
Abstract
The non-structural protein nsp3 from SARS-CoV-2 plays an essential role in the viral replication transcription complex. Nsp3a constitutes the N-terminal domain of nsp3, comprising a ubiquitin-like folded domain and a disordered acidic chain. This region of nsp3a has been linked to interactions with the viral nucleoprotein and the structure of double membrane vesicles. Here, we report the backbone resonance assignment of both domains of nsp3a. The study is carried out in the context of the international covid19-nmr consortium, which aims to characterize SARS-CoV-2 proteins and RNAs, providing for example NMR chemical shift assignments of the different viral components. Our assignment will provide the basis for the identification of inhibitors and further functional and interaction studies of this essential protein.
Similar content being viewed by others


Biological context
Severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) represents a significant threat to human health and to the stability of contemporary societies. The development of vaccines or potential inhibitors is essential if the disease is to be eradicated. One important step in the development of effective therapeutic strategies is the characterization of potentially druggable targets that constitute the functioning virus. The international covid19-nmr research consortium is dedicated to the measurement and rapid dissemination of NMR-based parameters that will further deepen our understanding of the viral life cycle and in particular facilitate the identification of potential drug binding sites.
With over 1900 amino acids the non-structural protein nsp3 is the largest of the coronavirus proteins (Snijder et al. 2003). It comprises a large number of structured domains and disordered linker regions in SARS-CoV (Neuman 2016), and is an essential component of the replication-transcription complex (Lei et al. 2018). As such this protein represents a promising target for inhibitory strategies. Nsp3a constitutes the N-terminal section of nsp3, comprising an N-terminal ubiquitin-like domain (Ubl1) and a hypervariable acidic intrinsically disordered region (IDR) (Neuman et al. 2014).
Nsp3a has been shown to interact with the nucleoprotein (N) in the related beta-coronavirus mouse hepatitis virus (MHV) (Keane and Giedroc 2013). The acidic IDR domain has a high Glu/Asp content and is predicted to be 23 amino acids longer in SARS-CoV-2 compared to SARS-CoV. In SARS-CoV, pulldown experiments show nsp3a to associate with viral proteins nsp8, nsp9, components of the replication/transcription complex, as well as different domains of nsp3 (Imbert et al. 2008). The ubiquitin-like domain colocalizes with the nucleoprotein in viral liquid droplets in SARS-CoV-2 (Carlson et al. 2020) and has been shown to bind single stranded RNA in SARS-CoV (Serrano et al. 2007). Recently nsp3 was shown to play a structural role in the formation of molecular pores in double membrane vesicles associated with the endoplasmic reticulum (Stertz et al. 2007; Wolff et al. 2020).
Here, we present the near-complete backbone resonance assignment of the two domains of nsp3a, providing the basic data required for screening of interaction partners and detailed mapping of the interaction using the homologous structures of nsp3a from SARS-CoV (Serrano et al. 2007) and MHV (Keane and Giedroc 2013).
Methods and experiments
Construct design
The primary sequence of nsp3a (amino acids 1-206 of nsp3) from SARS-CoV-2 was extracted from NCBI genome entry NC_045512.2 [GenBank entry MN908947.3]. A commercially synthesized gene (GenScript Biotech) was codon-optimized for expression in Escherichia coli and subcloned in a pET21b( +) vector. Hexa-histidine and TEV-cleavage tags were included at the N-terminus to facilitate protein purification. After protease cleavage, the proteins contain N-terminal GAM- extensions.
Sample preparation
The nsp3a plasmid was transformed into BL21 (DE3) E. coli cells and the protein expressed heterologously with an N-terminal His6 tag. Cells were grown at 37 °C until OD600 of 0.6–0.8, at which point protein expression was induced with IPTG and incubated for 5 h at 37 °C. Bacteria were harvested by centrifugation and the cell pellet resuspended in buffer A (50 mM Tris–HCl pH 8.0 and 250 mM NaCl) with protease inhibitors (complete, Roche). Cell lysis was performed by sonication, followed by centrifugation (45 min, 18,000 rpm at 5 °C).
