Use of Taxonomic and Trait-Based Approaches to Evaluate the Effect of Bt maize Expressing Cry1Ie Protein on Non-Target Collembola: A Case Study in Northeast China

 ,
, 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Source of Maize Varieties
2.2. Experimental Design
2.3. Sample Collection
2.4. Statistical Analysis
3. Results
3.1. Collembola Composition
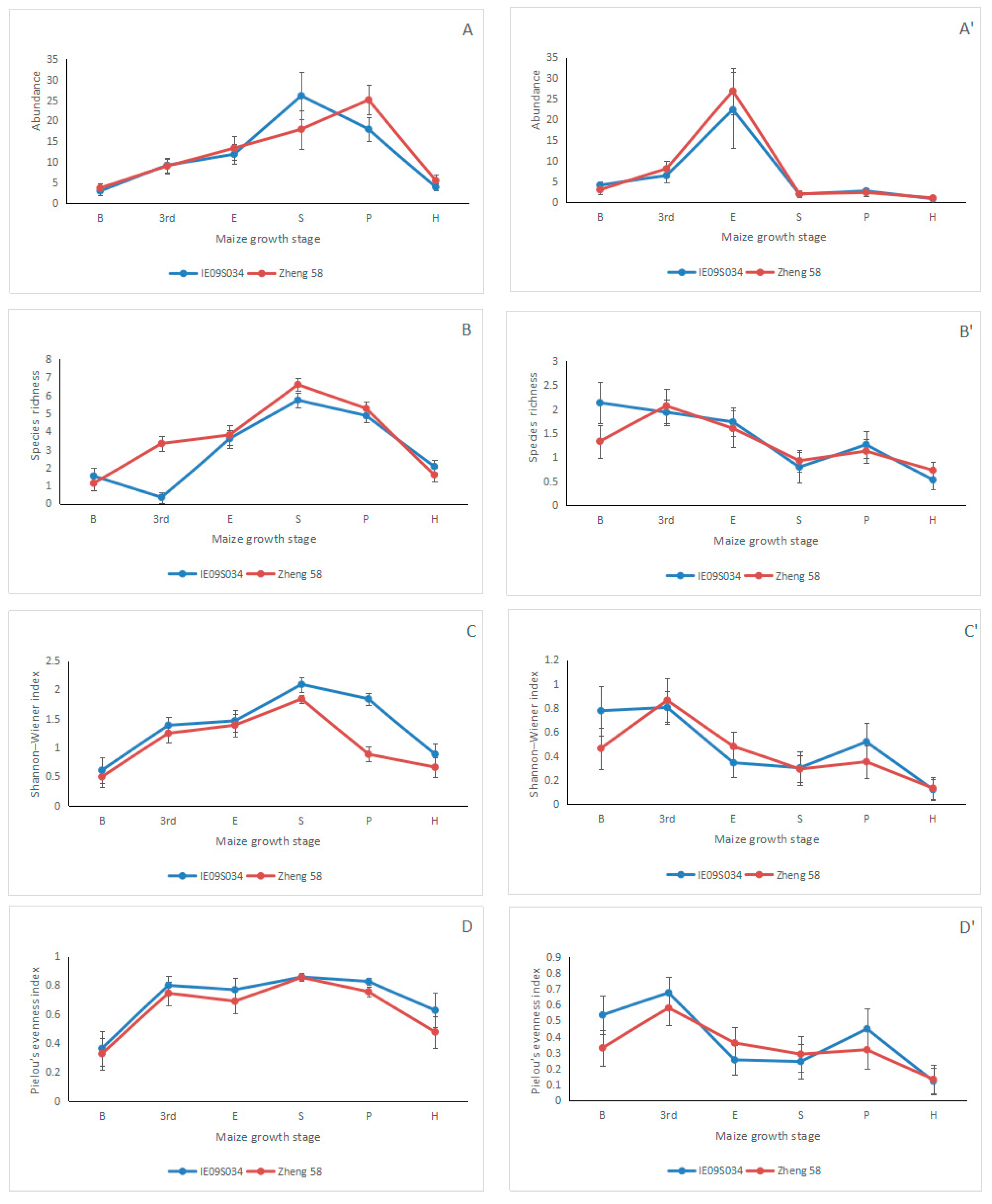
3.2. Effects of Maize Variety and Maize Growth Stage on Collembolan Abundance and Diversity
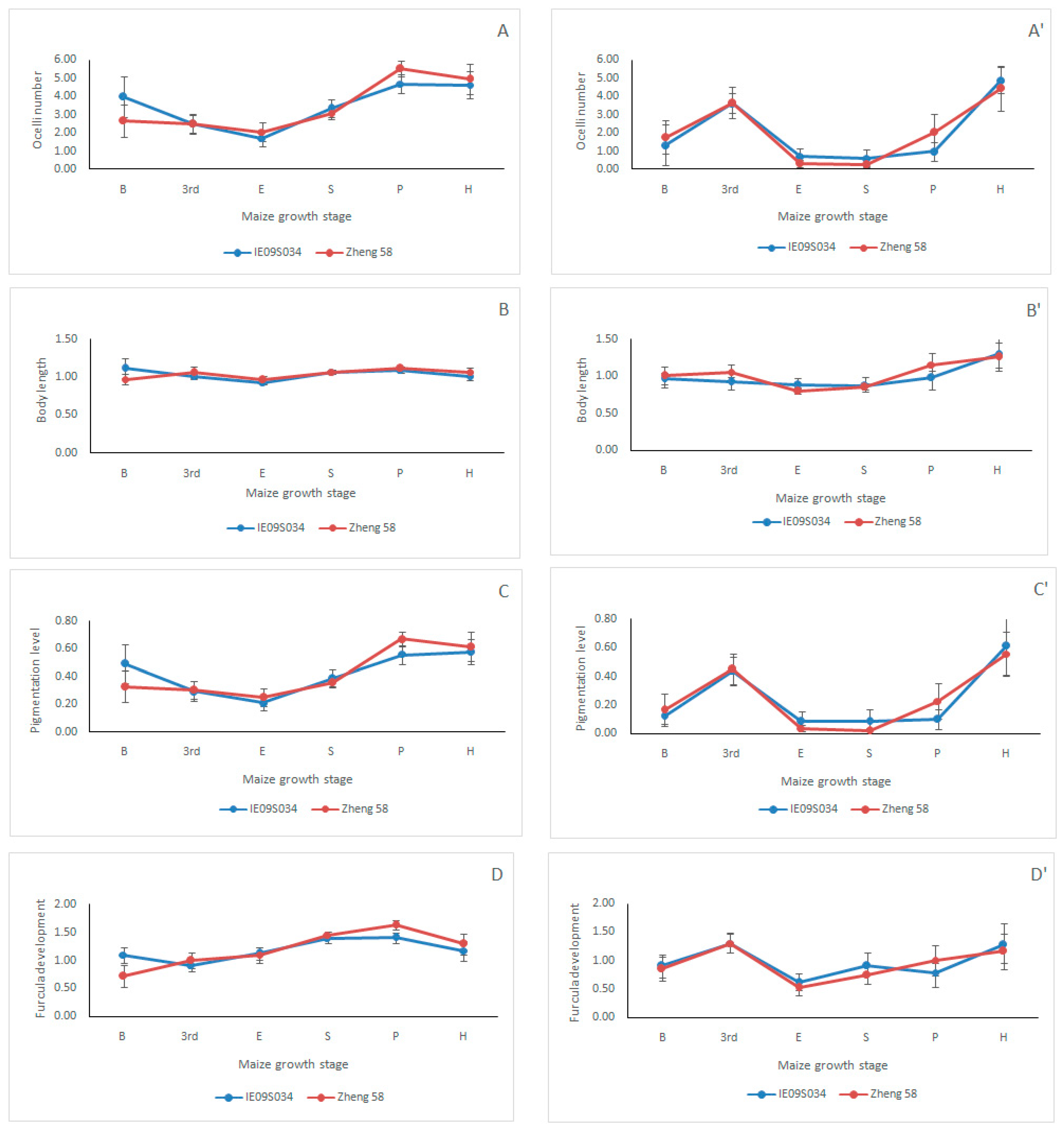
3.3. Effects of Maize Variety and Maize Growth Stage on Collembolan Morphological TraitAttribute Value (CWM)
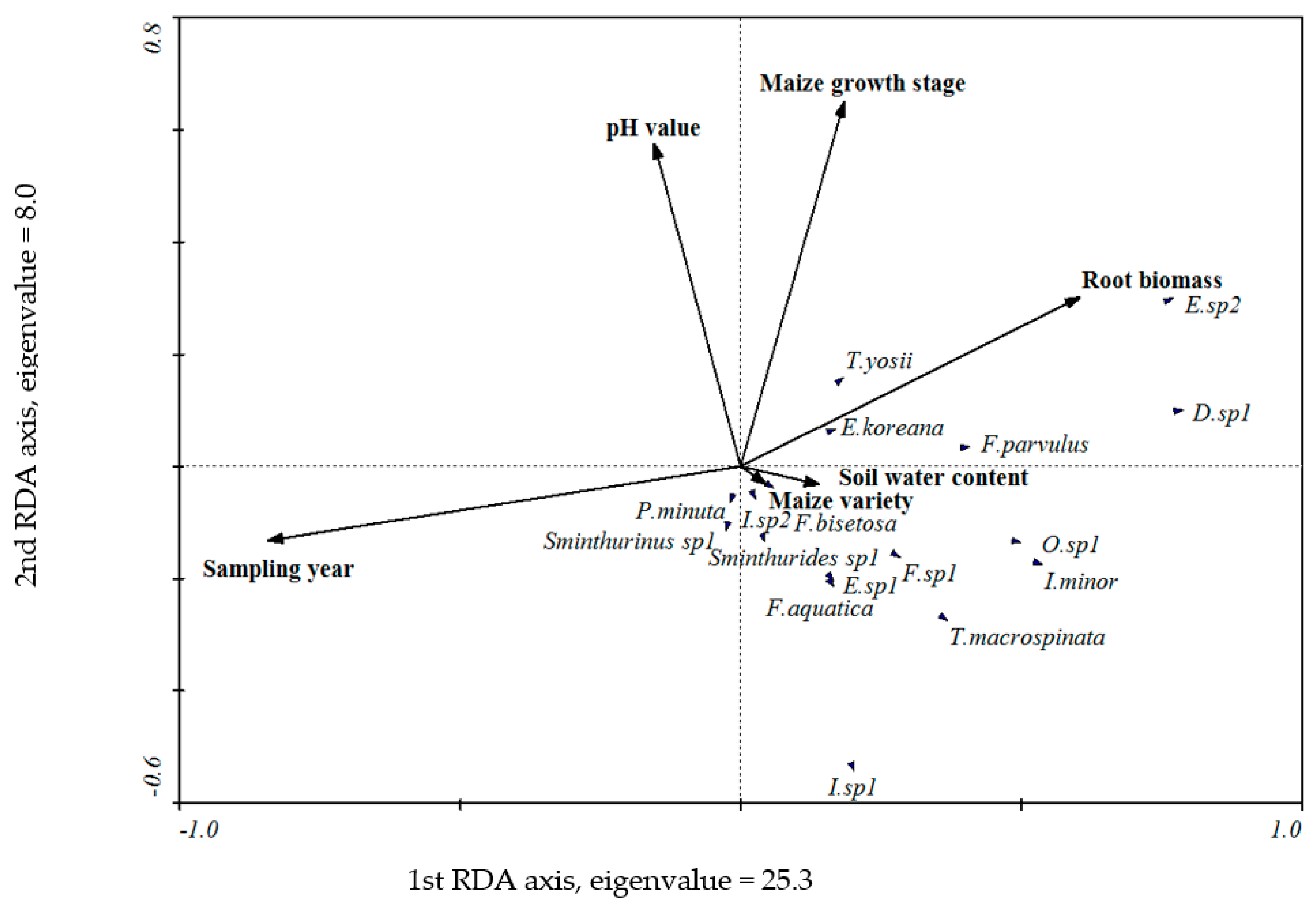
3.4. The Relationship between Environmental Variables and the Collembolan Community Composition
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ishida, Y.; Saito, H.; Ohta, S.; Hiei, Y.; Komari, T.; Kumashiro, T. High efficiency transformation of maize (Zea mays L.) mediated by Agrobacterium tumefaciens. Nat. Biotechnol. 1996, 14, 745–750. [Google Scholar] [CrossRef] [PubMed]
- Saxena, D.; Flores, S.; Stotzky, G. Insecticidal toxin in root exudates from Bt corn. Nature 1999, 402, 480. [Google Scholar] [CrossRef] [PubMed]
