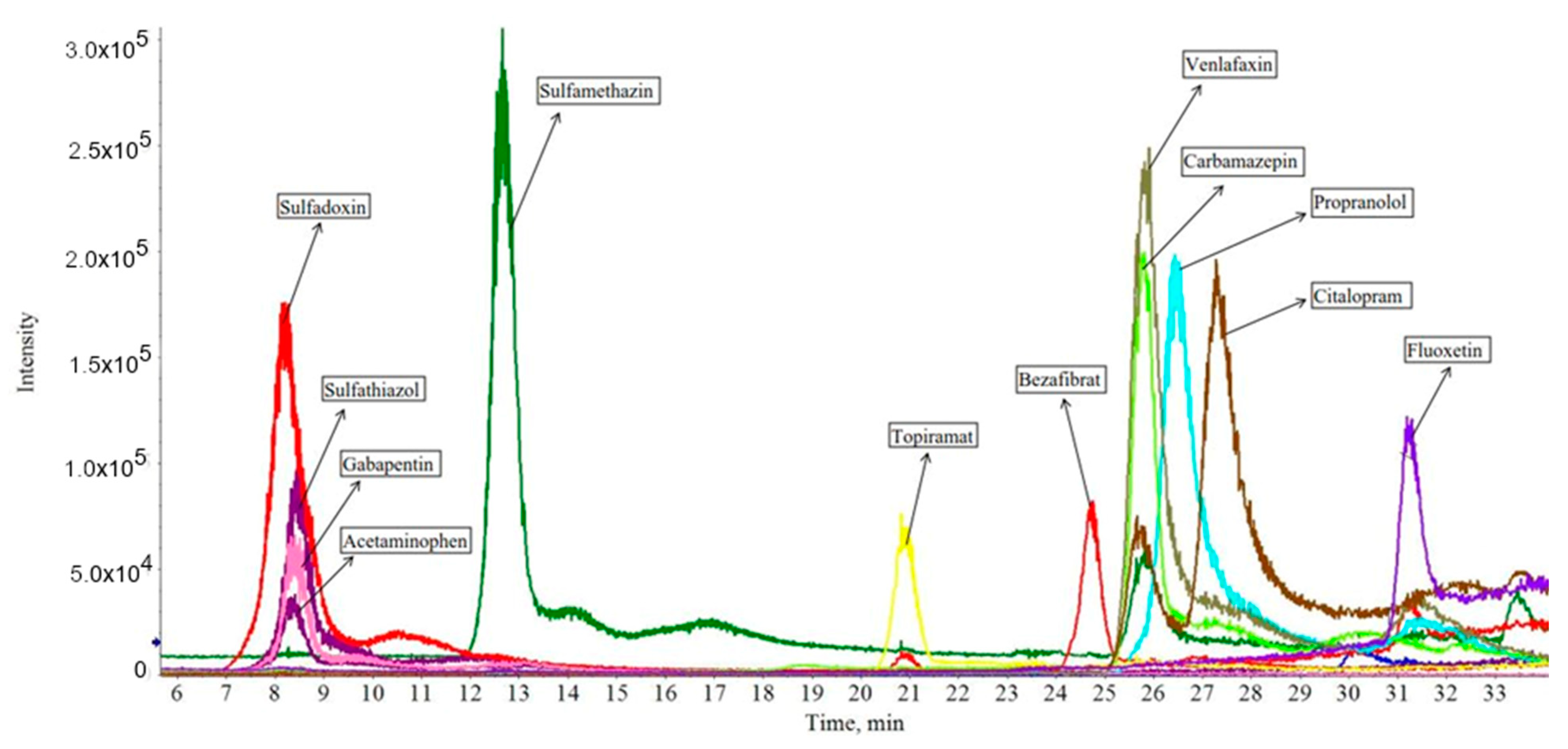
3.1. Standards Mass Spectra Analysis
3.2. Xenobiotic Concentration in Estuarine Water Samples
3.3. Exposure of the Halophyte Tripolium Pannonicum to Xenobiotics
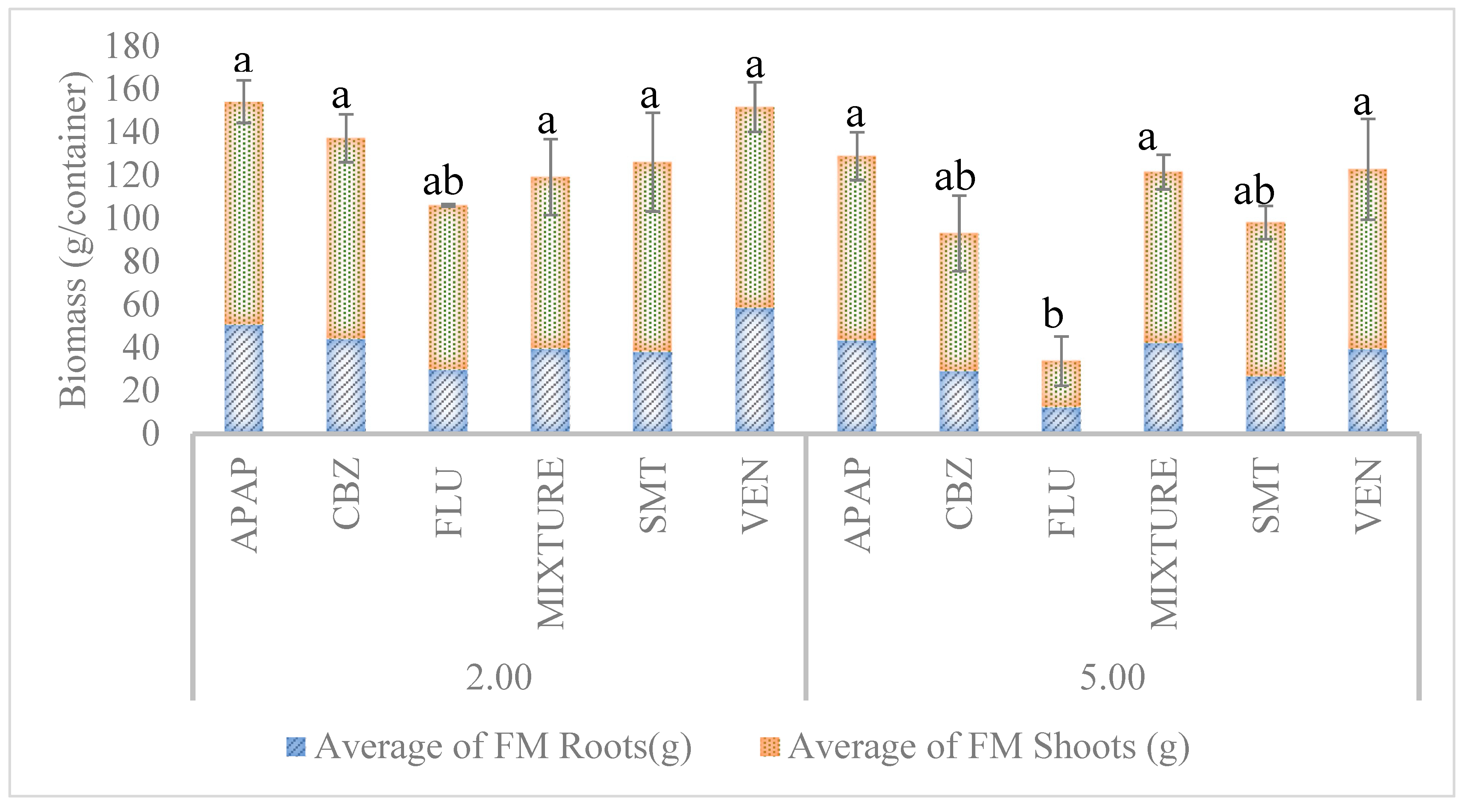
3.3.1. Influence of Xenobiotics on Biomass Yield
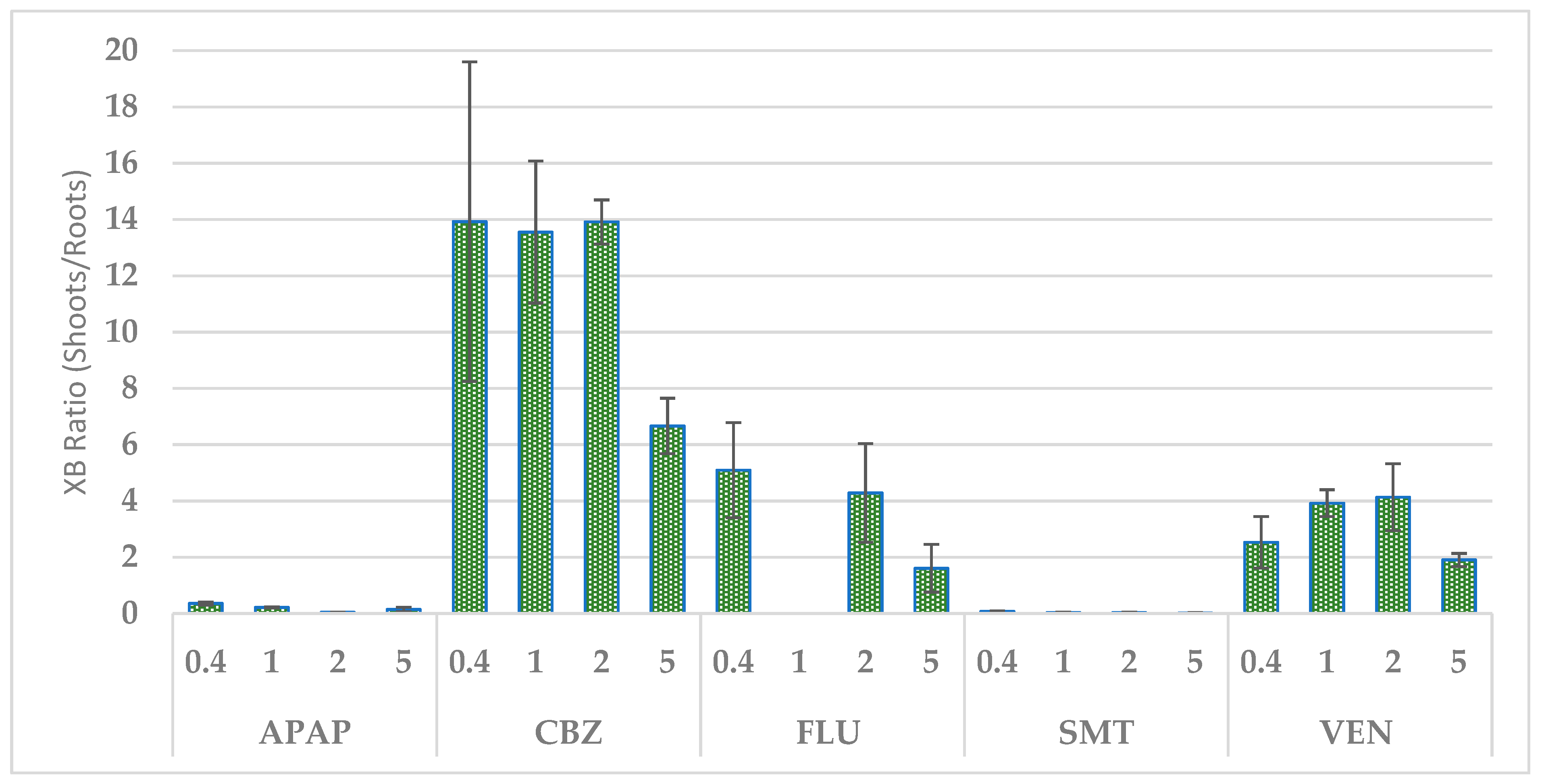
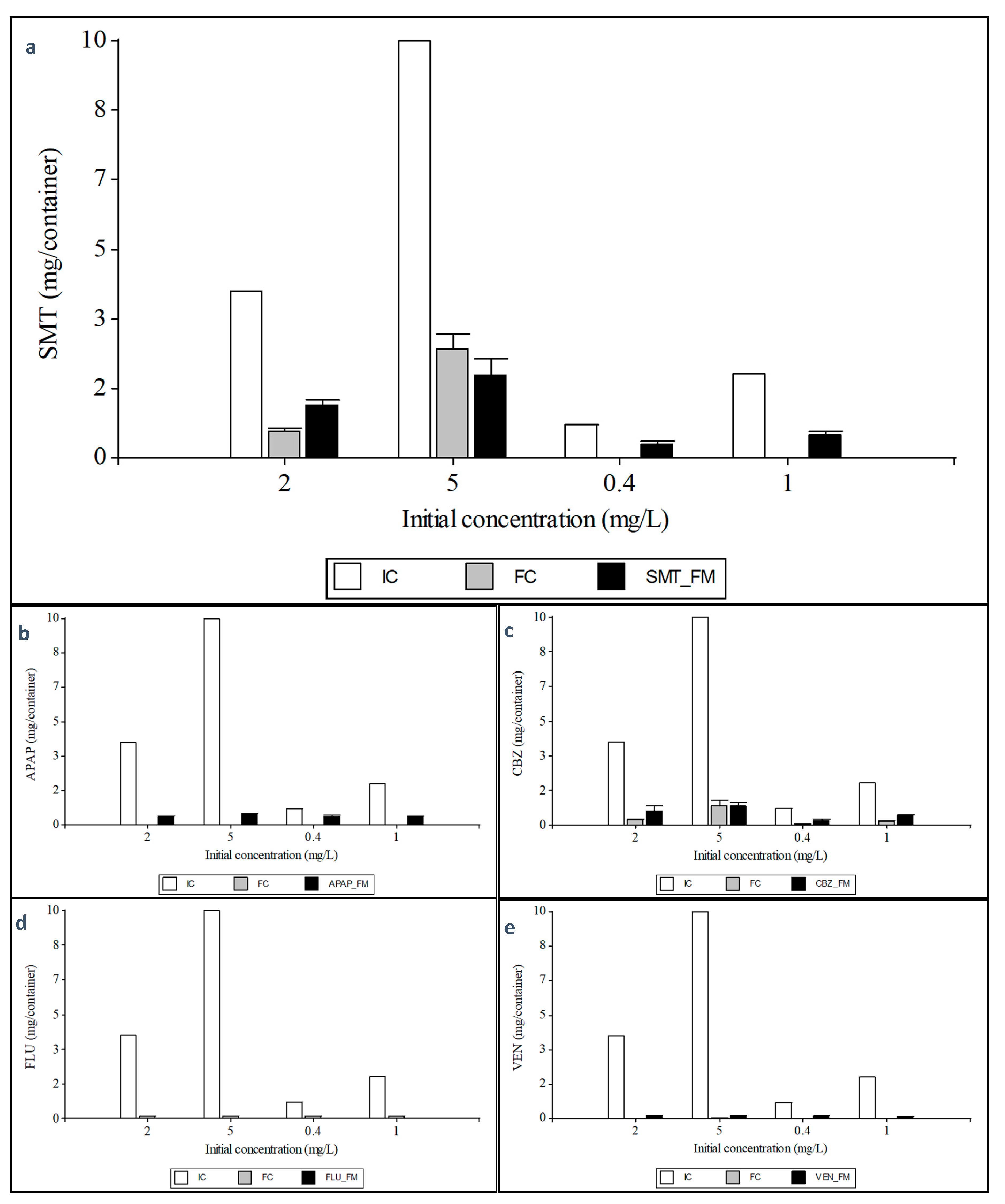
3.3.2. Uptake of Xenobiotics by Tripolium Pannonicum and Their Distribution in the Plant
3.3.3. Mass Balance
3.4. Degradation of Xenobiotics by Crude Plant Extracts
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gavrilescu, M.; Demnerová, K.; Aamand, J.; Agathos, S.; Fava, F. Emerging Pollutants in the Environment: Present and Future Challenges in Biomonitoring, Ecological Risks and Bioremediation. New Biotechnol. 2015, 32, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, A.R.M.; Cardoso, V.V.; Rodrigues, A.; Ferreira, E.; Benoliel, M.J.; Duarte, E.A. Occurrence and Analysis of Endocrine-Disrupting Compounds in a Water Supply System. Environ. Monit. Assess 2015, 187, 139. [Google Scholar] [CrossRef]
- Reis-Santos, P.; Pais, M.; Duarte, B.; Caçador, I.; Freitas, A.; Vila Pouca, A.S.; Barbosa, J.; Leston, S.; Rosa, J.; Ramos, F.; et al. Screening of Human and Veterinary Pharmaceuticals in Estuarine Waters: A Baseline Assessment for the Tejo Estuary. Mar. Pollut. Bull. 2018, 135, 1079–1084. [Google Scholar] [CrossRef]
- Chemicals Production and Consumption Statistics—Statistics Explained. Available online: https://ec.europa.eu/eurostat/statistics-explained/index.php/Chemicals_production_and_consumption_statistics (accessed on 14 December 2020).
- Roma, J.; Matos, A.R.; Vinagre, C.; Duarte, B. Engineered Metal Nanoparticles in the Marine Environment: A Review of the Effects on Marine Fauna. Mar. Environ. Res. 2020, 161, 105110. [Google Scholar] [CrossRef]
- Lopes, D.G.; Duarte, I.A.; Antunes, M.; Fonseca, V.F. Effects of Antidepressants in the Reproduction of Aquatic Organisms: A Meta-Analysis. Aquat. Toxicol. 2020, 227, 105569. [Google Scholar] [CrossRef]
