
Cannabinoid and Cannabinoid-Related Receptors in the Myenteric Plexus of the Porcine Ileum

 , , , and
, , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Tissue Collection
2.3. Immunofluorescence
2.4. Specificity of the Primary Antibodies
2.5. Specificity of the Secondary Antibodies
2.6. Fluorescence Microscopy
3. Results
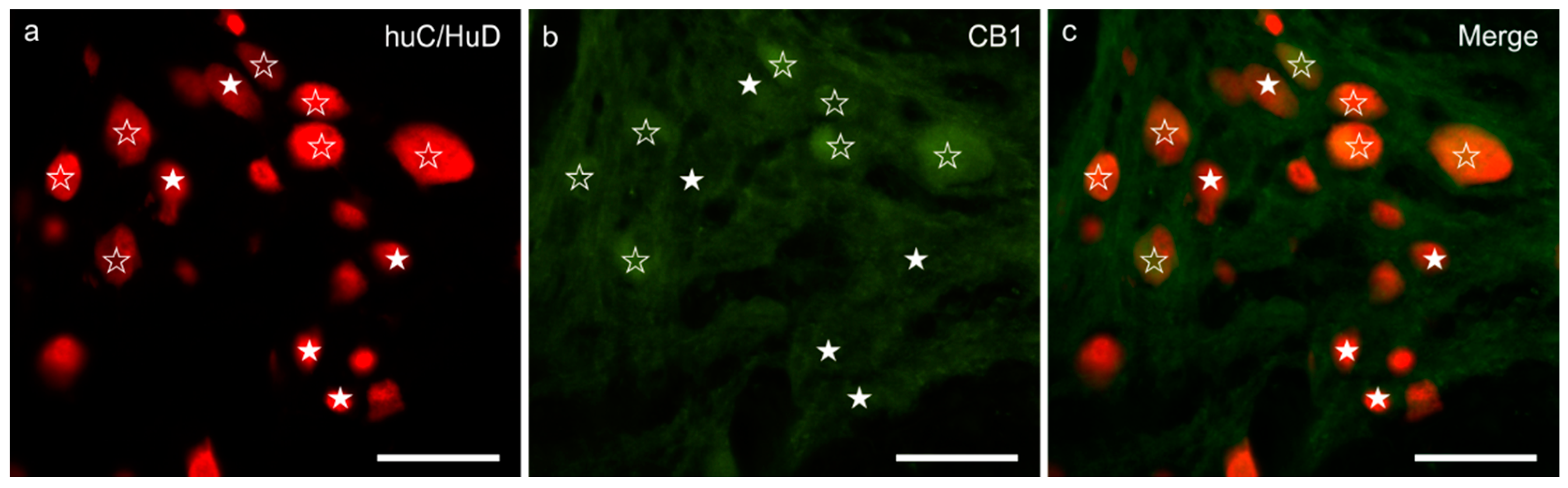
3.1. CB1R Immunoreactivity
3.2. CB2R Immunoreactivity
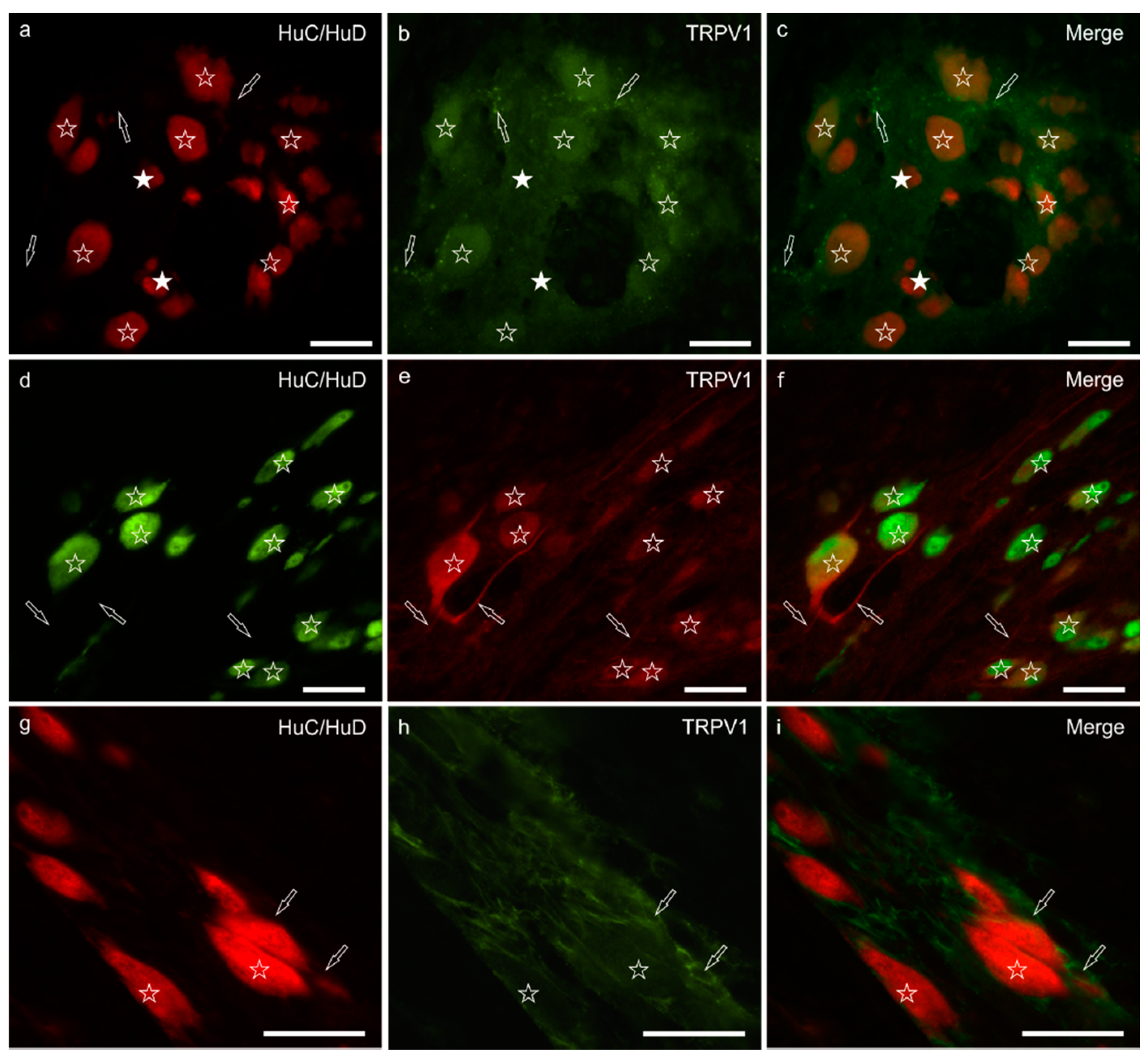
3.3. TRPV1 Immunoreactivity
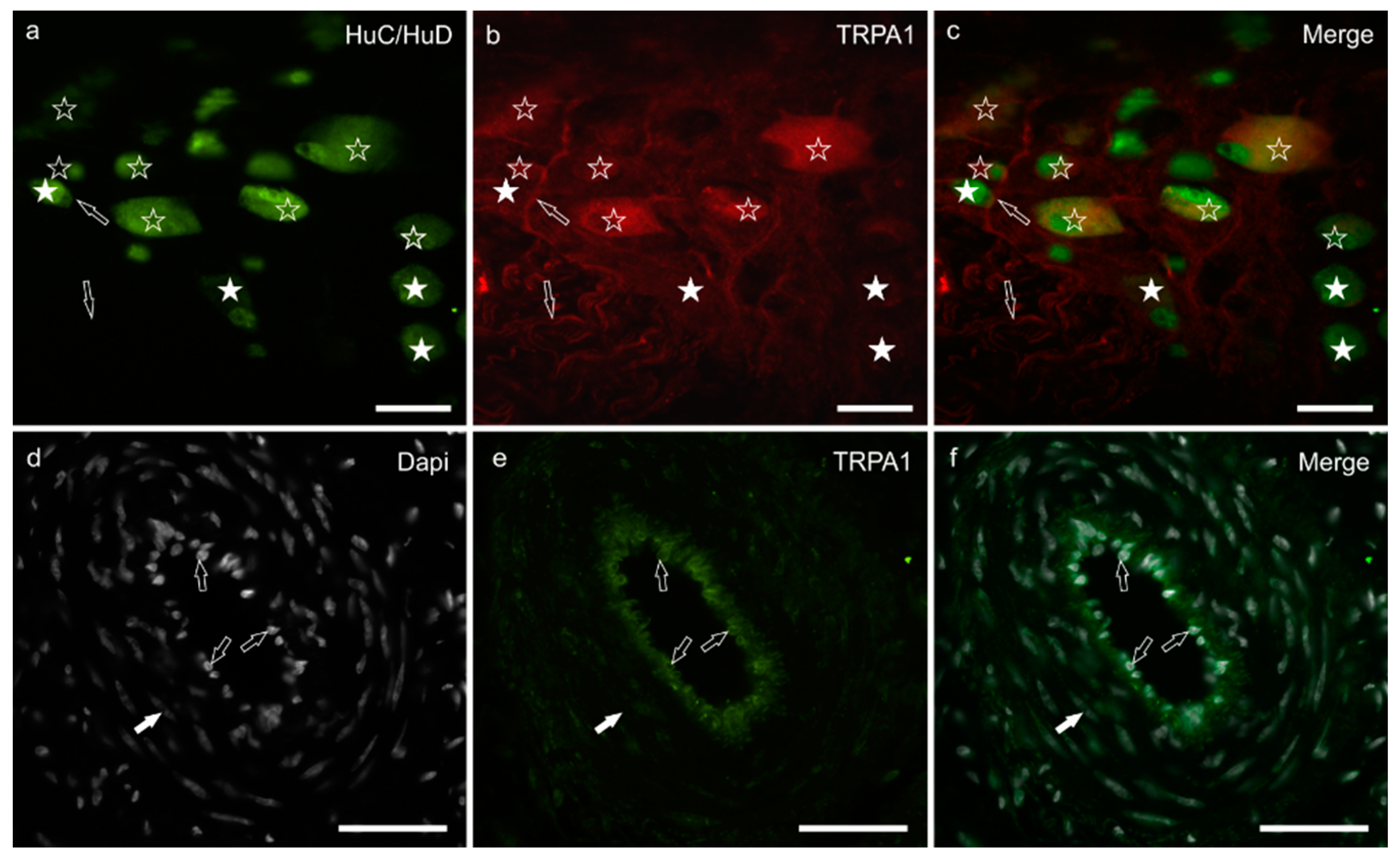
3.4. TRPA1 Immunoreactivity
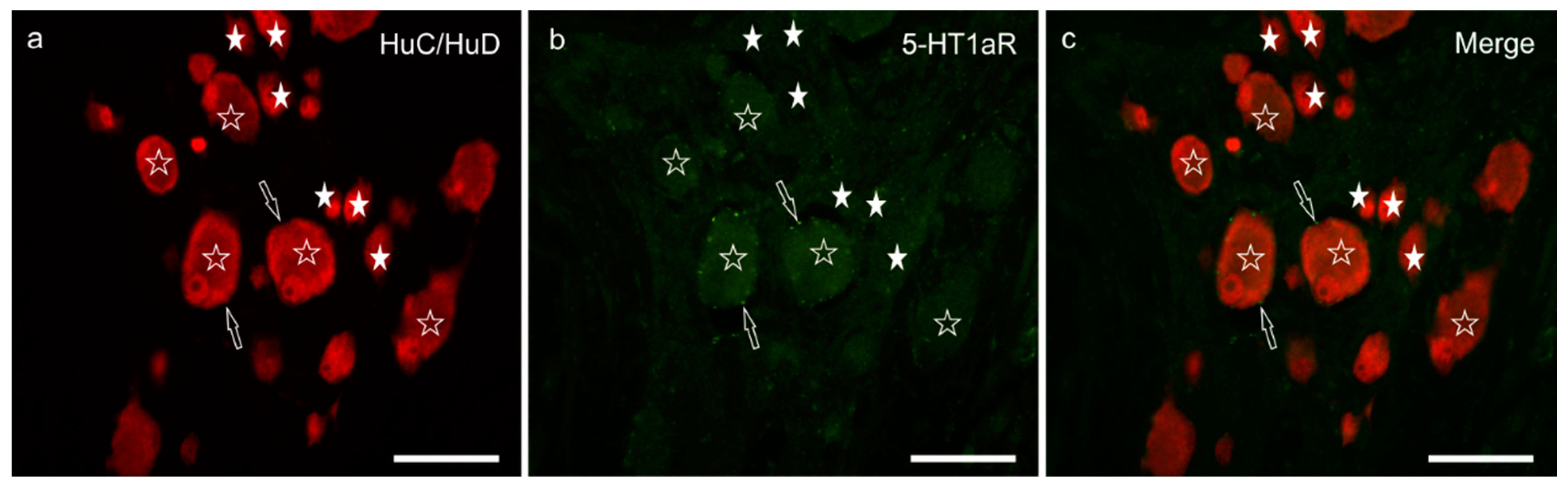
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wood, J.D. Enteric nervous system. In Encyclopedia of Gastroenterology; Johnson, L.R., Ed.; Elsevier: New York, NY, USA, 2004; pp. 701–706. ISBN 978-0-12-386860-2. [Google Scholar]
- Costa, M.; Brookes, S.J.H.; Hennig, G.W. Anatomy and physiology of the enteric nervous system. Gut 2000, 47, iv15–iv19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furness, J.B.; Callaghan, B.P.; Rivera, L.R.; Cho, H.-J. The enteric nervous system and gastrointestinal innervation: Integrated local and central control. In Microbial Endocrinology: The Microbiota-Gut-Brain Axis in Health and Disease; Lyte, M., Cryan, J.F., Eds.; Springer: New York, NY, USA, 2014; pp. 39–71. ISBN 978-1-4939-0897-4. [Google Scholar]
- Stella, N. Cannabinoid signaling in glial cells. Glia 2004, 48, 267–277. [Google Scholar] [CrossRef] [PubMed]
- Ligresti, A.; De Petrocellis, L.; Di Marzo, V. From phytocannabinoids to cannabinoid receptors and endocannabinoids: Pleiotropic physiological and pathological roles through complex pharmacology. Physiol. Rev. 2016, 96, 1593–1659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iannotti, F.A.; Di Marzo, V.; Petrosino, S. Endocannabinoids and endocannabinoid-related mediators: Targets, metabolism and role in neurological disorders. Prog. Lipid Res. 2016, 62, 107–128. [Google Scholar] [CrossRef]
- Lu, Y.; Anderson, H.D. Cannabinoid signaling in health and disease. Can. J. Physiol. Pharmacol. 2017, 95, 311–327. [Google Scholar] [CrossRef]
- Maccarrone, M.; Bab, I.; Bíró, T.; Cabral, G.A.; Dey, S.K.; Di Marzo, V.; Konje, J.C.; Kunos, G.; Mechoulam, R.; Pacher, P.; et al. Endocannabinoid signaling at the periphery: 50 years after THC. Trends Pharmacol. Sci. 2015, 36, 277–296. [Google Scholar] [CrossRef] [Green Version]
- Mackie, K. Distribution of cannabinoid receptors in the central and peripheral nervous system. In Handbook of Experimental Pharmacology; Cannabinoids; Pertwee, R.G., Ed.; Springer Berlin Heidelberg: Berlin, Heidelberg, 2005; pp. 299–325. ISBN 978-3-540-26573-3. [Google Scholar]
- Klegeris, A.; Bissonnette, C.J.; McGeer, P.L. Reduction of human monocytic cell neurotoxicity and cytokine secretion by ligands of the cannabinoid-type CB2 receptor. Br. J. Pharmacol. 2003, 139, 775–786. [Google Scholar] [CrossRef] [Green Version]
- Turcotte, C.; Blanchet, M.-R.; LaViolette, M.; Flamand, N. The CB2 receptor and its role as a regulator of inflammation. Cell. Mol. Life Sci. 2016, 73, 4449–4470. [Google Scholar] [CrossRef] [Green Version]
