Symmetry Transformations in Metazoan Evolution and Development

1
A.N. Severtsov Institute of Ecology and Evolution of the Russian Academy of Science, 119071 Moscow, Russia
2
A.V. Zhirmunsky National Scientific Center of Marine Biology, Far Eastern Branch, Russian Academy of Sciences, 690041 Vladivostok, Russia
3
Institute of Theory and History of Architecture and Town Planning, 105264 Moscow, Russia
*
Author to whom correspondence should be addressed.
Symmetry 2021, 13(2), 160; https://doi.org/10.3390/sym13020160
Submission received: 9 December 2020
/
Revised: 15 January 2021
/
Accepted: 18 January 2021
/
Published: 20 January 2021
(This article belongs to the Section Life Sciences)
Abstract
:In this review, we consider transformations of axial symmetry in metazoan evolution and development, the genetic basis, and phenotypic expressions of different axial body plans. In addition to the main symmetry types in metazoan body plans, such as rotation (radial symmetry), reflection (mirror and glide reflection symmetry), and translation (metamerism), many biological objects show scale (fractal) symmetry as well as some symmetry-type combinations. Some genetic mechanisms of axial pattern establishment, creating a coordinate system of a metazoan body plan, bilaterian segmentation, and left–right symmetry/asymmetry, are analysed. Data on the crucial contribution of coupled functions of the Wnt, BMP, Notch, and Hedgehog signaling pathways (all pathways are designated according to the abbreviated or full names of genes or their protein products; for details, see below) and the axial Hox-code in the formation and maintenance of metazoan body plans are necessary for an understanding of the evolutionary diversification and phenotypic expression of various types of axial symmetry. The lost body plans of some extinct Ediacaran and early Cambrian metazoans are also considered in comparison with axial body plans and posterior growth in living animals.
1. Introduction
The symmetry of an animal body is one of the basic features characterizing the body plan (Bauplan) in different metazoan clades [1,2,3,4,5,6,7,8,9,10,11,12,13]. Symmetry of biological structures can be defined as the repetition of parts in different positions and orientations to each other [14]. In addition to the rotation (radial symmetry), translation (metamerism), and reflection (bilateral symmetry) including mirror and glide reflection, many biological objects show scale (fractal) symmetry [3,4,14,15,16,17,18,19,20,21,22]. M. Manuel [8] considered asymmetrical, spherical, cylindrical, n-radial, and bilateral symmetry as main types of symmetry in metazoan body plans; the author reasoned that the particular cases of rotational and reflection symmetries are sufficient for biologists, although a more general theory of organism geometry would require other kinds of symmetries, for example, translational and helicoidal symmetry, to be investigated. G. Holló [10] assumed only a few types of overall metazoan body symmetry: radial, biradial, and bilateral symmetry. Such complex symmetry types as combinations of radial and bilateral symmetry in some cnidarians, bilateral and pentaradial symmetry in echinoderms [8,23], and bilateral and spiral (helical) symmetry [11,14,19] were also considered. Metazoan body plans combine well-defined primary, secondary, and, in many bilaterians, tertiary body axes [13].
Body axes form the basis of animal body plans; polarity establishment requires a symmetry-breaking event, so body polarity is a breaking of symmetry [8,24,25]. Symmetry breaking inevitably arising during early development plays a key role in biological morphogenesis. Symmetry breaking is one of the fundamental processes of development [25,26]. Symmetry breaking and “symmetry propagation” [27] are inevitable in evolution and development [3,4,20,25,27,28]. Biological morphogenesis has a dynamical character involving qualitative discontinuities as topological bifurcations tightly coupled with symmetry breaking, so developmental and evolutionary transformations of symmetry are discrete steps in biological morphogenesis inevitably disrupting a preexisting pattern of symmetry [17,18,19,20,29,30]. Both phylogeny and ontogeny include transitions from symmetry to dissymmetry, and the process is generally shifted towards symmetries decreasing [8,20,31]. In all metazoan taxa, random deviations from perfect symmetry in the form of fluctuating asymmetry are common and inevitable (see [14,19] and Section 6.1).
Developmental gene transcriptional regulatory cascades and the signaling pathway responsible for the patterning of the metazoan body plan and underlying the establishment of body polarity are remarkably similar throughout Metazoa; there are a limited number of major signaling pathways, primarily Wnt, BMP, Notch, and Hedgehog cascades, which determine metazoan body axes [8,9,10,12,32,33,34,35]. In Metazoa, the main body axes are patterning by the same pathways and gradients [9,12,36,37,38].
Molecular components of the signaling systems as diffusible morphogens carry positional information, which determines axial coordinates in morphogenetic fields of an egg, embryo, and future adult animal through instructive clues for genes to be expressed differently in cells located in different spatial areas [8,9,17,18,33,39,40,41,42]. The signaling pathways are just the major highways of information flow [33]. It is possible now to establish the relation between morphogenetic fields with positional information provided by gradients of different morphogens (transcription and translation factors, components of signaling systems including Wnt, BMP, Nodal, and Hedgehog) and integral developmental patterns to understand many genetic and cellular mechanisms of symmetry breakdown in metazoan development and evolution [9,13,41,42].
Understanding how diverse body plans evolved remains one of the most exciting and challenging goals for evolutionary and developmental biologists [43]. In this review, we try to provide a better understanding of correlations between genetic basis and some phenotypic expression of axial body plans in different metazoans, taking into consideration recent advances in this research field.
2. Phylogenomic Data: The Genetic Background of Axial Patterning
Modern phylogenomics makes it possible to associate the genomic diversity with phenotypic biodiversity in Metazoa by revealing the genomic correlates of different morphological complexity (see [36,44,45,46]). Sequencing the genomes of many animals at different levels of complexity indicates that the genomic “toolkit” that controls the development of animals and the formation of a bilaterian body plan originated in the early evolution of Metazoa [36,44,45,46,47,48]. A significant, clearly pronounced correlation between the regulatory complexity of the genome (determined by the total number of transcription factors and other transcription regulators) and the morphological complexity of the organism (measured by the number of cell types) in Metazoa as a causal evolutionary relationship between the regulatory and phenotypic complexity was found [36,49]. On the other hand, it was shown that many extant metazoan taxa have undergone genomic and morphophysiological simplification in the course of their evolution; it has become apparent that regression is a common evolutionary trend that cannot be ignored (see [4,46,47]). Progressive evolution is characterized by the addition of traits and stages of development, whereas regressive evolution is characterized by the reduction of ancestral structures and stages of development [4]. We can say that evolutionary progress can be characterized by an increase in the amount of genomic information and phenotypic complexity, and regressive evolution can be characterized by the reduction of ancestral genomic information and secondary simplification of the phenotype [46]. The data of phylogenomics and evolutionary developmental biology indicate that some representatives of Cnidaria contain a number of transcription factors and signaling pathway genes, which are characteristic of Bilateria and, probably, of a common ancestor of Protostomia and Deuterostomia [36,47,48,50]. It was proposed that modern Cnidaria, as well as Placozoa, represent “depauperated” remains of the early Ediacaran fauna [47]. According to Srivastava [36], the four non-bilaterian animal lineages (i.e., cnidarians, ctenophores, placozoans, and sponges) diverged from bilaterians early in animal evolution (i.e., they are early-branching relative to lineages within the bilaterian clade); these lineages have been considered by some to be morphologically “simple”, because they lack many bilaterian features.
Comparative genomic data provide evidence that all families of the signaling pathways and most of the molecular developmental toolkit that is used to build bilaterian body plans were already established in the common metazoan ancestor [8,12,36,37]. The ancestral metazoan complexity is deduced from the high degree of conservation of the gene family complements in contemporary animals [35]. This complexity is reflected by the presence of almost full gene family complement, such as the major toolkit genes Wnt, and in modern representatives of the earliest branching lineages, such as sponges, Trichoplax, and cnidarians [13,51,52,53,54]. These data supported the proposal that ancient metazoans in the pre-Cambrian were already genetically complex, with most of the modern gene families and genomic organization already present [35]. The canonical Wnt pathway presents in all extant metazoans; for example, in cnidarians, it specifies the oral–aboral embryonic axis, and thus, the eumetazoan sister clades of cnidarians and bilaterians employ homologous “developmental toolkits” for body-axis specification [13,51,52,53,54]. The formation of a localized Wnt signaling center at the site of gastrulation determines the orientation of the primary body axis in both bilaterians and non-bilaterians, in deuterostomes and protostomes, indicating that the conserved Wnt signaling pathway is at the base of polarized development in all metazoans [5,34,37,52,54,55]. Thus, the Wnt pathway arose concurrently with the first metazoans [51]. A core set of Wnt genes was already present in sponges, but cnidarians are the only pre-bilaterian animals with a complete set of Wnt genes and a complete functional repertoire of Wnt ligands; in Protostomia and Deuterostomia, Wnt signaling is essential for posterior patterning [5,34,54]. Wnt patterning across both the oral–aboral and anterior–posterior axes may suggest that the primary axis across Eumetazoa is homologous, and similar Wnt patterning across the primary body axis of sponges suggests that the primary body axis across all Metazoa may be homologous [37]. Data revealed by comparative genomics have shown that members of the Wnt signaling pathway are specific for metazoans as important regulators of embryogenesis and cell differentiation [12,54]. In both bilaterians and cnidarians, the Wnt signaling pathway determines not only patterning of the primary axis but also gastrulation movements, germ-layer specification, and axial differences in cell fate [51,52,55]. In segmented bilaterian animals, the Wnt signaling pathway is a key component of posterior segment addition from the posterior growth zone (see [54,56,57,58] and Section 4.4). The proposed ancestral role of Wnt in early Metazoa was to establish the posterior pole of the primary body axis, thus breaking symmetry, positioning the site of gastrulation, and organizing the primary axis, without which the body could not form; the pole of the primary body axis specified by high Wnt signals becomes the blastopore [37,51,54].
