Oxidative Stress and Gene Expression Modifications Mediated by Extracellular Vesicles: An In Vivo Study of the Radiation-Induced Bystander Effect


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Irradiation and Sample Collection
2.3. EV Isolation, Characterisation and Bystander Experiment
2.4. Lipid Peroxidation
2.5. Plasma Oxidative Damage
2.6. Gene Expression
2.7. Statistical Analyses
3. Results
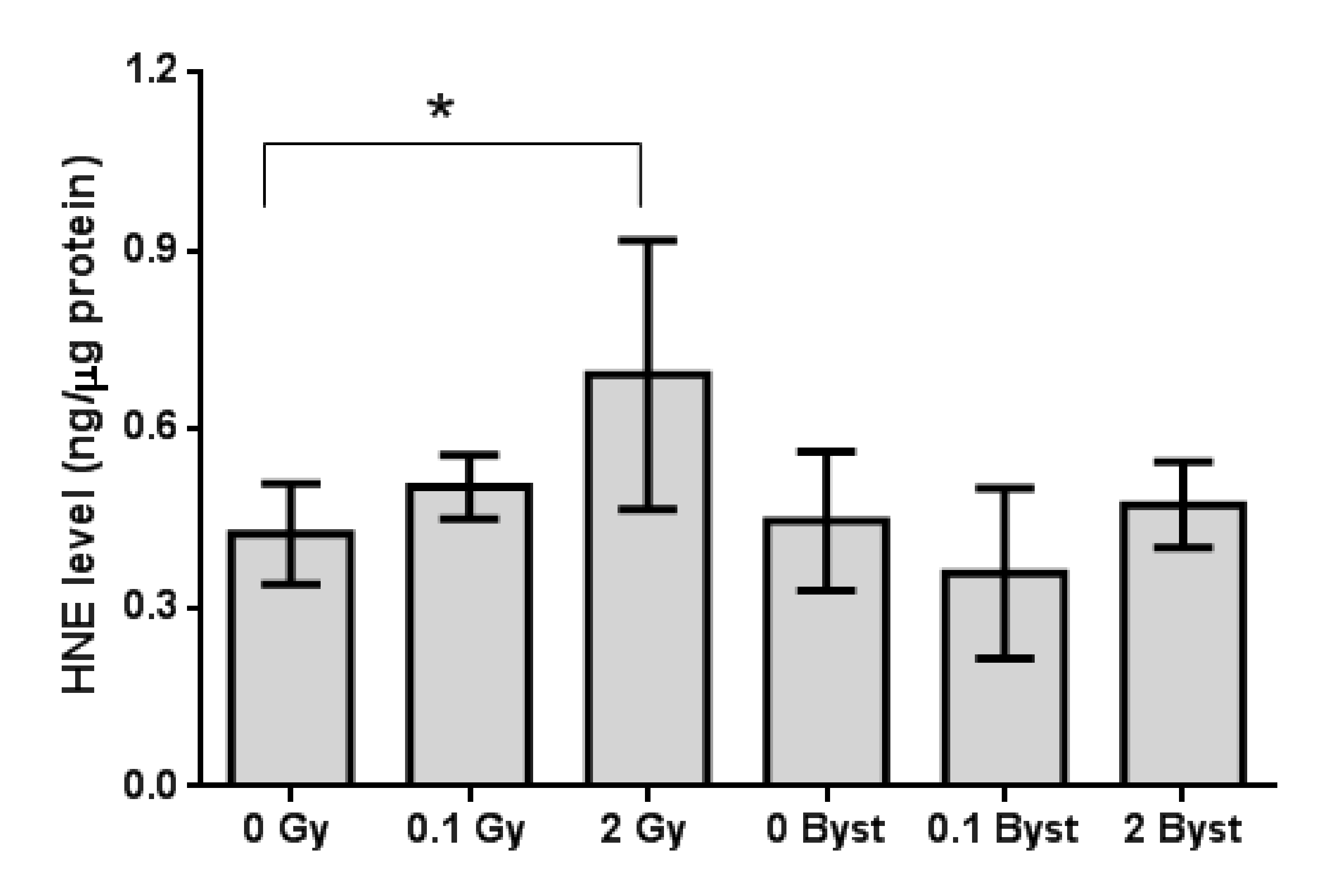
3.1. Effects of Direct Irradiation and EV-Transfer from Irradiated Mice on Lipid Peroxidation in the Spleen
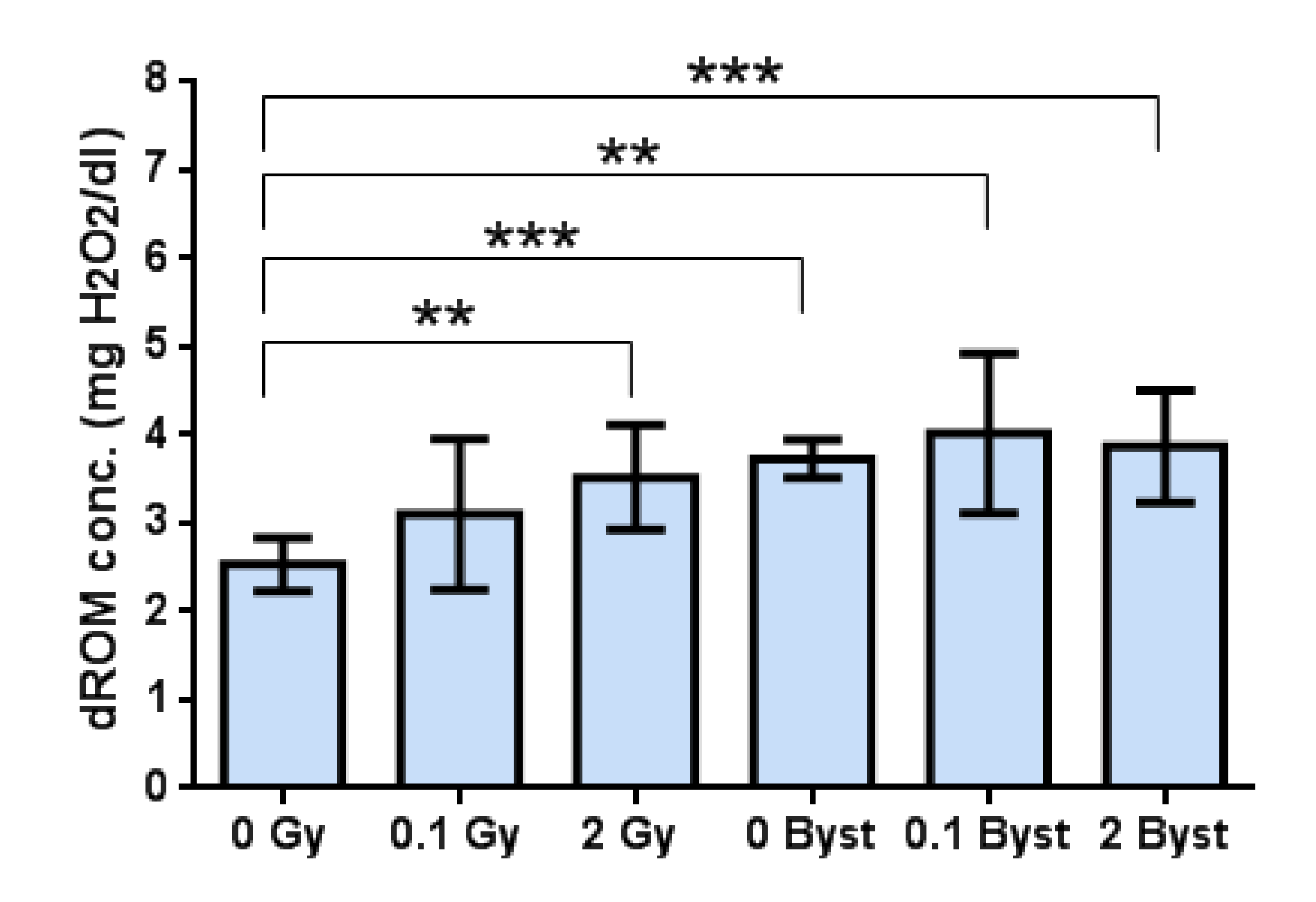
3.2. Effects of Direct Irradiation and EV-Transfer from Irradiated Mice on the Plasma Level of Oxidative Damage
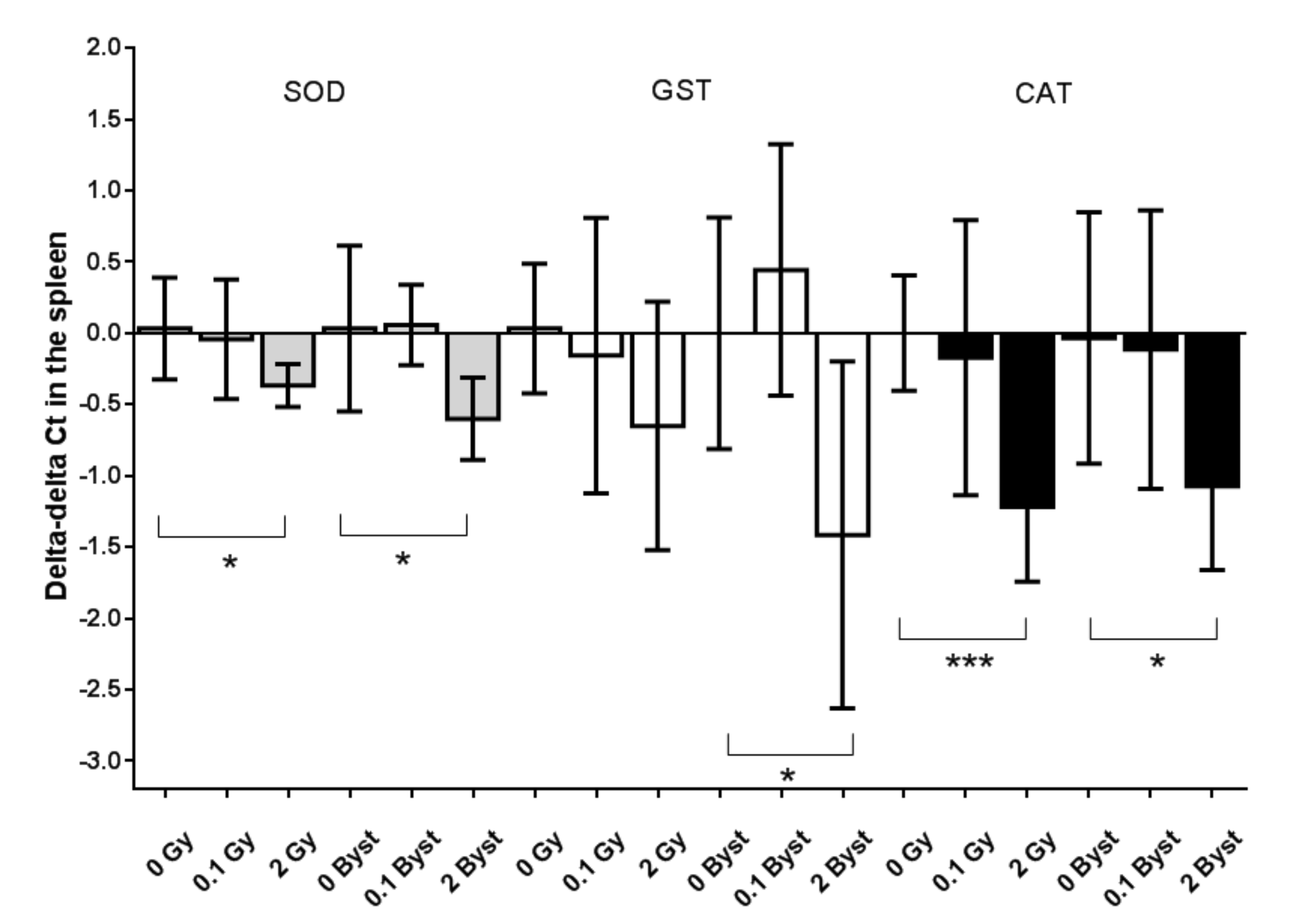
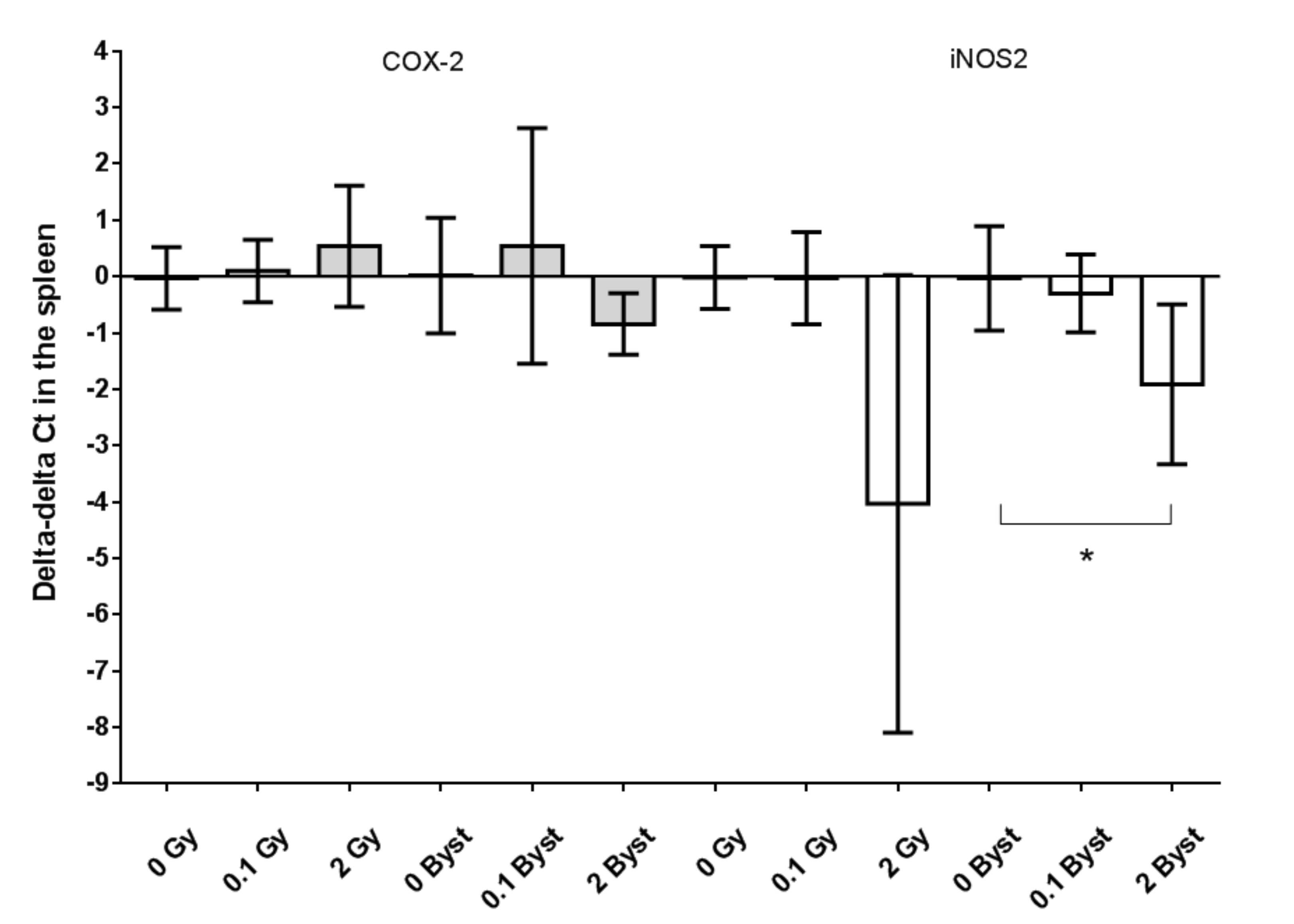
3.3. Effects of Direct Irradiation and EV Transfer from Irradiated Mice on the Antioxidant and Pro-Oxidant Gene Expressions
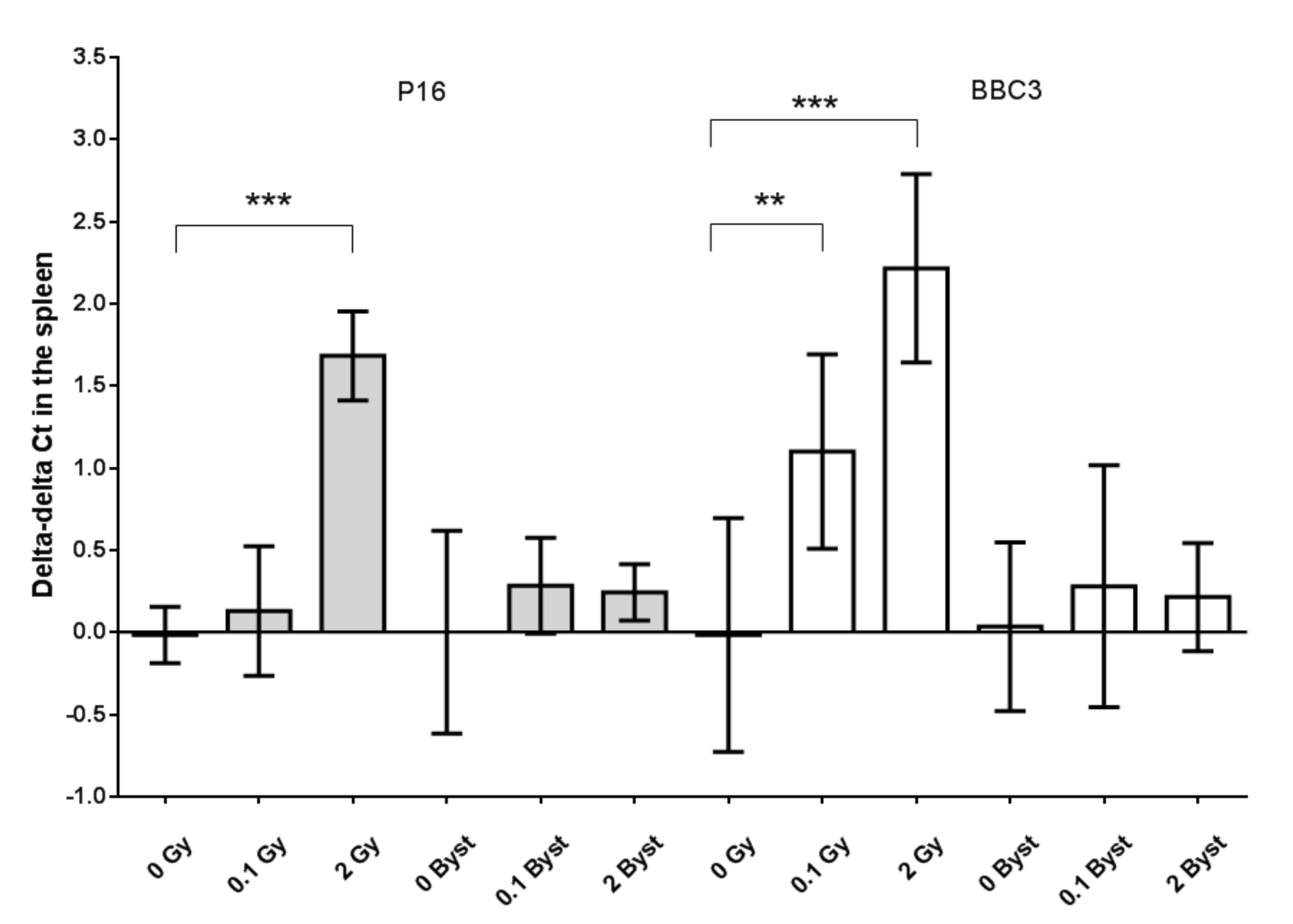
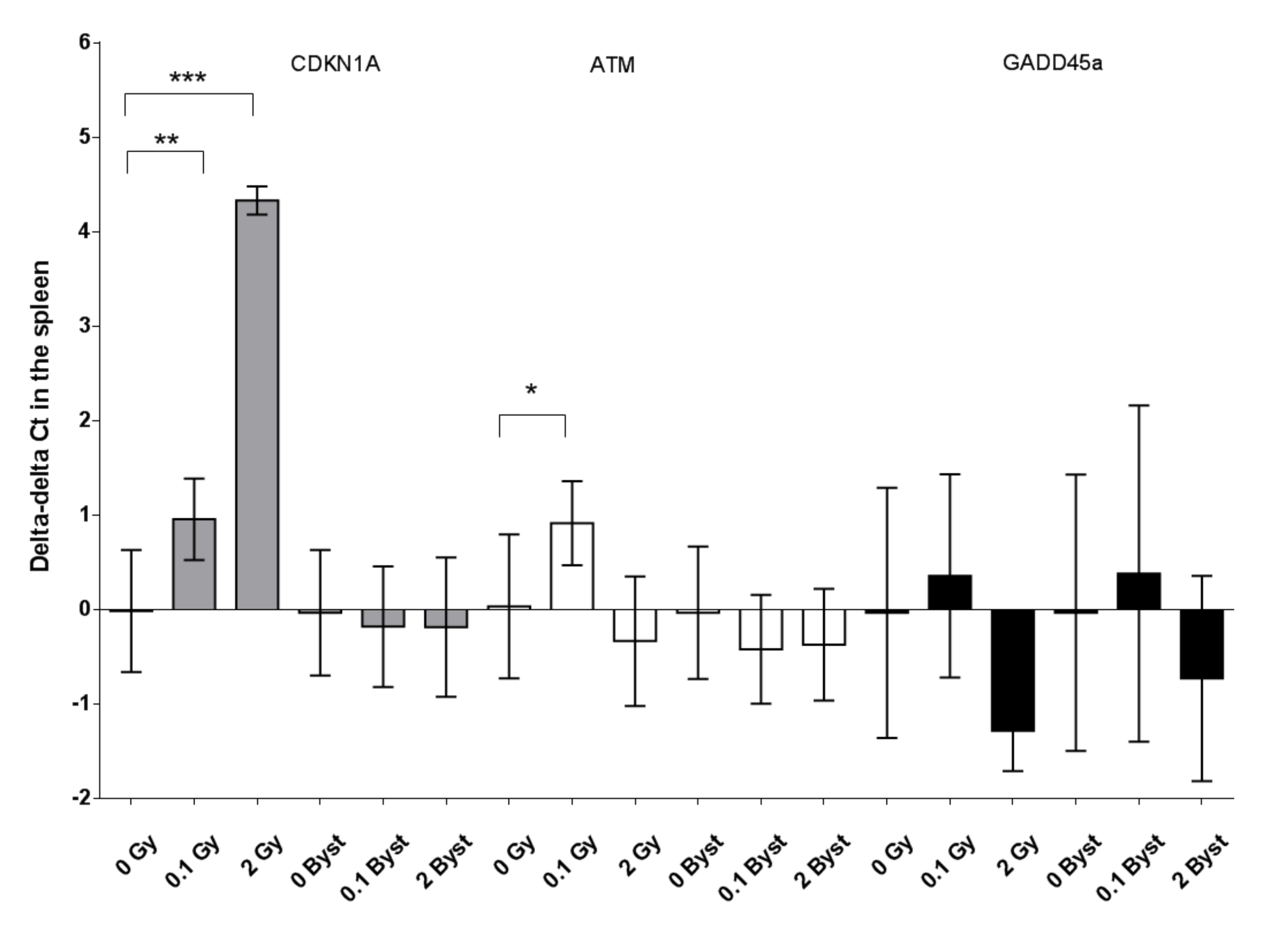
3.4. Effects of Direct Irradiation and EV Transfer from Irradiated Mice on the Cell Cycle, Senescence and Apoptosis Regulator Gene Expression
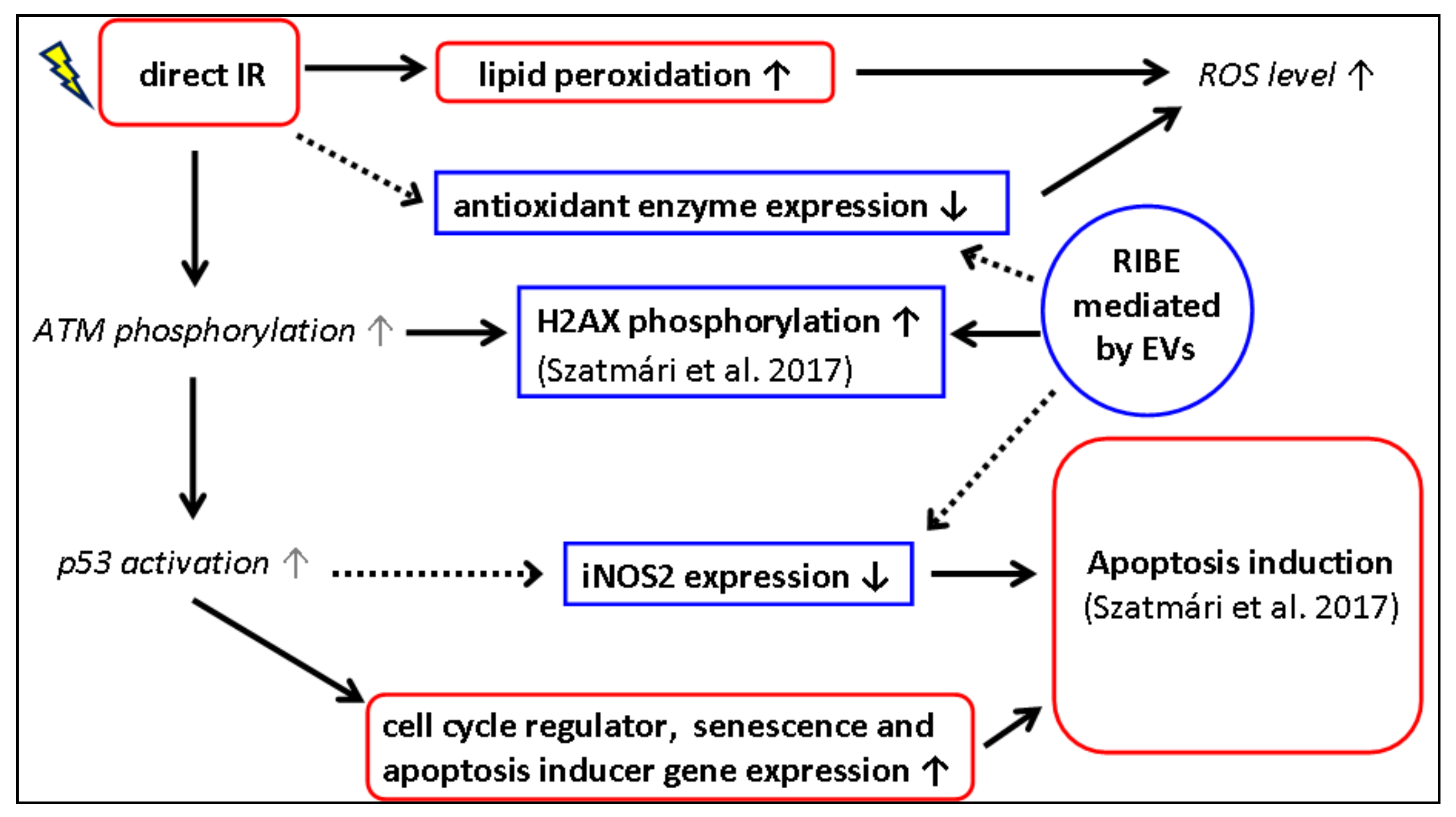
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Clutton, S.; Townsend, K.; Walker, C.; Ansell, J.; Wright, E. Radiation-induced genomic instability and persisting oxidative stress in primary bone marrow cultures. Carcinogenesis 1996, 17, 1633–1639. [Google Scholar] [CrossRef] [PubMed]
- Azzam, E.I.; Jay-Gerin, J.-P.; Pain, D. Ionizing radiation-induced metabolic oxidative stress and prolonged cell injury. Cancer Lett. 2012, 327, 48–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reisz, J.A.; Bansal, N.; Qian, J.; Zhao, W.; Furdui, C.M. Effects of ionizing radiation on biological molecules—mechanisms of damage and emerging methods of detection. Antioxid. Redox Signal. 2014, 21, 260–292. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Robbins, M.E. Inflammation and chronic oxidative stress in radiation-induced late normal tissue injury: Therapeutic implications. Curr. Med. Chem. 2009, 16, 130–143. [Google Scholar] [CrossRef]
- Buonanno, M.; De Toledo, S.M.; Pain, D.; Azzam, E.I. Long-term consequences of radiation-induced bystander effects depend on radiation quality and dose and correlate with oxidative stress. Radiat. Res. 2011, 175, 405–415. [Google Scholar] [CrossRef] [Green Version]
- Devi, G.S.; Prasad, M.H.; Saraswathi, I.; Raghu, D.; Rao, D.; Reddy, P. Free radicals antioxidant enzymes and lipid peroxidation in different types of leukemias. Clin. Chim. Acta 2000, 293, 53–62. [Google Scholar] [CrossRef]
- Flora, S. Role of free radicals and antioxidants in health and disease. Cell. Mol. Biol. 2007, 53, 1–2. [Google Scholar]
- Kryston, T.B.; Georgiev, A.B.; Pissis, P.; Georgakilas, A.G. Role of oxidative stress and DNA damage in human carcinogenesis. Mutat. Res. Fundam. Mol. Mech. Mutagenesis 2011, 711, 193–201. [Google Scholar] [CrossRef]
- Georgakilas, A.G. Oxidative stress, DNA damage and repair in carcinogenesis: Have we established a connection? Cancer Lett. 2012, 327, 3–4. [Google Scholar] [CrossRef]
- Sjostedt, S.; Bezak, E. Non-targeted effects of ionising radiation and radiotherapy. Australas. Phys. Eng. Sci. Med. 2010, 33, 219–231. [Google Scholar] [CrossRef]
- Tamminga, J.; Kovalchuk, O. Role of DNA damage and epigenetic DNA methylation changes in radiation-induced genomic instability and bystander effects in germline in vivo. Curr. Mol. Pharmacol 2011, 4, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Nikitaki, Z.; Mavragani, I.V.; Laskaratou, D.A.; Gika, V.; Moskvin, V.P.; Theofilatos, K.; Vougas, K.; Stewart, R.D.; Georgakilas, A.G. Systemic mechanisms and effects of ionizing radiation: A new old paradigm of how the bystanders and distant can become the players. Semin. Cancer Biol. 2016, 37–38, 77–95. [Google Scholar] [CrossRef] [PubMed]
- Morgan, W.F.; Sowa, M.B. Non-targeted effects induced by ionizing radiation: Mechanisms and potential impact on radiation induced health effects. Cancer Lett. 2015, 356, 17–21. [Google Scholar] [CrossRef] [Green Version]