The protein was purified by affinity chromatography on Ni–NTA agarose (ThermoFisher), washed with buffer A supplemented with 20 mM imidazole and eluted with buffer A supplemented with 500 mM imidazole. TEV cleavage was achieved by incubation with TEV protease at 4 °C coupled with dialysis into buffer A supplemented with 2 mM DTT, and the protein concentrated and subjected to size exclusion chromatography on a HiLoad 16/600 Superdex 75 column (GE Healthcare) in NMR buffer (50 mM Na-phosphate, pH 6.5, 150 mM NaCl). For 15N and 13C isotope labelling, cells were grown in M9-minimal medium supplemented with 15N-NH4-Cl and 13C6-d-glucose (1 g/L each).
NMR experiments
A suite of BEST- and BEST-TROSY (BT) double and triple resonance assignment experiments, including BEST-HNCA, BEST HN(CO)CA, BT-HNCO, BT-HN(CO)CACB and intraresidue BT-iHNCACB (Lescop et al. 2007; Solyom et al. 2013), were recorded on 15N,13C-labeled samples (632 μM) at 298 K using a Bruker Avance III spectrometer equipped with a cryoprobe at a 1H frequency of 850 MHz. The experimental parameters for these acquisitions are summarized in Table 1. All spectra were processed using NMRFx Analyst (Norris et al. 2016) and analysed using CCPNMR Analysis Assign (Skinner et al. 2016) and NMRFAM-SPARKY (Lee et al. 2015). Manual assignment of residues was assisted using I-PINE (Lee et al. 2019).
Assignments and data deposition
The 15N,1H-HSQC of Nsp3a(1-206) is typical of a two-domain protein comprising both folded and unfolded domains (Fig. 1). The folded domain is well-resolved while the unfolded domain has a more restricted chemical shift dispersion in the 1H dimension. Nevertheless, a high percentage of resonances could be assigned in both domains (92% 1HN, 92% 15NH, 93% 13Cα, 38% 13Cβ and 83% 13C’). 13Cβ assignments are low because of difficulty in detecting transfer to these nuclei in the 3D experiments. This assignment has been deposited in the biological magnetic resonance databank (BMRB ID: 50446).

1H, 15N-HSQC spectrum of SARS-CoV-2 nsp3a (1-206) of nsp3. The spectrum was acquired on an 850 MHz spectrometer at 298 K at a concentration of 632 μM in 50 mM Na-phosphate, pH 6.5, 250 mM NaCl. Assigned backbone NH peaks are labelled
Secondary structure comparison
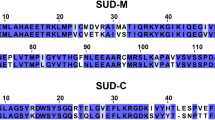
Secondary structural elements in Nsp3a from SARS-CoV-2, predicted on the basis of 1H, 15N and 13C secondary chemical shifts appear in the same regions as in the homologous protein from SARS-CoV (Serrano et al. 2007), suggesting that the three-dimensional fold is very similar (Fig. 2). Nevertheless, we note the presence of a short helical propensity around residue 110 and apparent extended or beta-sheet sampling between residues 185 and 206. Further investigation will be required to ascertain the nature of this apparent non-random coil behaviour.