- Moreira, F.A.; Grieb, M.; Lutz, B. Central side-effects of therapies based on CB1 cannabinoid receptor agonists and antagonists: Focus on anxiety and depression. Best Pract. Res. Clin. Endocrinol. Metab. 2009, 23, 133–144. [Google Scholar] [CrossRef]
- Parlar, A.; Arslan, S.O.; Doğan, M.F.; Elibol, E.; Kara, H. The exogenous administration of CB2 specific agonist, GW405833, inhibits inflammation by reducing cytokine production and oxidative stress. Exp. Ther. Med. 2018, 16, 4900–4908. [Google Scholar] [CrossRef] [Green Version]
- Izzo, A.A.; Sharkey, K.A. Cannabinoids and the gut: New developments and emerging concepts. Pharmacol. Ther. 2010, 126, 21–38. [Google Scholar] [CrossRef] [PubMed]
- DiPatrizio, N.V. Endocannabinoids in the Gut. Cannabis Cannabinoid Res. 2016, 1, 67–77. [Google Scholar] [CrossRef] [PubMed]
- Vera, G.; Fichna, J.; Abalo, R. Chapter 98—Cannabinoids and effects on the gastrointestinal tract: A focus on motility. In Handbook of Cannabis and Related Pathologies; Preedy, V.R., Ed.; Academic Press: San Diego, CA, USA, 2017; pp. 947–957. ISBN 978-0-12-800756-3. [Google Scholar]
- Izzo, A.A. Cannabinoids and intestinal motility: Welcome to CB2 receptors. Br. J. Pharmacol. 2004, 142, 1201–1202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Marzo, V. Endocannabinoid overactivity and intestinal inflammation. Gut 2006, 55, 1373–1376. [Google Scholar] [CrossRef] [Green Version]
- Morales, P.; Hurst, D.P.; Reggio, P.H. Molecular Targets of the Phytocannabinoids: A complex picture. Prog. Chem. Org. Nat. Prod. 2017, 103, 103–131. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.; Yu, M.; Liu, Y.; Yu, S. TRP channel functions in the gastrointestinal tract. Semin. Immunopathol. 2016, 38, 385–396. [Google Scholar] [CrossRef]
- Kreitzer, F.R.; Stella, N. The therapeutic potential of novel cannabinoid receptors. Pharmacol. Ther. 2009, 122, 83–96. [Google Scholar] [CrossRef] [Green Version]
- Di Carlo, G.; Izzo, A.A. Cannabinoids for gastrointestinal diseases: Potential therapeutic applications. Expert Opin. Investig. Drugs 2003, 12, 39–49. [Google Scholar] [CrossRef]
- Hornby, P.J.; Prouty, S.M. Involvement of cannabinoid receptors in gut motility and visceral perception. Br. J. Pharmacol. 2004, 141, 1335–1345. [Google Scholar] [CrossRef]
- Duncan, M.; Davison, J.S.; Sharkey, K.A. Review article: Endocannabinoids and their receptors in the enteric nervous system. Aliment. Pharmacol. Ther. 2005, 22, 667–683. [Google Scholar] [CrossRef]
- Duncan, M.; Mouihate, A.; Mackie, K.; Keenan, C.M.; Buckley, N.E.; Davison, J.S.; Patel, K.D.; Pittman, Q.J.; Sharkey, K.A. Cannabinoid CB2 receptors in the enteric nervous system modulate gastrointestinal contractility in lipopolysaccharide-treated rats. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 295, G78–G87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Storr, M.A.; Sharkey, K.A. The endocannabinoid system and gut–brain signalling. Curr. Opin. Pharmacol. 2007, 7, 575–582. [Google Scholar] [CrossRef] [PubMed]