- Duarte, I.A.; Reis-Santos, P.; Novais, S.C.; Rato, L.D.; Lemos, M.F.L.; Freitas, A.; Pouca, A.S.V.; Barbosa, J.; Cabral, H.N.; Fonseca, V.F. Depressed, Hypertense and Sore: Long-Term Effects of Fluoxetine, Propranolol and Diclofenac Exposure in a Top Predator Fish. Sci. Total Environ. 2020, 712, 136564. [Google Scholar] [CrossRef]
- Cruz de Carvalho, R.; Feijão, E.; Matos, A.R.; Cabrita, M.T.; Novais, S.C.; Lemos, M.F.L.; Caçador, I.; Marques, J.C.; Reis-Santos, P.; Fonseca, V.F.; et al. Glyphosate-Based Herbicide Toxicophenomics in Marine Diatoms: Impacts on Primary Production and Physiological Fitness. Appl. Sci. 2020, 10, 7391. [Google Scholar] [CrossRef]
- Feijão, E.; Cruz de Carvalho, R.; Duarte, I.A.; Matos, A.R.; Cabrita, M.T.; Novais, S.C.; Lemos, M.F.L.; Caçador, I.; Marques, J.C.; Reis-Santos, P.; et al. Fluoxetine Arrests Growth of the Model Diatom Phaeodactylum Tricornutum by Increasing Oxidative Stress and Altering Energetic and Lipid Metabolism. Front. Microbiol. 2020, 11, 1803. [Google Scholar] [CrossRef]
- Silva, M.; Feijão, E.; da Cruz de Carvalho, R.; Duarte, I.A.; Matos, A.R.; Cabrita, M.T.; Barreiro, A.; Lemos, M.F.L.; Novais, S.C.; Marques, J.C.; et al. Comfortably Numb: Ecotoxicity of the Non-Steroidal Anti-Inflammatory Drug Ibuprofen on Phaeodactylum Tricornutum. Mar. Environ. Res. 2020, 161, 105109. [Google Scholar] [CrossRef]
- Duarte, B.; Prata, D.; Matos, A.R.; Cabrita, M.T.; Caçador, I.; Marques, J.C.; Cabral, H.N.; Reis-Santos, P.; Fonseca, V.F. Ecotoxicity of the Lipid-Lowering Drug Bezafibrate on the Bioenergetics and Lipid Metabolism of the Diatom Phaeodactylum Tricornutum. Sci. Total Environ. 2019, 650, 2085–2094. [Google Scholar] [CrossRef]
- Vos, J.G.; Dybing, E.; Greim, H.A.; Ladefoged, O.; Lambré, C.; Tarazona, J.V.; Brandt, I.; Vethaak, A.D. Health Effects of Endocrine-Disrupting Chemicals on Wildlife, with Special Reference to the European Situation. Crit. Rev. Toxicol. 2000, 30, 71–133. [Google Scholar] [CrossRef] [PubMed]
- Bergmann, A.; Fohrmann, R.U.; Weber, F. Zusammenstellung von Monitoringdaten Zu Umweltkonzentrationen von Arzneimitteln; UBA-Texte 66/2011; Umweltbundesamt: Dessau-Roßlau, Germany, 2011. [Google Scholar]
