
Excess Zinc Alters Cell Wall Class III Peroxidase Activity and Flavonoid Content in the Maize Scutellum


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
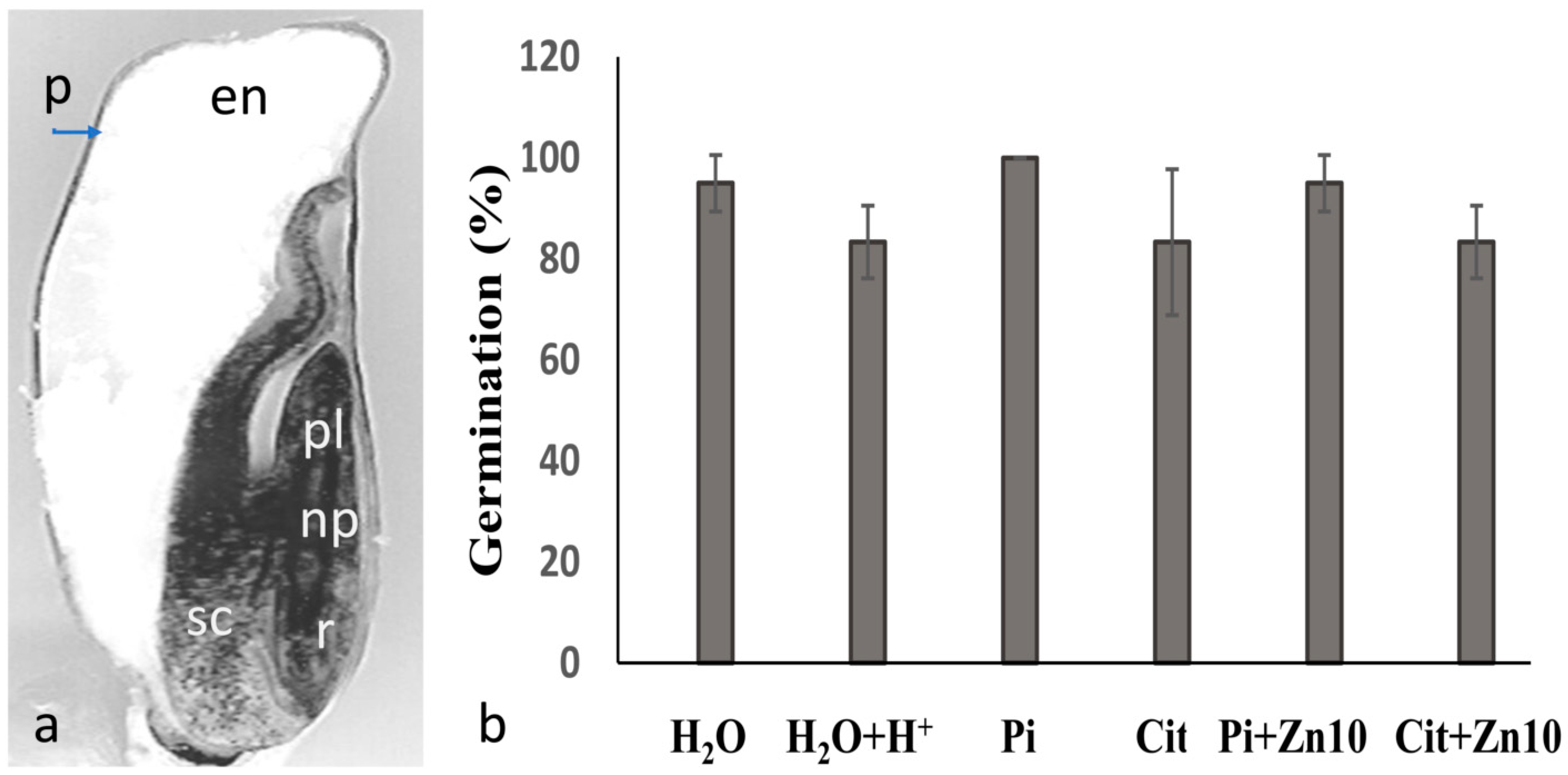
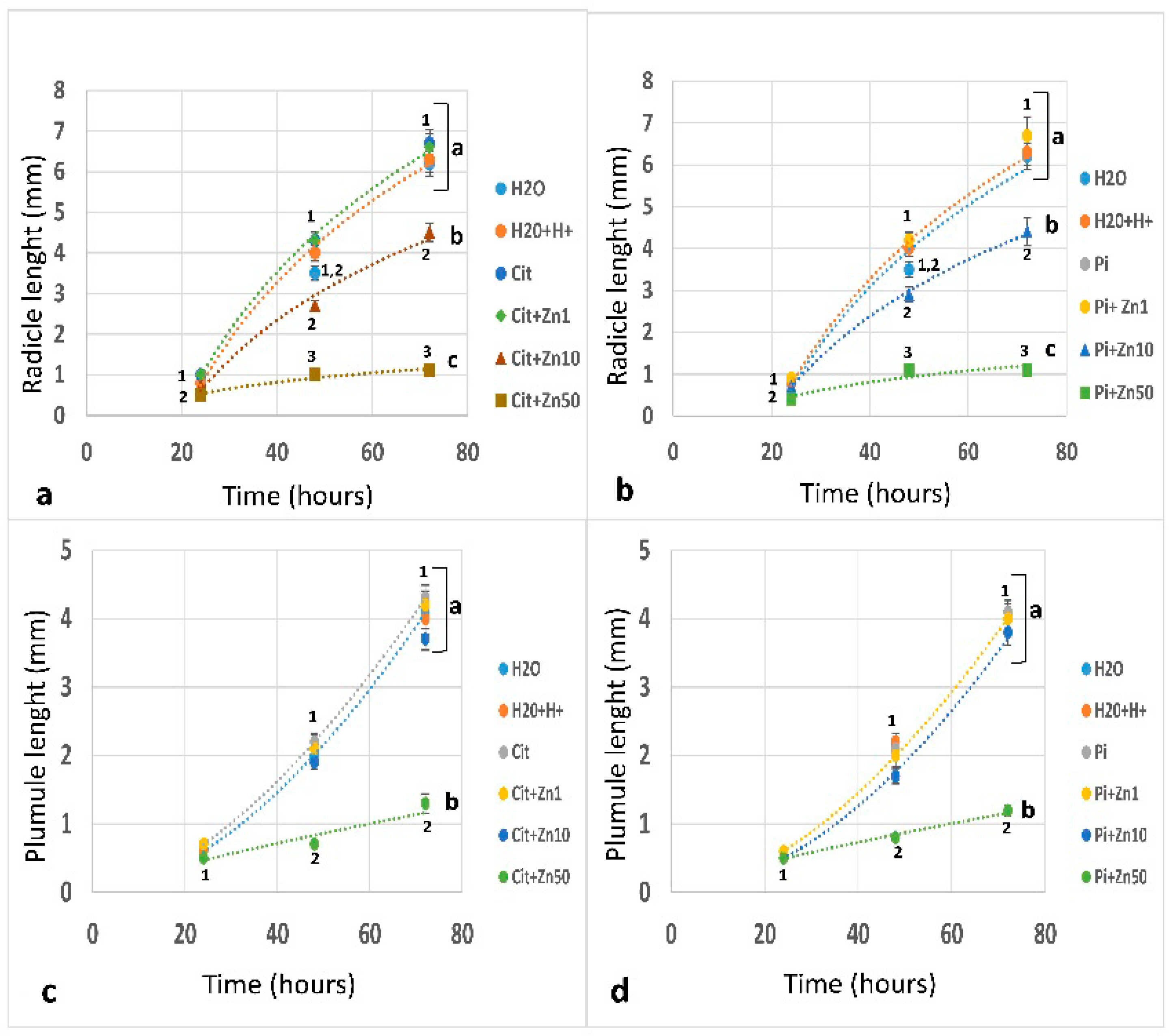
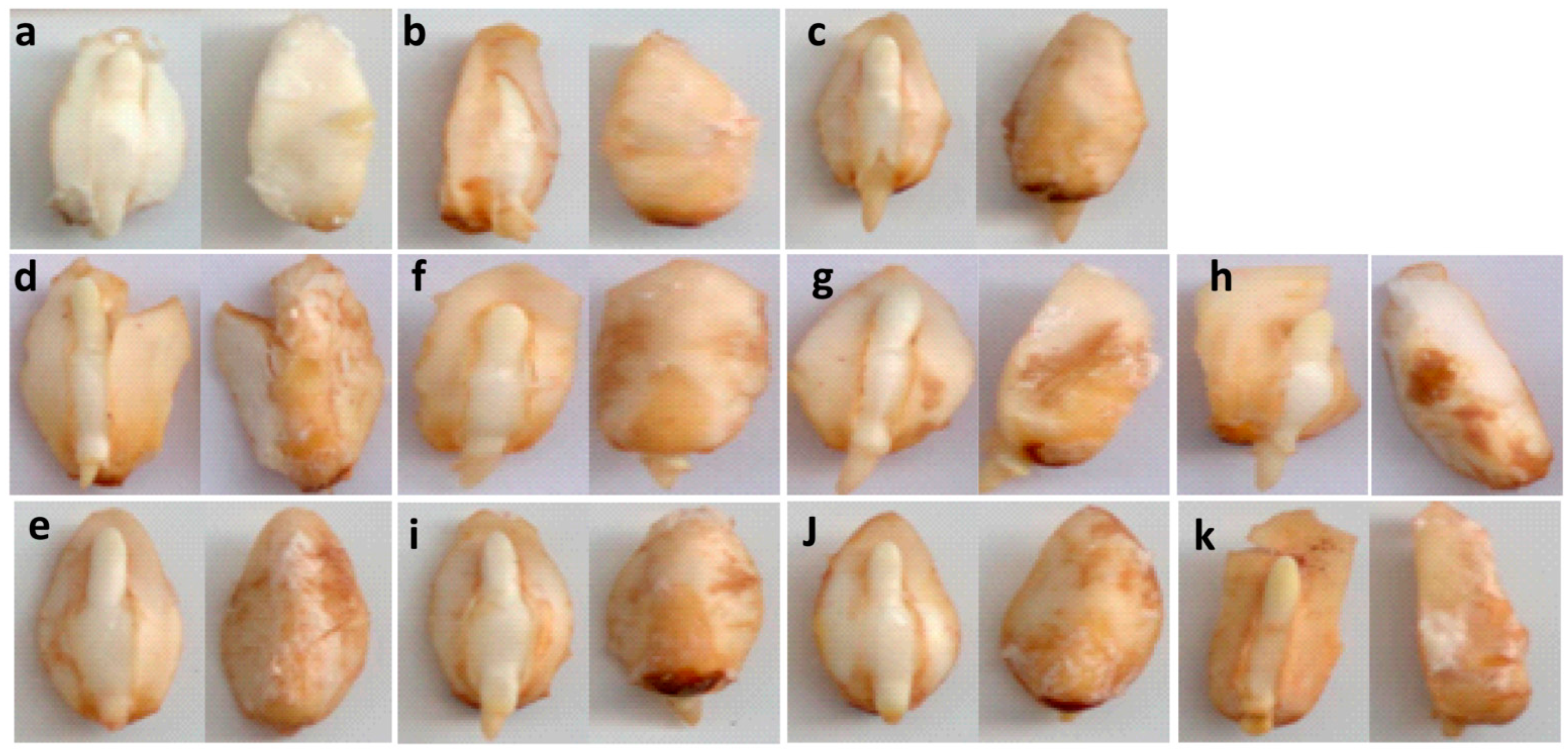
2.1. Germination and Growth
2.2. ROS Location
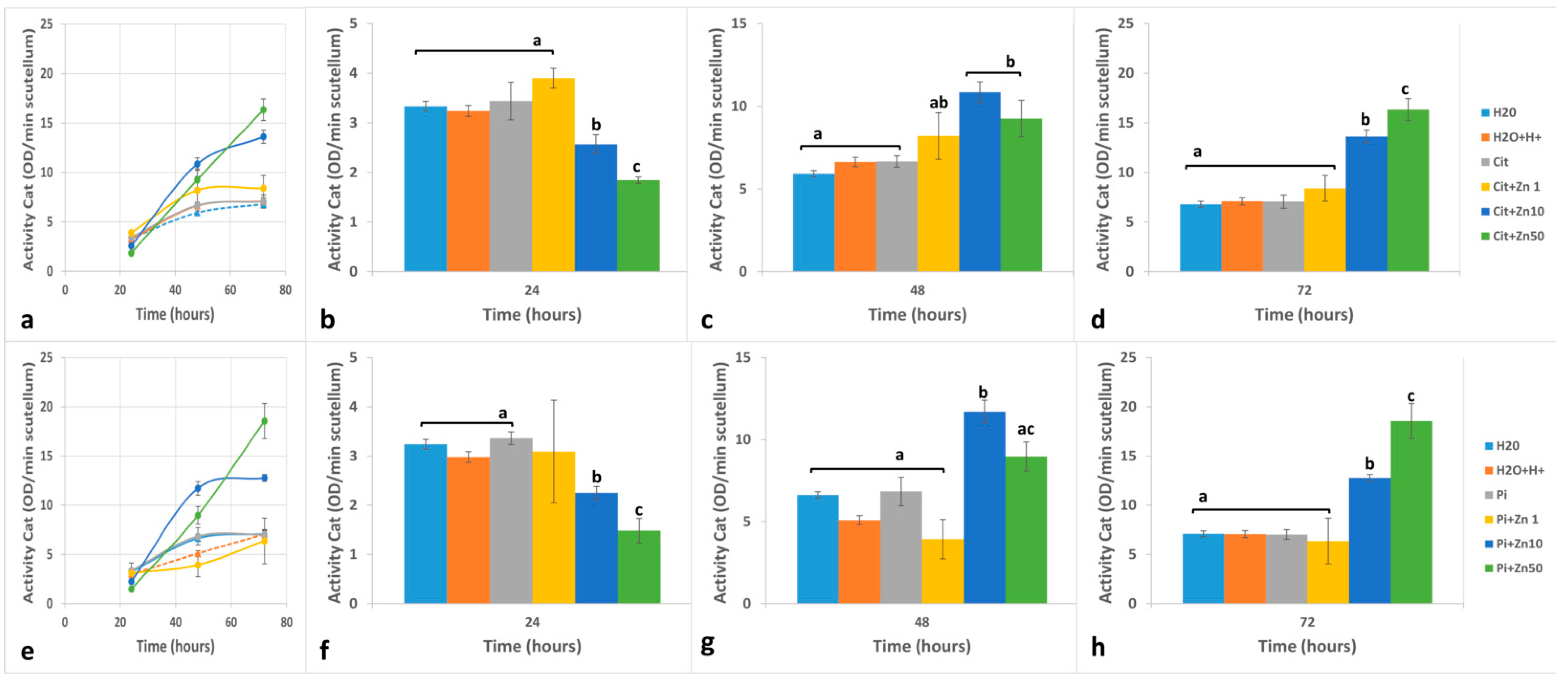
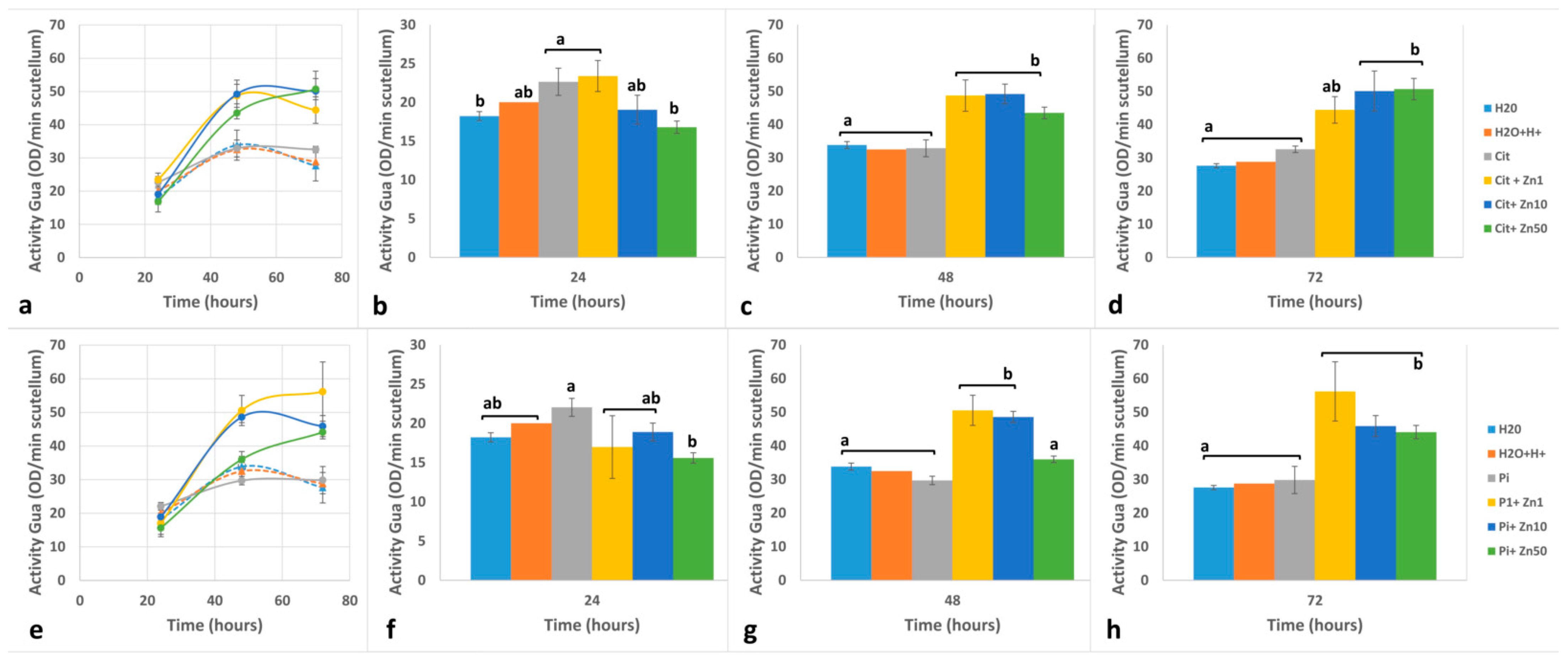
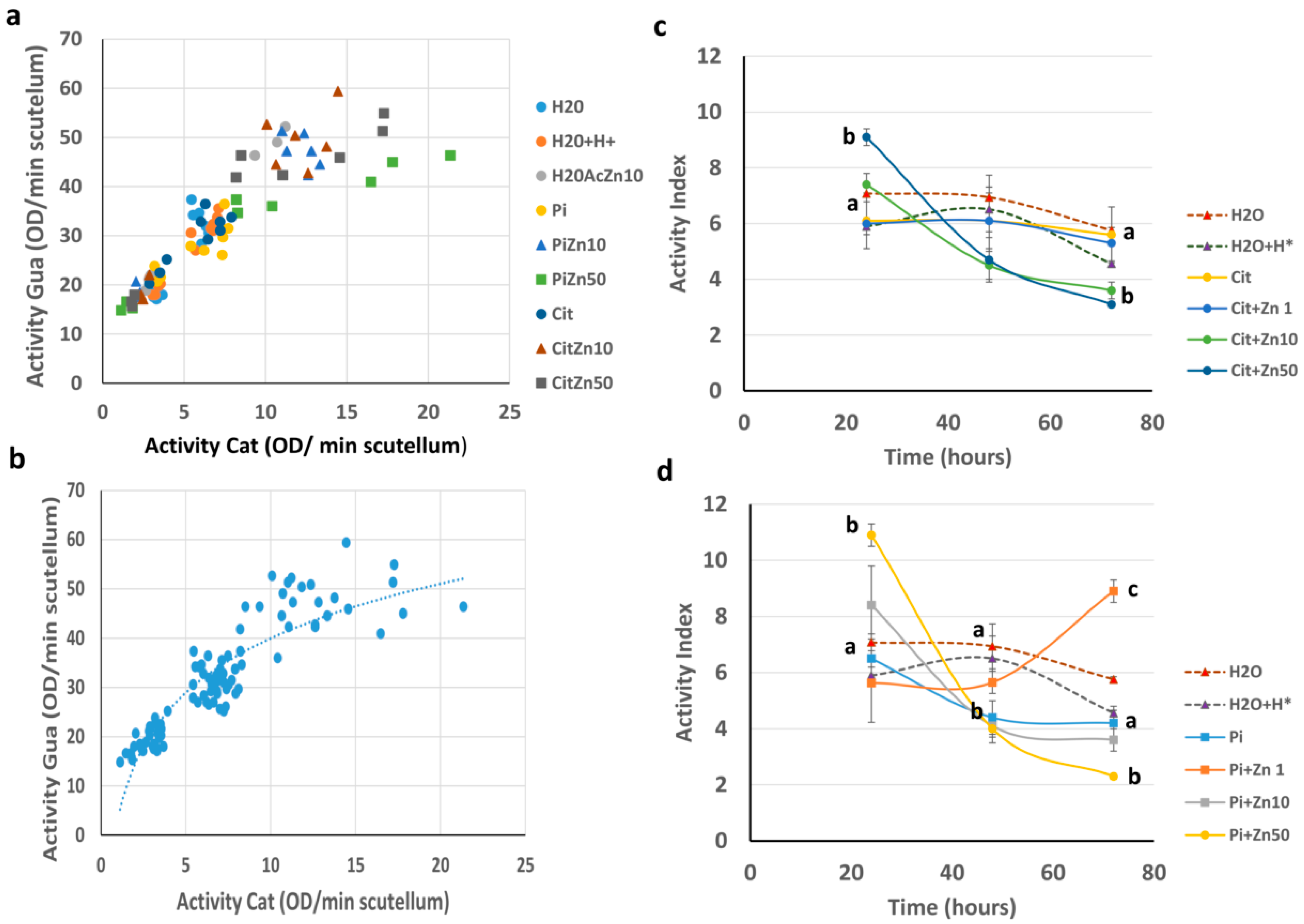
2.3. POD Activity