- Wright, K.L.; Duncan, M.; Sharkey, K.A. Cannabinoid CB2 receptors in the gastrointestinal tract: A regulatory system in states of inflammation. Br. J. Pharmacol. 2008, 153, 263–270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toschi, A.; Tugnoli, B.; Rossi, B.; Piva, A.; Grilli, E. Thymol modulates the endocannabinoid system and gut chemosensing of weaning pigs. BMC Vet. Res. 2020, 16, 289. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni-Narla, A.; Brown, D.R. Localization of CB 1 -cannabinoid receptor immunoreactivity in the porcine enteric nervous system. Cell Tissue Res. 2000, 302, 73–80. [Google Scholar] [CrossRef]
- Chiocchetti, R.; Giancola, F.; Mazzoni, M.; Sorteni, C.; Romagnoli, N.; Pietra, M. Excitatory and inhibitory enteric innervation of horse lower esophageal sphincter. Histochem. Cell Biol. 2015, 143, 625–635. [Google Scholar] [CrossRef]
- Giancola, F.; Torresan, F.; Repossi, R.; Bianco, F.; Latorre, R.; Ioannou, A.; Guarino, M.; Volta, U.; Clavenzani, P.; Mazzoni, M.; et al. Downregulation of neuronal vasoactive intestinal polypeptide in Parkinson’s disease and chronic constipation. Neurogastroenterol. Motil. 2017, 29, e12995. [Google Scholar] [CrossRef] [Green Version]
- Russo, D.; Clavenzani, P.; Sorteni, C.; Minelli, L.B.; Botti, M.; Gazza, F.; Panu, R.; Ragionieri, L.; Chiocchetti, R. Neurochemical features of boar lumbosacral dorsal root ganglion neurons and characterization of sensory neurons innervating the urinary bladder trigone. J. Comp. Neurol. 2013, 521, 342–366. [Google Scholar] [CrossRef]
- Stanzani, A.; Galiazzo, G.; Giancola, F.; Tagliavia, C.; De Silva, M.; Pietra, M.; Fracassi, F.; Chiocchetti, R. Localization of cannabinoid and cannabinoid related receptors in the cat gastrointestinal tract. Histochem. Cell Biol. 2020, 153, 339–356. [Google Scholar] [CrossRef]
- Van Sickle, M.D.; Oland, L.D.; Ho, W.; Hillard, C.J.; Mackie, K.; Davison, J.S.; Sharkey, K.A. Cannabinoids inhibit emesis through CB1 receptors in the brainstem of the ferret. Gastroenterology 2001, 121, 767–774. [Google Scholar] [CrossRef]
- Coutts, A.A.; Irving, A.J.; Mackie, K.; Pertwee, R.G.; Anavi-Goffer, S. Localisation of cannabinoid CB1 receptor immunoreactivity in the guinea pig and rat myenteric plexus. J. Comp. Neurol. 2002, 448, 410–422. [Google Scholar] [CrossRef] [PubMed]
- Wright, K.L.; Rooney, N.; Feeney, M.; Tate, J.; Robertson, D.; Welham, M.; Ward, S. Differential expression of cannabinoid receptors in the human colon: Cannabinoids promote epithelial wound healing. Gastroenterology 2005, 129, 437–453. [Google Scholar] [CrossRef] [PubMed]
- Galiazzo, G.; Giancola, F.; Stanzani, A.; Fracassi, F.; Bernardini, C.; Forni, M.; Pietra, M.; Chiocchetti, R. Localization of cannabinoid receptors CB1, CB2, GPR55, and PPARα in the canine gastrointestinal tract. Histochem. Cell Biol. 2018, 150, 187–205. [Google Scholar] [CrossRef] [PubMed]
- Coutts, A.A.; Pertwee, R.G. Inhibition by cannabinoid receptor agonists of acetylcholine release from the guinea-pig myenteric plexus. Br. J. Pharmacol. 1997, 121, 1557–1566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Izzo, A.A.; Mascolo, N.; Tonini, M.; Capasso, F. Modulation of peristalsis by cannabinoid CB(1) L1 ligands in the isolated guinea-pig ileum. Br. J. Pharmacol. 2000, 129, 984–990. [Google Scholar] [CrossRef] [Green Version]
