Musculoskeletal Growth Modulation in Gilthead Sea Bream Juveniles Reared at High Water Temperature and Fed with Palm and Rapeseed Oils-Based Diets

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
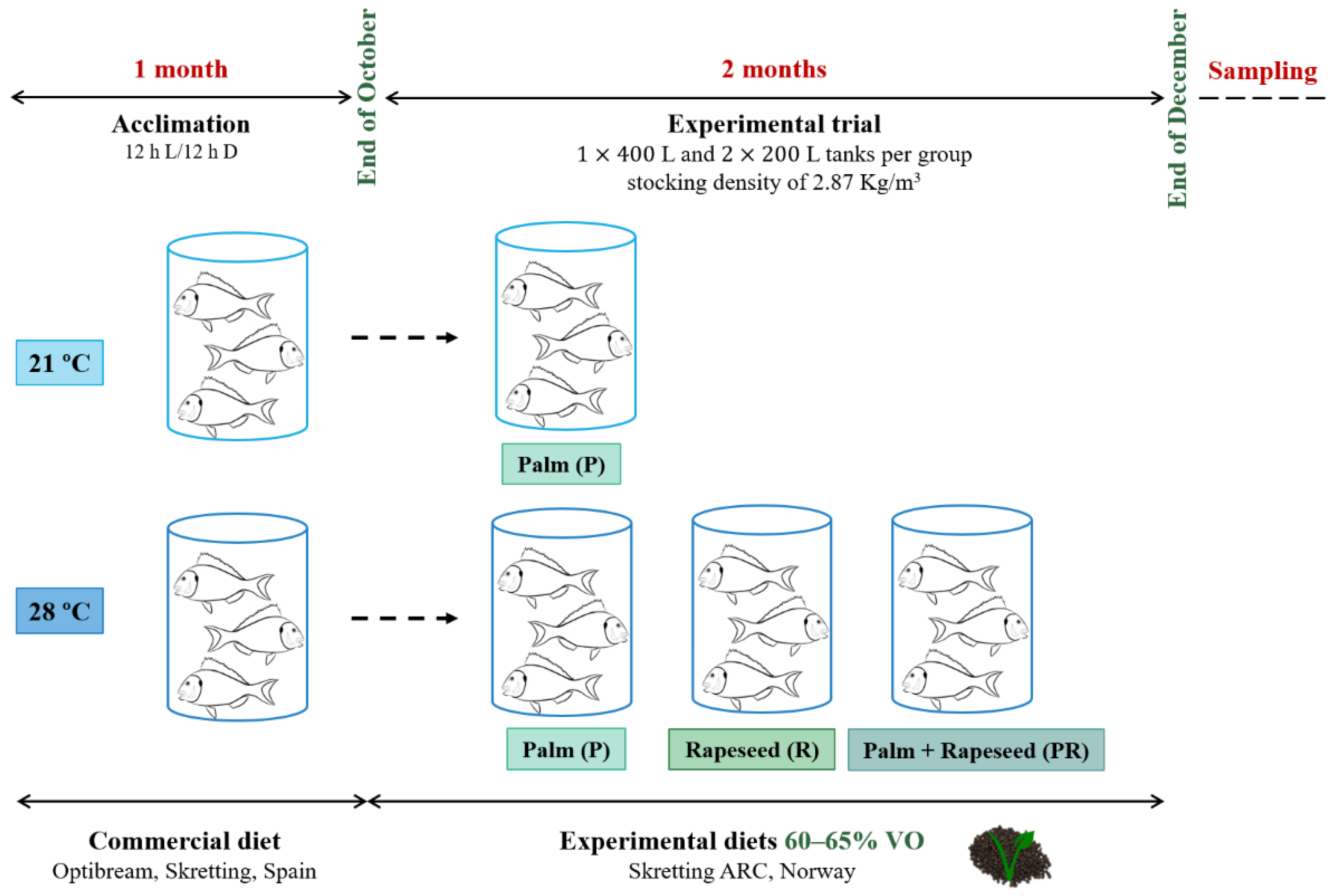
2.1. Animals, Experimental Diets and Ethics Statement
2.2. Biometric Parameters
2.3. Gene Expression Analyses
2.3.1. RNA Extraction and cDNA Synthesis
2.3.2. Real-Time Quantitative PCR (qPCR)