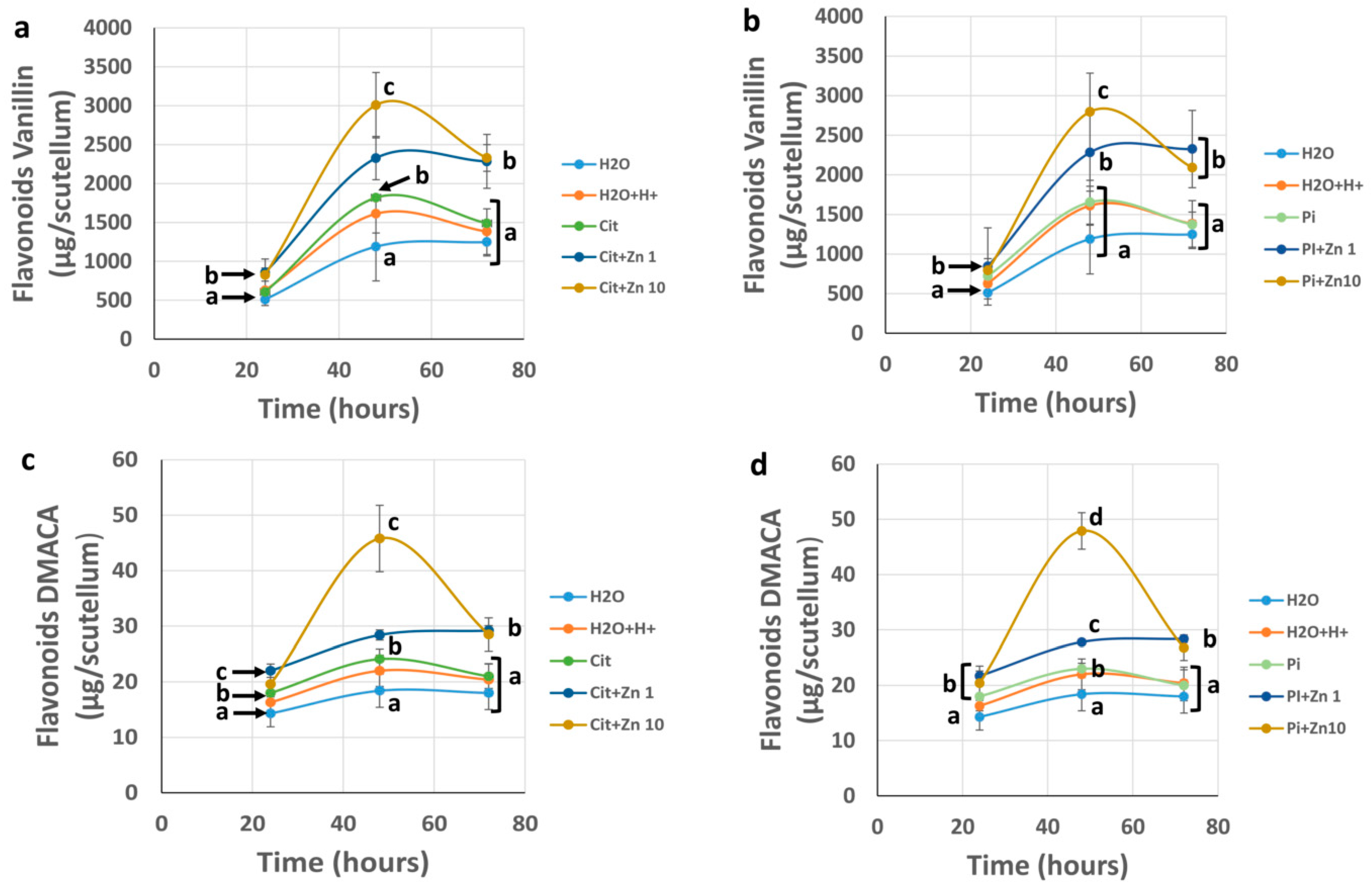
2.4. Concentration of Phenolic Compounds
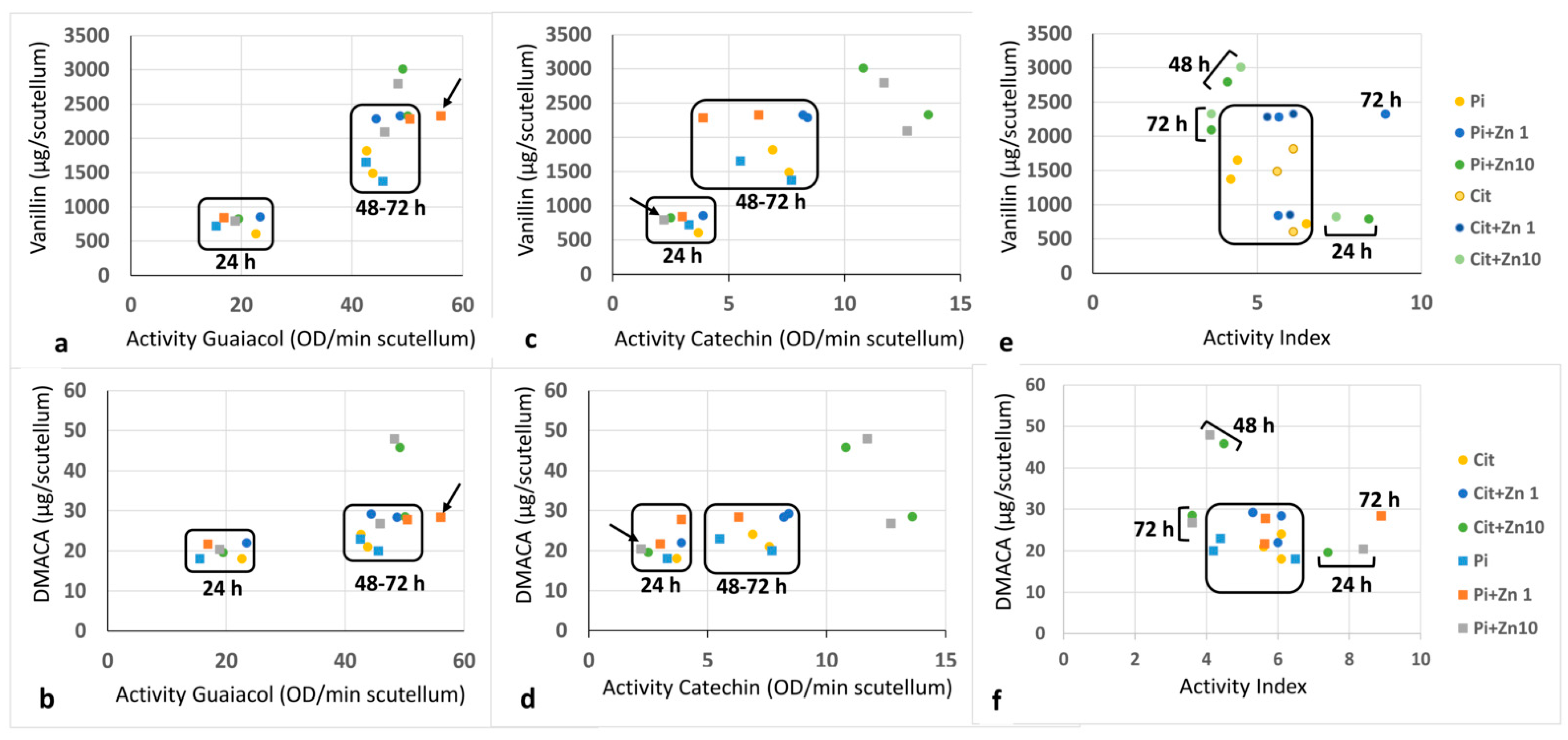
2.5. Relationship between POD Activity and Flavonoids
3. Discussion
3.1. Importance of the Scutellum in Maize
3.2. Zinc as a Nutrient and Its Disposal
3.3. Effects of Zinc on Germination, Vigor, Growth, and the Redox State
3.4. Effects of Zinc on the Amount of Phenols and POD Activity
3.5. Impact of the Study at the Food and Agronomic Levels
4. Material and Methods
4.1. Biological Material
4.2. Germination
4.3. ROS Location
4.4. POD Activity in the Scutellum
4.5. Concentration of Phenolic Compounds in the Scutellum
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AI | peroxidase activity index |
| Cat | catechin |
| Cit | germination in 10 mM citrates at pH 4.5 |
| Gua | guaiacol |
| H2O | germination in water pH 6.8 |
| H2O+H+ | germination in water at pH 4.5 |
| Pi | germination in 10 mM phosphate at pH 4.5 |
| POD | class III peroxidase |
| Zn1, Zn10, or Zn50 | ZnCl2 at 1, 10, or 50 mM |
References
- Salvador, R. Maíz, Publicaciones del Programa Nacional de Etnobotánica; Serie Traducciones: Chapingo, Mexico, 2001. [Google Scholar]
- Sistema de Información Agroalimentaria y Pesquera (SIAP-SAGARPA). Situación Actual y Perspectivas del Maíz en México 1996–2012. Available online: http://www.siap.gob.mx/estudios-2/ (accessed on 10 October 2015).