- Klein, E.Y.; Van Boeckel, T.P.; Martinez, E.M.; Pant, S.; Gandra, S.; Levin, S.A.; Goossens, H.; Laxminarayan, R. Global Increase and Geographic Convergence in Antibiotic Consumption between 2000 and 2015. Proc. Natl. Acad. Sci. USA 2018, 115, E3463–E3470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fonseca, V.F.; Reis-Santos, P.; Duarte, B.; Cabral, H.N.; Caçador, M.I.; Vaz, N.; Dias, J.M.; Pais, M.P. Roving Pharmacies: Modelling the Dispersion of Pharmaceutical Contamination in Estuaries. Ecol. Indic. 2020, 115, 106437. [Google Scholar] [CrossRef]
- Schröder, P.; Collins, C.D. (Eds.) Organic Xenobiotics and Plants: From Mode of Action to Ecophysiology. In Plant Ecophysiology; Springer: Dordrecht, The Netherlands, 2011; ISBN 978-90-481-9851-1. [Google Scholar]
- Farrington, J.W. Biogeochemical Processes Governing Exposure and Uptake of Organic Pollutant Compounds in Aquatic Organisms. Environ. Health Perspect. 1991, 90, 75–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turcios, A.E.; Weichgrebe, D.; Papenbrock, J. Uptake and Biodegradation of the Antimicrobial Sulfadimidine by the Species Tripolium Pannonicum Acting as Biofilter and Its Further Biodegradation by Anaerobic Digestion and Concomitant Biogas Production. Bioresour. Technol. 2016, 219, 687–693. [Google Scholar] [CrossRef]
- Li, C.; Zhou, K.; Qin, W.; Tian, C.; Qi, M.; Yan, X.; Han, W. A Review on Heavy Metals Contamination in Soil: Effects, Sources, and Remediation Techniques. Soil Sediment Contam. Int. J. 2019, 28, 380–394. [Google Scholar] [CrossRef]
- Turcios, A.; Papenbrock, J. Sustainable Treatment of Aquaculture Effluents—What Can We Learn from the Past for the Future? Sustainability 2014, 6, 836–856. [Google Scholar] [CrossRef] [Green Version]
- Cruz de Carvalho, R.; Feijão, E.; Kletschkus, E.; Marques, J.C.; Reis-Santos, P.; Fonseca, V.F.; Papenbrock, J.; Caçador, I.; Duarte, B. Halophyte Bio-Optical Phenotyping: A Multivariate Photochemical Pressure Index (Multi-PPI) to Classify Salt Marsh Anthropogenic Pressures Levels. Ecol. Indic. 2020, 119, 106816. [Google Scholar] [CrossRef]