- Baumgarte, S.; Tebbe, C.C. Field studies on the environmental fate of the Cry1Ab Bt-toxin produced by transgenic maize (MON810) and its effect on bacterial communities in the maize rhizosphere. Mol. Ecol. 2005, 14, 2539–2551. [Google Scholar] [CrossRef] [PubMed]
- Icoz, I.; Stotzky, G. Fate and effects of insect-resistant Bt crops in soil ecosystems. Soil Biol. Biochem. 2008, 40, 559–586. [Google Scholar] [CrossRef]
- Mendelsohn, M.J.; Kough, J.; Vaituzis, Z.; Matthews, K. Are Bt crops safe? Nat. Biotechnol. 2003, 21, 1003–1009. [Google Scholar] [CrossRef] [PubMed]
- Geng, L.L.; Cui, H.J.; Dai, P.L.; Lang, Z.H.; Shu, C.L.; Zhou, T.; Song, F.P.; Zhang, J. The influence of Bt-transgenic maize pollen on the bacterial diversity in the midgut of Apis mellifera ligustica. Apidologie 2013, 44, 198–208. [Google Scholar] [CrossRef]
- Saxena, D.; Stotzky, G. Bacillus thuringiensis (Bt) toxin released from root exudates and biomass of Bt corn has no apparent effect on earthworms, nematodes, protozoa, bacteria, and fungi in soil. Soil Biol. Biochem. 2001, 33, 1225–1230. [Google Scholar] [CrossRef]
- Zwahlen, C.; Hilbeck, A.; Gugerli, P.; Nentwig, W. Degradation of the Cry1Ab protein within transgenic Bacillus thuringiensis corn tissue in the field. Mol. Ecol. 2003, 12, 765–775. [Google Scholar] [CrossRef]