- Storr, M.A.; Bashashati, M.; Hirota, C.; Vemuri, V.K.; Keenan, C.M.; Duncan, M.; Lutz, B.; Mackie, K.; Makriyannis, A.; Macnaughton, W.K.; et al. Differential effects of CB1 N1 neutral antagonists and inverse agonists on gastrointestinal motility in mice. Neurogastroenterol. Motil. 2010, 22, 787–796.e233. [Google Scholar] [CrossRef] [Green Version]
- Donnerer, J.; Liebmann, I. Effect of CB1 ligands on neurogenic and myogenic contractile responses in the guinea-pig ileum. Pharmacology 2018, 101, 330–336. [Google Scholar] [CrossRef]
- Furness, J.B. The Enteric Nervous System; Blackwell Publishing: Malden, MA, USA, 2006; ISBN 978-1-4051-3376-0. [Google Scholar]
- Chiocchetti, R.; Bombardi, C.; Mongardi-Fantaguzzi, C.; Venturelli, E.; Russo, M.; Spadari, A.; Montoneri, C.; Romagnoli, N.; Grandis, A. Intrinsic innervation of the horse ileum. Res. Vet. Sci. 2009, 87, 177–185. [Google Scholar] [CrossRef]
- Mazzuoli-Weber, G.; Mazzoni, M.; Albanese, V.; Clavenzani, P.; Lalatta-Costerbosa, G.; Lucchi, M.L.; Furness, J.B.; Chiocchetti, R. Morphology and neurochemistry of descending and ascending myenteric plexus neurons of sheep ileum. Anat. Rec. (Hoboken) 2007, 290, 1480–1491. [Google Scholar] [CrossRef]
- Brehmer, A.; Schrödl, F.; Neuhuber, W.; Tooyama, I.; Kimura, H. Co-expression pattern of neuronal nitric oxide synthase and two variants of choline acetyltransferase in myenteric neurons of porcine ileum. J. Chem. Neuroanat. 2004, 27, 33–41. [Google Scholar] [CrossRef]
- Zwick, M.; Davis, B.M.; Woodbury, C.J.; Burkett, J.N.; Koerber, H.R.; Simpson, J.F.; Albers, K.M. Glial cell line-derived neurotrophic factor is a survival factor for isolectin B4-positive, but not vanilloid receptor 1-positive, neurons in the mouse. J. Neurosci. 2002, 22, 4057–4065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tominaga, M.; Caterina, M.J.; Malmberg, A.B.; Rosen, T.A.; Gilbert, H.; Skinner, K.; Raumann, B.E.; Basbaum, A.I.; Julius, D. The cloned capsaicin receptor integrates multiple pain-producing stimuli. Neuron 1998, 21, 531–543. [Google Scholar] [CrossRef] [Green Version]
- Poonyachoti, S.; Kulkarni-Narla, A.; Brown, D.R. Chemical coding of neurons expressing δ- and κ-opioid receptor and type I vanilloid receptor immunoreactivities in the porcine ileum. Cell Tissue Res. 2002, 307, 23–33. [Google Scholar] [CrossRef] [PubMed]
- Guo, A.; Vulchanova, L.; Wang, J.; Li, X.; Elde, R. Immunocytochemical localization of the vanilloid receptor 1 (VR1): Relationship to neuropeptides, the P2X3purinoceptor and IB4 binding sites. Eur. J. Neurosci. 1999, 11, 946–958. [Google Scholar] [CrossRef] [PubMed]
- Yiangou, Y.; Facer, P.; Dyer, N.; Chan, C.; Knowles, C.; Williams, N.; Anand, P. Vanilloid receptor 1 immunoreactivity in inflamed human bowel. Lancet 2001, 357, 1338–1339. [Google Scholar] [CrossRef]
- Schicho, R.; Florian, W.; Liebmann, I.; Holzer, P.; Lippe, I.T. Increased expression of TRPV1 receptor in dorsal root ganglia by acid insult of the rat gastric mucosa. Eur. J. Neurosci. 2004, 19, 1811–1818. [Google Scholar] [CrossRef] [PubMed]