2.4. Statistical Analyses
3. Results
3.1. Somatic Growth Parameters in Response to Temperature and Diet
3.2. Effects of a High Rearing Temperature in Bone and White Skeletal Muscle
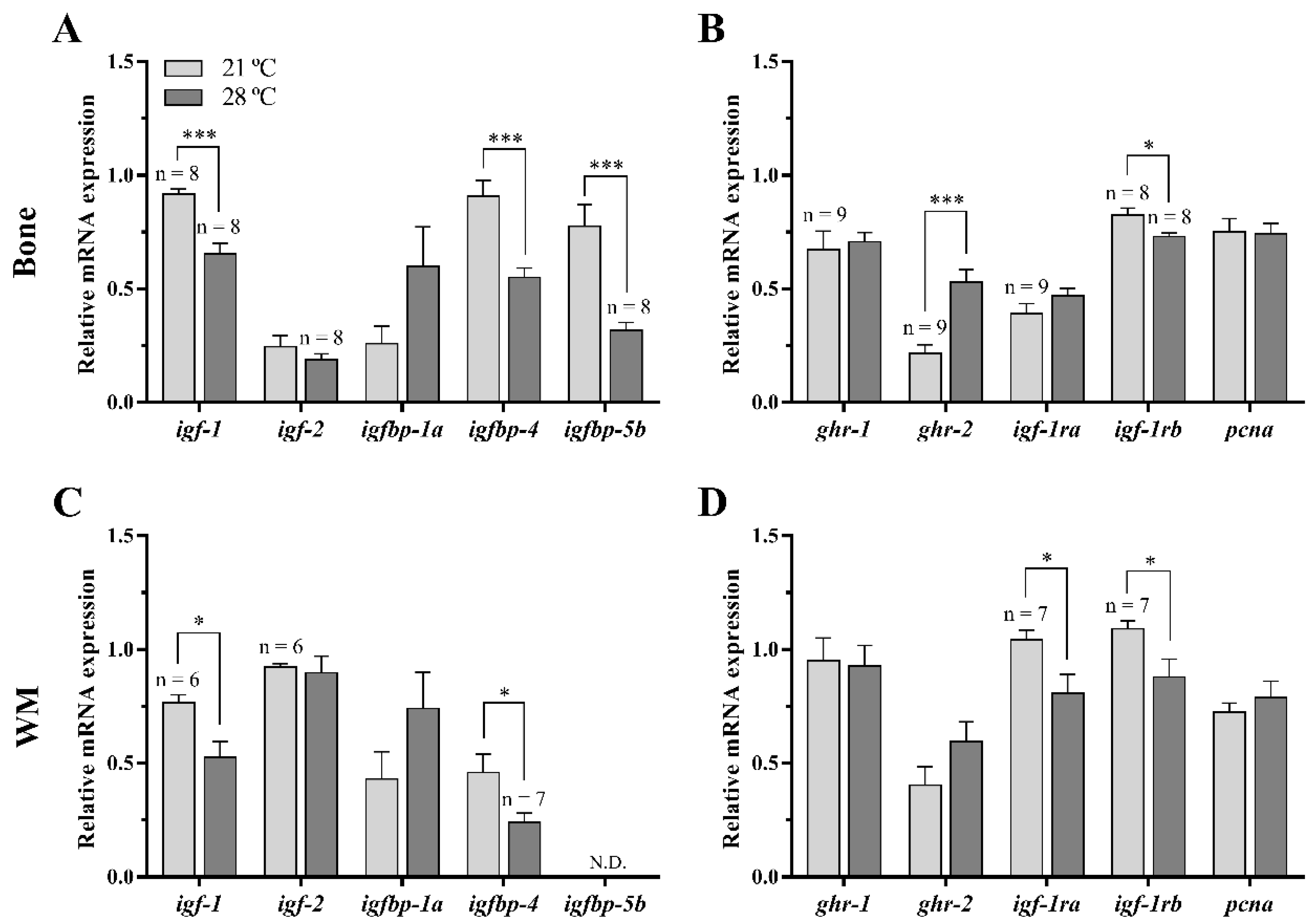
3.2.1. GH-IGFs Axis Members and a Proliferation Marker
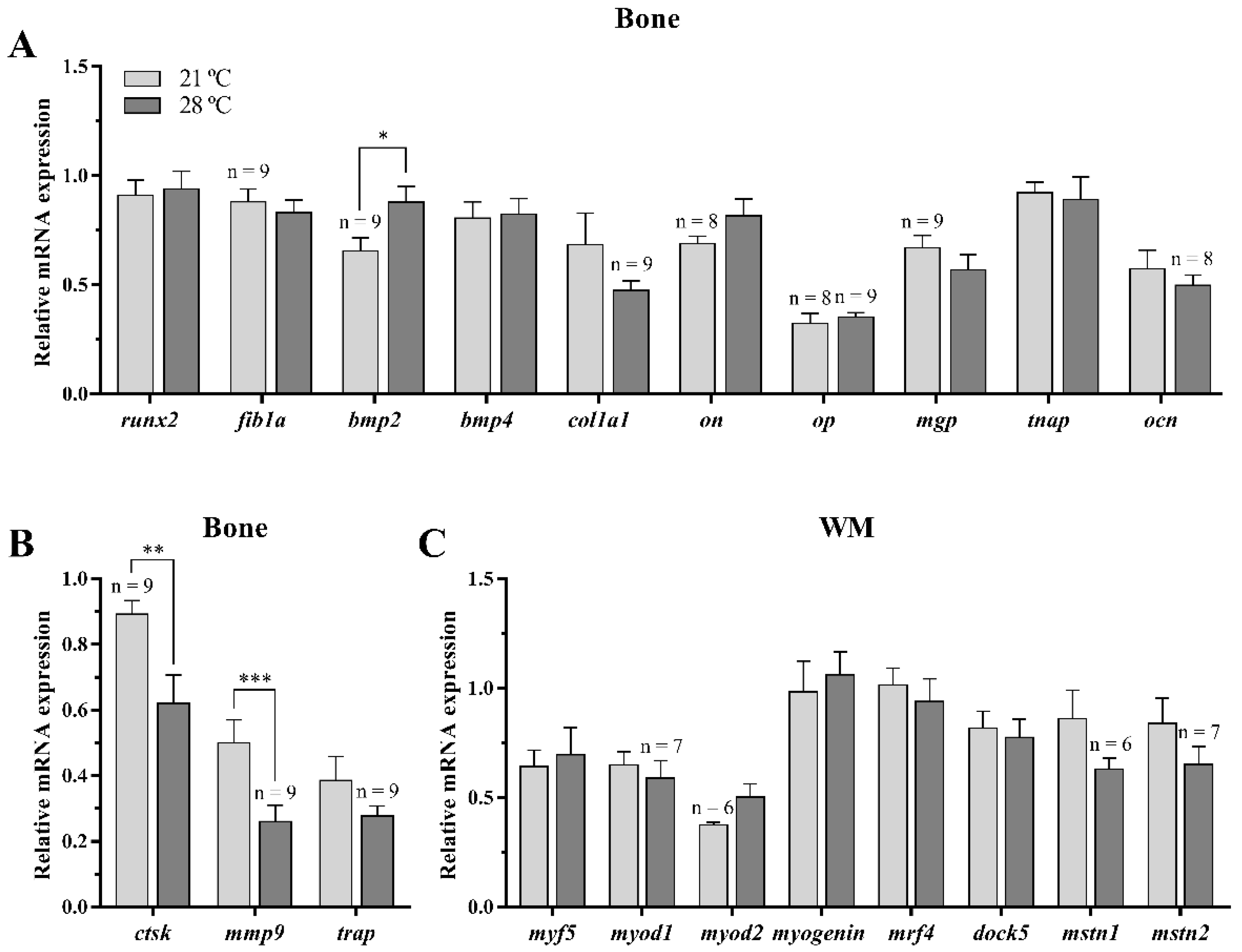
3.2.2. Osteogenic, Osteoclastic, and Myogenic Regulatory Factors
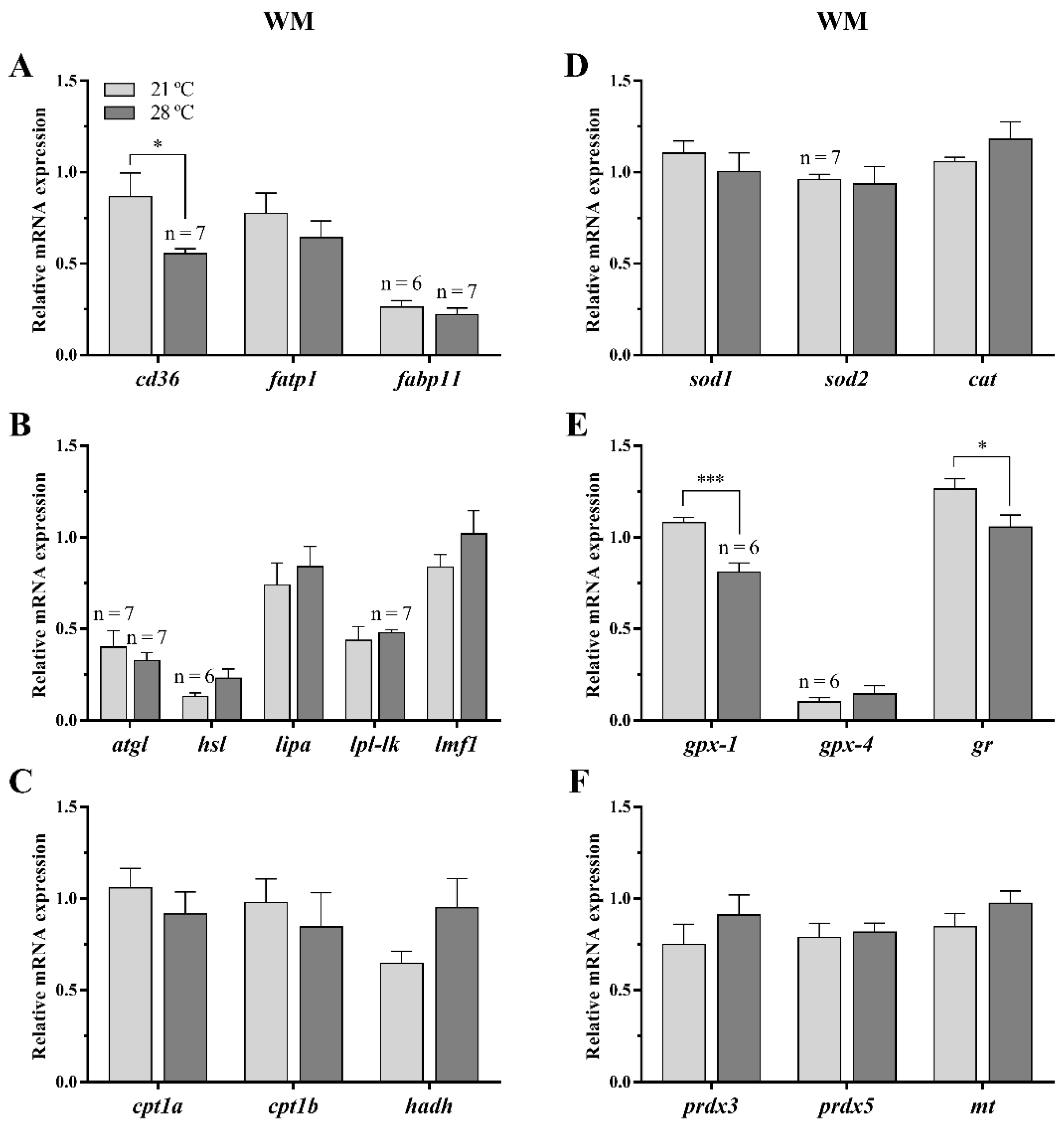
3.2.3. Lipid Metabolism and Oxidative Stress-Related Genes