- Caçador, I.; Costa, J.L.; Duarte, B.; Silva, G.; Medeiros, J.P.; Azeda, C.; Castro, N.; Freitas, J.; Pedro, S.; Almeida, P.R.; et al. Macroinvertebrates and Fishes as Biomonitors of Heavy Metal Concentration in the Seixal Bay (Tagus Estuary): Which Species Perform Better? Ecol. Indic. 2012, 19, 184–190. [Google Scholar] [CrossRef]
- Duarte, B.; Caetano, M.; Almeida, P.R.; Vale, C.; Caçador, I. Accumulation and Biological Cycling of Heavy Metal in Four Salt Marsh Species, from Tagus Estuary (Portugal). Environ. Pollut. 2010, 158, 1661–1668. [Google Scholar] [CrossRef]
- Caçador, I.; Neto, J.M.; Duarte, B.; Barroso, D.V.; Pinto, M.; Marques, J.C. Development of an Angiosperm Quality Assessment Index (AQuA-Index) for Ecological Quality Evaluation of Portuguese Water Bodies—A Multi-Metric Approach. Ecol. Indic. 2013, 25, 141–148. [Google Scholar] [CrossRef]
- Pereira, A.M.P.T.; Silva, L.J.G.; Meisel, L.M.; Lino, C.M.; Pena, A. Environmental Impact of Pharmaceuticals from Portuguese Wastewaters: Geographical and Seasonal Occurrence, Removal and Risk Assessment. Environ. Res. 2015, 136, 108–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sousa, M.A.; Gonçalves, C.; Cunha, E.; Hajšlová, J.; Alpendurada, M.F. Cleanup Strategies and Advantages in the Determination of Several Therapeutic Classes of Pharmaceuticals in Wastewater Samples by SPE–LC–MS/MS. Anal. Bioanal. Chem. 2011, 399, 807–822. [Google Scholar] [CrossRef]
- Buhmann, A.K.; Waller, U.; Wecker, B.; Papenbrock, J. Optimization of Culturing Conditions and Selection of Species for the Use of Halophytes as Biofilter for Nutrient-Rich Saline Water. Agric. Water Manag. 2015, 149, 102–114. [Google Scholar] [CrossRef]
- Turcios, A.E.; Papenbrock, J. Enzymatic Degradation of the Antibiotic Sulfamethazine by Using Crude Extracts of Different Halophytic Plants. Int. J. Phytoremediat. 2019, 21, 1104–1111. [Google Scholar] [CrossRef]
- Turcios, A.E.; Weichgrebe, D.; Papenbrock, J. Effect of Salt and Sodium Concentration on the Anaerobic Methanisation of the Halophyte Tripolium Pannonicum. Biomass Bioenergy 2016, 87, 69–77. [Google Scholar] [CrossRef]