- Daudu, C.K.; Muchaonyerwa, P.; Mnkeni, P.N.S. Litterbag decomposition of genetically modified maize residues and their constituent Bacillus thuringiensis protein (Cry1Ab) under field conditions in the central region of the Eastern Cape, South Africa. Agric. Ecosyst. Environ. 2009, 34, 153–158. [Google Scholar] [CrossRef]
- Li, X.G.; Liu, B. A 2-year field study shows little evidence that the long-term planting of transgenic insect-resistant cotton affects the community structure of soil nematodes. PLoS ONE 2013, 8, e61670. [Google Scholar] [CrossRef] [Green Version]
- Yang, B.; Chen, H.; Liu, X.H.; Ge, F.; Chen, Q.Y. Bt cotton planting does not affect the community characteristics of rhizosphere soil nematodes. Appl. Soil Ecol. 2014, 73, 156–164. [Google Scholar] [CrossRef]
- Arias-Martín, M.; García, M.; Luciáñez, M.J.; Ortego, F.; Castañera, P.; Farinós, G.P. Effects of three-year cultivation of Cry1Ab-expressing Bt maize on soil microarthropod communities. Agric. Ecosyst. Environ. 2016, 220, 125–134. [Google Scholar] [CrossRef]
- Shu, Y.; Zhang, Y.; Zeng, H.; Zhang, Y.; Wang, J. Effects of Cry1Ab Bt maize straw return on bacterial community of earthworm Eisenia fetida. Chemosphere 2017, 173, 1–13. [Google Scholar] [CrossRef]
- Saxena, D.; Stotzky, G. Bt corn has a higher lignin content than non-Bt corn. Am. J. Bot. 2001, 88, 1704–1706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, M.; Motavalli, P.P.; Kremer, R.J.; Nelson, K.A. Assessing changes in soil microbial communities and carbon mineralization in Bt and non-Bt corn residue-amended soils. Appl. Soil Ecol. 2007, 37, 150–160. [Google Scholar] [CrossRef]