- Lyng, F.; Seymour, C.; Mothersill, C. Production of a signal by irradiated cells which leads to a response in unirradiated cells characteristic of initiation of apoptosis. Br. J. Cancer 2000, 83, 1223–1230. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Randers-Pherson, G.; Suzuki, M.; Waldren, C.A.; K. Hei, T. Genotoxic damage in non-irradiated cells: Contribution from the bystander effect. Radiat. Prot. Dosim. 2002, 99, 227–232. [Google Scholar] [CrossRef] [Green Version]
- Nagasawa, H.; Little, J.B. Bystander effect for chromosomal aberrations induced in wild-type and repair deficient CHO cells by low fluences of alpha particles. Mutat. Res. Fundam. Mol. Mech. Mutagenesis 2002, 508, 121–129. [Google Scholar] [CrossRef]
- Zhou, H.; Ivanov, V.N.; Gillespie, J.; Geard, C.R.; Amundson, S.A.; Brenner, D.J.; Yu, Z.; Lieberman, H.B.; Hei, T.K. Mechanism of radiation-induced bystander effect: Role of the cyclooxygenase-2 signaling pathway. Proc. Natl. Acad. Sci. USA 2005, 102, 14641–14646. [Google Scholar] [CrossRef] [Green Version]
- Koturbash, I.; Rugo, R.; Hendricks, C.; Loree, J.; Thibault, B.; Kutanzi, K.; Pogribny, I.; Yanch, J.; Engelward, B.; Kovalchuk, O. Irradiation induces DNA damage and modulates epigenetic effectors in distant bystander tissue in vivo. Oncogene 2006, 25, 4267–4275. [Google Scholar] [CrossRef] [Green Version]
- Koturbash, I.; Loree, J.; Kutanzi, K.; Koganow, C.; Pogribny, I.; Kovalchuk, O. In vivo bystander effect: Cranial X-irradiation leads to elevated DNA damage, altered cellular proliferation and apoptosis, and increased p53 levels in shielded spleen. Radiat. Oncol. Biol. Phys. 2008, 70, 554–562. [Google Scholar] [CrossRef]
- Ilnytskyy, Y.; Koturbash, I.; Kovalchuk, O. Radiation-induced bystander effects in vivo are epigenetically regulated in a tissue-specific manner. Environ. Mol. Mutagenesis 2009, 50, 105–113. [Google Scholar] [CrossRef]
- Hatzi, V.I.; Laskaratou, D.A.; Mavragani, I.V.; Nikitaki, Z.; Mangelis, A.; Panayiotidis, M.I.; Pantelias, G.E.; Terzoudi, G.I.; Georgakilas, A.G. Non-targeted radiation effects in vivo: A critical glance of the future in radiobiology. Cancer Lett. 2015, 356, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Lyng, F.M.; Maguire, P.; McClean, B.; Seymour, C.; Mothersill, C. The involvement of calcium and MAP kinase signaling pathways in the production of radiation-induced bystander effects. Radiat. Res. 2006, 165, 400–409. [Google Scholar] [CrossRef] [PubMed]
- Dickey, J.S.; Zemp, F.J.; Martin, O.A.; Kovalchuk, O. The role of miRNA in the direct and indirect effects of ionizing radiation. Radiat. Environ. Biophys. 2011, 50, 491. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Du, S.; Liu, L.; Gan, F.; Jiang, X.; Wangrao, K.; Lyu, P.; Gong, P.; Yao, Y. Radiation-Induced Bystander Effect can be Transmitted Through Exosomes Using miRNAs as Effector Molecules. Radiat. Res. 2020, 89–100. [Google Scholar]
- Szatmári, T.; Hargitai, R.; Sáfrány, G.; Lumniczky, K. Extracellular vesicles in modifying the effects of ionizing radiation. Int. J. Mol. Sci. 2019, 20, 5527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shao, C.; Stewart, V.; Folkard, M.; Michael, B.D.; Prise, K.M. Nitric oxide-mediated signaling in the bystander response of individually targeted glioma cells. Cancer Res. 2003, 63, 8437–8442. [Google Scholar]
- Carruba, G.; Webber, M.M.; Quader, S.T.; Amoroso, M.; Cocciadiferro, L.; Saladino, F.; Trosko, J.E.; Castagnetta, L.A. Regulation of cell-to-cell communication in non-tumorigenic and malignant human prostate epithelial cells. Prostate 2002, 50, 73–82. [Google Scholar] [CrossRef]
- Szatmári, T.; Kis, D.; Bogdándi, E.N.; Benedek, A.; Bright, S.; Bowler, D.; Persa, E.; Kis, E.; Balogh, A.; Naszályi, L.N.; et al. Extracellular vesicles mediate radiation-induced systemic bystander signals in the bone marrow and spleen. Front. Immunol. 2017, 8, 347. [Google Scholar]
- Ariyoshi, K.; Miura, T.; Kasai, K.; Fujishima, Y.; Nakata, A.; Yoshida, M. Radiation-induced bystander effect is mediated by mitochondrial DNA in exosome-like vesicles. Sci. Rep 2019, 9, 1–14. [Google Scholar] [CrossRef] [Green Version]