- PubChem. Available online: https://pubchem.ncbi.nlm.nih.gov/ (accessed on 15 December 2020).
- Zhang, H.; Deng, X.; Miki, D.; Cutler, S.; La, H.; Hou, Y.-J.; Oh, J.; Zhu, J.-K. Sulfamethazine Suppresses Epigenetic Silencing in Arabidopsis by Impairing Folate Synthesis. Plant Cell 2012, 24, 1230–1241. [Google Scholar] [CrossRef] [Green Version]
- Sohel, A.J.; Shutter, M.C.; Molla, M. Fluoxetine. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2020. [Google Scholar]
- Kang, K.; Kim, Y.-S.; Park, S.; Back, K. Senescence-Induced Serotonin Biosynthesis and Its Role in Delaying Senescence in Rice Leaves. Plant Physiol. 2009, 150, 1380–1393. [Google Scholar] [CrossRef] [Green Version]
- Ford, A.T.; Hyett, B.; Cassidy, D.; Malyon, G. The Effects of Fluoxetine on Attachment and Righting Behaviours in Marine (Gibbula Unbilicalis) and Freshwater (Lymnea stagnalis) Gastropods. Ecotoxicology 2018, 27, 477–484. [Google Scholar] [CrossRef] [Green Version]
- Mathews, S.; Reinhold, D. Biosolid-Borne Tetracyclines and Sulfonamides in Plants. Environ. Sci. Pollut. Res. 2013, 20, 4327–4338. [Google Scholar] [CrossRef]
- Turcios, A.E.; Papenbrock, J. Biofiltration of the Antibacterial Drug Sulfamethazine by the Species Chenopodium Quinoa and Its Further Biodegradation through Anaerobic Digestion. J. Environ. Sci. 2019, 75, 54–63. [Google Scholar] [CrossRef] [PubMed]
- Burken, J.G. Uptake and Metabolism of Organic Compounds: Green-Liver Model. In A Wiley-Interscience Series of Texts and Monographs; McCutcheon, S.C., Schnoor, J.L., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2003; pp. 59–84. ISBN 978-0-471-39435-8. [Google Scholar]
- Phong, V.H.N.; Koottatep, T.; Chapagain, S.K.; Panuvatvanich, A.; Polprasert, C.; Ahn, K.-H. Removal of Acetaminophen from Wastewater by Constructed Wetlands with Scirpus Validus. Environ. Eng. Res. 2016, 21, 164–170. [Google Scholar] [CrossRef]