In Bilateria, dorsal–ventral axial patterning has similar molecular underpinnings as in all Metazoa; BMP signaling is a conserved mechanism for dorsal–ventral patterning across diverse bilaterians [36,37,38,56,59]. Genetic analyses have shown that the genes responsible for bilateral symmetry most likely appeared prior to the cnidarian–bilaterian split, in a common ancestor of Cnidaria and Bilateria; both molecular and anatomical evidence support a secondary (“directive”) axis in at least some cnidarians [9,10,13,51,52,60,61]. In addition, there are indications that also ctenophores, placozoans, and sponges diverge from urbilaterian ancestors in early evolution [36].
Hox gene transcription factors are important regulators of positional identity along the main axis, not only in Bilateria [62,63,64], but also in Cnidaria (the sister group to the Bilateria), which possess genes related to both anterior and central/posterior Hox genes [52,61]. Generally, both Wnt (Section 3) and Hox cascades (Section 4.1) pattern the primary body axis in Bilateria, but Wnt proteins operate as primordial symmetry-breaking signals [34,52,55]. Hox gene evidence leaves little doubt that the anterior–posterior axis of all bilaterians is homologous [65]. Phylogenetic reconstructions of the bilaterian Hox gene set indicate that the common ancestor of most extant animals had a Hox cluster containing anterior, central, and posterior Hox genes [36,48]. Cnidarians probably retain components of a simple ancestral Hox cluster consisting of at least two types of Hox genes, related to anterior and central/posterior genes functioned to organize the primary axis in the cnidarian ancestor (see [61]). Before the emergence of bilaterian animals, three distinct clusters of ANTP class homeobox genes arose: Hox, ParaHox, and NK homeobox genes; however, Hox genes are just one type of homeobox gene, they are “the tip of the homeobox iceberg” [66] (p. 3).
The left–right differences in bilaterally symmetrical animals are genetically determined by the conserved Nodal signaling system [26,33,59,67,68,69,70,71]. Although many factors were shown to regulate the left–right asymmetry, including members of Wnt, Notch, FGF, or BMP signaling pathways, the central pathway that is directly linked to left–right axis specification is that involving Nodal and regulating body asymmetry in a plethora of animals [71]. The control of axial asymmetry along the main body axes by the Nodal pathway might be ancestral in eumetazoans; the Nodal signaling probably dates back to the common ancestor of bilaterians and cnidarians and thus predates the origin of bilaterality [71]; it is also possible that the gene Nodal appeared after the cnidarian–bilaterian split in the common ancestor of protostomes and deuterostomes [59,68,70].
There is evidence of crosstalk between gene regulatory networks of different signaling pathways and other developmental regulatory cascades (see [12,33,42,68]. The evolution in Bilateria is determined not only by signaling pathways and the Hox gene system, but also by the entire genome—an integrated, hierarchical network system including numerous intragenomic connections [35,72,73]. In addition, the morphological and developmental evolution in bilaterians is due not just to coding DNA; studies of the “dark matter” [35] (p. 5) of the animal genome, DNA that does not encode proteins, reveals its regulatory role. A significant part of this DNA encodes various RNAs, including long and short noncoding RNAs, transposons, and microRNAs. The key regulatory functions of some of these RNAs were established [12,35,36,73,74]. Non-coding RNAs, including microRNAs and long non-coding RNAs, and all complex gene networks are very important in development and evolution [36,72,73,74].
3. Primary Body Axis: The Wnt Signaling Pathway
Metazoan body plans are organized by a primary body axis (referred to “anterior/posterior” “apical/basal”, and “oral/aboral” in different taxa) [1,5]. The generation of the primary embryonic axes is a crucial first step in metazoan development [7]. The first symmetry breaking takes place along the animal–vegetal (anterior–posterior) axis of an egg or early embryo, and the primary axis is established in most Metazoa before gastrulation. Establishment of the animal–vegetal (oral–aboral) axis in Metazoa is guided by the Wnt signaling pathway, and the Wnt signaling pathway may be the ancestral metazoan axial patterning system predating the “Hox code” and other developmental patterning signals, and thus appears to constitute the original ancestral program for primary body axis specification, presumably homologous and similar across Eumetazoa [5,8,9,10,13,32,34,37,51,52,53,54,55]. Data revealed by comparative genomics have shown that members of the Wnt signaling pathway are specific for metazoans as important regulators of embryogenesis and cell differentiation [13,54,57].
Wnt signalling is crucial for animal development, as it is involved in the regulation of numerous developmental processes such as pattern formation, cell proliferation and cell migration, posterior elongation, and organogenesis (see Section 4.4). Wnt genes are subdivided into 13 classes, of which 12 are present in protostomian animals; the name Wnt comes from fusing wingless (the name of the Drosophila segment polarity gene) with integrated (the name of one of its vertebrate homologues) [12,33,54]. The Wnt pathway has been shown to be very dynamic, exhibiting both loss and gain patterns in various lineages (see [35]). The available evidence indicates that Wnt proteins are asymmetrically distributed along the future primary body axis; in the early embryo, posterior Wnt signaling is often a maternally imposed developmental directive [51]. The body pole specified by high Wnt signals establishes the posterior pole of the primary body axis determining its orientation in all metazoans and positioning the site of gastrulation (becoming the blastopore) [37,51,52,55].
In the canonical Wnt pathway, binding of Wnt(s) to their receptors induces intracellular gene cascades that lead to the release of β-catenin, which then regulates the transcription of Wnt target genes. β-catenin is the key intracellular effector of the canonical Wnt signalling pathway and plays an important role in the early polarization of the animal–vegetal axis of the embryo [12,34,54,55]. An ancient component of the Wnt-imposed posterior program is the transcription factor Brachyury, which is present in virtually all extant metazoa just like Wnt itself. In diverse animals ranging from Hydra to mice, Wnt directly activates Brachyury expression at the posterior pole of the primary body axis [51]. In Deuterostomia, the cascade of Wnt-dependent pathways is required for the specification of the endomesoderm and regulates subsequent Nodal and BMP signaling [8,54,59].
In planarians with their extraordinary morphological plasticity, the Wnt/β-catenin pathway provides positional information pivotal for proper head regeneration [75]. Iglesias et al. [75] studied the role of the β-catenin homologues from the planarian Schmidtea mediterranea (Smed β-catenin1). Like many freshwater planarians, S. mediterranea can regenerate a head or tail, and experimental amputation of either produces trunk fragments usually regenerating both anterior and posterior structures; when β-catenin is depleted by RNA interference, a head is generated instead of a tail, resulting in two-headed or multi-headed planarians [13,51,55,75]. Thus, silencing of Smed β-catenin1 in regenerating and intact planarians induces a gradual anteriorization of the animals that finally leads to a radial-like hypercephalized phenotype, demonstrating the requirement for Smed β-catenin1 in anterior–posterior axis reestablishment and maintenance [75].
Unique examples of striking body symmetry transformation were observed in the most extreme cases, when planarians not only lose their tail, but ectopic heads replete with eyespots begin sprouting around the periphery of the body, eventually forming almost a radially transformed organism whose perimeter essentially consists of multiple heads [9,13,51,75] (Figure 1).
The radial-like hypercephalized planarian phenotype demonstrates the requirement for Smed β-catenin1 expression in anterior–posterior axis re-establishment and maintenance [75]. In normal planarians, the central nervous system is composed of two anterior cephalic ganglia located at one end of two ventral nerve cords, while in radial-like hypercephalized planarians, the cephalic ganglia and ventral nerve cords are transformed into a thick ring from which cephalic branches ectopically differentiate all around the body, and all photoreceptor cells appeared to be connected through their visual axonal projections to the circular brain [75]. Moreover, global abrogation of Wnt signaling (by β-catenin RNAi) not only in regenerating but also in steady-state planarians leads to the relinquishment of posterior identities and can generate ectopic heads, while hyperactivation of Wnt signaling in regenerating planarians leads to the loss of anterior fates and can generate “two-tailed” animals in which the head has been converted into a tail (see [51,75]). Thus, in the asexual freshwater planaria, specific experimental manipulations can symmetrize the anterior–posterior axis or as the limiting case eliminate this axis to produce a radially symmetric, anteriorized form around the remaining dorsal–ventral axis [13]. Holló and Novák [60] suggested that radial and bilateral body plans could have been generated with the same or similar genetic toolkit but with different regulatory networks.