- Kadowaki, M.; Kuramoto, H.; Takaki, M. Combined determination with functional and morphological studies of origin of nerve fibers expressing transient receptor potential vanilloid 1 in the myenteric plexus of the rat jejunum. Auton. Neurosci. 2004, 116, 11–18. [Google Scholar] [CrossRef]
- Faussone-Pellegrini, M.S.; Taddei, A.; Bizzoco, E.; Lazzeri, M.; Vannucchi, M.G.; Bechi, P. Distribution of the vanilloid (capsaicin) receptor type 1 in the human stomach. Histochem. Cell Biol. 2005, 124, 61–68. [Google Scholar] [CrossRef]
- Matsumoto, K.; Hosoya, T.; Tashima, K.; Namiki, T.; Murayama, T.; Horie, S. Distribution of transient receptor potential vanilloid 1 channel-expressing nerve fibers in mouse rectal and colonic enteric nervous system: Relationship to peptidergic and nitrergic neurons. Neuroscience 2011, 172, 518–534. [Google Scholar] [CrossRef]
- Sharrad, D.; Hibberd, T.J.; Kyloh, M.; Brookes, S.J.H.; Spencer, N.J. Quantitative immunohistochemical co-localization of TRPV1 and CGRP in varicose axons of the murine oesophagus, stomach and colorectum. Neurosci. Lett. 2015, 599, 164–171. [Google Scholar] [CrossRef]
- Chan, C.; Facer, P.; Davis, J.; Smith, G.; Egerton, J.; Bountra, C.; Williams, N.; Anand, P. Sensory fibres expressing capsaicin receptor TRPV1 in patients with rectal hypersensitivity and faecal urgency. Lancet 2003, 361, 385–391. [Google Scholar] [CrossRef]
- Anavi-Goffer, S.; McKay, N.G.; Ashford, M.L.J.; Coutts, A.A. Vanilloid receptor type 1-immunoreactivity is expressed by intrinsic afferent neurones in the guinea-pig myenteric plexus. Neurosci. Lett. 2002, 319, 53–57. [Google Scholar] [CrossRef]
- Filippova, L.V.; Fedorova, A.V.; Nozdrachev, A.D. Mechanism of activation of enteric nociceptive neurons via interaction of TLR4 and TRPV1 receptors. Dokl. Biol. Sci. 2018, 479, 44–46. [Google Scholar] [CrossRef]
- Akiba, Y.; Nakamura, M.; Ishii, H. Immunolocalization of vanilloid receptor-1 (VR-1) in CGRP-postive neurons and inter-stitial cells of cajal in the myenteric ptexus of the rat gastrointestinal tract. Gastroenterology 2001, 120. [Google Scholar] [CrossRef]
- Penuelas, A.; Tashima, K.; Tsuchiya, S.; Matsumoto, K.; Nakamura, T.; Horie, S.; Yano, S. Contractile effect of TRPA1 receptor agonists in the isolated mouse intestine. Eur. J. Pharmacol. 2007, 576, 143–150. [Google Scholar] [CrossRef]
- Schmidt, P.T.; Holst, J.J. Use of antagonists to define tachykininergic control of intestinal motility in pigs. Peptides 1997, 18, 373–379. [Google Scholar] [CrossRef]
- Buckinx, R.; Van Nassauw, L.; Avula, L.R.; Alpaerts, K.; Adriaensen, D.; Timmermans, J.-P. Transient receptor potential vanilloid Type 1 channel (TRPV1) immunolocalization in the murine enteric nervous system is affected by the targeted C-terminal epitope of the applied antibody. J. Histochem. Cytochem. 2013, 61, 421–432. [Google Scholar] [CrossRef]