- Phong Vo, H.N.; Le, G.K.; Hong Nguyen, T.M.; Bui, X.-T.; Nguyen, K.H.; Rene, E.R.; Vo, T.D.H.; Thanh Cao, N.-D.; Mohan, R. Acetaminophen Micropollutant: Historical and Current Occurrences, Toxicity, Removal Strategies and Transformation Pathways in Different Environments. Chemosphere 2019, 236, 124391. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Zhang, L.; Chen, J. Paracetamol in the Environment and Its Degradation by Microorganisms. Appl. Microbiol. Biotechnol. 2012, 96, 875–884. [Google Scholar] [CrossRef]
- Riemenschneider, C.; Seiwert, B.; Moeder, M.; Schwarz, D.; Reemtsma, T. Extensive Transformation of the Pharmaceutical Carbamazepine Following Uptake into Intact Tomato Plants. Environ. Sci. Technol. 2017, 51, 6100–6109. [Google Scholar] [CrossRef]
- Goldstein, M.; Malchi, T.; Shenker, M.; Chefetz, B. Pharmacokinetics in Plants: Carbamazepine and Its Interactions with Lamotrigine. Environ. Sci. Technol. 2018, 52, 6957–6964. [Google Scholar] [CrossRef]
- Huber, C.; Bartha, B.; Harpaintner, R.; Schröder, P. Metabolism of Acetaminophen (Paracetamol) in Plants—Two Independent Pathways Result in the Formation of a Glutathione and a Glucose Conjugate. Environ. Sci. Pollut. Res. 2009, 16, 206. [Google Scholar] [CrossRef] [Green Version]
- Rühmland, S.; Wick, A.; Ternes, T.A.; Barjenbruch, M. Fate of Pharmaceuticals in a Subsurface Flow Constructed Wetland and Two Ponds. Ecol. Eng. 2015, 80, 125–139. [Google Scholar] [CrossRef]
- Lam, M.W.; Young, C.J.; Mabury, S.A. Aqueous Photochemical Reaction Kinetics and Transformations of Fluoxetine. Environ. Sci. Technol. 2005, 39, 513–522. [Google Scholar] [CrossRef]
- Redshaw, C.H.; Cooke, M.P.; Talbot, H.M.; McGrath, S.; Rowland, S.J. Low Biodegradability of Fluoxetine HCl, Diazepam and Their Human Metabolites in Sewage Sludge-Amended Soil. J. Soils Sediments 2008, 8, 217–230. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Standard | Molar Mass [g/mol] | Precursor (ESI+) | Retention Time [min] |
|---|---|---|---|
| Acetaminophen | 151.16 | 152.07 | 8.34 |
| Azithromycin (dehydrate) | 748.99 (785.03) | 749.51 | 31.63 |
| Bezafibrate | 361.82 | 362.11 | 25.04 |
| Carbamazepine | 236.27 | 237.10 | 26.34 |