- González, H.A.; Vázquez, G.L.M.; Castellanos, J.S.; Rodríguez, P.J.E. Phenotypic diversity of maize varieties and hybrids in the Toluca-Atlacomulco Valley, Mexico. Rev. Fitotec. Mex. 2008, 31, 67–76. [Google Scholar]
- Comisión Nacional Para El Conocimiento y Uso de la Biodiversidad (CONABIO). Biodiversidad Mexicana. Available online: http://www.biodiversidad.gob.mx/usos/maices/maiz.html (accessed on 25 September 2015).
- Scanlon, M.J.; Takacs, E.M. Kernel biology. Kernel Biology en Handbook of Maize: Its Biology; Bennetzen, J.L., Hake, S.C., Eds.; Springer: New York, NY, USA, 2009; pp. 121–143. [Google Scholar]
- Dolfini, S.; Consonni, G.; Viotti, C.; Prà, M.D.; Saltini, G.; Giulini, A.; Pilu, R.; Malgioglio, A.; Gavazzi, G. A mutational approach to the study of seed development in maize. J. Exp. Bot. 2007, 58, 1197–1205. [Google Scholar] [CrossRef] [PubMed]
- Lombi, E.; Smith, E.; Hansen, T.H.; Paterson, D.; de Jonge, M.D.; Howard, D.L.; Persson, D.P.; Husted, S.; Ryan, C.; Schjoerring, J.K. Megapixel imaging of (micro)nutrients in mature barley grains. J. Exp. Bot. 2011, 62, 273–282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Consonni, G.; Gavazzi, G.; Dolfini, S. Genetic analysis as a tool to investigate the molecular mechanisms underlying seed development in maize. Ann. Bot. 2005, 96, 353–362. [Google Scholar] [CrossRef] [Green Version]
- Shewry, P.R.; Morell, M. Manipulating cereal endosperm structure, development and composition to improve end-use properties. Adv. Bot. Res. 2001, 34, 165–235. [Google Scholar] [CrossRef]
- Negbi, M. The structure and function of the scutellum of the Gramineae. Bot. J. Linn. Soc. 1984, 8, 205–222. [Google Scholar] [CrossRef]
- Bityutskii, N.P.; Magnitskiy, S.V.; Korobeynikova, L.P.; Lukina, E.I.; Soloviova, A.N.; Patsevitch, V.G.; Lapshina, I.N.; Matveeva, G.V. Distribution of iron, manganese, and zinc in mature grain and their mobilization during germination and early seedling development in maize. J. Plant Nutr. 2002, 25, 635–653. [Google Scholar] [CrossRef]
- Bityutskii, N.P.; Davydovskaya, E.N.; Malyuga, E.A.; Yakkonen, K.L. Mechanisms underlying iron and zinc transport to axis organs in grain during early seedling development of maize. J. Plant Nutr. 2004, 27, 1525–1541. [Google Scholar] [CrossRef]
- Sánchez-Linares, L.; Gavilanes-Ruíz, M.; Díaz-Pontones, D.; Guzmán-Chávez, F.; Calzada-Alejo, V.; Zurita-Villegas, V.; Sánchez-Nieto, S. Early carbon mobilization and radicle protrusion in maize germination. J. Exp. Bot. 2012, 63, 4513–4526. [Google Scholar] [CrossRef] [Green Version]
- Corona-Carrillo, J.I.; Flores-Ponce, M.; Chávez-Nájera, G.; Díaz-Pontones, D.M. Peroxidase activity in scutella of maize in association with anatomical changes during germination and grain storage. Springerplus 2014, 3, 399. [Google Scholar] [CrossRef] [Green Version]
- Evers, A.D.; O’Brien, L.; Blakeney, A.B. Cereal structure and composition. Aust. J. Agric. Res. 1999, 50, 629. [Google Scholar] [CrossRef] [Green Version]
- Fincher, G.B. Molecular and cellular biology associated with endosperm mobilization in germinating cereal grains. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1989, 40, 305–346. [Google Scholar] [CrossRef]
- Okamoto, K.; Murai, T.; Eguchi, G.; Okamoto, M.; Akazawa, T. Enzymic mechanism of starch breakdown in germinating rice seeds: 11. Ultrastructural changes in scutellar epithelium. Plant Physiol. 1982, 70, 905–911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szcziparev, S.M. Scutellum and its role in germination. In Embriology of Flowering Plants: Terminology and Concepts, Vol 2: Seed; Batygina, T.B., Ed.; CRC Press: Boca Raton, FL, USA, 2006; pp. 443–446. [Google Scholar]
- Tanaka, Y.; Ito, T.; Akazawa, T. Enzymic Mechanism of Starch Breakdown in Germinating Rice Seeds: III. alpha-Amylase Isozymes. Plant Physiol. 1970, 46, 650–654. [Google Scholar] [CrossRef] [PubMed]
- Whalley, R.D.B.; McKell, C.M.; Green, L.R. Seedling vigor and the early nonphotosynthetic stage of seedling growth in grasses. Crop Sci. 1966, 6, 147–150. [Google Scholar] [CrossRef]
- Domínguez, F.; Moreno, J.; Cejudo, F.J. The scutellum of germinated wheat grains undergoes programmed cell death: Identification of an acidic nuclease involved in nucleus dismantling. J. Exp. Bot. 2012, 63, 5475–5485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Domínguez, F.; Cejudo, F.J. Programmed cell death (PCD): An essential process of cereal seed development and germination. Front. Plant Sci. 2014, 5, 366. [Google Scholar] [CrossRef] [Green Version]
- Webb, E.C. Enzyme Nomenclature, Recommendations of the Nomenclature Committee of the International Union of Biochemistry and Molecular Biology; Academic Press: New York, NY, USA, 1992. [Google Scholar]
- Auld, D.S. Zinc coordination sphere in biochemical zinc sites. Biometals 2001, 14, 271–313. [Google Scholar] [CrossRef]
- Maret, W. Zinc coordination environments in proteins determine zinc functions. J. Trace Elem. Med. Biol. 2005, 19, 7–12. [Google Scholar] [CrossRef]
- Broadley, M.R.; White, P.J.; Hammond, J.P.; Zelko, I.; Lux, A. Zinc in plants. New Phytol. 2007, 173, 677–702. [Google Scholar] [CrossRef]
- Broadley, M.; Brown, P.; Cakmak, I.; Rengel, Z.; Zhao, F. Function of nutrients: Micronutrients. In Marschner’s Mineral Nutrition of Higher Plants, 3rd ed.; Marschner, P., Ed.; Academic Press: London, UK, 2012; pp. 191–248. [Google Scholar]
- Ciftci-Yilmaz, S.; Mittler, R. The zinc finger network of plants. Cell. Mol. Life Sci. 2008, 65, 1150–1160. [Google Scholar] [CrossRef] [PubMed]
- Takatsuji, H. Zinc-finger transcription factors in plants. Cell. Mol. Life Sci. 1998, 54, 582–596. [Google Scholar] [CrossRef] [PubMed]
- Alloway, B.J. Zinc the Vital Micronutrient for Healthy, High-Value Crop; International Zinc Association: Brussels, Belgium, 2002. [Google Scholar]
- Cakmak, I. Enrichment of cereal grains with zinc: Agronomic or genetic biofortification? Plant Soil 2008, 302, 1–17. [Google Scholar] [CrossRef]
- Sadeghzadeh, B. A review of zinc nutrition and plant breeding. J. Soil Sci. Plant Nutr. 2013, 13, 905–927. [Google Scholar] [CrossRef] [Green Version]