- György, B.; Szabó, T.G.; Pásztói, M.; Pál, Z.; Misják, P.; Aradi, B.; László, V.; Pállinger, E.; Pap, E.; Kittel, Á.; et al. Membrane vesicles, current state-of-the-art: Emerging role of extracellular vesicles. Cell. Mol. Life Sci. 2011, 68, 2667–2688. [Google Scholar]
- Valadi, H.; Ekström, K.; Bossios, A.; Sjöstrand, M.; Lee, J.J.; Lötvall, J.O. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Mayah, A.H.; Irons, S.L.; Pink, R.C.; Carter, D.R.; Kadhim, M.A. Possible role of exosomes containing RNA in mediating nontargeted effect of ionizing radiation. Radiat. Res 2012, 177, 539–545. [Google Scholar] [CrossRef] [PubMed]
- Szatmári, T.; Persa, E.; Kis, E.; Benedek, A.; Hargitai, R.; Sáfrány, G.; Lumniczky, K. Extracellular vesicles mediate low dose ionizing radiation-induced immune and inflammatory responses in the blood. Int. J. Radiat. Biol. 2019, 95, 12–22. [Google Scholar] [CrossRef] [PubMed]
- Hinzman, C.P.; Baulch, J.E.; Mehta, K.Y.; Girgis, M.; Bansal, S.; Gill, K.; Li, Y.X.; Limon, C.L.; Cheema, A.K. Plasma-derived extracellular vesicles yield predictive markers of cranial irradiation exposure in mice. Sci. Rep. 2019, 9. [Google Scholar] [CrossRef] [PubMed]
- Jabbari, N.; Karimipour, M.; Khaksar, M.; Akbariazar, E.; Heidarzadeh, M.; Mojarad, B.; Aftab, H.; Rahbarghazi, R.; Rezaie, J. Tumor-derived extracellular vesicles: Insights into bystander effects of exosomes after irradiation. Laser Med. Sci. 2020, 35, 531–545. [Google Scholar] [CrossRef]
- Xu, S.; Wang, J.F.; Ding, N.; Hu, W.T.; Zhang, X.R.; Wang, B.; Hua, J.R.; Wei, W.J.; Zhu, Q.Y. Exosome-mediated microRNA transfer plays a role in radiation-induced bystander effect. RNA Biol. 2015, 12, 1355–1363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petkau, A. Role of superoxide dismutase in modification of radiation injury. Br. J. Cancer Suppl. 1987, 8, 87–95. [Google Scholar]
- Limón-Pacheco, J.; Gonsebatt, M.E. The role of antioxidants and antioxidant-related enzymes in protective responses to environmentally induced oxidative stress. Mutat. Res. Genet. Toxicol. Environ. Mutagenesis 2009, 674, 137–147. [Google Scholar] [CrossRef]
- Jagetia, G.C.; Rajanikant, G.K.; Rao, S.K.; Baliga, M.S. Alteration in the glutathione, glutathione peroxidase, superoxide dismutase and lipid peroxidation by ascorbic acid in the skin of mice exposed to fractionated gamma radiation. Clin. Chim. Acta 2003, 332, 111–121. [Google Scholar] [CrossRef]
- Yang, H.Y.; Asaad, N.; Held, K.D. Medium-mediated intercellular communication is involved in bystander responses of X-ray-irradiated normal human fibroblasts. Oncogene 2005, 24, 2096–2103. [Google Scholar] [CrossRef] [Green Version]
- Strange, R.C.; Spiteri, M.A.; Ramachandran, S.; Fryer, A.A. Glutathione-S-transferase family of enzymes. Mutat. Res. Mol. Mech. Mutagen. 2001, 482, 21–26. [Google Scholar] [CrossRef]
- Li, M.J.; Wang, W.W.; Chen, S.W.; Shen, Q.; Min, R. Radiation dose effect of DNA repair-related gene expression in mouse white blood cells. Med. Sci. Monit. 2011, 17, Br290–Br297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, C.M.; Park, K.P.; Song, J.E.; Jeoung, D.I.; Cho, C.K.; Kim, T.H.; Bae, S.; Lee, S.J.; Lee, Y.S. Possible biomarkers for ionizing radiation exposure in human peripheral blood lymphocytes. Radiat. Res. 2003, 159, 312–319. [Google Scholar] [CrossRef]
- Kabacik, S.; Mackay, A.; Tamber, N.; Manning, G.; Finnon, P.; Paillier, F.; Ashworth, A.; Bouffler, S.; Badie, C. Gene expression following ionising radiation: Identification of biomarkers for dose estimation and prediction of individual response. Int. J. Radiat. Biol. 2011, 87, 115–129. [Google Scholar] [CrossRef]
- Han, J.W.; Flemington, C.; Houghton, A.B.; Gu, Z.M.; Zambetti, G.P.; Lutz, R.J.; Zhu, L.; Chittenden, T. Expression of bbc3, a pro-apoptotic BH3-only gene, is regulated by diverse cell death and survival signals. Proc. Natl. Acad. Sci. USA 2001, 98, 11318–11323. [Google Scholar] [CrossRef] [Green Version]