4. Symmetry in Bilateria
The majority of Metazoa are bilaterally symmetric and fall in a group called the Bilateria, which are characterized as having a primary body axis (head to tail), a dorsal–ventral axis (top to bottom), and a left–right axis; Bilateria possess a central nervous system with a brain and sense organs concentrated at the anterior end of the body (cephalization), three germ layers, true muscle, and a through gut [25,36,66,76]. Bilaterian animals are patterned along two major body axes perpendicular to each other: the primary anterior–posterior axis and the secondary dorsal–ventral axis [9,33]. Bilateral symmetry dominates the animal world with more than 99% of species showing this symmetry type [9,10,60].
Bilateral symmetry with two body axes arose early in animal evolution, probably in slow, flat, worm-like organisms moving on a substrate [1,10,60]. The Cambrian is marked by the evolution of active, directed, exploratory locomotion in a three-dimensional world [36,66]. In early Bilateria, their “ecological engineering” [66] (p. 7) included also an epithelial through gut with two openings, ingesting food through a mouth and passing food unidirectionally before ejection of waste through an anus [36,66]. In evolutionary terms, bilateral symmetry is highly adaptive [25]. Locomotion in three-dimensional macro-world space is itself sufficient to explain the maintenance of bilateral symmetry in animal evolution, and bilaterality is the only type of symmetry that can maximize this force; thus, an actively moving bilateral body can have the maximal maneuverability as compared to other symmetry types, providing obvious selective advantages in the bilateral animal [25,60]. The combination of bilaterality with the centralization of the nervous system and cephalization allowed the evolution of successful body plans, ensuring precise locomotion and rapid information processing [36,60,66]. Symmetry is more readily abandoned in the internal organs than in other parts of the body, including the brain and limbs; asymmetry in the internal organs of the body is more adaptive, as it provides efficient packaging [25]. In humans, the tradeoff between symmetry and asymmetry results in individual differences in brain asymmetries and handedness [25].
4.1. Axial Hox Code
It is well known that the genes of the Hox cluster encoding transcription factors of the ANTP class play a central role in the determination of the anterior–posterior axis and the entire complex of the main features of the bilaterian body plan. Hox gene expression is initiated in early embryogenesis, and Wnt signaling usually precedes zygotic Hox activity; Hox genes are not involved in the early establishment of the anterior–posterior axis, a process in which various maternally localized determinants and signaling molecules participate, and they act in controlling the correct positioning of structures along an already established axis [5,8,44]. Wnt signaling might induce and regulate Hox gene expression in anterior–posterior axis formation, and the functional coupling of these two mechanisms in primary axis formation might also represent an ancient feature [5,9].
It is known that genetic mechanisms determining the anterior–posterior body pattern in Bilateria are associated with the physical arrangement of Hox genes in their cluster on the chromosome: genes located closer to the 3′ end of the cluster are expressed closer to the anterior end of the embryo and earlier than their 5′-associated neighbors, thus showing spatial and temporal colinearity. The colinearity of Hox gene expression, i.e., the correspondence between the position of the gene in the cluster and the place and time of its expression in the developing organism, forms the basis for creating a vector gradient of positional information in the process of individual development [36,66,72,77,78,79]. The colinearity of expression and conservatism of the Hox system in the determination of anterior–posterior polarity and segmentation (“an axial Hox code”: [61], p.1) is a very important innovation of Bilateria, a plesiomorphism of all bilateral animals, the common ancestor of which already possessed a sufficient set of Hox genes in its cluster (see [36,66]).
Hox genes, which are required for patterning of the anterior–posterior axis in Bilateria, were found to be expressed in tandem domains along this axis, and to be encoded in tandem (forming the Hox complex) in the genomes [36]. This ability of the Hox cluster to coordinate the functioning of the entire complex of Hox genes and the multitude of their target genes with the performance of the integral function of coding the axial pattern of the organism enabled D. Duboule to consider the Hox cluster as a metagene [78,80]. The phenomenon of the colinearity of the cluster Hox genes is the only unambiguous example of topological mapping as a translation of the linear vector of spatial order of the gene localization in the Hox cluster (axial symmetry) into the spatial–temporal order of their expression and axial morphology of the organism [3,18,45]. Thus, the Hox cluster in Bilateria is the metagene encoding an axial body pattern.
Although the emergence of Hox genes precedes radiation from Bilateria, the Hox system, as a complex of coordinated Hox genes, is specific for Bilateria. The emergence and evolutionary expansion of Hox genes, and their clusters, as well as the related genes of the ParaHox and NK clusters, accompanied by the loss of some genes, provided the genetic basis for the evolutionary transformations and diversifications of body plans in Bilateria [35,36,66,72,79]. There is a correlation between an increase in the number of Hox genes with an increase in the complexity of the body plan (see [36,66]). It is proposed that diversification of ANTP class genes played an important role in the Cambrian explosion by contributing to the patterning systems used to build animal bodies capable of high-energy directed locomotion and increasing body complexity; “without this molecular evolution, there could be no Cambrian explosion” [66] (p. 7). The patterning Hox, ParaHox, and NK homeobox genes were supposedly recruited for roles in patterning the ectoderm, gut, and mesoderm, and this permitted the evolution of animal body plans capable of active, directed locomotion and, due to evolution of a through-gut with a distinct anus, active burrowing and feeding in sediments [66].
An ancient Hox code may have evolved to regulate both tissue segmentation and body patterning in the bilaterian–cnidarian common ancestor [61]. D. Duboule succeeded in associating the organization of the Hox cluster with morphological transformations through the heterochrony of early embryogenesis with the body plan of an adult organism by controlling the growth rate in early embryogenesis [77,78,80,81]. It is known that Hox genes are involved in the functioning of an extensive genetic cascade with tens or hundreds of target genes that regulate a wide range of key developmental processes, including the passage of the cell cycle and cell proliferation (see [45,82]).
Comparison of the organization of Hox clusters and the pattern of Hox gene expression with the morphology of the organism makes it possible to trace the emergence of macroevolutionary innovations as changes in the morphogenesis and body plan of Bilateria depending on the pattern of the Hox code. A correlation knot is considered that includes a particular organization of Hox genes, a type of early embryogenesis, and cellular developmental resources, and thus determines alternative evolutionary trajectories of various taxa of Bilateria [45,46]. Radical, macroevolutionary transformations of the body plan and axial body pattern are associated with the expansion or loss of individual Hox genes of the ancestral cluster and with the topological reorganization of the Hox cluster, suggesting a leading role of the Hox code in determining the axial body plan of bilateral animals [63,64,66]. A certain reduction and simplification of genome and phenotype quite often occurred in the evolution of multicellular animals (see [46,66]). Examples of a striking restructuring of the body plan, coupled with the loss of some Hox genes, were found among Dicyemida and Orthonectida, formerly “Mezozoa” and now Lophotrochozoa (see [83,84]). Apparently, ancient dorsal–ventral axis is lost during regressive evolution in Dicyemida and Orthonectida. Among Ecdysozoa, dramatic regressive evolutionary transformations occurred in crustacean Rhizocephala (see [85]). In Rhizocephala and all studied cirripedian crustaceans, the absence of the AbdA gene correlates with the loss of the abdominal part of the body (see [45,85]). In some Rhizocephala (Crustacea: Cirripedia), the blastogenesis and coloniality at the parasitic, sexually mature stage involved a complete loss of the body plan and morphology of arthropods, including main body axes common in Panarthropoda (see [86,87] and Section 6). Moreover, it is reasonable to suppose that non-bilaterian animal lineages (i.e., morphologically “simple” cnidarians, ctenophores, placozoans, and sponges) diverged from bilaterians early in animal evolution and they are early branching relatives to lineages within the bilaterian clade [36], which lost some ancient body axes.
4.2. Dorsal–Ventral Axis: The BMP Signaling Pathway
In Bilateria, the BMP family (bone morphogenetic proteins, which were originally discovered by their ability to induce bone formation) is exploited to create the dorsal–ventral axis—the secondary axis that runs perpendicularly to the primary body axis established by Wnt; the BMP pathway also regulates important interactions in development including cell division, apoptosis, cell migration, and differentiation [6,8,9,13,33,34,51,52]. The third, left–right asymmetry appears in some arthropods and is ubiquitous in vertebrates (see [13] and Section 4.5 and Section 5).
Wnt and BMP signaling pathways specify main body axes creating a near-Cartesian coordinate system of positional information, which was set up by gradients of perpendicular Wnt and BMP signaling gradients representing a conserved ancestral panbilaterian program [7,9].