- Alloway, B.J. Zinc in Soils and Crop Nutrition, 2nd ed.; International Zinc Association: Brussels, Belgium; International Fertilizer Industry Association: Paris, France, 2008. [Google Scholar]
- Escudero-Almanza, D.J.; Ojeda-Barrios, D.L.; Hernández-Rodríguez, O.A.; Sánchez Chávez, E.; Ruíz-Anchondo, T.; Sida-Arreola, J.P. Carbonic anhydrase and zinc in plant physiology. Chil. J. Agric. Res. 2012, 72, 140–146. [Google Scholar] [CrossRef] [Green Version]
- Rout, G.R.; Das, P. Effect of metal toxicity on plant growth and metabolism: I. Zinc. Agronomie 2003, 23, 3–11. [Google Scholar] [CrossRef]
- Kirkby, E.; Romheld, V. Micronutrients in plant physiology: Functions, uptake and mobility. In Proceedings of Proceedings 543; International Fertiliser Society: York, UK, 2004; pp. 1–51. [Google Scholar]
- Chaney, R.L. Zinc phytotoxicity. In Zinc in Soils and Plants; Robson, A.D., Ed.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1993; pp. 135–150. [Google Scholar]
- Baker, A.J.M. Metal tolerance. New Phytol. 1987, 106, 93–111. [Google Scholar] [CrossRef]
- Tsonev, T.; Cebola Lidon, F.J. Zinc in plants—An overview. Emir. J. Food Agric. 2012, 24, 322–333. [Google Scholar]
- Doncheva, S.; Stoynova, Z.; Velikova, V. Influence of succinate on zinc toxicity of pea plants. J. Plant Nutr. 2001, 24, 789–804. [Google Scholar] [CrossRef]
- Bonnet, M.; Camares, O.; Veisseire, P. Effects of zinc and influence of Acremonium lolii on growth parameters, chlorophyll a fluorescence and antioxidant enzyme activities of ryegrass (Lolium perenne L. cv Apollo). J. Exp. Bot. 2000, 51, 945–953. [Google Scholar] [CrossRef]
- Ajouri, A.; Asgedom, H.; Becker, M. Seed priming enhances germination and seedling growth of barley under conditions of P and Zn deficiency. J. Plant Nutr. Soil Sci. 2004, 167, 630–636. [Google Scholar] [CrossRef]
- Prom-u-thai, C.; Rerkasem, B.; Yazici, A.; Cakmak, I. Zinc priming promotes seed germination and seedling vigor of rice. J. Plant Nutr. Soil Sci. 2012, 175, 482–488. [Google Scholar] [CrossRef]
- Raboy, V.; Below, F.E.; Dickinson, D.B. Alteration of maize kernel phytic acid levels by recurrent selection for protein and oil. J. Hered. 1989, 80, 311–315. [Google Scholar] [CrossRef]
- Raboy, V. Myo-Inositol-1,2,3,4,5,6-hexakisphosphate. Phytochemistry 2003, 64, 1033–1043. [Google Scholar] [CrossRef]
- Gupta, R.K.; Gangoliya, S.S.; Singh, N.K. Reduction of phytic acid and enhancement of bioavailable micronutrients in food grains. J. Food Sci. Technol. 2015, 52, 676–684. [Google Scholar] [CrossRef] [Green Version]
- Guttieri, M.; Bowen, D.; Dorsch, J.A.; Raboy, V.; Souza, E. Identification and characterization of a low phytic acid wheat. Crop Sci. 2004, 44, 418–424. [Google Scholar] [CrossRef]
- Lott, J.N.A.; Greenwood, J.S.; Batten, G.D. Mechanisms and Regulation of Mineral Nutrient Storage During Seed Development; Marcel Dekker: New York, NY, USA, 1995. [Google Scholar]
- Park, S.H.; Sung, J.K.; Lee, S.Y.; Park, J.H.; Lee, J.Y.; Jang, B.C.; Lee, B.H.; Kim, T.W. Early growth, carbohydrate, and phytic acid contents of germinating rice seeds under NaCl stress. Korean J. Crop Sci. 2006, 51, 137–141. [Google Scholar]
- Hall, J.R.; Hodges, T.K. Phosphorus metabolism of germinating oat seeds. Plant Physiol. 1966, 41, 1459–1464. [Google Scholar] [CrossRef] [Green Version]
- Lott, J.N.A. Accumulation of seed reserves of phosphorus and other minerals. In Seed Physiology; Murray, D.R., Ed.; Academic Press: New York, NY, USA, 1984; pp. 139–166. [Google Scholar]
- Raboy, V. Accumulation and storage of phosphate and minerals. In Cellular and Molecular Biology of Plant Seed Development; Larkins, B.A., Vasil, I.K., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1997; pp. 441–477. [Google Scholar]
- Loewus, F.A.; Murthy, P.P. Myo-Inositol metabolism in plants. Plant Sci. 2000, 150, 1–19. [Google Scholar] [CrossRef]
- Raboy, V.; Noaman, M.M.; Taylor, G.A.; Pickett, S.G. Grain phytic acid and protein are highly correlated in winter wheat. Crop Sci. 1991, 31, 631–635. [Google Scholar] [CrossRef]
- Nadeem, M.; Mollier, A.; Morel, C.; Vives, A.; Prud’homme, L.; Pellerin, S. Relative contribution of seed phosphorus reserves and exogenous phosphorus uptake to maize (Zea mays L.) nutrition during early growth stages. Plant Soil 2011, 346, 231–244. [Google Scholar] [CrossRef]
- Nadeem, M.; Mollier, A.; Morel, C.; Vives, A.; Prud’homme, L.; Pellerin, S. Maize (Zea mays L.) endogenous seed phosphorus remobilization is not influenced by exogenous phosphorus availability during germination and early growth stages. Plant Soil 2012, 357, 13–24. [Google Scholar] [CrossRef]
- Bolland, M.D.A.; Baker, M.J. High phosphorus concentrations in seed of wheat and annual medic are related to higher rates of dry matter production of seedlings and plants. Aust. J. Exp. Agric. 1988, 28, 765–770. [Google Scholar] [CrossRef]
- Ros, C.; Bell, R.W.; White, P.F. Effect of seed phosphorus and soil phosphorus applications on early growth of rice (Oryza sativa L.) cv. IR66. Soil Sci. Plant Nutr. 1997, 43, 499–509. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Nyborg, M.; McGill, W.B. Phosphorus concentration in barley (Hordeum vulgare L.) seed: Influence on seedling growth and dry matter production. Plant Soil 1990, 122, 79–83. [Google Scholar] [CrossRef]
- Zhu, Y.G.; Smith, S.E. Seed phosphorus (P) content affects growth, and P uptake of wheat plants and their association with arbuscular mycorrhizal (AM) fungi. Plant Soil 2001, 231, 105–112. [Google Scholar] [CrossRef]
- Kisko, M.; Bouain, N.; Safi, A.; Medici, A.; Akkers, R.C.; Secco, D.; Fouret, G.; Krouk, G.; Aarts, M.G.; Busch, W.; et al. LPCAT1 controls phosphate homeostasis in a zinc-dependent manner. Elife 2018, 7, e32077. [Google Scholar] [CrossRef]
- Sagardoy, R.; Morales, F.; López-Millán, A.F.; Abadía, A.; Abadía, J. Effects of zinc toxicity on sugar beet (Beta vulgaris L.) plants grown in hydroponics. Plant Biol. 2008, 11, 339–350. [Google Scholar] [CrossRef]
- Tsuji, N.; Hirayanagi, N.; Okada, M.; Miyasaka, H.; Hirata, K.; Zenk, M.H.; Miyamoto, K. Enhancement of tolerance to heavy metals and oxidative stress in Dunaliella tertiolecta by Zn-induced phytochelatin synthesis. Biochem. Biophys. Res. Commun. 2002, 293, 653–659. [Google Scholar] [CrossRef]