- Dutto, I.; Tillhon, M.; Cazzalini, O.; Stivala, L.A.; Prosperi, E. Biology of the cell cycle inhibitor p21(CDKN1A): Molecular mechanisms and relevance in chemical toxicology. Arch. Toxicol. 2015, 89, 155–178. [Google Scholar] [CrossRef]
- Xiong, Y.; Hannon, G.J.; Zhang, H.; Casso, D.; Kobayashi, R.; Beach, D. P21 Is a Universal Inhibitor of Cyclin Kinases. Nature 1993, 366, 701–704. [Google Scholar] [CrossRef]
- Kastan, M.B.; Zhan, Q.M.; Eldeiry, W.S.; Carrier, F.; Jacks, T.; Walsh, W.V.; Plunkett, B.S.; Vogelstein, B.; Fornace, A.J. A Mammalian-Cell Cycle Checkpoint Pathway Utilizing P53 and Gadd45 Is Defective in Ataxia-Telangiectasia. Cell 1992, 71, 587–597. [Google Scholar] [CrossRef]
- Shrivastav, M.; De Haro, L.P.; Nickoloff, J.A. Regulation of DNA double-strand break repair pathway choice. Cell Res. 2008, 18, 134–147. [Google Scholar] [CrossRef] [Green Version]
- Gately, D.P.; Hittle, J.C.; Chan, G.K.T.; Yen, T.J. Characterization of ATM expression, localization, and associated DNA-dependent protein kinase activity. Mol. Biol. Cell 1998, 9, 2361–2374. [Google Scholar] [CrossRef] [Green Version]
- Collado, M.; Blasco, M.A.; Serrano, M. Cellular senescence in cancer and aging. Cell 2007, 130, 223–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baker, D.J.; Jin, F.; van Deursen, J.M. The yin and yang of the Cdkn2a locus in senescence and aging. Cell Cycle 2008, 7, 2795–2802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campisi, J. Senescent cells, tumor suppression, and organismal aging: Good citizens, bad neighbors. Cell 2005, 120, 513–522. [Google Scholar] [CrossRef] [PubMed]
- Campisi, J.; di Fagagna, F.D. Cellular senescence: When bad things happen to good cells. Nat. Rev. Mol. Cell Biol. 2007, 8, 729–740. [Google Scholar] [CrossRef] [PubMed]
- Chai, Y.; Lam, R.; Calaf, G.; Zhou, H.; Amundson, S.; Hei, T. Radiation-induced non-targeted response in vivo: Role of the TGFβ-TGFBR1-COX-2 signalling pathway. Br. J. Cancer 2013, 108, 1106–1112. [Google Scholar] [CrossRef] [Green Version]
- Deeb, R.S.; Upmacis, R.K.; Lamon, B.D.; Gross, S.S.; Hajjar, D.P. Maintaining equilibrium by selective targeting of cyclooxygenase pathways - Promising offensives against vascular injury. Hypertension 2008, 51, 1–7. [Google Scholar] [CrossRef]
- Meziani, F.; Tesse, A.; David, E.; Martinez, M.C.; Wangesteen, R.; Schneider, F.; Andriantsitohaina, R. Shed membrane particles from preeclamptic women generate vascular wall inflammation and blunt vascular contractility. Am. J. Pathol. 2006, 169, 1473–1483. [Google Scholar] [CrossRef] [Green Version]
- Lorimore, S.A.; Coates, P.J.; Scobie, G.E.; Milne, G.; Wright, E.G. Inflammatory-type responses after exposure to ionizing radiation in vivo: A mechanism for radiation-induced bystander effects? Oncogene 2001, 20, 7085–7095. [Google Scholar] [CrossRef] [Green Version]
- MacMicking, J.; Xie, Q.W.; Nathan, C. Nitric oxide and macrophage function. Annu. Rev. Immunol. 1997, 15, 323–350. [Google Scholar] [CrossRef]
- Sawal, H.A.; Asghar, K.; Bureik, M.; Jalal, N. Bystander signaling via oxidative metabolism. Oncotargets Ther 2017, 10, 3925–3940. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, H.; Hayashi, S.; Hatashita, M.; Ohnishi, K.; Shioura, H.; Ohtsubo, T.; Kitai, R.; Ohnishi, T.; Kano, E. Induction of radioresistance by a nitric oxide-mediated bystander effect. Radiat. Res. 2001, 155, 387–396. [Google Scholar] [CrossRef]
- Azzam, E.I.; De Toledo, S.M.; Spitz, D.R.; Little, J.B. Oxidative metabolism modulates signal transduction and micronucleus formation in bystander cells from alpha-particle-irradiated normal human fibroblast cultures. Cancer Res. 2002, 62, 5436–5442. [Google Scholar] [PubMed]