- Henriksen, T.M.; Breland, T.A. Nitrogen availability effects on carbon mineralization, fungal and bacterial growth, and enzyme activities during decomposition of wheat straw in soil. Soil Biol. Biochem. 1999, 31, 1121–1134. [Google Scholar] [CrossRef]
- Vikman, M.; Karjomaa, S.; Kapanen, A.; Wallenius, K.; Itävaara, M. The influence of lignin content and temperature on the biodegradation of lignocellulose in composting conditions. Appl. Microbiol. Biotechnol. 2002, 59, 591–598. [Google Scholar] [CrossRef]
- Güsewell, S.; Gessner, M.O. N:P ratios influence litter decomposition and colonization by fungi and bacteria in microcosms. Funct. Ecol. 2009, 23, 211–219. [Google Scholar] [CrossRef]
- Peterson, H.; Luxton, M. A comparative analysis of soil fauna populations and their role in decomposition processes. Oikos 1982, 39, 287–388. [Google Scholar] [CrossRef]
- Rusek, J. Biodiversity of Collembola and their functional role in the ecosystem. Biodivers. Conserv. 1998, 7, 1207–1219. [Google Scholar] [CrossRef]
- Bellinger, P.F.; Christiansen, K.A.; Janssens, F. Checklist of the Collembola of the World. 1996–2019. Available online: http://www.collembola.org (accessed on 25 September 2019).