3.3. Effects of VO-Based Diets at a High Rearing Temperature in Bone and White Skeletal Muscle
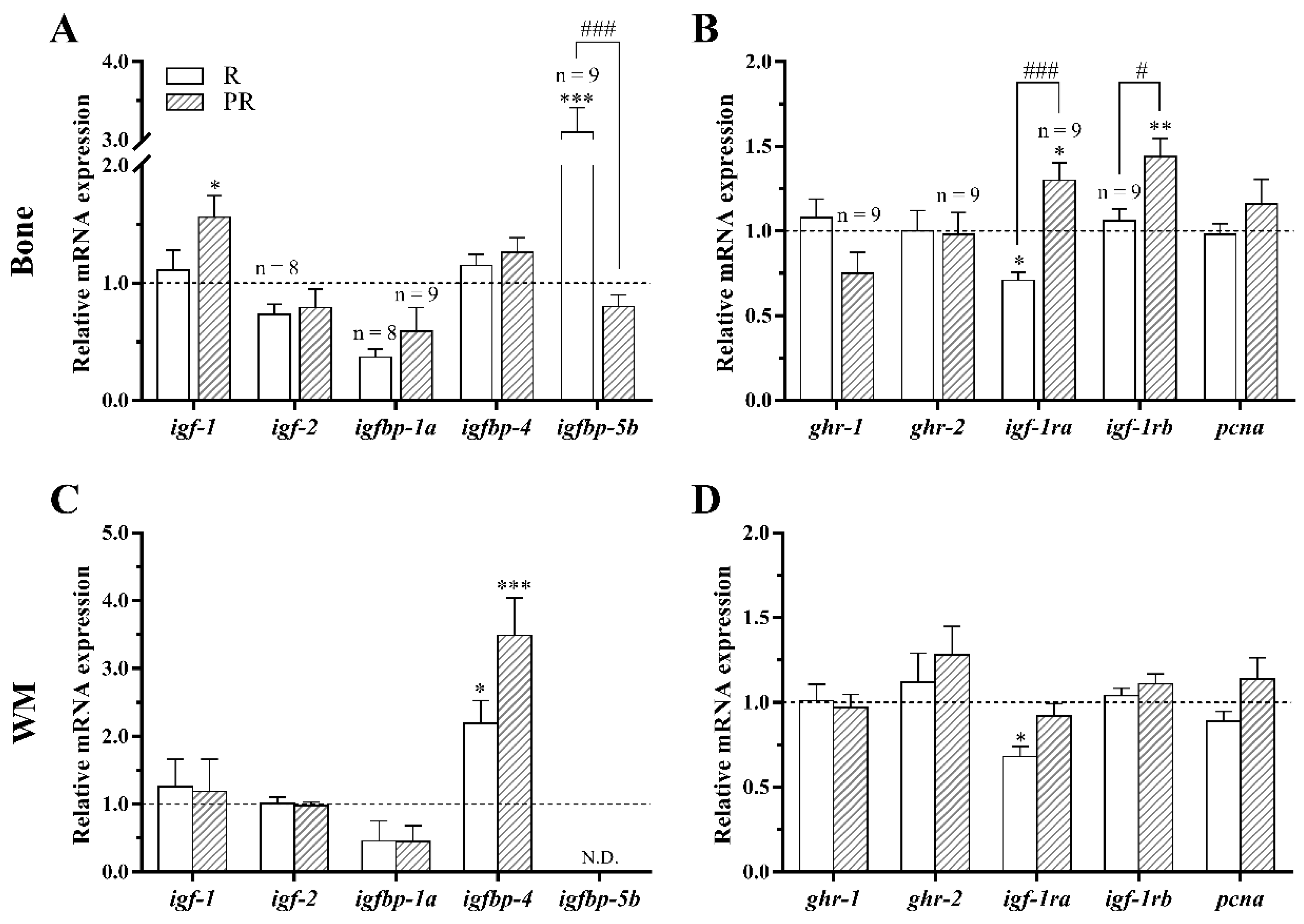
3.3.1. GH-IGFs Axis Members and a Proliferation Marker
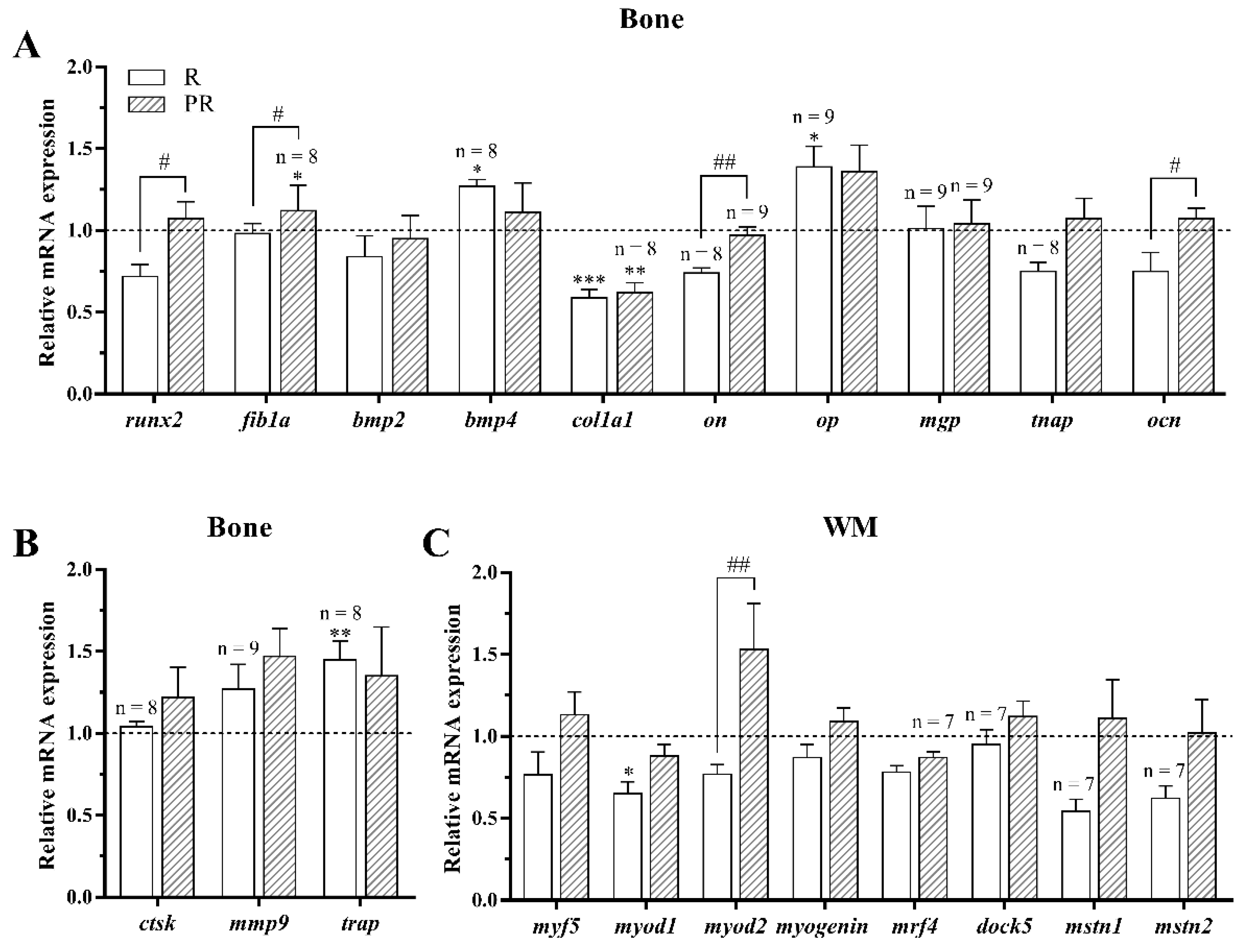
3.3.2. Osteogenic, Osteoclastic and Myogenic Regulatory Factors
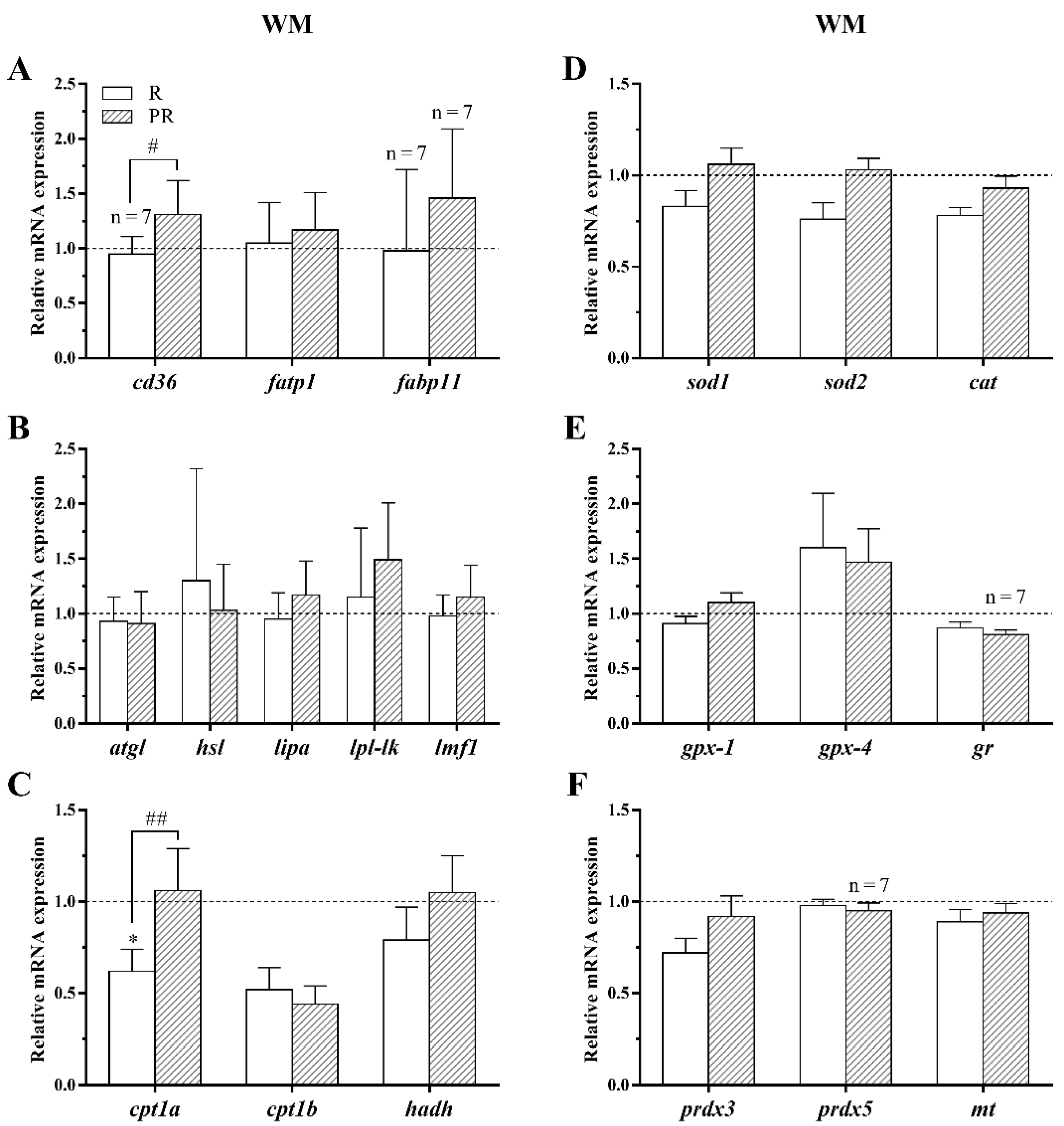
3.3.3. Lipid Metabolism and Oxidative Stress-Related Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- De Silva, S.S.; Francis, D.S.; Tacon, A.G.J. Fish oils in aquaculture: In retrospect. In Fish Oil Replacement and Alternative Lipid Sources in Aquaculture Feeds; Turchini, G.M., Ng, W.-K., Tocher, D.R., Eds.; CRC Press: Boca Raton, FL, USA, 2010; pp. 1–20. ISBN 9781439808634. [Google Scholar]
- The State of World Fisheries and Aquaculture 2020, Sustainability in Action; FAO: Rome, Italy, 2020; ISBN 9789251326923.