- Havaki, S.; Kotsinas, A.; Chronopoulos, E.; Kletsas, D.; Georgakilas, A.; Gorgoulis, V.G. The role of oxidative DNA damage in radiation induced bystander effect. Cancer Lett. 2015, 356, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Dutta, S.; Warshall, C.; Bandyopadhyay, C.; Dutta, D.; Chandran, B. Interactions between Exosomes from Breast Cancer Cells and Primary Mammary Epithelial Cells Leads to Generation of Reactive Oxygen Species Which Induce DNA Damage Response, Stabilization of p53 and Autophagy in Epithelial Cells. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larson, M.C.; Hillery, C.A.; Hogg, N. Circulating membrane-derived microvesicles in redox biology. Free Radic. Biol. Med. 2014, 73, 214–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, M.; Drwal, G.; Bourgeois, T.; Saltz, J.; Wu, H.F.M. Distinct proteome features of plasma microparticles. Proteomics 2005, 5, 1940–1952. [Google Scholar] [CrossRef]
- Horn, P.; Cortese-Krott, M.M.; Amabile, N.; Hundsdoerfer, C.; Kroencke, K.D.; Kelm, M.; Heiss, C. Circulating Microparticles Carry a Functional Endothelial Nitric Oxide Synthase That Is Decreased in Patients With Endothelial Dysfunction. J. Am. Heart Assoc. 2013, 2, e003764. [Google Scholar] [CrossRef] [Green Version]
- Janiszewski, M.; do Carmo, A.O.; Pedro, M.A.; Silva, E.; Knobel, E.; Laurindo, F.R. Platelet-derived exosomes of septic individuals possess proapoptotic NAD (P) H oxidase activity: A novel vascular redox pathway. Crit. Care Med. 2004, 32, 818–825. [Google Scholar] [CrossRef]
- Miguet, L.; Pacaud, K.; Felden, C.; Hugel, B.; Martinez, M.; Freyssinet, J.; Herbrecht, R.; Potier, N.; van Dorsselaer, A.; Mauvieux, L. Proteomic analysis of malignant lymphocyte membrane microparticles using double ionization coverage optimization. Proteomics 2006, 6, 153. [Google Scholar] [CrossRef]
- Peterson, D.B.; Sander, T.; Kaul, S.; Wakim, B.T.; Halligan, B.; Twigger, S.; Pritchard, K.A., Jr.; Oldham, K.T.; Ou, J.S. Comparative proteomic analysis of PAI-1 and TNF-alpha-derived endothelial microparticles. Proteomics 2008, 8, 2430–2446. [Google Scholar] [CrossRef] [Green Version]
- Ermakov, A.V.; Konkova, M.S.; Kostyuk, S.V.; Izevskaya, V.L.; Baranova, A.; Veiko, N.N. Oxidized Extracellular DNA as a Stress Signal in Human Cells. Oxidative Med. Cell. Longev. 2013, 2013, 649747. [Google Scholar] [CrossRef] [PubMed]
- Elzanowska, J.; Semira, C.; Costa-Silva, B. DNA in extracellular vesicles: Biological and clinical aspects. Mol. Oncol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Jansen, F.; Yang, X.Y.; Franklin, B.S.; Hoelscher, M.; Schmitz, T.; Bedorf, J.; Nickenig, G.; Werner, N. High glucose condition increases NADPH oxidase activity in endothelial microparticles that promote vascular inflammation. Cardiovasc. Res. 2013, 98, 94–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pilch, D.R.; Sedelnikova, O.A.; Redon, C.; Celeste, A.; Nussenzweig, A.; Bonner, W.M. Characteristics of γ-H2AX foci at DNA double-strand breaks sites. Biochem. Cell Biol. 2003, 81, 123–129. [Google Scholar] [CrossRef]
- Rogakou, E.P.; Pilch, D.R.; Orr, A.H.; Ivanova, V.S.; Bonner, W.M. DNA double-stranded breaks induce histone H2AX phosphorylation on serine 139. J. Biol. Chem. 1998, 273, 5858–5868. [Google Scholar] [CrossRef] [Green Version]
- Sokolov, M.V.; Dickey, J.S.; Bonner, W.M.; Martin, O.A. γH2AX in Bystander Cells: Not Just a Radiation-Triggered Event, a Cellular Response to Stress Mediated by Intercellular Communication. Cell Cycle 2007, 6, 2210–2212. [Google Scholar] [CrossRef] [Green Version]
- Kis, D.; Persa, E.; Szatmari, T.; Antal, L.; Bota, A.; Csordas, I.B.; Hargitai, R.; Jezso, B.; Kis, E.; Mihaly, J.; et al. The effect of ionising radiation on the phenotype of bone marrow-derived extracellular vesicles. Br. J. Radiol. 2020, 93. [Google Scholar] [CrossRef]
- Bonisoli-Alquati, A.; Mousseau, T.A.; Moller, A.P.; Caprioli, M.; Saino, N. Increased oxidative stress in barn swallows from the Chernobyl region. Comp. Biochem. Phys. A 2010, 155, 205–210. [Google Scholar] [CrossRef]
- Sato, T.; Kinoshita, M.; Yamamoto, T.S.; Ito, M.; Nishida, T.; Takeuchi, M.; Saitoh, D.Z.; Seki, S.H.; Mukai, Y.S. Treatment of Irradiated Mice with High-Dose Ascorbic Acid Reduced Lethality. PLoS ONE 2015, 10. [Google Scholar] [CrossRef]
- Costantini, D. Oxidative stress ecology and the d-ROMs test: Facts, misfacts and an appraisal of a decade’s work. Behav. Ecol. Sociobiol. 2016, 70, 809–820. [Google Scholar] [CrossRef] [Green Version]