- Filser, J. The role of Collembola in carbon and nitrogen cycling in soil. Pedobiologia 2002, 46, 234–245. [Google Scholar] [CrossRef]
- Endlweber, K.; Scheu, S. Effects of Collembola on root properties of two competing ruderal plant species. Soil Biol. Biochem. 2006, 38, 2025–2031. [Google Scholar] [CrossRef]
- Rebek, E.J.; Hogg, D.B.; Young, D.K. Effect of four cropping systems on the abundance and diversity of epedaphic Springtails (Hexapoda: Parainsecta: Collembola) in southern Wisconsin. Environ. Entomol. 2002, 31, 37–46. [Google Scholar] [CrossRef] [Green Version]
- Santorufo, L.; Cortet, J.; Arena, C.; Goudon, R.; Rakoto, A.; Morel, J.L.; Maisto, G. An assessment of the influence of the urban environment on collembolan communities in soils using taxonomy- and trait-based approaches. Appl. Soil Ecol. 2014, 78, 48–56. [Google Scholar] [CrossRef]
- Rossetti, I.; Bagella, S.; Cappai, C.; Caria, M.C.; Lai, R.; Roggero, P.P.; Martins da Silva, P.; Sousa, J.P.; Querner, P.; Seddaiu, G. Isolated cork oak trees affect soil properties and biodiversity in a Mediterranean wooded grassland. Agric. Ecosyst. Environ. 2015, 202, 203–216. [Google Scholar] [CrossRef]
- Hopkin, S.P. Biology of the Springtails: (Insecta: Collembola); Oxford University Press: New York, NY, USA, 1997; pp. 1–201. [Google Scholar]
- Silva, P.M.D.; Carvalho, F.; Dirilgen, T.; Stone, D.; Creamer, R.; Bolger, T.; Sousa, J.P. Traits of collembolan life-form indicate land use types and soil properties across an European transect. Appl. Soil Ecol. 2016, 97, 69–77. [Google Scholar] [CrossRef]
- Winck, B.R.; Sá, E.L.S.D.; Rigotti, V.M.; Chauvat, M. Relationship between land-use types and functional diversity of epigeicCollembola in Southern Brazil. Appl. Soil Ecol. 2017, 109, 49–59. [Google Scholar] [CrossRef]
- Makkonen, M.; Berg, M.P.; Hal, J.R.V.; Callaghan, T.V.; Press, M.C.; Aerts, R. Traits explain the responses of a sub-arctic Collembola community to climate manipulation. Soil Biol. Biochem. 2011, 43, 377–384. [Google Scholar] [CrossRef]
- Joimel, S.; Capiaux, H.; Schwartz, C.; Hedde, M.; Lebeau, T.; Guern, C.; Nahmani, J.; Pernin, C.; Salmon, S.; Santorufo, L.; et al. Effect of geogenic lead on fungal and collembolan communities in garden topsoil. Pedosphere 2018, 28, 215–226. [Google Scholar] [CrossRef] [Green Version]
- Song, X.Y.; Chang, L.; Reddy, G.V.P.; Zhang, L.; Fan, C.M.; Wang, B.F. Use of Taxonomic and trait-based approaches to evaluate the effects of transgenic Cry1Ac Corn on the community characteristics of soil collembola. Environ. Entomol. 2019, 48, 263–269. [Google Scholar] [CrossRef]
- Song, F.; Zhang, J.; Gu, A.; Wu, Y.; Han, L.; He, K.; Chen, Z.; Yao, J.; Hu, Y.; Li, G.; et al. Identification of cry1I-Type Genes from Bacillus thuringiensis Strains and Characterization of a Novel cry1I-Type Gene. Appl. Environ. Microb. 2003, 69, 5207–5211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Liu, Y.; Ren, Y.; Liu, Y.; Liang, G.; Song, F.; Bai, S.X.; Wang, J.H.; Wang, G.Y. Overexpression of a novel Cry1Ie gene confers resistance to Cry1Ac-resistant cotton bollworm in transgenic lines of maize. Plant Cell Tissue Organ Cult. 2013, 115, 151–158. [Google Scholar] [CrossRef]