- Oliva-Teles, A.; Enes, P.; Peres, H. Replacing fishmeal and fish oil in industrial aquafeeds for carnivorous fish. In Feed and Feeding Practices in Aquaculture; David, D.A., Ed.; Woodhead Publishing: Cambridge, UK, 2015; pp. 203–233. ISBN 9780081005064. [Google Scholar]
- Nasopoulou, C.; Zabetakis, I. Benefits of fish oil replacement by plant originated oils in compounded fish feeds. A review. Lebensm. Wiss. Technol. Food Sci. Technol. 2012, 47, 217–224. [Google Scholar] [CrossRef]
- Turchini, G.M.; Torstensen, B.E.; Ng, W.-K. Fish oil replacement in finfish nutrition. Rev. Aquac. 2009, 1, 10–57. [Google Scholar] [CrossRef]
- Basiron, Y. Palm oil production through sustainable plantations. Eur. J. Lipid Sci. Technol. 2007, 109, 289–295. [Google Scholar] [CrossRef]
- Ayompe, L.M.; Schaafsma, M.; Egoh, B.N. Towards sustainable palm oil production: The positive and negative impacts on ecosystem services and human wellbeing. J. Clean. Prod. 2021, 278, 123914. [Google Scholar] [CrossRef]
- Bell, J.G.; McEvoy, J.; Tocher, D.R.; McGhee, F.; Campbell, P.J.; Sargent, J.R. Replacement of fish oil with rapeseed oil in diets of Atlantic salmon (Salmo salar) affects tissue lipid compositions and hepatocyte fatty acid metabolism. J. Nutr. 2001, 131, 1535–1543. [Google Scholar] [CrossRef] [Green Version]
- Komilus, C.; Shichi, N.; Koshio, S.; Ishikawa, M.; Yokoyama, S.; Michael, F.; Gao, J.; Makita, C. Influences of palm oil blended with fish oil on growth performances and lipid profiles of red sea bream Pagrus major. Aquac. Sci. 2008, 56, 317–326. [Google Scholar] [CrossRef]
- Fountoulaki, E.; Vasilaki, A.; Hurtado, R.; Grigorakis, K.; Karacostas, I.; Nengas, I.; Rigos, G.; Kotzamanis, Y.; Venou, B.; Alexis, M.N. Fish oil substitution by vegetable oils in commercial diets for gilthead sea bream (Sparus aurata L.); effects on growth performance, flesh quality and fillet fatty acid profile. Recovery of fatty acid profiles by a fish oil finishing diet under fluctuating water temperatures. Aquaculture 2009, 289, 317–326. [Google Scholar] [CrossRef]
- Ayisi, C.L.; Zhao, J.; Rupia, E.J. Growth performance, feed utilization, body and fatty acid composition of Nile tilapia (Oreochromis niloticus) fed diets containing elevated levels of palm oil. Aquac. Fish. 2017, 2, 67–77. [Google Scholar] [CrossRef]
- Caballero, M.J.; Obach, A.; Rosenlund, G.; Montero, D.; Gisvold, M.; Izquierdo, M.S. Impact of different dietary lipid sources on growth, lipid digestibility, tissue fatty acid composition and histology of rainbow trout, Oncorhynchus mykiss. Aquaculture 2002, 214, 253–271. [Google Scholar] [CrossRef]
- Izquierdo, M.S.; Montero, D.; Robaina, L.; Caballero, M.J.; Rosenlund, G.; Ginés, R. Alterations in fillet fatty acid profile and flesh quality in gilthead seabream (Sparus aurata) fed vegetable oils for a long term period. Recovery of fatty acid profiles by fish oil feeding. Aquaculture 2005, 250, 431–444. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Moya, A.; García-Meilán, I.; Riera-Heredia, N.; Vélez, E.J.; Lutfi, E.; Fontanillas, R.; Gutiérrez, J.; Capilla, E.; Navarro, I. Effects of different dietary vegetable oils on growth and intestinal performance, lipid metabolism and flesh quality in gilthead sea bream. Aquaculture 2020, 519. [Google Scholar] [CrossRef]
- Betancor, M.B.; Sprague, M.; Montero, D.; Usher, S.; Sayanova, O.; Campbell, P.J.; Napier, J.A.; Caballero, M.J.; Izquierdo, M.; Tocher, D.R. Replacement of marine fish oil with de novo Omega-3 oils from transgenic camelina sativa in feeds for gilthead sea bream (Sparus aurata L.). Lipids 2016, 51, 1171–1191. [Google Scholar] [CrossRef] [PubMed]
- Mourente, G.; Bell, J.G. Partial replacement of dietary fish oil with blends of vegetable oils (rapeseed, linseed and palm oils) in diets for European sea bass (Dicentrarchus labrax L.) over a long term growth study: Effects on muscle and liver fatty acid composition and effectiveness of a fish oil finishing diet. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2006, 145, 389–399. [Google Scholar] [CrossRef] [PubMed]
- Piedecausa, M.A.; Mazón, M.J.; García García, B.; Hernández, M.D. Effects of total replacement of fish oil by vegetable oils in the diets of sharpsnout seabream (Diplodus puntazzo). Aquaculture 2007, 263, 211–219. [Google Scholar] [CrossRef]
- Torstensen, B.E.; Espe, M.; Sanden, M.; Stubhaug, I.; Waagbø, R.; Hemre, G.I.; Fontanillas, R.; Nordgarden, U.; Hevrøy, E.M.; Olsvik, P.; et al. Novel production of Atlantic salmon (Salmo salar) protein based on combined replacement of fish meal and fish oil with plant meal and vegetable oil blends. Aquaculture 2008, 285, 193–200. [Google Scholar] [CrossRef]
- Benedito-Palos, L.; Saera-Vila, A.; Calduch-Giner, J.A.; Kaushik, S.; Pérez-Sánchez, J. Combined replacement of fish meal and oil in practical diets for fast growing juveniles of gilthead sea bream (Sparus aurata L.): Networking of systemic and local components of GH/IGF axis. Aquaculture 2007, 267, 199–212. [Google Scholar] [CrossRef] [Green Version]
- Bouraoui, L.; Sánchez-Gurmaches, J.; Cruz-Garcia, L.; Gutiérrez, J.; Benedito-Palos, L.; Pérez-Sánchez, J.; Navarro, I. Effect of dietary fish meal and fish oil replacement on lipogenic and lipoprotein lipase activities and plasma insulin in gilthead sea bream (Sparus aurata). Aquac. Nutr. 2011, 17, 54–63. [Google Scholar] [CrossRef] [Green Version]