| Citalopram (hydrobromide) | 324.39 (405.31) | 325.17 | 28.34 |
| Fluoxetine (hydrochloride) | 309.33 (345.79) | 310.14 | 31.91 |
| Gabapentin | 171.24 | 172.13 | 9.40 |
| Gemfibrozil | 250.33 | 251.2 | 35.5 |
| Ibuprofen | 206.28 | n.d. | n.d. |
| Oxytetracycline (hydrochloride) | 460.43 (496.90) | n.d. | n.d. |
| Propranolol (hydrochloride) | 259.34 (295.80) | 260.17 | 27.48 |
| Sulfadoxin | 310.33 | 311.08 | 8.25 |
| Sulfamethazine | 278.33 | 279.16 | 12.63 |
| Sulfathiazole | 255.32 | 256.02 | 8.52 |
| Tetracycline (hydrochloride) | 444.44 (480.90) | n.d. | n.d. |
| Topiramate | 339.36 | 357.13 | 21.26 |
| Venlafaxine (hydrochloride) | 277.40 (313.86) | 278.2 | 27.83 |
| Xenobiotic | Xenobiotic Concentration [ng L−1] | ||
|---|---|---|---|
| Seixal | Rosario | Alcochete | |
| Acetaminophen | 81.0 | 6.04 | <LOD |
| Azithromycin dhydrate | <LOD | <LOD | <LOD |
| Bezafibrate | <LOD | <LOD | <LOD |
| Carbamazepine | 5.86 | <LOD | <LOD |
| Citalopram hydrobromide | <LOD | <LOD | <LOD |
| Fluoxetine hydrochloride | 88.46 | 68.16 | 68.28 |
| Gabapentin | <LOD | <LOD | <LOD |
| Propranolol hydrochloride | <LOD | <LOD | <LOD |
| Sulfadoxin | <LOD | <LOD | <LOD |
| Sulfamethazin | <LOD | <LOD | <LOD |
| Sulfathiazole | <LOD | <LOD | <LOD |
| Topiramate | <LOD | <LOD | <LOD |
| Venlafaxine hydrochloride | 25.8 | 13.68 | <LOD |
| Shoots | Initial Concentration (mg L−1) | |||
|---|---|---|---|---|
| Xenobiotic | 5 (Singly) | 2 (Singly) | 1 (Mixture) | 0.4 (Mixture) |
| Sulfamethazine | 1.58 ± 0.30 C | 1.09 ± 0.15 C | 0.47 ± 0.05 C | 0.53 ± 0.07 C |
| Acetaminophen | 1.19 ± 0.44 C | 0.33 ± 0.01 C | 1.42 ± 0.26 C | 1.93 ± 0.31 C |
| Carbamazepine | 13.57 ± 0.71 A | 6.98 ± 1.85 B | 4.99 ± 0.18 B | 2.23 ± 0.83 C |
| Venlafaxine | 0.59 ± 0.22 C | 0.47 ± 0.08 C | 0.34 ± 0.07 C | 0.09 ± 0.03 C |
| Fluoxetine | 0.20 ± 0.02 C | 0.13 ± 0.01 C | 0.03 ± 0.00 C | 0.02 ± 0.00 C |
| Roots | Initial Concentration (mg L−1) | |||
|---|---|---|---|---|
| Xenobiotic | 5 (Singly) | 2 (Singly) | 1 (Mixture) | 0.4 (Mixture) |
| Sulfamethazine | 68.96 ± 3.28 A | 30.20 ± 1.56 B | 12.66 ± 1.65 C | 6.41 ± 0.64 DE |