- Bashashati, M.; Fichna, J.; Piscitelli, F.; Capasso, R.; Izzo, A.A.; Sibaev, A.; Timmermans, J.-P.; Cenac, N.; Vergnolle, N.; Di Marzo, V.; et al. Targeting fatty acid amide hydrolase and transient receptor potential vanilloid-1 simultaneously to modulate colonic motility and visceral sensation in the mouse: A pharmacological intervention with N-arachidonoyl-serotonin (AA-5-HT). Neurogastroenterol. Motil. 2017, 29, e13148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lai, N.Y.; Musser, M.A.; Pinho-Ribeiro, F.A.; Baral, P.; Jacobson, A.; Ma, P.; Potts, D.E.; Chen, Z.; Paik, D.; Soualhi, S.; et al. Gut-Innervating Nociceptor Neurons Regulate Peyer’s Patch Microfold Cells and SFB levels to mediate salmonella host defense. Cell 2020, 180, 33–49.e22. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, M.; Nishiyama, M.; Iizuka, S.; Suzuki, S.; Suzuki, N.; Aiso, S.; Nakahara, J. Transient receptor potential vanilloid 1-immunoreactive signals in murine enteric glial cells. World J. Gastroenterol. 2016, 22, 9752–9764. [Google Scholar] [CrossRef]
- Ives, S.J.; Park, S.Y.; Kwon, O.S.; Gifford, J.R.; Andtbacka, R.H.I.; Hyngstrom, J.R.; Richardson, R.S. TRPV1channels in human skeletal muscle feed arteries: Implications for vascular function. Exp. Physiol. 2017, 102, 1245–1258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poole, D.P.; Pelayo, J.C.; Cattaruzza, F.; Kuo, Y.; Gai, G.; Chiu, J.V.; Bron, R.; Furness, J.B.; Grady, E.F.; Bunnett, N.W. Transient receptor potential ankyrin 1 is expressed by inhibitory motoneurons of the mouse intestine. Gastroenterology 2011, 141, 565–575.e4. [Google Scholar] [CrossRef] [PubMed]
- Kaji, I.; Yasuoka, Y.; Karaki, S.-I.; Kuwahara, A. Activation of TRPA1 by luminal stimuli induces EP4-mediated anion secretion in human and rat colon. Am. J. Physiol. Gastrointest. Liver Physiol. 2012, 302, G690–G701. [Google Scholar] [CrossRef] [Green Version]
- Cho, E.; Callaghan, B.; Bron, R.; Bravo, D.M.; Furness, J.B. Identification of enteroendocrine cells that express TRPA1 channels in the mouse intestine. Cell Tissue Res. 2014, 356, 77–82. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.P.; Buber, M.T.; Yang, Q.; Cerne, R.; Cortés, R.Y.; Sprous, D.G.; Bryant, R.W. Thymol and related alkyl phenols activate the hTRPA1 channel. Br. J. Pharmacol. 2008, 153, 1739–1749. [Google Scholar] [CrossRef] [Green Version]
- Kurganov, E.; Zhou, Y.; Saito, S.; Tominaga, M. Heat and AITC activate green anole TRPA1 in a membrane-delimited manner. Pflügers Arch. 2014, 466, 1873–1884. [Google Scholar] [CrossRef]
- Nozawa, K.; Kawabata-Shoda, E.; Doihara, H.; Kojima, R.; Okada, H.; Mochizuki, S.; Sano, Y.; Inamura, K.; Matsushime, H.; Koizumi, T.; et al. TRPA1 regulates gastrointestinal motility through serotonin release from enterochromaffin cells. Proc. Natl. Acad. Sci. USA 2009, 106, 3408–3413. [Google Scholar] [CrossRef] [Green Version]
- Sandor, Z.I.; Bencsik, T.; Dekany, A.; Bartho, L. Effects of cinnamaldehyde on smooth muscle preparations. Pharmacology 2019, 104, 207–211. [Google Scholar] [CrossRef]
- Capasso, R.; Aviello, G.; Romano, B.; Borrelli, F.; De Petrocellis, L.; Di Marzo, V.; Izzo, A.A. Modulation of mouse gastrointestinal motility by allyl isothiocyanate, a constituent of cruciferous vegetables (Brassicaceae): Evidence for TRPA1-independent effects. Br. J. Pharmacol. 2012, 165, 1966–1977. [Google Scholar] [CrossRef] [Green Version]