- Guo, J.F.; He, K.L.; Bai, S.X.; Zhang, T.T.; Liu, Y.J.; Wang, F.X.; Wang, Z.Y. Effects of transgenic cry1Ie maize on non-lepidopteran pest abundance, diversity and community composition. Transgenic Res. 2016, 25, 761–772. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.F.; He, K.L.; Hellmich, R.L.; Bai, S.X.; Zhang, T.T.; Liu, Y.J.; Ahmed, T.; Wang, Z.Y. Field trials to evaluate the effects of transgenic Cry1Ie maize on the community characteristics of arthropod natural enemies. Sci. Rep. UK 2016, 6, 22102. [Google Scholar] [CrossRef] [PubMed]
- Fan, C.M.; Wu, F.C.; Dong, J.Y.; Wang, B.F.; Yin, J.Q.; Song, X.Y. No impact of transgenic cry1Ie maize on the diversity, abundance and composition of soil fauna in a 2-year field trial. Sci. Rep. UK 2019, 9, 10333. [Google Scholar] [CrossRef] [Green Version]
- Macfadyen, A. Improved funnel-type extractors for soil arthropods. J. Anim. Ecol. 1961, 30, 171–184. [Google Scholar] [CrossRef]
- Christiansen, K.A.; Bellinge, P.F. The Collembola of North America, North of the Rio Grande: A Taxonomic Analysis, 2nd ed.; Grinnell College: Grinnell, IA, USA, 1998. [Google Scholar]
- Yin, W.Y. Pictorial Keys to Soil Animals of China; Science Press: Beijing, China, 1998. [Google Scholar]
- Potapov, M. Synopses on Palaearctic Collembola: Isotomidae; Abhandlungen und Berichte des Naturkundemuseums: Görlitz, Poland, 2001; Volume 73, pp. 1–603. [Google Scholar]
- Fjellberg, A. The Collembola of Fennoscandia and Denmark. Part Ⅱ: Entomobryomorpha and Symphypleona. In Fauna Entomologica Scandinavica; Koninklijke Brill NV: Leiden, The Netherlands, 2007; Volume 42, pp. 1–264. [Google Scholar] [CrossRef]
- Braak, C.J.F.T.; Smilaue, P. CANOCO Reference Manual and CanoDraw for Windows User’s Guide: Software for Canonical Community Ordination (Version 4.5); Microcomputer Power: New York, NY, USA, 2002. [Google Scholar]
- Höss, S.; Menzel, R.; Gessler, F.; Nguyen, H.T.; Jehle, J.A.; Traunspurger, W. Effects of insecticidal crystal proteins (Cry proteins) produced by genetically modified maize (Bt maize) on the nematode Caenorhabditis elegans. Environ. Pollut. 2013, 178, 147–151. [Google Scholar] [CrossRef]
- Szabó, B.; Seres, A.; Bakonyi, G. Long-term consumption and food replacement of near-isogenic by Bt-maize alter life-history traits of Folsomia candida Willem 1902 (Collembola). Appl. Ecol. Environ. Res. 2017, 15, 1275–1286. [Google Scholar] [CrossRef]
- Fiera, C. Biodiversity of Collembola in urban soils and their use as bioindicators for pollution. Pesqui. Agropecu. Bras. 2009, 44, 868–873. [Google Scholar] [CrossRef] [Green Version]
- Fountain, M.T.; Hopkin, S.P. Biodiversity of Collembola in urban soils and the use of Folsomia candida to assess soil ‘quality’. Ecotoxicology 2004, 13, 555–572. [Google Scholar] [CrossRef]
- Bitzer, R.J.; Rice, M.E.; Pilcher, C.D.; Pilcher, C.L.; Lam, W.F. Biodiversity and community structure of epedaphic and euedaphic springtails (Collembola) in transgenic rootworm Bt maize. Environ. Entomol. 2005, 34, 1346–1376. [Google Scholar] [CrossRef] [Green Version]