In Bilateria, the patterning mechanisms of the dorsal–ventral polarity involve a conserved protein network that comprises, among others, secreted molecules of the BMP family (e.g., encoded by the gene decapentaplegic in Drosophila and its orthologue BMP4 in vertebrates) and their antagonist ligands (e.g., encoded by the gene short gastrulation in Drosophila and its orthologue chordin in vertebrates) [6,8,9,10,34]. Thus, the dorsal–ventral axis is patterned by the conserved BMP-Chordin network in Bilateria [6,34].
The primary and secondary axes are established in most bilaterians during oogenesis or in early embryogenesis, before gastrulation [9,22,33,39]. During the development of different bilaterians, the dorsal–ventral axis can be specified before fertilization, during fertilization, or during early egg cleavage (see [3,33]). In chordates, the dorsal–ventral axis is usually established after the sperm entry; this symmetry breaking often involves an asymmetric flow of the cortical actin network and ooplasmic rearrangement specifying the dorsal–ventral polarity of a future animal [2,6,26,28,39,88]. Thus, symmetry transformations of the egg yield the most important information that determines the axial pattern of the future body.
BMP signaling is a conserved mechanism for dorsal–ventral patterning across diverse bilaterians; however, in chordates, BMP activity is high on the ventral side, where the mouth is located, while in other bilaterians, BMP signaling is active on the dorsal side, i.e., organization along the dorsal–ventral axis is inverted between arthropods and vertebrates (see [37,38]). A defining feature of chordates is the unique presence of a dorsal hollow neural tube that forms by internalization of the ectodermal neural plate specified via inhibition of BMP signaling during gastrulation by antagonists, such as Chordin; thus, the chordate central nervous system exists in a dorsoventrally inverse orientation relative to other bilaterally symmetric animals [38]. Using hemichordate and sea urchin larvae, Su et al. [38] found that BMP signaling controls dorsal–ventral and neural patterning. Alterations of BMP levels result in pronounced morphological changes reminiscent of a proposed intermediate stage in the emergence of the chordate body plan. Transient over-activation of BMP signaling during gastrulation blocked mouth formation, resulting in loss of the larval mouth and centralization of the neurogenic ectoderm on the ancestral ventral side devoid of BMP activity, and thus displayed an organization similar to that of the chordate neural plate [38]. These morphological changes are similar to a proposed intermediate stage leading to the emergence of chordates with a dorsally located central nervous system in an inverse dorsal–ventral body axis relative to their ancestors [38]. Su et al. [38] proposed that evolutionary “tinkering” [89] (p. 1161) with BMP signaling level may have driven dramatic morphological changes, leading to the emergence of chordates. Such changes in BMP levels during gastrulation resulted in loss of the deuterostome ancestral mouth, and the use of one of the dorsal openings (perhaps a pharyngeal gill slit) as a new mouth would have allowed the chordate ancestor to take on an inverse dorsal–ventral orientation relative to Ambulacraria and the deuterostome ancestor [38].
4.3. Metamerism in Bilateria: Wnt and Notch Signaling Pathways
Translational symmetry in biology is referred to as metamerism, or segmentation [1,3,8,14]. Most metazoans are segmented along their anterior–posterior axis, and many evolutionary rearrangements in the body plan are associated with the multiplication of body parts, polymerization, which creates a redundancy of elements and subsequent oligomerization, forming the basis for the progressive evolution of many metazoans [1,14]. Metamerism is always based on the repetition of body parts, serially homologous to each other, arranged along the longitudinal axis or the plane of the symmetry [14,90]; segments are usually regarded as body modules [3]. As a basic feature of the body plan, metamerism dominates among the recent metazoans, thus proving to be an extremely valuable evolutionary invention [14,42,90].
The bilateral symmetry of most animals is associated with the repetition of parts as pairs, located on the left and right sides as mirror images of each other; for each pair of parts, a reflection about the median plane maps the left part onto the right part and vice versa, so this reflection about the median plane is fundamental for bilateral symmetry and can be used to characterize it [14]. Modern Bilateria possess external reflection (mirror) symmetry, which inevitably includes some degree of fluctuating asymmetry as well as genetically determined asymmetry of the internal organs of the left and right sides in many bilaterians. Mirror symmetry can be defined by configurations in which one half represents the mirror image of the other; left and right halves are separated by a symmetry axis. In Bilateria, the two sides of the body are to a high degree left–right mirror images, where limbs and sense organs are arranged in mirrored pairs on opposite sides of the body [14,25,91]. Bilaterial symmetry is an innovation often thought to be associated with the regionalization of sensory structures and the eventual elaboration of a central nervous system [48].
In vertebrate embryos, the segmented anatomy is evident in the two bilaterally symmetrical rows of somites—Paired epithelial spheres budding off from an undifferentiated paraxial mesoderm [3,24,66,68,92,93,94]. The paraxial mesoderm is progressively segmenting in an anterior to posterior direction, forming bilaterally symmetrical somites that will give rise to dermis, an axial skeleton, and skeletal muscles [3,68,92]. Signaling gradients in the presomitic mesoderm are believed to supply positional information that regulates segment length [42,94]. In such segmented animals as vertebrates, annelids, and panarthropods, Wnt signaling is a key component of posterior segment addition (Section 4.4), and one of the key functions of the Wnt signaling pathway is its general involvement in posterior growth in animals [54]. In annelids and arthropods, Wnt signaling is likely involved in segment border formation and regionalisation of the segments; the most known Wnt gene, wingless (wg/Wnt1), is a key factor maintaining parasegmental boundaries in concert with other segment polarity genes in Drosophila [54,56]. While the specific gene regulatory networks controlling posterior growth (Section 4.4) and segmentation can differ between different groups of animals, and in some cases even between closely related species of the same group, Wnt signalling seems to always be involved [54].
In vertebrates, somitogenesis is a regularly repeated, rhythmic morphogenetic process, in which each bilateral somitic pair is consequently segregated from the undifferentiated presomitic mesoderm at regular time intervals controlled by a complex genetic network functioning as a “molecular clock” of segmentation with oscillatory expression of a gene set of the Wnt, Notch, and FGF (fibroblas growth factors) signaling pathways [7,42,54,65,68,92,93,94,95]. Oscillatory activity of the homeobox genes Hes1 and Hes7, which are downstream targets of Notch signaling, is translated into periodic changes in the fate of cells, which are grouped in cohorts subsequently forming somites [54,93]. There is clear evidence of crosstalk between the FGF, Wnt, and Notch signaling pathways in the control of the oscillations [92]; in addition, the role of retinoic acid seems to synchronize the oscillation of clock gene expression between the left and right sides of a vertebrate embryo [42,68]. Interactions between these signaling pathways permit the synchronized activation of segmentation genes in the presomitic mesoderm in response to the “segmentation clock”, which is defined by periodic waves of expression of genes of the FGF, Wnt, and Notch pathways that are travelling along the presomitic mesoderm [42,68,92,94]. Reiterative translation symmetry of segmentation controlled by the “segmentation clock” based on a mechanism characterized by oscillating gene expression was also found in insects [56,93,96,97]. In Drosophila and other hexapods, activation and repression of Hox genes by gap genes, which are part of the segmentation cascade, set up regionalized expression of the Hox genes [98]. It is possible that pair rule gene orthologs are part of an ancestral segmentation clock mechanism [56]. Spatial-temporal colinearity as an ordered expression of genes in the Hox cluster and a multitude of target genes probably ensured the synchronization of the appearance of paired rudiments of segments on the left and right sides of the body and, as a consequence, the appearance of mirror symmetry in Bilatera and a more perfect coordination of functions of the nervous, muscle, and all other systems of a directionally moving bilaterian organism. Moreover, Hox genes themselves appear to oscillate along with the segmentation clock during this process [24,99].
Thus, molecular oscillators represent a common feature of segmentation in divergent animals [42,75,91,93,97]; a clock-based mechanism underlies segmentation in animals as widely separated as arthropods and vertebrates. Moreover, a similar “clock” may be a general principle that governs patterning in growing tissues, but the discussion about the mono- vs. polyphyletic origin of segmentation in the arthropod, annelid, and vertebrate lineages is far from settled [4,75,97,100]. It is probably that segmentation arose independently in different animal clades [12].
In chordates and other deuterostomes, in addition to somitomery, another type of segmentation is manifested, associated with the subdivision of the pharyngeal endoderm and the formation of a series of gill slits, i.e., chordates have an additional variant of translational symmetry: branchiomery [3,4]. Recently, a conserved deuterostome-specific genomic microsyntenic group, the so-called pharyngeal gene cluster including four transcription factor genes (nk2.1, nk2.2, pax1/9, and foxA1/2) that are expressed during development of the pharyngeal gill slits and the branchial apparatus, was described as the most prominent morphological innovation of the deuterostome ancestor (see [12,35]). Molecular and paleontological evidence supports the idea that pharyngeal gill slits were present in the deuterostome ancestor (see [35,38]). Thus, evolution of Deuterostomia included a greater number of symmetry transformations in comparison with Ecdysozoa and Lophotrochozoa.