- Haghdoost, S.; Czene, S.; Naslund, I.; Skog, S.; Harms-Ringdahl, M. Extracellular 8-oxo-dG as a sensitive parameter for oxidative stress in vivo and in vitro. Free Radic. Res 2005, 39, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Barshishat-Kupper, M.; Tipton, A.J.; McCart, E.A.; McCue, J.; Mueller, G.P.; Day, R.M. Effect of ionizing radiation on liver protein oxidation and metabolic function in C57BL/6J mice. Int. J. Radiat. Biol. 2014, 90, 1169–1178. [Google Scholar] [CrossRef] [PubMed]
- El-Din, A.A.M.; Abdelrazzak, A.B.; Ahmed, M.T.; El-Missiry, M.A. Radiation induced bystander effects in the spleen of cranially-irradiated rats. Br. J. Radiol. 2017, 90. [Google Scholar] [CrossRef]
- Umegaki, K.; Sugisawa, A.; Shin, S.J.; Yamada, K.; Sano, M. Different onsets of oxidative damage to DNA and lipids in bone marrow and liver in rats given total body irradiation. Free Radic. Biol. Med. 2001, 31, 1066–1074. [Google Scholar] [CrossRef]
- Yamamoto, T.; Kinoshita, M.; Shinomiya, N.; Hiroi, S.; Sugasawa, H.; Matsushita, Y.; Majima, T.; Saitoh, D.; Seki, S. Pretreatment with Ascorbic Acid Prevents Lethal Gastrointestinal Syndrome in Mice Receiving a Massive Amount of Radiation. J. Radiat. Res. 2010, 51, 145–156. [Google Scholar] [CrossRef] [Green Version]
- La Torre, F.; Orlando, A.; Silipigni, A.; Giacobello, T.; Pergolizzi, S.; Aragona, M. Increase of oxygen free radicals and their derivatives in chemo-and radiation treated neoplasm patients. Minerva Med. 1997, 88, 121. [Google Scholar] [PubMed]
- Sakhi, A.K.; Russnes, K.M.; Thoresen, M.; Bastani, N.E.; Karlsen, A.; Smeland, S.; Blomhoff, R. Pre-radiotherapy plasma carotenoids and markers of oxidative stress are associated with survival in head and neck squamous cell carcinoma patients: A prospective study. BMC Cancer 2009, 9, 458. [Google Scholar] [CrossRef] [Green Version]
- Ushakova, T.; Melkonyan, H.; Nikonova, L.; Afanasyev, V.; Gaziev, A.I.; Mudrik, N.; Bradbury, R.; Gogvadze, V. Modification of gene expression by dietary antioxidants in radiation-induced apoptosis of mice splenocytes. Free Radic. Biol. Med. 1999, 26, 887–891. [Google Scholar] [CrossRef]
- Nenoi, M.; Ichimura, S.; Mita, K.; Yukawa, O.; Cartwright, I.L. Regulation of the catalase gene promoter by Sp1, CCAAT-recognizing factors, and a WT1/Egr-related factor in hydrogen peroxide-resistant HP100 cells. Cancer Res. 2001, 61, 5885–5894. [Google Scholar]
- Kostyuk, S.V.; Ermakov, A.V.; Alekseeva, A.Y.; Smirnova, T.D.; Glebova, K.V.; Efremova, L.V.; Baranova, A.; Veiko, N.N. Role of extracellular DNA oxidative modification in radiation induced bystander effects in human endotheliocytes. Mutat. Res. Mol. Mech. Mutagen. 2012, 729, 52–60. [Google Scholar] [CrossRef]
- Narayanan, P.K.; Goodwin, E.H.; Lehnert, B.E. Alpha particles initiate biological production of superoxide anions and hydrogen peroxide in human cells. Cancer Res. 1997, 57, 3963–3971. [Google Scholar] [PubMed]
- Khan, M.A.; Van Dyk, J.; Yeung, I.W.T.; Hill, R.P. Partial volume rat lung irradiation; assessment of early DNA damage in different lung regions and effect of radical scavengers. Radiother. Oncol. 2003, 66, 95–102. [Google Scholar] [CrossRef]
- Przybyszewski, W.M.; Widel, M.; Szurko, A.; Lubecka, B.; Matulewicz, L.; Maniakowski, Z.; Polaniak, R.; Birkner, E.; Rzeszowska-Wolny, J. Multiple bystander effect of irradiated megacolonies of melanoma cells on non-irradiated neighbours. Cancer Lett. 2004, 214, 91–102. [Google Scholar] [CrossRef] [PubMed]
- Buttke, T.M.; Sandstrom, P.A. Oxidative Stress as a Mediator of Apoptosis. Immunol. Today 1994, 15, 7–10. [Google Scholar] [CrossRef]
- Lu, T.; Finkel, T. Free radicals and senescence. Exp. Cell Res. 2008, 314, 1918–1922. [Google Scholar] [CrossRef] [PubMed]
- Day, R.M.; Snow, A.L.; Panganiban, R.A.M. Radiation-induced accelerated senescence: A fate worse than death? Cell Cycle 2014, 13, 2011–2012. [Google Scholar] [CrossRef] [Green Version]