- Bai, Y.Y.; Yan, R.H.; Ye, G.Y.; Huang, F.N.; Cheng, J.A. Effects of transgenic rice expressing Bacillus thuringiensis Cry1Ab protein on ground-dwelling collembolan community in postharvest seasons. Environ. Entomol. 2010, 39, 243–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, L.; Liu, X.H.; Ge, F. Effect of elevated O3 associated with Bt cotton on the abundance, diversity and community structure of soil Collembola. Appl. Soil Ecol. 2011, 47, 45–50. [Google Scholar] [CrossRef]
- Al-Deeb, M.; Wilde, G.E.; Blair, J.M.; Todd, T.C. Effect of bt corn for corn rootworm control on nontarget soil microarthropods and nematodes. Environ. Entomol. 2003, 32, 859–865. [Google Scholar] [CrossRef] [Green Version]
- Bakonyi, G.; Szira, F.; Kiss, I.; Villányi, I.; Seres, A.; Székacs, A. Preference tests with collembolas on isogenic and bt-maize. Eur. J. Soil Biol. 2006, 42 (Suppl. S1), S132–S135. [Google Scholar] [CrossRef]
- Coulson, S.J.; Hodkinson, I.D.; Wooley, C.; Webb, N.R.; Block, W.; Worland, M.R.; Bale, J.S.; Strathdee, A.T. Effects of experimental temperature elevation on high-arctic soil microarthropod populations. Polar Biol. 1996, 16, 147–153. [Google Scholar] [CrossRef]
- Ruess, L.; Michelsen, A.; Schmidt, I.K.; Jonasson, S. Simulated climate change affecting microorganisms, nematode density and biodiversity in subarctic soils. Plant Soil 1999, 212, 63–73. [Google Scholar] [CrossRef]
- Maraun, M.; Alphei, J.; Beste, P.; Bonkowski, M.; Buryn, R.; Migge, S.; Peter, M.; Schaefer, M.; Scheu, S. Indirect effects of carbon and nutrient amendments on the soil meso- and microfauna of a beechwood. Biol. Fertil. Soils 2001, 34, 222–229. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Trait | Data Type | Attribute Value |
|---|---|---|
| Ocelli number | Ordinal | 0–8 |
| Body length | Quantitative | in mm, to the nearest 0.1mm |
| Pigmentation level | Ordinal | 0 = neither colored nor patterned 1 = colored with no patterns 2 = colored with distinct patterns |
| Furcula development | Ordinal | 0 = absent 1 = shorter than head 2 = longer than head |
| Collembola Group | 2014 | 2015 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Number | Percentage (%) | Number | Percentage (%) | |||||||
| Family | Genus | Species | Bt | CK | Bt | CK | Bt | CK | Bt | CK |
| Onychiuridae | Thalassaphorura | macrospinata | 278 | 252 | 25.69 | 17.91 | 265 | 296 | 45.93 | 45.47 |
| Tullbergiidae | Tullbergia | yosii | 4 | 7 | 0.37 | 0.50 | 1 | 1 | 0.17 | 0.15 |
| Isotomidae | Isotomodes | sp.1 | 119 | 179 | 11.00 | 12.72 | 189 | 223 | 32.76 | 34.25 |
| Isotomodes | sp.2 | 2 | 0 | 0.18 | 0 | 0 | 0 | 0 | 0 | |
| Isotomiella | minor | 73 | 93 | 6.75 | 6.61 | 11 | 9 | 1.91 | 1.38 | |
| Folsomia | aquatica | 60 | 68 | 5.55 | 4.83 | 29 | 23 | 5.03 | 3.53 | |
| Folsomia | bisetosa | 7 | 23 | 0.65 | 1.63 | 4 | 1 | 0.69 | 0.15 | |
| Folsomides | parvulus | 29 | 24 | 2.68 | 1.71 | 9 | 0 | 1.56 | 0 | |
| Folsomides | sp.1 | 4 | 6 | 0.37 | 0.43 | 1 | 1 | 0.17 | 0.15 | |
| Proisotoma | minuta | 13 | 22 | 1.20 | 1.56 | 6 | 7 | 1.04 | 1.08 | |
| Desoria | sp.1 | 251 | 272 | 23.20 | 19.33 | 3 | 9 | 0.52 | 1.38 | |
| Entomobryidae | Orchesellides | sp.1 | 60 | 107 | 5.55 | 7.60 | 15 | 26 | 2.60 | 3.99 |
| Entomobrya | koreana | 22 | 43 | 2.03 | 3.06 | 10 | 22 | 1.73 | 3.38 | |