- Bailly, C. Active oxygen species and antioxidants in seed biology. Seed Sci. Res. 2004, 14, 93–107. [Google Scholar] [CrossRef]
- Bailly, C.; El-Maarouf-Bouteau, H.; Corbineau, F. From intracellular signaling networks to cell death: The dual role of reactive oxygen species in seed physiology. C. R. Biol. 2008, 331, 806–814. [Google Scholar] [CrossRef]
- Lin, Y.F.; Aarts, M.G.M. The molecular mechanism of zinc and cadmium stress response in plants. Cell. Mol. Life Sci. 2012, 69, 3187–3206. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Patni, B.; Shankhdhar, D.; Shankhdhar, S.C. Zinc–an indispensable micronutrient. Physiol. Mol. Biol. Plants 2013, 19, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive oxygen species, oxidative damage, and antioxidative defense mechanism in plants under stressful conditions. J. Bot. 2012, 2012, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Jia, L.; Xu, W.; Li, W.; Ye, N.; Liu, R.; Shi, L.; Bin, A.N.M.R.; Fan, M.; Zhang, J. Class III peroxidases are activated in proanthocyanidin-deficient Arabidopsis thaliana seeds. Ann. Bot. 2013, 111, 839–847. [Google Scholar] [CrossRef] [Green Version]
- Agati, G.; Azzarello, E.; Pollastri, S.; Tattini, M. Flavonoids as antioxidants in plants: Location and functional significance. Plant Sci. 2012, 196, 67–76. [Google Scholar] [CrossRef]
- Sakihama, Y.; Cohen, M.; Grace, S.; Yamasaki, Y. Plant phenolic antioxidant and prooxidant activities: Phenolics-induced oxidative damage mediated by metals in plants. Toxicology 2002, 177, 67–80. [Google Scholar] [CrossRef]
- Fini, A.; Brunetti, C.; Di Ferdinando, M.; Ferrini, F.; Tattini, M. Stress-induced flavonoid biosynthesis and the antioxidant machinery of plants. Plant Signal. Behav. 2011, 6, 709–711. [Google Scholar] [CrossRef]
- Heim, K.E.; Tagliaferro, A.R.; Bobilya, D.J. Flavonoid antioxidants: Chemistry, metabolism and structure-activity relationships. J. Nutr. Biochem. 2002, 13, 572–584. [Google Scholar] [CrossRef]
- Kumar, S.; Pandey, A.K. Chemistry and biological activities of flavonoids: An overview. Sci. World J. 2013, 2013, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michalak, A. Phenolic compounds and their antioxidant activity. Pol. J. Environ. Stud. 2006, 15, 523–530. [Google Scholar]
- Pourcel, L.; Routaboul, J.M.; Cheynier, V.; Lepiniec, L.; Debeaujon, I. Flavonoid oxidation in plants: From biochemical properties to physiological functions. Trends Plant Sci. 2007, 12, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Passardi, F.; Cosio, C.; Penel, C.; Dunand, C. Peroxidases have more functions than a Swiss army knife. Plant Cell Rep. 2005, 24, 255–265. [Google Scholar] [CrossRef] [PubMed]
- Takahama, U.; Oniki, T. Flavonoids and some other phenolics as substrates of peroxidase: Physiological significance of the redox reactions. J. Plant Res. 2000, 113, 301–309. [Google Scholar] [CrossRef]
- Yamasaki, H.; Sakihama, Y.; Ikehara, N. Flavonoid-peroxidase reaction as a detoxification mechanism of plant cells against H2O2. Plant Physiol. 1997, 115, 1405–1412. [Google Scholar] [CrossRef] [Green Version]
- Panchaksharappa, M.G.; Rudramuniyappa, C.K. Scutellum and endosperm in the seedlings of Gramineae—A histochemical study. Proc. Indian Acad. Sci. 1975, 82, 211–217. [Google Scholar]
- Chang, S.S. Micronutrients in crop production of Taiwan. In Proceedings of the International Workshop on Micronutrient in Crop Production; National Taiwan University: Taipei, Taiwan, 1999. [Google Scholar]
- Foth, H.D.; Ellis, B.G. Soil Fertility, 2nd ed.; Lewis Publishers: New York, NY, USA, 1997. [Google Scholar]
- Manara, A. Plant Responses to Heavy Metal Toxicity. In Plants and Heavy Metals; Furini, A., Ed.; Springer: Dordrecht, The Netherlands, 2012. [Google Scholar] [CrossRef]
- López-Millán, A.F.; Ellis, D.R.; Grusak, M.A. Effect of zinc and manganese supply on the activities of superoxide dismutase and carbonic anhydrase in Medicago truncatula wild type and raz mutant plants. Plant Sci. 2005, 168, 1015–1022. [Google Scholar] [CrossRef]
- Kappus, H. Lipid Peroxidation: Mechanisms, Analysis, Enzymology and Biological Relevance. In Oxidative Stress; Sies, H., Ed.; Academic Press: London, UK, 1985; pp. 273–310. [Google Scholar]
- Weckx, J.E.J.; Clijsters, H.M.M. Zn phytotoxicity induces oxidative stress in primary leaves of Phaseolus vulgaris. Plant Physiol. Bioch. 1997, 35, 405–410. [Google Scholar]
- Cakmak, I. Possible roles of zinc in protecting plant cells from damage by reactive oxygen species. New Phytol. 2000, 146, 185–205. [Google Scholar] [CrossRef]
- Madhava Rao, K.V.; Sresty, T.V.S. Antioxidative parameters in the seedlings of pigeonpea (Cajanus cajan (L.) Millspaugh) in response to Zn and Ni stresses. Plant Sci. 2000, 157, 113–128. [Google Scholar] [CrossRef]
- Morina, F.; Jovanovic, L.; Mojovic, M.; Vidovic, M.; Pankovic, D.; Veljovic, J.S. Zinc-induced oxidative stress in Verbascum thapsus L. is caused by an accumulation of reactive oxygen species and quinhydrone in the cell wall. Physiol. Plant. 2010, 140, 209–224. [Google Scholar] [CrossRef] [PubMed]
- Kukavica, B.; Mojović, M.; Vucčinić, Ž.; Maksimović, V.; Takahama, U.; Jovanović, S.V. Generation of hydroxyl radical in isolated pea root cell wall, and the role of cell wall-bound peroxidase, Mn-SOD and phenolics in their production. Plant Cell Physiol. 2009, 50, 304–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mongkhonsina, B.; Nakbanpotea, W.; Hokurab, A.; Nuengchamnongc, N.; Maneechaia, S. Phenolic compounds responding to zinc and/or cadmium treatments in Gynura pseudochina (L.) DC. extracts and biomass. Plant Physiol. Biochem. 2016, 109, 549–560. [Google Scholar] [CrossRef]
- Yang, X.; Feng, Y.; He, Z.; Stoffella, P.J. Molecular mechanisms of heavy metal hyperaccumulation and phytoremediation. J. Trace Elem. Med. Biol. 2005, 18, 339–353. [Google Scholar] [CrossRef] [PubMed]
- Food and Agriculture Organization. Datos de Seguridad Alimetaria. America Latina y Caribe. Available online: http://www.fao.org/faostat/es/#data/FSI (accessed on 4 July 2019).