4.4. Posterior Growth Mechanism in Bilateria
In bilaterians as well as cnidarians, the “Wnt code” determines not only patterning the primary, anterior–posterior (oral–aboral) axis during early development and axial differences in cell fate, but also gastrulation movements and germ-layer specification [8,51,52,55]. Three major metazoan clades, vertebrates, arthropods, and lophotrochozoans, have common aspects of molecules and morphology in the segmentation process, developing their segments through a process of sequential budding from the posterior growth zone, so the sequential patterning of segments depends critically on posterior elongation of the growing embryo [57,65,91]. Axial extension from a posterior growth zone is a common feature and the basal mode of body axis elongation in Bilateria; presumably, the terminal addition was the ancestral growth mode [54,57,58,65,101]. Wnt genes are involved in posterior elongation in both vertebrates and invertebrates, which provides evidence suggesting that the genetic network controlling posterior growth was already present in the last common ancestor of the Bilateria [54,57]. In addition, Hox gene evidence leaves little doubt that the anterior–posterior axis of all bilaterians is homologous [65]. By coupling cell fate to position, symmetry-breaking Wnt signaling activities play crucial roles in development, controlling cell divisions, cell polarity, convergent cell extension, directed cell migration, and therefore coordinating lineage specification with spatial morphogenesis [51,54,57,58].
A pivotal evolutionary innovation was the extension of the metazoan body through the terminal addition of posteriorly located progenitor cells, and Wnt signals may keep posterior cells in a pluripotent state and sustain posterior proliferative progenitors [51,57]. Posterior growth is accomplished by a group of progenitor cells located in a growth zone at the posterior-most end of the embryo, which continuously provides cells to the growing body [101]. During elongation of the vertebrate embryo, a pool of proliferating precursor cells that are continuously added to the caudal zone is maintained in the most posterior part, the tailbud, creating an anteriorward cell flow through the presomitic mesoderm [42,68,101]. Similarly, in arthropods, the ancestral mode of segmentation is through the sequential addition of segments from the posterior end of the embryonic rudiment, which forms the segment addition zone, i.e., the growth zone [56,57]. Transcription factors of several families including homeodomain proteins have proven to be instrumental for regulating the axial progenitor niche in the embryonic growth zone in vertebrates; a complex genetic network linking these transcription factors and signaling molecules underlies the phenomenon of tissue lengthening from the axial stem cells [58]. At least part of the paraxial mesoderm and neuro-ectoderm of the trunk and tail is generated from a population of progenitors residing within the posterior growth zone [58].
Canonical Wnt signaling dynamically controls multiple stem cell fate decisions during vertebrate body formation; loss of Wnt signaling results in a severely truncated body, which forms only the head and anterior part of the trunk [58,101]. The evolution of Wnt genes has therefore not only an impact for our understanding of stem cell self-renewal, cell differentiation, and apoptosis, but also for the evolution and diversification of the metazoan body plans [9,34,51,54,101]. Generally, posterior growth is a common feature of body formation throughout the animal kingdom [101]. However, posterior growth and terminal addition is certainly not universal in all bilaterian groups: the evolutionary loss, reduction, or modification of terminal addition is documented in several lineages, which have been derived from a basal condition in which a single bilaterian axis generates the body plan [65]. For example, the typical posterior growth zone is absent in Nematoda and Urochordata, having secondary derived modes of development [65,101]. All modern echinoderms have pentaradial symmetry with five ambulacra; if this body pattern is considered a result of main body axis replication, terminal addition refers to growth and patterning in distal subterminal growth zones of each radial arm [65]—for example, five terminal growth zones localized in the arm tips in asteroids [102]. Partial replication of the main body axis also occurs in the pectoral and pelvic limb buds of gnathostome vertebrates (see [65]). These gnathostome limb axes incorporate copies of some but not all Hox genes expressed on the anterior–posterior axis complemented with additional duplicates of the most posterior axial Hox gene; thus, some aspects of the terminal addition process are likely maintained here in the development of these supernumerary axes [65]. Earlier, Minelli considered vertebrate limbs as additional, paramorphic axes [3], i.e., partial duplicates of the main body axis.
4.5. Left–Right Symmetry/Asymmetry in Bilateria: Nodal and Hedgehog Signaling Pathways
To describe a bilaterally symmetrical animal, three systems of spatial coordinates are needed: the position along the anterior–posterior and dorso-ventral body axes, and the position along the medio-lateral axis (left–right disruption of the bilateral symmetry corresponds to the addition of a fourth coordinate) [8]. Morphological asymmetry is common in bilaterians [25,68]. The differences between the right and left body sides in bilaterally symmetrical animals, including the asymmetry of internal organs, are genetically determined by the conserved Nodal signaling system [3,26,33,39,67,68,69].
In bilaterians, the difference in the site of action of the Nodal pathway (left in chordates, right in non-chordates) is most likely due to the dorso-ventral inversion of body axes occurring at the base of the chordate phylogeny and resulting in a concomitant flipping of left and right sides [71]. In the amphioxus (Cephalochordata), the Nodal pathway is conserved in its entirety and similarly to vertebrates, and its members are expressed on the left side [71].
In vertebrates, the first left–right asymmetries can be traced back to an early embryonic period when morphological asymmetries are preceded by molecular asymmetries driven by the action of the Nodal signaling pathway [26,33,39,67,69,71]. Left–right asymmetry in vertebrates is directly linked to the presence of the left–right organizer (for example, the so-named node in mammals) formed during gastrulation at the ventral midline surface of early embryos. A clockwise rotation of motile cilia in the left–right vertebrate organizer generates an asymmetric, leftward flow of surrounding extraembryonic fluid that breaks bilateral left–right symmetry, forming a gradient of the extracellular molecular signaling [26,33,67,69,71]. The gene Lefty1 expresses in the embryonic midline, restricting the expression of Nodal to the left lateral mesodermal plate; ciliar activity in the midline are required to supplement Lefty1 expression and establish the functional midline barrier. Lefty is a Nodal antagonist, and the Nodal/Lefty interaction controlling left–right axis specification amplifies small initial differences between the left and right sides [26,35,67,69].
The maintenance of left–right asymmetry in vertebrates involves Sonic Hedgehog (SHH) signaling, which is necessary for the proper formation of the notochord and the floor plate; Hedgehog signaling pathways is known to regulate many fundamental cellular processes of patterning, cell identity, and environmental responsiveness in all Eumetazoa (see [33,69,103]). In vertebrates, the Hedgehog pathway is extremely important in limb and neural differentiation; besides left–right asymmetry establishment, it is also implicated in a number of roles during embryonic development, including neural crest cell survival and the development of bone, muscle, gonads, and germ cells [104]. Vertebrates possess three paralogues of the Drosophila hedgehog gene: Sonic hedgehog (Shh), Indian hedgehog, and Desert hedgehog; Shh has the broadest range of developmental functions (see [33,103]). The Shh is the most widely used of the three vertebrate homologues and is expressed along the axial mesoderm and the neural floorplate in the embryonic midline, which functions as an organizer region; ciliar motility in the midline is required to establish the midline barrier and maintain left–right asymmetry [33,69,103,105]. The Shh as a midline signal induces symmetry breaking in the presomitic mesoderm; the many roles of the SHH signaling pathway during vertebrate development include morphogenetic processes in the brain, eye, and limb formation [95,106]. The Shh gene is involved in the separation of the single eye field into two bilateral fields. If the expression of the Shh gene is lost or inhibited, the result may be cyclopia (a single eye in the center of the face) and other ocular malformations in vertebrates, including humans [33,103,104,106,107,108]. Such malformations as holoprosencephaly and cyclopia reveal a morphological midline seam between left and right sides, demonstrating these disorders of the midline [103,104,105,107,108].
An essential difference was found in the left–right asymmetry, particularly in the control of somitogenesis between vertebrates and cephalochordates [66,68,71,109]. In the amphioxus, unlike in vertebrates, the left–right asymmetry affects development of the whole body, involving not only the internal, but also the external morphology; all three amphioxus genera (Branchiostoma, Epigonichthys, and Asymmetron) show prominent left–right asymmetry at larval and, to a lesser extent, adult stages [71]. In the cephalochordate Branchiostoma, in contrast to vertebrates, a left/right asymmetry in somite (and later myomeres) arrangement is observed: while the three most anterior somite pairs seem to form simultaneously and appear to be symmetric, the subsequent posterior somites form alternatively on the left and right sides of the embryo, with the left somites forming before the right ones, resulting in the left row being displaced half a segment anteriorly with respect to that on the right side [68,100,109]. Thus, the body segments containing muscle blocks show an offset arrangement, with the left set positioned more anteriorly than the right set [71]. Accordingly, the axons of peripheral nerves that innervate these segments, as well as the neurons within the neural tube, show a comparable offset arrangement; following this asymmetry, axons coming out from the nerve cord slide through the myotome boundaries with a left–right offset until they reach their final destination in peripheral tissues [109]. Neuronal innervation of segmented paraxial structures such as muscle blocks show an offset arrangement between the sides, making it difficult to propose any adaptivity of such state [71].