| Entomobrya | sp.1 | 13 | 38 | 1.20 | 2.70 | 28 | 26 | 4.85 | 3.99 | |
| Entomobrya | sp.2 | 145 | 272 | 13.40 | 19.33 | 4 | 4 | 0.69 | 0.61 | |
| Sminthurididae | Sminthurinus | sp.1 | 2 | 0 | 0.18 | 0 | 0 | 2 | 0 | 0.31 |
| Sminthurides | sp.1 | 0 | 1 | 0 | 0.07 | 2 | 1 | 0.35 | 0.15 | |
| Total | 1082 | 1407 | 100 | 100 | 577 | 651 | 100 | 100 | ||
| Year. | Variable | Abundance (N) | Species Richness (S) | Shannon-Wiener index (H’) | Pielou’s Evenness index (J) | ||||
|---|---|---|---|---|---|---|---|---|---|
| F | P | F | P | F | P | F | P | ||
| 2014 | Maize variety | 2.422 | 0.122 | 0.020 | 0.887 | 0.482 | 0.489 | 1.872 | 0.173 |
| Maize growth stage | 35.702 | 0.000 *** | 41.94 | 0.000 *** | 29.62 | 0.000 *** | 10.925 | 0.000 *** | |
| Maize variety×Maize growth stage | 0.437 | 0.822 | 0.903 | 0.480 | 0.445 | 0.816 | 0.203 | 0.961 | |
| 2015 | Maize variety | 0.202 | 0.654 | 0.331 | 0.566 | 0.331 | 0.566 | 0.528 | 0.469 |
| Maize growth stage | 15.075 | 0.000 *** | 6.360 | 0.000 *** | 6.039 | 0.000 *** | 5.014 | 0.000 *** | |
| Maize variety×Maize growth stage | 0.602 | 0.699 | 0.763 | 0.577 | 0.667 | 0.649 | 0.612 | 0.691 | |
| Year | Variable | Ocelli Number (O) | Body Length (B) | Pigmentation Level (P) | Furcula Development (F) | ||||
|---|---|---|---|---|---|---|---|---|---|
| F | P | F | P | F | P | F | P | ||
| 2014 | Maize variety | 0.002 | 0.964 | 0.005 | 0.946 | 0.000 | 0.993 | 0.043 | 0.836 |
| Maize growth stage | 9.687 | 0.000 *** | 2.408 | 0.039 * | 9.388 | 0.000 *** | 7.252 | 0.000 *** | |
| Maize variety×Maize growth stage | 0.637 | 0.672 | 0.847 | 0.518 | 0.659 | 0.655 | 1.062 | 0.384 | |
| 2015 | Maize variety | 0.018 | 0.893 | 0.252 | 0.617 | 0.002 | 0.963 | 0.076 | 0.783 |
| Maize growth stage | 8.593 | 0.000 *** | 3.088 | 0.012 * | 8.741 | 0.000 *** | 3.449 | 0.006 ** | |
| Maize variety×Maize growth stage | 0.272 | 0.928 | 0.399 | 0.848 | 0.269 | 0.929 | 0.192 | 0.965 | |
| Environmental Variable | Variance Explained (%) | F | p |
|---|---|---|---|
| Sampling year | 19 | 16.11 | 0.002 ** |
| Root biomass | 7 | 6.55 | 0.002 ** |
| Maize growth stage | 6 | 6.56 | 0.002 ** |
| Soil water content | 4 | 3.40 | 0.010 * |
| pH value | 1 | 1.49 | 0.166 |
| Maize variety | 1 | 0.66 | 0.666 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, B.-F.; Wu, F.-C.; Yin, J.-Q.; Jiang, Z.-L.; Song, X.-Y.; Reddy, G.V.P. Use of Taxonomic and Trait-Based Approaches to Evaluate the Effect of Bt maize Expressing Cry1Ie Protein on Non-Target Collembola: A Case Study in Northeast China. Insects 2021, 12, 88. https://doi.org/10.3390/insects12020088
Wang B-F, Wu F-C, Yin J-Q, Jiang Z-L, Song X-Y, Reddy GVP. Use of Taxonomic and Trait-Based Approaches to Evaluate the Effect of Bt maize Expressing Cry1Ie Protein on Non-Target Collembola: A Case Study in Northeast China. Insects. 2021; 12(2):88. https://doi.org/10.3390/insects12020088
Chicago/Turabian StyleWang, Bai-Feng, Feng-Ci Wu, Jun-Qi Yin, Zhi-Lei Jiang, Xin-Yuan Song, and Gadi V. P. Reddy. 2021. "Use of Taxonomic and Trait-Based Approaches to Evaluate the Effect of Bt maize Expressing Cry1Ie Protein on Non-Target Collembola: A Case Study in Northeast China" Insects 12, no. 2: 88. https://doi.org/10.3390/insects12020088