- Montero, D.; Mathlouthi, F.; Tort, L.; Afonso, J.M.; Torrecillas, S.; Fernández-Vaquero, A.; Negrin, D.; Izquierdo, M.S. Replacement of dietary fish oil by vegetable oils affects humoral immunity and expression of pro-inflammatory cytokines genes in gilthead sea bream Sparus aurata. Fish Shellfish Immunol. 2010, 29, 1073–1081. [Google Scholar] [CrossRef]
- Riera-Heredia, N.; Sánchez-Moya, A.; Balbuena-Pecino, S.; Fontanillas, R.; Gutiérrez, J.; Capilla, E.; Navarro, I. The combination of palm and rapeseed oils emerges as a good dietary alternative for optimal growth and balanced lipid accumulation in juvenile gilthead sea bream reared at an elevated temperature. Aquaculture 2020, 526. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2014: Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Pachauri, R.K., Meyer, L.A., Eds.; Core Writing Team: Geneva, Switzerland, 2015. [Google Scholar]
- Pettersson, A.; Pickova, J.; Brännäs, E. Swimming performance at different temperatures and fatty acid composition of Arctic charr (Salvelinus alpinus) fed palm and rapeseed oils. Aquaculture 2010, 300, 176–181. [Google Scholar] [CrossRef]
- Glencross, B.D. Exploring the nutritional demand for essential fatty acids by aquaculture species. Rev. Aquac. 2009, 1, 71–124. [Google Scholar] [CrossRef]
- Balbuena-Pecino, S.; Riera-Heredia, N.; Vélez, E.J.; Gutiérrez, J.; Navarro, I.; Riera-Codina, M.; Capilla, E. Temperature affects musculoskeletal development and muscle lipid metabolism of gilthead sea bream (Sparus aurata). Front. Endocrinol. 2019, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT–PCR. Nucleic Acids Res. 2001, 29, 2002–2007. [Google Scholar] [CrossRef] [PubMed]
- Reindl, K.M.; Sheridan, M.A. Peripheral regulation of the growth hormone-insulin-like growth factor system in fish and other vertebrates. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2012, 163, 231–245. [Google Scholar] [CrossRef] [PubMed]
- Beckman, B.R. Perspectives on concordant and discordant relations between insulin-like growth factor 1 (IGF1) and growth in fishes. Gen. Comp. Endocrinol. 2011, 170, 233–252. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Sánchez, J.; Le Bail, P.Y. Growth hormone axis as marker of nutritional status and growth performance in fish. Aquaculture 1999, 177, 117–128. [Google Scholar] [CrossRef]
- Vélez, E.J.; Perelló, M.; Azizi, S.; Moya, A.; Lutfi, E.; Pérez-Sánchez, J.; Calduch-Giner, J.A.; Navarro, I.; Blasco, J.; Fernández-Borràs, J.; et al. Recombinant bovine growth hormone (rBGH) enhances somatic growth by regulating the GH-IGF axis in fingerlings of gilthead sea bream (Sparus aurata). Gen. Comp. Endocrinol. 2018, 257, 192–202. [Google Scholar] [CrossRef]
- Gabillard, J.C.; Yao, K.; Vandeputte, M.; Guttierez, J.; Le Bail, P.Y. Differential expression of two GH receptor mRNAs following temperature change in rainbow trout (Oncorhynchus mykiss). J. Endocrinol. 2006, 190, 29–37. [Google Scholar] [CrossRef]
- Li, M.; Greenaway, J.; Raine, J.; Petrik, J.; Hahnel, A.; Leatherland, J. Growth hormone and insulin-like growth factor gene expression prior to the development of the pituitary gland in rainbow trout (Oncorhynchus mykiss) embryos reared at two temperatures. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2006, 143, 514–522. [Google Scholar] [CrossRef]
- Duan, C.; Ren, H.; Gao, S. Insulin-like growth factors (IGFs), IGF receptors, and IGF-binding proteins: Roles in skeletal muscle growth and differentiation. Gen. Comp. Endocrinol. 2010, 167, 344–351. [Google Scholar] [CrossRef]
- Witten, P.E.; Huysseune, A. A comparative view on mechanisms and functions of skeletal remodelling in teleost fish, with special emphasis on osteoclasts and their function. Biol. Rev. 2009, 84, 315–346. [Google Scholar] [CrossRef] [PubMed]
- Crane, J.L.; Cao, X. Function of matrix IGF-1 in coupling bone resorption and formation. J. Mol. Med. 2014, 92, 107–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohlsson, C.; Bengtsson, B.-A.; Isaksson, O.G.P.; Andreassen, T.T.; Slootweg, M.C. Growth hormone and bone. Endocr. Rev. 1998, 19, 55–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abe, E.; Yamamoto, M.; Taguchi, Y.; Lecka-Czernik, B.; O’Brien, C.A.; Economides, A.N.; Stahl, N.; Jilka, R.L.; Manolagas, S.C. Essential requirement of BMPs-2/4 for both osteoblast and osteoclast formation in murine bone marrow cultures from adult mice: Antagonism by noggin. J. Bone Miner. Res. 2000, 15, 663–673. [Google Scholar] [CrossRef] [PubMed]
- Itoh, K.; Udagawa, N.; Katagiri, T.; Iemura, S.; Ueno, N.; Yasuda, H.; Higashio, K.; Quinn, J.M.W.; Gillespie, M.T.; Martin, T.J.; et al. Bone morphogenetic protein 2 stimulates osteoclast differentiation and survival supported by receptor activator of nuclear factor-κB ligand. Endocrinology 2001, 142, 3656–3662. [Google Scholar] [CrossRef]
- Yamaguchi, A.; Katagiri, T.; Ikeda, T.; Wozney, J.M.; Rosen, V.; Wang, E.A.; Kahn, A.J.; Suda, T.; Yoshiki, S. Recombinant human bone morphogenetic protein-2 stimulates osteoblastic maturation and inhibits myogenic differentiation in vitro. J. Cell Biol. 1991, 113, 681–687. [Google Scholar] [CrossRef] [Green Version]
- Kamiya, Y.; Kamiya, M.; Tanaka, M. The effect of high ambient temperature on Ca, P and Mg balance and bone turnover in high-yielding dairy cows. Anim. Sci. J. 2010, 81, 482–486. [Google Scholar] [CrossRef]