| Acetaminophen | 9.74 ± 1.74 CD | 7.37 ± 0.41 CDE | 6.42 ± 0.69 DE | 5.40 ± 0.16 DEF |
| Carbamazepine | 2.14 ± 0.35 EFG | 0.49 ± 0.11 FG | 0.40 ± 0.09 FG | 0.17 ± 0.01 FG |
| Venlafaxine | 0.28 ± 0.21 FG | 0.13 ± 0.03 FG | 0.05 ± 0.03 FG | 0.04 ± 0.01 FG |
| Fluoxetine | 0.23 ± 0.13 FG | 0.06 ± 0.00 FG | 0.00 ± 0.00 G | 0.01 ± 0.00 FG |
| Xenobiotic | Single Treatment | Mixture of XBs | ||
|---|---|---|---|---|
| Not Denatured | Denatured | Not Denatured | Denatured | |
| Sulfamethazine | 1.4969 ± 0.12 c | 5.4227 ± 0.18 a | 0.7748 ± 0.14 d | 4.1400 ± 0.08 b |
| Acetaminophen | 3.8860 ± 0.67 a | 4.2600 ± 0.76 a | 5.0660 ± 0.34 a | 4.4993 ± 0.70 a |
| Carbamazepine | 7.0553 ± 0.07 a | 7.0927 ± 0.03 a | 6.7460 ± 0.11 a | 6.7933 ± 0.10 a |
| Venlafaxine HCl | 0.0397 ± 9.4 × 10−5 a | 0.0341 ± 9.7 × 10−4 b | 0.0368 ± 1.2 × 10−3 ab | 0.0391 ± 1.4 × 10−3 a |
| Fluoxetine HCl | 0.0205 ± 2.2 × 10−4 ab | 0.0207 ± 9.6 × 10−5 ab | 0.0201 ± 8.5 × 10−4 b | 0.0225 ± 3.4 × 10−4 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Turcios, A.E.; Hielscher, M.; Duarte, B.; Fonseca, V.F.; Caçador, I.; Papenbrock, J. Screening of Emerging Pollutants (EPs) in Estuarine Water and Phytoremediation Capacity of Tripolium pannonicum under Controlled Conditions. Int. J. Environ. Res. Public Health 2021, 18, 943. https://doi.org/10.3390/ijerph18030943
Turcios AE, Hielscher M, Duarte B, Fonseca VF, Caçador I, Papenbrock J. Screening of Emerging Pollutants (EPs) in Estuarine Water and Phytoremediation Capacity of Tripolium pannonicum under Controlled Conditions. International Journal of Environmental Research and Public Health. 2021; 18(3):943. https://doi.org/10.3390/ijerph18030943
Chicago/Turabian StyleTurcios, Ariel E., Marie Hielscher, Bernardo Duarte, Vanessa F. Fonseca, Isabel Caçador, and Jutta Papenbrock. 2021. "Screening of Emerging Pollutants (EPs) in Estuarine Water and Phytoremediation Capacity of Tripolium pannonicum under Controlled Conditions" International Journal of Environmental Research and Public Health 18, no. 3: 943. https://doi.org/10.3390/ijerph18030943