- Jabs, T. Reactive oxygen intermediates as mediators of programmed cell death in plants and animals. Biochem. Pharmacol. 1999, 57, 231–245. [Google Scholar] [CrossRef]
- Hinkal, G.W.; Gatza, C.E.; Parikh, N.; Donehower, L.A. Altered senescence, apoptosis, and DNA damage response in a mutant p53 model of accelerated aging. Mech. Ageing Dev. 2009, 130, 262–271. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Schulte, B.A.; LaRue, A.C.; Ogawa, M.; Zhou, D. Total body irradiation selectively induces murine hematopoietic stem cell senescence. Blood 2006, 107, 358–366. [Google Scholar] [CrossRef] [Green Version]
- Palacio, L.; Krishnan, V.; Le, N.L.O.; Sharpless, N.E.; Beausejour, C.M. Sustained p16(INK4a) expression is required to prevent IR-induced tumorigenesis in mice. Oncogene 2017, 36, 1309–1314. [Google Scholar] [CrossRef] [Green Version]
- Velarde, M.C.; Flynn, J.M.; Day, N.U.; Melov, S.; Campisi, J. Mitochondrial oxidative stress caused by Sod2 deficiency promotes cellular senescence and aging phenotypes in the skin. Aging 2012, 4, 3–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borghesan, M.; Fafian-Labora, J.; Eleftheriadou, O.; Carpintero-Fernandez, P.; Paez-Ribes, M.; Vizcay-Barrena, G.; Swisa, A.; Kolodkin-Gal, D.; Ximenez-Embun, P.; Lowe, R.; et al. Small Extracellular Vesicles Are Key Regulators of Non-cell Autonomous Intercellular Communication in Senescence via the Interferon Protein IFITM3. Cell Rep. 2019, 27, 3956–3971.e6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mensa, E.; Guescini, M.; Giuliani, A.; Bacalini, M.G.; Ramini, D.; Corleone, G.; Ferracin, M.; Fulgenzi, G.; Graciotti, L.; Prattichizzo, F.; et al. Small extracellular vesicles deliver miR-21 and miR-217 as pro-senescence effectors to endothelial cells. J. Extracell. Vesicles 2020, 9. [Google Scholar] [CrossRef]
- Takahashi, A.; Ohnishi, K.; Asakawa, I.; Kondo, N.; Nakagawa, H.; Yonezawa, M.; Tachibana, A.; Matsumoto, H.; Ohnishi, T. Radiation response of apoptosis in C57BL/6N mouse spleen after whole-body irradiation. Int. J. Radiat. Biol. 2001, 77, 939–945. [Google Scholar] [CrossRef] [PubMed]
- Bogdandi, E.N.; Balogh, A.; Felgyinszki, N.; Szatmari, T.; Persa, E.; Hildebrandt, G.; Safrany, G.; Lumniczky, K. Effects of Low-Dose Radiation on the Immune System of Mice after Total-Body Irradiation. Radiat. Res. 2010, 174, 480–489. [Google Scholar] [CrossRef]
- Amundson, S.A.; Grace, M.B.; McLeland, C.B.; Epperly, M.W.; Yeager, A.; Zhan, Q.; Greenberger, J.S.; Fornace, A.J., Jr. Human in vivo radiation-induced biomarkers: Gene expression changes in radiotherapy patients. Cancer Res. 2004, 64, 6368–6371. [Google Scholar] [CrossRef] [Green Version]
- Furlong, H.; Mothersill, C.; Lyng, F.M.; Howe, O. Apoptosis is signalled early by low doses of ionising radiation in a radiation-induced bystander effect. Mutat. Res. Mol. Mech. Mutagen. 2013, 741, 35–43. [Google Scholar] [CrossRef]
- Amundson, S.A.; Do, K.T.; Fornace Jr, A.J. Induction of stress genes by low doses of gamma rays. Radiat. Res. 1999, 152, 225–231. [Google Scholar] [CrossRef]
- Zhan, Q.M.; Carrier, F.; Fornace, A.J. Induction of Cellular P53 Activity by DNA-Damaging Agents and Growth Arrest. Mol. Cell Biol. 1993, 13, 4242–4250. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.M.; Stamler, J.S. NO: An inhibitor of cell death. Cell Death Differ. 1999, 6, 937–942. [Google Scholar] [CrossRef]
- Wu-Hsieh, B.A.; Chen, W.; Lee, H.-J. Nitric Oxide Synthase Expression in Macrophages ofHistoplasma capsulatum-Infected Mice Is Associated with Splenocyte Apoptosis and Unresponsiveness. Infect. Immun. 1998, 66, 5520–5526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, B.P.C.; Li, M.X.; Asaithamby, A. New insights into the roles of ATM and DNA-PKcs in the cellular response to oxidative stress. Cancer Lett. 2012, 327, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Baltimore, D. Dual roles of ATM in the cellular response to radiation and in cell growth control. Gene Dev. 1996, 10, 2401–2410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, T.; Huang, X.; Halicka, H.D.; Zhao, H.; Traganos, F.; Albino, A.P.; Dai, W.; Darzynkiewicz, Z. Cytometry of ATM activation and histone H2AX phosphorylation to estimate extent of DNA damage induced by exogenous agents. Cytom. Part A 2007, 71A, 648–661. [Google Scholar] [CrossRef]