Thus, in the amphioxus, a left/right asymmetry of most somatic pairs arises during somite formation, and this gliding somitic arrangement is maintained in adult animals [68,100], which is morphologically to some degree similar to the glide reflection symmetry in the metamerim of Ediacaran (Vendian) metazoans (Section 7.1). Larvae develop asymmetrically, as evidenced by the appearance of the mouth on the left side of their bodies [109]. In addition, the anterior somites are formed enterocoelically, and the remainder by schizocoely [68,100]. In addition, the amphioxus has a single, cyclopean eye [100,109]. The situation in the amphioxus may reflect an ancestral deuterostome state [71]. Bertrand et al. [68] also proposed that the ancestral chordate embryo was morphologically close to amphioxus, and left/right somite asymmetry was under the control of the asymmetric expression of the Nodal pathway in posterior somites. In vertebrates, the recruitment of retinoic acid in the control of somitogenesis permitted the acquisition of symmetry through the buffering of the left/right machinery controlled by Nodal [68]. Nodal pathway and Shh midline signaling in the amphioxus are probably less evolutionarily advanced than that in Vertebrata.
5. Helicoidal Symmetry: Nodal Pathway
Helical (spiral) symmetry involves rotation combined with translation along the axis of rotation [19] (i.e., in spiral symmetry, rotation and translation occur together with an expansion); each rotation has an order as the number of repetitions of parts that are arranged around a full circle [14].
In Spiralia, which include annelids, molluscs, and flatworms, “spiral cleavage” produces a highly stereotypic, spiral-like arrangement of blastomeres and swimming trochophore-type larvae with rotational (spiral) symmetry [33,110,111]. Mosaic development, where most blastomeres inherit distinct maternal determinants, is also considered characteristic for the Spiralia [33,111]. Within molluscs, the gastropods represent a drastic example of an asymmetrical body plan with chirality, which is manifested by a dextrally (right-handed or clockwise) versus sinistrally (left-handed or anticlockwise) coiled shell and gut. The Nodal pathway, known to act in the correct left–right patterning in deuterostomes, is involved in defining handedness in gastropods [70,71,110,112]. In snails, the Nodal pathway regulates coiling of the shell, so that it either acts on the right side in dextrally coiling species or on the left side in sinistrally coiling species, with dextrality probably being the ancestral state [70,71,112]. Expression of the genes nodal and Pitx (encoding the downstream acting transcription factor) in gastropod embryos is asymmetric and correlates with their dextrality or synistrality: in embryos of the Lottia gigantea with a dextral body plan, both genes are expressed on the right side of the embryo, while their expression is to the left in sinistral embryos of Biomphalaria glabrata [70,110]. When Nodal signaling was inhibited in embryos of Biomphalaria up to the blastula stage, the expression of the Pitx gene was lost, and tubular, non-coiled shells developed in juvenile snails, while a later exposure (at the trochophore stage) does not prevent the development of normal shells [70,110]. It has been proposed that asymmetric expression of nodal signaling is influenced by the stereotyped positioning of blastomeres (see [43]).
In some pulmonates, such as the snails Lymnaea, Biomphalaria, and Partula, both dextrally and sinistrally coiled individuals occur in the same wild population. In these species, this break in symmetry is maternally inherited [33,110]. This phenomenon is traditionally viewed as a “maternal effect”, i.e., the control of gene expression in the embryo by maternal gene products stored in the ooplasm; thus, in gastropod molluscs, the maternal morphogenetic information determines both the direction of blastomere displacement in early spiral cleavage and the direction of the shell coiling. In the pond snail Lymnaea, the displacement of the blastomere quartet during the third division, clockwise (dextral direction) or counterclockwise (sinistral), coincides with the corresponding direction of dextral or sinistral twisting of the shell. This correlation was shown by T. Morgan in his book “Experimental Embryology” [113] (Figure 2).
Thus, the phenotype of the animal and its shell is determined by the genotype of the mother [110,111,114]. Spatial distribution of maternal molecular information (e.g., mRNA) specifies that the axial polarity of the future body is controlled by components of the egg cytoskeletal-motor system using such motor proteins as kinesin, dynein, and myosins (see [28,33]). The egg cytoskeleton acts as a global morphogenetic determinant, directing and maintaining anisotropy of molecular information distributed in the ooplasm and specifying the axial polarity of egg and future body [17]. The actin filaments and microtubules as components of the cytoskeletal-motor system convert the initial egg asymmetry into global cell polarity; the polarized cell architectonics specifies the embryonic axes [28]. These early events are accomplished by the localization or sequestration of maternally synthesized proteins and mRNAs; such localizations are mediated by specific cytoskeletal structures during late oogenesis or early development [39]. The egg morphogenetic field as an anizotropic distribution of maternal mRNA and transcription factors provides a system of axial coordinates, an egg “regulatory architecture”; this maternal information regulates the differential activity of zygotic target genes in different parts of the developing embryo [22,33,40]. In the frog Xenopus, a maternally inherited, actin filament-dependent chirality was experimentally demonstrated within the egg cortex; a brief disruption of cortical actin during the first cell cycle randomizes the left–right orientation of the tadpole heart and gut [39]. Chiral patterns arising from the self-organization of the actin (actomyosin) cytoskeleton were also shown [115]. In embryogenesis, an ooplasmic “maternal anisotropy” translates into differential zygotic gene expression [33,40].
Spiral-to-Bilateral Transition in Spiralia
The spiral-like displacement of blastomeres in early cleavage demonstrates rotational symmetry [33,111,113,114]. For example, the whole early embryo of the marine annelid Platynereis dumerilii shows a four-fold rotational symmetry around the animal–vegetal axis. Later, spiralian larvae acquire elements of bilateral symmetry, before they metamorphose into fully bilateral juveniles. This transition from embryonic and larval rotational symmetry to the bilateral symmetry of the juvenile was studied by the annelid P. dumerilii in development [111]. It was found that the bilaterally symmetrical parts of the head and larval body emerge from paired bilateral founder cells, which produce bilaterally symmetrical clonal progeny with similar lineage tree topology on the right and left body sides. In particular, the larval brain and head sensory organs in P. dumerilii develop from 11 pairs of bilateral founders, each giving rise to identical clones on the right and left body sides [111]. The conserved head patterning homeobox genes otx and six3 are expressed in bilateral founders giving rise to early differentiating cholinergic neurons and head sensory organs, respectively [111]. Thus, a complex transition from the embryonic spiral cleavage pattern with rotational symmetry to the bilateral symmetry of the early juvenile, involving the interplay between cellular lineage and gene regulatory networks in spiralian development, was found [111].
6. Scale Symmetry
Fractal symmetry is symmetry of scale; scale (fractal) symmetry is also referred to as the symmetry of self-similarity, or as the symmetry of extension [16,19]. The scale symmetry as self-similarity across scaling is a typical feature of fractal hierarchical forms [15,16,19,116]. Various biological structures and processes have been shown to display fractal properties, namely, scale invariance (self-similarity) and fractal dimension [15,21,117,118,119]. The development of repeatedly branched fractal structures is an important mechanism of metazoan morphogenesis, and the scale symmetry as self-similarity across scaling is a typical feature of biological fractal morphogenesis [15,21,22,113,114,115,116,117,118,119]. A complex spatial organization of biological fractal structures can be quantified by the fractal dimension (D or Df) that provides a quantitative view of the spatial complexity of a structure as well as a measure of the degree to which a fractal structure fills the space [15,21,22,117,118,119]. Unlike mathematical fractals, in the living world, fractals are finite, random fractals [19], i.e., “truncated” fractals.
Colonial, clonal animals with repeating modular elements are fractal organisms. For example, the parasitic colonial barnacles (Crustacea: Cirripedia: Rhizocephala) infesting other Crustacea (Decapoda) have a life cycle that includes parasitic and free-living larval stages, with irregular fractal organization at the endoparasitic stage. An adult rhizocephalan parasite (externa) lacks such arthropod features as segmentation, appendages, a digestive tract, and sensory and excretory organs; i.e., the main morphological characteristics of arthropods are absent (see [85,120,121]). The complicated rhizocephalan life cycle resulting from the parasitic mode of life leads to the development of colonial organization in some species of the families Peltogastridae, Sacculinidae, and Duplorbidae, and in all representatives of Thompsoniidae and Polysaccidae [85,120,121]. The colonial organization in the rhizocephalan species emerges at the endoparasitic stage of their life cycle as a result of asexual reproduction by the budding of the interna (root system) with the subsequent development of multiple mature externae [85,120,121]; however, the budding was unambiguously demonstrated only in a few rhizocephalan species.