Secondary structural propensity in nsp3a derived from backbone chemical shifts. Position of secondary structural elements in SARS-CoV-2 nsp3a predicted on the basis of the current assignment, using TALOS-N (Shen and Bax 2013) (top) (+ 1 equivalent to 100% helical propensity, − 1 to 100% beta-sheet propensity). Structural elements observed in the NMR structure of the nsp3a domain from SARS-CoV (Serrano et al. 2007) (bottom) are shown in ribbon representation above the plot
References
Carlson CR, Asfaha JB, Ghent CM et al (2020) Phosphorylation modulates liquid-liquid phase separation of the SARS-CoV-2 N protein. bioRxiv. https://doi.org/10.1101/2020.06.28.176248
Imbert I, Snijder EJ, Dimitrova M et al (2008) The SARS-coronavirus PLnc domain of nsp3 as a replication/transcription scaffolding protein. Virus Res 133:136–148. https://doi.org/10.1016/j.virusres.2007.11.017
Keane SC, Giedroc DP (2013) Solution structure of mouse hepatitis virus (MHV) nsp3a and determinants of the interaction with MHV nucleocapsid (N) protein. J Virol 87:3502–3515. https://doi.org/10.1128/JVI.03112-12
Lee W, Tonelli M, Markley JL (2015) NMRFAM-SPARKY: enhanced software for biomolecular NMR spectroscopy. Bioinformatics 31:1325–1327. https://doi.org/10.1093/bioinformatics/btu830
Lee W, Bahrami A, Dashti HT et al (2019) I-PINE web server: an integrative probabilistic NMR assignment system for proteins. J Biomol NMR 73:213–222. https://doi.org/10.1007/s10858-019-00255-3
Lei J, Kusov Y, Hilgenfeld R (2018) Nsp3 of coronaviruses: structures and functions of a large multi-domain protein. Antivir Res 149:58–74. https://doi.org/10.1016/j.antiviral.2017.11.001
Lescop E, Schanda P, Brutscher B (2007) A set of BEST triple-resonance experiments for time-optimized protein resonance assignment. J MagnReson 187:163–169. https://doi.org/10.1016/j.jmr.2007.04.002
Neuman BW (2016) Bioinformatics and functional analyses of coronavirus nonstructural proteins involved in the formation of replicative organelles. Antivir Res 135:97–107. https://doi.org/10.1016/j.antiviral.2016.10.005
Neuman BW, Chamberlain P, Bowden F, Joseph J (2014) Atlas of coronavirus replicase structure. Virus Res 194:49–66. https://doi.org/10.1016/j.virusres.2013.12.004
Norris M, Fetler B, Marchant J, Johnson BA (2016) NMRFx processor: a cross-platform NMR data processing program. J Biomol NMR 65:205–216. https://doi.org/10.1007/s10858-016-0049-6
Serrano P, Johnson MA, Almeida MS et al (2007) Nuclear magnetic resonance structure of the N-terminal domain of nonstructural protein 3 from the severe acute respiratory syndrome coronavirus. J Virol 81:12049–12060. https://doi.org/10.1128/JVI.00969-07
Shen Y, Bax A (2013) Protein backbone and sidechain torsion angles predicted from NMR chemical shifts using artificial neural networks. J Biomol NMR 56:227–241. https://doi.org/10.1007/s10858-013-9741-y
Skinner SP, Fogh RH, Boucher W et al (2016) CcpNmr analysis assign: a flexible platform for integrated NMR analysis. J Biomol NMR 66:111–124. https://doi.org/10.1007/s10858-016-0060-y
Snijder EJ et al (2003) Unique and conserved features of genome and proteome of SARS-coronavirus, an early split-off from the coronavirus group 2 lineage. J MolBiol 331:991–1004. https://doi.org/10.1016/s0022-2836(03)00865-9
Solyom Z, Schwarten M, Geist L et al (2013) BEST-TROSY experiments for time-efficient sequential resonance assignment of large disordered proteins. J Biomol NMR 55:311–321. https://doi.org/10.1007/s10858-013-9715-0
Stertz S, Reichelt M, Spiegel M et al (2007) The intracellular sites of early replication and budding of SARS-coronavirus. Virology 361:304–315. https://doi.org/10.1016/j.virol.2006.11.027
Wolff G, Melia CE, Snijder EJ, Bárcena M (2020) Double-membrane vesicles as platforms for viral replication. Trends Microbiol. https://doi.org/10.1016/j.tim.2020.05.009
Acknowledgements
This work used the platforms of the Grenoble Instruct-ERIC center (ISBG; UMS 3518 CNRS-CEA-UGA-EMBL) within the Grenoble Partnership for Structural Biology (PSB), supported by FRISBI (ANR-10-INBS-05-02) and GRAL, financed within the University Grenoble Alpes graduate school (Ecoles Universitaires de Recherche) CBH-EUR-GS (ANR-17-EURE-0003). IBS acknowledges integration into the Interdisciplinary Research Institute of Grenoble (IRIG CEA). Work at BMRZ is supported by the state of Hesse. Covid19 work was supported by the German research foundation in CRC902.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Salvi, N., Bessa, L.M., Guseva, S. et al. 1H, 13C and 15N backbone chemical shift assignments of SARS-CoV-2 nsp3a. Biomol NMR Assign 15, 173–176 (2021). https://doi.org/10.1007/s12104-020-10001-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12104-020-10001-8