- Doihara, H.; Nozawa, K.; Kawabata-Shoda, E.; Kojima, R.; Yokoyama, T.; Ito, H. Molecular cloning and characterization of dog TRPA1 and AITC stimulate the gastrointestinal motility through TRPA1 in conscious dogs. Eur. J. Pharmacol. 2009, 617, 124–129. [Google Scholar] [CrossRef]
- Earley, S.; Gonzales, A.L.; Crnich, R. Endothelium-dependent cerebral artery dilation mediated by TRPA1 and Ca2+-activated K+channels. Circ. Res. 2009, 104, 987–994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Earley, S. TRPA1 channels in the vasculature. Br. J. Pharmacol. 2012, 167, 13–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sullivan, M.N.; Gonzales, A.L.; Pires, P.W.; Bruhl, A.; Leo, M.D.; Li, W.; Oulidi, A.; Boop, F.A.; Feng, Y.; Jaggar, J.H.; et al. Localized TRPA1 channel Ca2+signals stimulated by reactive oxygen species promote cerebral artery dilation. Sci. Signal. 2015, 8, ra2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blackshaw, L.A.; Brierley, S.M.; Hughes, P.A.; Harrington, A.M. The hot mustard receptor’s role in gut motor function. Gastroenterology 2011, 141, 423–427. [Google Scholar] [CrossRef] [PubMed]
- Barbiers, M.; Timmermans, J.-P.; Adriansen, D.; Groodt-Lasseel, M.H.A.; Scheuermann, D.W.; Adriaensen, D. Projections of neurochemically specified neurons in the porcine colon. Histochem. Cell Biol. 1995, 103, 115–126. [Google Scholar] [CrossRef] [PubMed]
- Cornelissen, W.; De Laet, A.; Kroese, A.B.; Van Bogaert, P.-P.; Scheuermann, D.W.; Timmermans, J.-P. Excitatory synaptic inputs on myenteric Dogiel type II neurones of the pig ileum. J. Comp. Neurol. 2001, 432, 137–154. [Google Scholar] [CrossRef] [PubMed]
- Gershon, M.D. Review article: Serotonin receptors and transporters—roles in normal and abnormal gastrointestinal motility. Aliment. Pharmacol. Ther. 2004, 20, 3–14. [Google Scholar] [CrossRef]
- Russo, E.B.; Burnett, A.; Hall, B.; Parker, K.K. Agonistic properties of cannabidiol at 5-HT1a receptors. Neurochem. Res. 2005, 30, 1037–1043. [Google Scholar] [CrossRef]
- Alexander, S.P.; Christopoulos, A.; Davenport, A.P.; Kelly, E.; Mathie, A.; Peters, J.A.; Veale, E.L.; Armstrong, J.F.; Faccenda, E.; Harding, S.D.; et al. The concise guide to pharmacology 2019/20: G protein-coupled receptors. Br. J. Pharmacol. 2019, 176, S21–S141. [Google Scholar] [CrossRef] [Green Version]
- Jesus, C.H.A.; Redivo, D.D.B.; Gasparin, A.T.; Sotomaior, B.B.; De Carvalho, M.C.; Genaro, K.; Zuardi, A.W.; Hallak, J.E.C.; Crippa, J.A.; Zanoveli, J.M.; et al. Cannabidiol attenuates mechanical allodynia in streptozotocin-induced diabetic rats via serotonergic system activation through 5-HT1A receptors. Brain Res. 2019, 1715, 156–164. [Google Scholar] [CrossRef]
- De Almeida, D.L.; Devi, L.A. Diversity of molecular targets and signaling pathways for CBD. Pharmacol. Res. Perspect. 2020, 8, e00682. [Google Scholar] [CrossRef] [PubMed]