- Rube, C.E.; Grudzenski, S.; Kuhne, M.; Dong, X.R.; Rief, N.; Loebrich, M.; Rube, C. DNA Double-Strand Break Repair of Blood Lymphocytes and Normal Tissues Analysed in a Preclinical Mouse Model: Implications for Radiosensitivity Testing. Clin. Cancer Res. 2008, 14, 6546–6555. [Google Scholar] [CrossRef] [Green Version]
- Khanna, K.K.; Keating, K.E.; Kozlov, S.; Scott, S.; Gatei, M.; Hobson, K.; Taya, Y.; Gabrielli, B.; Chan, D.; Lees-Miller, S.P.; et al. ATM associates with and phosphorylates p53: Mapping the region of interaction. Nat. Genet. 1998, 20, 398–400. [Google Scholar] [CrossRef] [PubMed]
- Bakkenist, C.J.; Kastan, M.B. DNA damage activates ATM through intermolecular autophosphorylation and dimer dissociation. Nature 2003, 421, 499–506. [Google Scholar] [CrossRef]
- Li, L.; Zou, L. Sensing, signaling, and responding to DNA damage: Organization of the checkpoint pathways in mammalian cells. J. Cell Biochem. 2005, 94, 298–306. [Google Scholar] [CrossRef]
- Forrester, K.; Ambs, S.; Lupold, S.E.; Kapust, R.B.; Spillare, E.A.; Weinberg, W.C.; FelleyBosco, E.; Wang, X.W.; Geller, D.A.; Tzeng, E.; et al. Nitric oxide-induced p53 accumulation and regulation of inducible nitric oxide synthase expression by wild-type p53. Proc. Natl. Acad. Sci. USA 1996, 93, 2442–2447. [Google Scholar] [CrossRef] [Green Version]
- Sandstrom, P.A.; Buttke, T.M. Autocrine production of extracellular catalase prevents apoptosis of the human CEM T-cell line in serum-free medium. Proc. Natl. Acad. Sci. USA 1993, 90, 4708–4712. [Google Scholar] [CrossRef] [Green Version]
- Yabuki, M.; Kariya, S.; Ishisaka, R.; Yasuda, T.; Yoshioka, T.; Horton, A.A.; Utsumi, K. Resistance to nitric oxide-mediated apoptosis in HL-60 variant cells is associated with increased activities of Cu,Zn-superoxide dismutase and catalase. Free Radical. Biol. Med. 1999, 26, 325–332. [Google Scholar] [CrossRef]
- He, L.; Hannon, G.J. Micrornas: Small RNAs with a big role in gene regulation. Nat. Rev. Genet. 2004, 5, 522–531. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D.P. MicroRNAs: Target Recognition and Regulatory Functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghosh, R.; Guha, D.; Bhowmik, S.; Karmakar, S. Antioxidant enzymes and the mechanism of the bystander effect induced by ultraviolet C irradiation of A375 human melanoma cells. Mutat. Res.-Gen. Tox. Environ. 2013, 757, 83–90. [Google Scholar] [CrossRef]
- Bewicke-Copley, F.; Mulcahy, L.A.; Jacobs, L.A.; Samuel, P.; Akbar, N.; Pink, R.C.; Carter, D.R.F. Extracellular vesicles released following heat stress induce bystander effect in unstressed populations. J. Extracell. Vesicles 2017, 6. [Google Scholar] [CrossRef]
- Gebremedhn, S.; Gad, A.; Aglan, H.S.; Laurincik, J.; Prochazka, R.; Salilew-Wondim, D.; Hoelker, M.; Schellander, K.; Tesfaye, D. Extracellular vesicles shuttle protective messages against heat stress in bovine granulosa cells. Sci. Rep. 2020, 10. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hargitai, R.; Kis, D.; Persa, E.; Szatmári, T.; Sáfrány, G.; Lumniczky, K. Oxidative Stress and Gene Expression Modifications Mediated by Extracellular Vesicles: An In Vivo Study of the Radiation-Induced Bystander Effect. Antioxidants 2021, 10, 156. https://doi.org/10.3390/antiox10020156
Hargitai R, Kis D, Persa E, Szatmári T, Sáfrány G, Lumniczky K. Oxidative Stress and Gene Expression Modifications Mediated by Extracellular Vesicles: An In Vivo Study of the Radiation-Induced Bystander Effect. Antioxidants. 2021; 10(2):156. https://doi.org/10.3390/antiox10020156
Chicago/Turabian StyleHargitai, Rita, Dávid Kis, Eszter Persa, Tünde Szatmári, Géza Sáfrány, and Katalin Lumniczky. 2021. "Oxidative Stress and Gene Expression Modifications Mediated by Extracellular Vesicles: An In Vivo Study of the Radiation-Induced Bystander Effect" Antioxidants 10, no. 2: 156. https://doi.org/10.3390/antiox10020156