- Couto, A.; Enes, P.; Peres, H.; Oliva-Teles, A. Effect of water temperature and dietary starch on growth and metabolic utilization of diets in gilthead sea bream (Sparus aurata) juveniles. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2008, 151, 45–50. [Google Scholar] [CrossRef]
- Handeland, S.O.; Björnsson, B.T.; Arnesen, A.M.; Stefansson, S.O. Seawater adaptation and growth of post-smolt Atlantic salmon (Salmo salar) of wild and farmed strains. Aquaculture 2003, 220, 367–384. [Google Scholar] [CrossRef]
- Handeland, S.O.; Imsland, A.K.; Stefansson, S.O. The effect of temperature and fish size on growth, feed intake, food conversion efficiency and stomach evacuation rate of Atlantic salmon post-smolts. Aquaculture 2008, 283, 36–42. [Google Scholar] [CrossRef]
- Guerreiro, I.; Peres, H.; Castro-Cunha, M.; Oliva-Teles, A. Effect of temperature and dietary protein/lipid ratio on growth performance and nutrient utilization of juvenile Senegalese sole (Solea senegalensis). Aquac. Nutr. 2012, 18, 98–106. [Google Scholar] [CrossRef]
- Person-Le Ruyet, J.; Mahé, K.; Le Bayon, N.; Le Delliou, H. Effects of temperature on growth and metabolism in a Mediterranean population of European sea bass, Dicentrarchus labrax. Aquaculture 2004, 237, 269–280. [Google Scholar] [CrossRef] [Green Version]
- Neuheimer, A.B.; Thresher, R.E.; Lyle, J.M.; Semmens, J.M. Tolerance limit for fish growth exceeded by warming waters. Nat. Clim. Chang. 2011, 1, 110–113. [Google Scholar] [CrossRef]
- Wang, T.; Overgaard, J. The heartbreak of adapting to global warming. Science 2007, 315, 49–50. [Google Scholar] [CrossRef]
- Montero, D.; Robaina, L.; Caballero, M.J.; Ginés, R.; Izquierdo, M.S. Growth, feed utilization and flesh quality of European sea bass (Dicentrarchus labrax) fed diets containing vegetable oils: A time-course study on the effect of a re-feeding period with a 100% fish oil diet. Aquaculture 2005, 248, 121–134. [Google Scholar] [CrossRef] [Green Version]
- Ng, W.K.; Sigholt, T.; Bell, J.G. The influence of environmental temperature on the apparent nutrient and fatty acid digestibility in Atlantic salmon (Salmo salar L.) fed finishing diets containing different blends of fish oil, rapeseed oil and palm oil. Aquac. Res. 2004, 35, 1228–1237. [Google Scholar] [CrossRef]
- Peng, X.; Li, F.; Lin, S.; Chen, Y. Effects of total replacement of fish oil on growth performance, lipid metabolism and antioxidant capacity in tilapia (Oreochromis niloticus). Aquac. Int. 2016, 24, 145–156. [Google Scholar] [CrossRef]
- Sun, S.; Ye, J.; Chen, J.; Wang, Y.; Chen, L. Effect of dietary fish oil replacement by rapeseed oil on the growth, fatty acid composition and serum non-specific immunity response of fingerling black carp, Mylopharyngodon piceus. Aquac. Nutr. 2011, 17, 441–450. [Google Scholar] [CrossRef]
- Riera-Heredia, N.; Martins, R.; Mateus, A.P.; Costa, R.A.; Gisbert, E.; Navarro, I.; Gutiérrez, J.; Power, D.M.; Capilla, E. Temperature responsiveness of gilthead sea bream bone; an in vitro and in vivo approach. Sci. Rep. 2018, 8, 1–14. [Google Scholar] [CrossRef]
- García de la serrana, D.; Codina, M.; Capilla, E.; Jiménez-Amilburu, V.; Navarro, I.; Du, S.J.; Johnston, I.A.; Gutiérrez, J. Characterisation and expression of myogenesis regulatory factors during in vitro myoblast development and in vivo fasting in the gilthead sea bream (Sparus aurata). Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2014, 167, 90–99. [Google Scholar] [CrossRef] [PubMed]
- Norambuena, F.; Morais, S.; Emery, J.A.; Turchini, G.M. Arachidonic acid and eicosapentaenoic acid metabolism in juvenile Atlantic salmon as affected by water temperature. PLoS ONE 2015, 10, e0143622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menoyo, D.; Izquierdo, M.S.; Robaina, L.; Ginés, R.; Lopez-Bote, C.J.; Bautista, J.M. Adaptation of lipid metabolism, tissue composition and flesh quality in gilthead sea bream (Sparus aurata) to the replacement of dietary fish oil by linseed and soyabean oils. Br. J. Nutr. 2004, 92, 41–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ofori-Mensah, S.; Yıldız, M.; Arslan, M.; Eldem, V. Fish oil replacement with different vegetable oils in gilthead seabream, Sparus aurata diets: Effects on fatty acid metabolism based on whole-body fatty acid balance method and genes expression. Aquaculture 2020, 529. [Google Scholar] [CrossRef]
- Stubhaug, I.; Frøyland, L.; Torstensen, B.E. β-oxidation capacity of red and white muscle and liver in Atlantic salmon (Salmo salar L.)—Effects of increasing dietary rapeseed oil and olive oil to replace capelin oil. Lipids 2005, 40, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Torstensen, B.E.; Øyvind, L.; Frøyland, L. Lipid metabolism and tissue composition in Atlantic salmon (Salmo salar L.)—Effects of capelin oil, palm oil, and oleic acid-enriched sunflower oil as dietary lipid sources. Lipids 2000, 35, 653–664. [Google Scholar] [CrossRef] [PubMed]
- Torstensen, B.E.; Tocher, D.R. The effects of fish oil replacement on lipid metabolism of fish. In Fish Oil Replacement and Alternative Lipid Sources in Aquaculture Feeds; Turchini, G.M., Ng, W.-K., Tocher, D.R., Eds.; CRC Press: Boca Raton, FL, USA, 2010; pp. 405–437. ISBN 9781439808634. [Google Scholar]
- Sies, H.; Berndt, C.; Jones, D.P. Oxidative stress. Annu. Rev. Biochem. 2017, 86, 715–748. [Google Scholar] [CrossRef]