- Youn, Y.H.; Choi, E.J.; Lee, Y.H.; Oshima, T.; Miwa, H.; Park, H. The effects of 5-hydroxytryptamine1a receptor agonist, buspirone on the gastric fundus accommodation in an animal model using guinea pigs. Neurogastroenterol. Motil. 2015, 27, 532–541. [Google Scholar] [CrossRef] [PubMed]
- Delesalle, C.; Acker, N.; Claes, P.; Deprez, P.; Smet, I.; Dewulf, J.; Lefebvre, R.A. Contractile effects of 5-hydroxytryptamine (5-HT) in the equine jejunum circular muscle: Functional and immunohistochemical identification of a 5-HT1A-like receptor. Equine Vet. J. 2008, 40, 313–320. [Google Scholar] [CrossRef] [PubMed]
- Johnson, S.M.; Katayama, Y.; North, R.A. Multiple actions of 5-hydroxytryptamine on myenteric neurones of the guinea-pig ileum. J. Physiol. (Lond.) 1980, 304, 459–470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mylecharane, E. Mechanisms involved in serotonin-induced vasodilatation. Blood Vessel. 1990, 27, 116–126. [Google Scholar] [CrossRef]
- Pazos, M.R.; Mohammed, N.; Lafuente, H.; Santos, M.; Martínez-Pinilla, E.; Moreno, E.; Valdizán, E.M.; Romero, J.; Pazos, A.; Franco, R.; et al. Mechanisms of cannabidiol neuroprotection in hypoxic–ischemic newborn pigs: Role of 5HT1A and CB2 receptors. Neuropharmacology 2013, 71, 282–291. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primary Antibodies | Host | Code | Dilution | Source |
|---|---|---|---|---|
| CB1R | Rabbit | ab23703 | 1:100 | Abcam |
| CB2R | Rabbit | ab45942 | 1:200 | Abcam |
| CB2R | Mouse | sc-293188 | 1:50 | Santa Cruz |
| TRPV1 | Rabbit | ACC-030 | 1:200 | Alomone |
| TRPA1 | Rabbit | ab58844 | 1:100 | Abcam |
| 5-HT1aR | Rabbit | ab85615 | 1:100 | Abcam |
| HuC/HuD | Mouse | A21271 | 1:200 | Life Technologies |
| Secondary Antibodies | Host | Code | Dilution | Source |
|---|---|---|---|---|
| Anti-rabbit 488 | Donkey | A-21206 | 1:1000 | Thermo Fisher |
| Anti-rabbit 594 | Donkey | ab150076 | 1:1000 | Abcam |
| Anti-mouse 594 | Donkey | A-21203 | 1:500 | Thermo Fisher |
| Anti-mouse 488 | Donkey | A-21202 | 1:500 | Thermo Fisher |
| Receptors | MP Neurons | Nerve Fibers | EGCs | Tunica Muscularis | Blood Vessels |
|---|---|---|---|---|---|
| CB1R | C +/++ | + | − | + | − |
| CB2R | − | − | − | − | − |
| TRPV1 | ++/+++ | +/++ | −/+++ | − | E ++ |
| TRPA1 | C ++ | ++ | − | − | E +++ |
| 5-HT1aR | C + | − | − | + | SMC + |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Toschi, A.; Galiazzo, G.; Piva, A.; Tagliavia, C.; Mazzuoli-Weber, G.; Chiocchetti, R.; Grilli, E. Cannabinoid and Cannabinoid-Related Receptors in the Myenteric Plexus of the Porcine Ileum. Animals 2021, 11, 263. https://doi.org/10.3390/ani11020263
Toschi A, Galiazzo G, Piva A, Tagliavia C, Mazzuoli-Weber G, Chiocchetti R, Grilli E. Cannabinoid and Cannabinoid-Related Receptors in the Myenteric Plexus of the Porcine Ileum. Animals. 2021; 11(2):263. https://doi.org/10.3390/ani11020263
Chicago/Turabian StyleToschi, Andrea, Giorgia Galiazzo, Andrea Piva, Claudio Tagliavia, Gemma Mazzuoli-Weber, Roberto Chiocchetti, and Ester Grilli. 2021. "Cannabinoid and Cannabinoid-Related Receptors in the Myenteric Plexus of the Porcine Ileum" Animals 11, no. 2: 263. https://doi.org/10.3390/ani11020263