- Welch, R.M.; Combs, G.F., Jr.; Duxbury, J.M. Toward a “greener” revolution. Issues Sci. Technol. 1997, 14, 50–58. [Google Scholar]
- Consejo Nacional de Evaluación de la Política de Desarrollo Social. Dimensiones de la Seguridad Alimentaria: Evaluación Estratégica de Nutrición y Abasto; Coneval: Ciuda de Mexico, Mexico, 2010. [Google Scholar]
- Gutiérrez, J.P.; Rivera-Dommarco, J.; Shamah-Levy, T.; Villalpando-Hernández, S.; Franco, A.; Cuevas-Nasu, L.; Romero-Martinez, M.; Hernandez-Avila, M. Encuesta Nacional de Salud y Nutrición 2012, Resultados Nacionales; Instituto Nacional de Salud Pública: Cuernavaca, Mexico, 2012. [Google Scholar]
- Urquía-Fernández, N. La seguridad alimentaria en México. Salud Pública Méx. 2014, 56, s92–s98. [Google Scholar] [CrossRef] [Green Version]
- Assunção, A.G.L.; Schat, H.; Aarts, M.G.M. Regulation of the adaptation to zinc deficiency in plants. Plant Signal. Behav. 2010, 5, 1553–1555. [Google Scholar] [CrossRef] [Green Version]
- Stein, A.J. Global impacts of human mineral malnutrition. Plant Soil 2010, 335, 133–154. [Google Scholar] [CrossRef]
- Welch, R.M.; Graham, R.D. Breeding crops for enhanced micronutrient content. Plant Soil 2002, 245, 205–214. [Google Scholar] [CrossRef]
- Welch, R.M.; Graham, R.D. Breeding for micronutrients in staple food crops from a human nutrition perspective. J. Exp. Bot. 2004, 55, 353–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uauy, C.; Distelfeld, A.; Fahima, T.; Blechl, A.; Dubcovsky, J. A NAC gene regulating senescence improves grain protein, zinc, and iron content in wheat. Science 2006, 314, 1298–1301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Genc, Y.; McDonald, G.K.; Graham, R.D. Effect of seed zinc content on early growth of barley (Hordeum vulgare L.) under low and adequate soil zinc supply. Aust. J. Agric. Res. 2000, 51, 37–46. [Google Scholar] [CrossRef]
- Harris, D.; Rashid, A.; Miraj, G.; Arif, M.; Shah, H. ‘On-farm’ seed priming with zinc sulphate solution—A cost-effective way to increase the maize yields of resource-poor farmers. Field Crops Res. 2007, 102, 119–127. [Google Scholar] [CrossRef]
- Johnson, S.E.; Lauren, J.G.; Welch, R.M.; Duxbury, J.M. A comparison of the effects of micronutrient seed priming and soil fertilization on the mineral nutrition of chickpea (Cicer arietinum), lentil (Lens culinaris), rice (Oryza sativa) and wheat (Triticum aestivum) in Nepal. Exp. Agric. 2005, 41, 427–448. [Google Scholar] [CrossRef] [Green Version]
- Corpas, F.J.; Barroso, J.B. Reactive sulfur species (RSS): Possible new players in the oxidative metabolism of plant peroxisomes. Front. Plant Sci. 2015, 6, 116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Filipovic, M.R.; Jovanovic, V.M. More than just an intermediate: Hydrogen sulfide signalling in plants. J. Exp. Bot. 2017, 68, 4733–4736. [Google Scholar] [CrossRef]
- Harmens, H.; Den Hartog, P.R.; Bookum, W.M.T.; Verkleij, J.A.C. Increased zinc tolerance in silene vulgaris (moench) garcke is not due to increased production of phytochelatins. Plant Physiol. 1993, 103, 1305–1309. [Google Scholar] [CrossRef] [Green Version]
- Jambunathan, N. Determination and detection of reactive oxygen species (ROS), lipid peroxidation, and electrolyte leakage in plants. In Plant Stress Tolerance Methods in Molecular Biology (Methods and Protocols); Sunkar, R., Ed.; Humana Press: Totowa, NJ, USA, 2010; Volume 639, pp. 291–297. [Google Scholar] [CrossRef]
- Maxson, E.D.; Rooney, L.W. Evaluation of methods for tannin analysis in sorghum grain. Am. Assoc. Cereal Chem. 1972, 49, 719–729. [Google Scholar]
- Walton, M.F.; Haskins, A.; Gorz, H. False positive results in the vanillin-hel assay of tannins in sorghum forage. Crop Sci. 1983, 23, 197–200. [Google Scholar] [CrossRef]
- Ivanova, V.; Stefova, M.; Chinnici, F. Determination of the polyphenol contents in Macedonian grapes and wines by standardized spectrophotometric methods. J. Serb. Chem. Soc. 2010, 75, 45–59. [Google Scholar] [CrossRef]
- Kuskoski, E.M.; Asuero, A.G.; Troncoso, A.M.; Fett, R. Antioxidant capacity (ORACFL) of frozen fruit´s pulps. Nutr. Rev. Soc. 2006, 31, 53–64. [Google Scholar] [CrossRef] [Green Version]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Díaz-Pontones, D.M.; Corona-Carrillo, J.I.; Herrera-Miranda, C.; González, S. Excess Zinc Alters Cell Wall Class III Peroxidase Activity and Flavonoid Content in the Maize Scutellum. Plants 2021, 10, 197. https://doi.org/10.3390/plants10020197
Díaz-Pontones DM, Corona-Carrillo JI, Herrera-Miranda C, González S. Excess Zinc Alters Cell Wall Class III Peroxidase Activity and Flavonoid Content in the Maize Scutellum. Plants. 2021; 10(2):197. https://doi.org/10.3390/plants10020197
Chicago/Turabian StyleDíaz-Pontones, David Manuel, José Isaac Corona-Carrillo, Carlos Herrera-Miranda, and Sandra González. 2021. "Excess Zinc Alters Cell Wall Class III Peroxidase Activity and Flavonoid Content in the Maize Scutellum" Plants 10, no. 2: 197. https://doi.org/10.3390/plants10020197