Using careful dissection of internae under a microscope in the host hemolymph and the subsequent short-term in vitro culture of the internae outside the host organism, we have visualized the fractal internal structure of the barnacles Polyascus polygenea, Peltogasterella gracilis, and Peltogaster reticulatus at their endoparasitic stage, with multiple modular units of trophic (“roots” or “lamp brush”) and reproductive systems integrated into the colonial organism [86,87,122,123]. There is a reduced, chaotic organization of the rhizocephalan body at this stage, including the loss of limbs, segmentation, most tissue systems’ arthropod organization, and the whole axial Bauplan specific of Arthropoda. The budding stolon with several primordial externae and many early buds is evidence of asexual reproduction in P. polygenea without separation of blastozooids. Such clonal, colonial organization is a unique phenomenon among Crustacea, all Arthropoda, and all Ecdysozoa [86,87,122,123]. The colonial organization of the endoparasitic organism of the rhizocephalans P. polygenea is schematically depicted in (Figure 3).
Fractal-like branching structures are typical not only for colonial sedentary organisms. In many metazoan animals, branching is not so evident, being hidden in the internal anatomy [119]; nevertheless, most animal organisms are filled with branching fractal systems, such as the respiratory, excretory, and circulation systems and various glands. Networks of a tubular structure are widespread throughout the animal kingdom [3,124,125,126,127]. Ramifying fractal systems, such as the blood vascular system [126], embedded within the outer epithelial envelope of an organism, can be represented as a complex, high-order manifold with the numerous anastomoses between branches of this embedded system [18]. The development of branching respiratory trees is a classic example of fractal morphogenesis [15,125,126,127,128]. Conserved rules and a conserved genetic machinery seem to control morphogenetic events such as branching and anastomosing of tubes in widely diverging animal clades [3,4,125,126,128].
Genetic studies have led to the identification of key signaling pathways and regulatory genes that control lung fractal morphogenesis, most importantly FGF, BMP, and SHH [119,124,125,129,130]. The human bronchial tree contains millions of branches; in mammalian lungs, the branching process is remarkably stereotyped, and genetic programs are ideally suited to encoding the complex networks of the lung and other branched organs [124]. It was found that the dichotomous branching of epithelial bronchial tubes is partially controlled by a conserved genetic mechanism; each of several initial steps of branching is regulated by an individual gene, but the interaction of the FGF with its receptor is subsequently re-used in determination at many sequential steps of branching both in flies and mammals [124,127,128]. Thus, a fractal algorithm of sequential development of a set of similar scale-invariant modules is an efficient way of morphogenesis based on a relatively small, compressed genetic program [21,22]. However, in the development of the respiratory structures, a complex genetic control has been superimposed on the simple iterativity of a self-similar process [3,4]. In addition, complex branched epithelial structures develop as a self-organized process, reliant upon a strikingly simple generic rule, without a rigid deterministic sequence of genetically programmed events.
The iterated self-similar morphogenesis of branching epithelial channels in the jellyfish Aurelia aurita gastro-vascular system, the gill tracheal system in the larvae of mayflies Siphlonurus immanis and Parameletus chelifer, and neurons of the central nervous system of the fishes Pholidapus dybowskii, Oncorhynchus keta, and O. masou have been studied [17,18,21,22,123,131,132,133] (see also Section 6.1). For the quantitative characteristics of the morphology of the groups of spinal cord neurons in the cherry salmon O. masou during early ontogenesis, the fractal dimension and some more traditional morphometric indices were determined from the first to the second year of fish life [133]. The correlation between the fractal dimension and the main morphometric values of the dendritic trees was found in the neurons investigated. It was shown that the fractal dimension as well as morphometric indices of neuronal morphological complexity increased during the second year of fish life (Figure 4).
This is only a simplified, superficial view of the branching of vertebrate neurons. A branching system of neuron processes is used for neural signal collection, transmission, processing, and integration [119,133]); however, many aspects of branching morphogenesis in neurons remain poorly understood.
The epithelial surface is an interface between the internal medium of an organism and the outside environmental medium (or between two internal compartments), and fractal morphogenesis during metazoan evolution and development increases this interface providing a better adaptation of the organism to the environment as well as a compact packaging of a structure within a small volume, thus providing a certain functional advantage for the organism [18,21,22,119]. Topological and fractal transformations of epithelial systems in the evolution and individual development of multicellular animals increase the interface separating the internal body medium from its environment, thereby enhancing the adaptation of the organism; thus, the fractal structures may be considered as a functionally optimized design in Metazoa (see [21,22]). An important feature of the fractal morphogenesis is its plasticity, which makes possible adaptive responses, in particular, after an injury.
6.1. Fluctuating Asymmetry in Biological Fractal Structures
Biological symmetry is always imperfect. Random non-heritable deviations from perfect symmetry, i.e., stochastic differences among repeated homologous structures within an organism or in populations of organisms, are known as fluctuating asymmetry [14,19,134]. Fluctuating asymmetry usually were considered a measure of developmental noise and developmental instability [19] and/or a useful indicator of stress [14]. Fluctuating asymmetry was studied mainly in bilaterally symmetrical animals [14,19,135]. In Bilateria, fluctuating asymmetry denotes small differences between the left and right sides, i.e., random deviations from left–right symmetry [14,134]. For segmented or radial organisms, other types of body symmetry, more specific of their body architecture, can be used to study developmental stability through the quantification of fluctuating asymmetry [134]. For example, translational fluctuating asymmetry was studied in centipedes (Chilopoda and Geophilomorpha) with high numbers of serially homologous segments, and differences in size and shape patterns were found [134].
Biological fractal structures display stochastic variation, and the biological scale symmetry in branching fractal structures inevitably exhibits irregularity and fluctuating asymmetry. For example, in the ontogenesis of the jellyfish Aurelia aurita (Cnidaria: Scyphozoa), the larvae (Ephyrae) have a primitive digestive system with simple (non-branching) radial canals. In the course of their metamorphosis, differentiation of the radial channels into eight simple and eight branching canals takes place; in this process, each of the branching canals acquires the characteristic form of a trident fork (see [50,131]). Two branching of radial canals in the metaephyra (post-ephyra stage) is uniform and stereotyped. Apparently, only the most general developmental features are strictly specified in the A. aurita gastro-vascular system, i.e., tetra-radial rotation symmetry and the development of eight branching and eight non-branching canals. The essential feature of subsequent branching in gastro-vascular canals is variability during ontogenesis leading to the emergence of fluctuating asymmetry (Figure 5).
In the jellyfish A. aurita, an irregular chaotic branching of its gastro-vascular system results from the asynchronous and topographically variable development of new branches [131]. To evaluate the relationship between order and chaos in the structure of branching fractal canals (known as perradial canals) of the gastro-vascular system in the jellyfish A. aurita, these patterns in the symmetrical parts of each organism (cell clone) were compared with model graphs of deterministic fractal trees [17,21,22,132]. The natural branching pattern of four perradial canals in an individual jellyfish and its representation by standard fractal trees with regular dichotomous branching are shown in Figure 6.
The irregularity of branching (Figure 6) is evident. During ontogenesis, the spatial and temporal variability of branching increases and results in the emergence of chaotic patterns and the breaking of radial symmetry in each individual medusa. Thus, a boundary between order and chaos is revealed in the structural organization of the branching gastro-vascular canals in A. aurita [17,18,21,22,131,132].
Among bilateral animals, data on the fractal-like organization of the respiratory system in the larval tracheal gills of the mayflies Siphlonurus immanis and Parameletus chelifer were collected. The larval tracheal gills are arranged on both sides of the abdomen in mayfly larvae as several pairs of segmental flat petals with a fractal tracheal pattern. The presence of several symmetrical pairs of gill petals in each mayfly larva provides abundant data for quantitative analysis of fluctuating asymmetry in the respiratory system, demonstrating an increasing chaos in the branching tracheal trees [17,21,132]. A chaotic pattern of the larval gill tracheal system is quite evident; the differences in the branching pattern of each pair of gill petals disrupt the bilateral symmetry of the mayfly larval body (Figure 7).
Thus, fractal-like structures of an organism are a record of morphogenetic dynamics including elements of chaos. Spatial and temporal branching variability increases during development, and the cascade of bifurcations leads to irregular branching resulting in a breaking of radial symmetry in each individual medusa and bilateral symmetry in each mayfly larva, with the emergence of fluctuating asymmetry.