- Feidantsis, K.; Pörtner, H.O.; Vlachonikola, E.; Antonopoulou, E.; Michaelidis, B. Seasonal changes in metabolism and cellular stress phenomena in the gilthead sea bream (Sparus aurata). Physiol. Biochem. Zool. 2018, 91, 878–895. [Google Scholar] [CrossRef]
- Bagnyukova, T.V.; Lushchak, O.V.; Storey, K.B.; Lushchak, V.I. Oxidative stress and antioxidant defense responses by goldfish tissues to acute change of temperature from 3 to 23 °C. J. Therm. Biol. 2007, 32, 227–234. [Google Scholar] [CrossRef]
- Brigelius-Flohé, R. Tissue-specific functions of individual glutathione peroxidases. Free Radic. Biol. Med. 1999, 27, 951–965. [Google Scholar] [CrossRef]
- Martin, H.L.; Teismann, P. Glutathione—A review on its role and significance in Parkinson’s disease. FASEB J. 2009, 23, 3263–3272. [Google Scholar] [CrossRef] [Green Version]
- Kaur, M.; Atif, F.; Ali, M.; Rehman, H.; Raisuddin, S. Heat stress-induced alterations of antioxidants in the freshwater fish Channa punctata Bloch. J. Fish Biol. 2005, 67, 1653–1665. [Google Scholar] [CrossRef]
- Ploch, S.A.; Lee, Y.P.; MacLean, E.; Di Giulio, R.T. Oxidative stress in liver of brown bullhead and channel catfish following exposure to tert-butyl hydroperoxide. Aquat. Toxicol. 1999, 46, 231–240. [Google Scholar] [CrossRef]
- Monahan, F. Oxidation of lipids in muscle foods: Fundamentals and applied concerns. In Antioxidants in Muscle Foods: Nutritional Strategies to Improve Quality; Decker, E., Faustman, C., Lopez-Bote, C., Eds.; Wiley Interscience: New York, NY, USA, 2000; pp. 3–24. [Google Scholar]
- Grim, J.M.; Simonik, E.A.; Semones, M.C.; Kuhn, D.E.; Crockett, E.L. The glutathione-dependent system of antioxidant defense is not modulated by temperature acclimation in muscle tissues from striped bass, Morone saxatilis. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2013, 164, 383–390. [Google Scholar] [CrossRef]
- Vinagre, C.; Madeira, D.; Narciso, L.; Cabral, H.N.; Diniz, M. Effect of temperature on oxidative stress in fish: Lipid peroxidation and catalase activity in the muscle of juvenile seabass, Dicentrarchus labrax. Ecol. Indic. 2012, 23, 274–279. [Google Scholar] [CrossRef]
- Lushchak, V.I.; Bagnyukova, T.V. Temperature increase results in oxidative stress in goldfish tissues. 1. Indices of oxidative stress. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2006, 143, 30–35. [Google Scholar] [CrossRef]
- Castro, C.; Firmino, A.; Coutinho, F.; Panserat, S.; Corraze, G.; Pérez-jiménez, A.; Peres, H.; Oliva-teles, A. Liver and intestine oxidative status of gilthead sea bream fed vegetable oil and carbohydrate rich diets. Aquaculture 2016, 464, 665–672. [Google Scholar] [CrossRef]
- Mu, H.; Wei, C.; Zhang, Y.; Zhou, H.; Pan, Y.; Chen, J.; Zhang, W.; Mai, K. Impacts of replacement of dietary fish oil by vegetable oils on growth performance, anti-oxidative capacity, and inflammatory response in large yellow croaker Larimichthys crocea. Fish Physiol. Biochem. 2020, 46, 231–245. [Google Scholar] [CrossRef]
- Kutluyer, F.; Sirkecioǧlu, A.N.; Aksakal, E.; Aksakal, F.I.; Tunç, A.; Günaydin, E. Effect of dietary fish oil replacement with plant oils on growth performance and gene expression in juvenile rainbow trout (Oncorhynchus mykiss). Ann. Anim. Sci. 2017, 17, 1135–1153. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients (%) | P | R | PR |
| Wheat | 6.83 | 6.83 | 6.83 |
| Corn meal | 10 | 10 | 10 |
| Wheat gluten | 14.79 | 14.78 | 14.78 |
| Soya protein | 25 | 25 | 25 |
| Broad beans | 10 | 10 | 10 |
| Fishmeal | 20 | 20 | 20 |
| Fish oil | 5.34 | 5.34 | 4.45 |
| Linseed oil | 1.33 | 2 | 1.33 |
| Rapeseed oil | 0 | 6.67 | 3.34 |
| Palm oil | 6.67 | 0 | 3.55 |
| Phosphate | 0.03 | 0.03 | 0.03 |
| Composition (%) | |||
| Moisture | 7.6 | 7.7 | 7.5 |
| Protein | 51.9 | 51.8 | 51.7 |
| Fat | 19.2 | 18.9 | 19.4 |
| Ash | 5.7 | 5.7 | 5.6 |
| 21 °C | 28 °C | |||
|---|---|---|---|---|
| Biometrics | P | P | R | PR |
| IBW (g) | 24.21 ± 0.57 | 22.89 ± 0.40 | 22.80 ± 0.75 | 22.24 ± 0.77 |
| FBW (g) | 54.32 ± 1.78 * | 68.02 ± 2.34 | 63.33 ± 2.09 | 62.21 ± 3.80 |
| WG (%) | 124.3 ± 5.19 * | 197.5 ± 13.27 | 177.8 ± 2.33 | 179.2 ± 7.62 |
| SGR (%) | 1.52 ± 0.04 * | 2.05 ± 0.08 | 1.93 ± 0.02 | 1.94 ± 0.05 |
| BL (cm) | 15.06 ± 0.02 * | 16.55 ± 0.11 | 16.37 ± 0.07 | 16.23 ± 0.22 |
| CF (%) | 1.59 ± 0.06 | 1.5 ± 0.04 | 1.44 ± 0.03 | 1.45 ± 0.03 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Balbuena-Pecino, S.; Riera-Heredia, N.; Gasch-Navalón, E.; Sánchez-Moya, A.; Fontanillas, R.; Gutiérrez, J.; Navarro, I.; Capilla, E. Musculoskeletal Growth Modulation in Gilthead Sea Bream Juveniles Reared at High Water Temperature and Fed with Palm and Rapeseed Oils-Based Diets. Animals 2021, 11, 260. https://doi.org/10.3390/ani11020260
Balbuena-Pecino S, Riera-Heredia N, Gasch-Navalón E, Sánchez-Moya A, Fontanillas R, Gutiérrez J, Navarro I, Capilla E. Musculoskeletal Growth Modulation in Gilthead Sea Bream Juveniles Reared at High Water Temperature and Fed with Palm and Rapeseed Oils-Based Diets. Animals. 2021; 11(2):260. https://doi.org/10.3390/ani11020260
Chicago/Turabian StyleBalbuena-Pecino, Sara, Natàlia Riera-Heredia, Esther Gasch-Navalón, Albert Sánchez-Moya, Ramon Fontanillas, Joaquim Gutiérrez, Isabel Navarro, and Encarnación Capilla. 2021. "Musculoskeletal Growth Modulation in Gilthead Sea Bream Juveniles Reared at High Water Temperature and Fed with Palm and Rapeseed Oils-Based Diets" Animals 11, no. 2: 260. https://doi.org/10.3390/ani11020260