7. Body Plan and Symmetry of Ediacaran (Vendian) Metazoans
Over the past several decades, paleontological research has been intensively developing. New local deposits containing interesting accumulations of fossils have been studied and a new paleontological panorama is becoming visible [12,48,136,137]. “The “pageant” of evolution”, “utterly unpredictable and quite unrepeatable” (p. 14 in [137]), included the appearance and disappearance of many creatures of unusual morphology, almost incredible for the modern animal world, and often of fantastic appearance (reflected, in particular, in the name Hallucigenia: e.g., see [138]). The design of these creatures arose as a result of evolutionary “natural experiments” in animal body-plan evolution, in which various possibilities of metazoan morphogenetic transformations were tested [48,136,137,138,139,140,141]. The well-known “Cambrian explosion”, a rapid increase in the diversity of bilaterian animals over a short period on a geological scale, led to the emergence of various body plans and most types of recent Bilateria, such as annelids, arthropods, and chordates [12,48,66,80,136,137,138,139,140]. The methods of comparative genomics make it possible to construct a phylogenetic system of the living world, revealing the connections of animal organisms and even determining the time of evolutionary divergence of different taxonomic branches (“molecular clock”), while the paleontological chronicle dated by radiometric methods allows one to “calibrate” the molecular clock [12,37,48,64,136,140,142,143]. The molecular clock indicates a Precambrian divergence of metazoan branches [48,143] or, more correctly, Ediacaran–Cambrian radiation [12].
For reconstructing deep evolutionary histories, morphology still remains an important source of phylogenetic information and the only means with which to place extinct lineages in a phylogenetic context [144]. Biota of the Ediacaran (Vendian) period immediately preceding the Cambrian consisted of phylogenetically diverse taxa, possibly ranging from microbial colonies, algae, fungi, and protists to animals including bilaterians; these fossil assemblages have been subjected to a variety of taxonomic interpretations [48,65,145]. Most Ediacaran macrofossils are referred to as the extinct kingdom Vendozoa [65] or Vendobionta [11,146] (the terms do not claim to have a taxonomic status). Ediacaran macrofossils were soft-bodied benthic multicellular organisms, mostly without appendages; although many Ediacaran multicellular organisms have been interpreted as coelenterates, there was also a large group of complex animals representing a higher, triploblastic grade of organization with anterior–posterior and dorsal–ventral polarity and composed of serially or fractally arranged elements, which have been termed modules, units, isomers, or segments [11,12,37,48,142,143,144,145,146,147,148,149].
Data on the Ediacaran Biota are very important for understanding the evolution of metazoan development and morphogenesis, including the specification of primary body axes and the appearance of metamerism and/or segmentation [37,50]. However, many Vendobionts had unique body plans, symmetry, and growth patterns, which were never to be repeated later in the Phanerozoic animals [90], and the unusual morphologies of some of these organisms have made it difficult to resolve their relationships to modern metazoan groups [37]. “The Ediacaran biota was not merely a prelude to the Cambrian ‘explosion’, but represents a singular state in the history of our biosphere—strange as life on another planet” [146] (p. 396). Taxonomic affiliation of Vendobionta is problematic and controversial, but in general, the fossils may include stem and crown groups of Metazoa [11,37,48,139,144,150,151], in addition to evolutionary “failed experiments” [151] (p. 250). The majority of Ediacaran fossils probably do not represent stem groups to modern metazoan phyla [144], but it is likely that the last common metazoan ancestor may be found within this “difficult-to-classify diverse group” of early Ediacarans, which “could well be the earliest animals, but with body plans that are rather distinct from modern-day metazoans” [72] (pp. 338–339).
The earliest Ediacaran macrofossils appeared after 570 Ma, with bilaterian metazoans appearing by 550 Ma, and the appearance of these presumed Eumetazoa represented a significant evolutionary transition in the history of life [12,48]. The development of bilateral symmetry was a critical step in the evolution of animal life; this transition from simple forms to the complex animal life and diversification of metazoans is recorded within soft-bodied fossils of the Ediacaran Biota [143]. Some ontogenetic data are available for fossils considered to belong to three morphogroups of the Ediacaran biota: the Rangeomorpha (e.g., Charnia masoni), Dickinsoniomorpha (e.g., Dickinsonia costata), and Erniettomorpha (e.g., Pteridinium simplex) [37,150,152,153]. The authors concluded that the affinities of the rangeomorphs and the dickinsoniomorphs lie within early metazoans [37]. Among Ediacaran fossils, the iconic species Dickinsonia costata was apparently a bilaterally symmetrical form [37,150]. Dickinsoniomorpha are defined as serially repetitive organisms with anterior–posterior differentiation probably capable of active locomotion [37]. Earlier data on the bilateral symmetry—mirror or sliding reflection symmetry—of D. costata were contradictory, but Evans, Droser, and Gehling [150] studied over 900 specimens of D. costata and clearly demonstrated its mirror bilateral symmetry. The D. costata had distally expanding units emanating from a visible central midline; these units are continuous across the midline, imparting a mirror bilateral symmetry [37]. The best-preserved specimens of D. costata clearly show that modules are continuous across the midline and symmetric along the long axis in all specimens, suggesting that this organism is bilaterally symmetrical [150]. The position of the smallest units has often been used to infer the position of a growth zone (or the generative zone), which has been described as being in a posterior, pre-terminal position [37,150]. Evans and coauthors [150] suggested that Dickinsonia represents a previously unrecognized lineage of Eumetazoa, located between sponges and the last common ancestor of Protostomes and Deuterostomes, which may be related to modern Placozoa and likely went extinct prior to the rise of more recognizable animal forms during the Cambrian. The most parsimonious placement for Dickinsonia is as an extinct lineage of Eumetazoa, although the possibility of Dickinsonia as a precursor to bilaterians cannot be ruled out [150]. Dickinsonia appears to possess one major body axis and one lateral axis [37]. If Dickinsoniomorpha are resolved as being segmented, it could be considered as a member of bilaterian metazoans that independently adopted a segmented form [37].
The mollusc-like Kimberella is also a bilaterally symmetric Ediacaran creature, which shows evidence for anterior–posterior and dorsal–ventral differentiation, satisfying some basic criteria of triploblastic bilaterian animals [12,48,90,142,145,148,150,151].
Recently, Evans et al. [143] found the oldest bilaterian Ikaria wariootia, a small, simple, worm-like organism with anterior–posterior differentiation. Authors have suggested that I. wariootia represented one of the earliest bilaterians, with a distinct head and tail, two openings at either end connected by a through gut, and two symmetrical sides. I. wariootia is probably the oldest fossil with this type of complexity [143].
7.1. Lost Ediacaran Body Plans
We consider here such “lost body plans” [154] (p.1), i.e., “lost life forms” [11] (p. 133), as gliding reflection symmetry and fractal branching fronds among Ediacaran metazoans, as well as helicoidal-bodied echinoderms from the early Cambrian (Section 7.2). Among Vendian benthic metazoans, such clades have been identified by M. Fedonkin [90,147] as the type Trilobozoa including the genera Tribrachidium, Albumares, Anfesta, and others, and the type Proarticulata [147], including the genera Dickinsonia, Yorgia, and Vendia [90,147,148]. However, the phylogenetic placement of Dickinsonia and other dickinsoniomorphs remains controversial [37,150]
Trilobozoans are considered diploblastic animals with triradial rotation symmetry, demonstrating a high complexity and morphological diversity [90,141,155]. Presumably, some metameric features were first realised at the diploblastic level of organization [141]. The triradial symmetry in Trilobozoans with three spiral arms [145,155] is sometimes manifested as gliding rotation symmetry, for example, Albumares brunsae (Figure 8). In A. brunsae, triradial spiral symmetry is combined with scale symmetry of fractal structures of a secondary order partitially including elements of fluctuating asymmetry (Figure 8).
Forms with a homonomic segmentation dominated among the fossils, which were interpreted as Bilateria (Triploblastica); there are no examples of heteronomic segmentation among Vendian bilaterian animals [90]. Proarticulata [147] is a type of extinct bilateral, mobile benthic metazoans of the Late Precambrian with a transversely divided body, including such genera as Dickinsonia, Yorgia, and Andiva. A distinctive feature of some representatives of this type is the symmetry of gliding reflection—when the segments are shifted relative to each other along the longitudinal axis [90,147,148,149]. Such repetition of homologous parts along the axis of their body cannot be classified as true segmentation because of the unusual arrangement of the left and right half-segments of the body—the symmetry of gliding reflection, as opposed to mirror symmetry common in true Bilateria [90]. For example, among Proarticalata, Yorgia sp. and Vendia sokolovi demonstrate a symmetry of gliding reflection (Figure 8b,c). In addition, the symmetry of gliding reflection was sometimes combined with the scale symmetry of fractal structures, as in Yorgia (Figure 8b). Similarly to Yorgia but unlike Dickinsonia, the dickinsoniomorph Andiva is not bilaterally symmetrical, bearing its body units aligned along a glide plane of symmetry [36]. Thus, among Proarticulata [90,148,149] (or Dickinsoniomorpha according to [36,78,148], an unusual mutual arrangement of metameres on two sides of the body is often observed, demonstrating not a mirror bilateral symmetry but a symmetry of gliding reflection with a mutual shift of right-sided and left-sided metameres along the anterior–posterior axis of the body [37,90,145,148,150].
Rangeomorphs had a similar glide reflection symmetry pattern: for example, Charnia masoni possessed a body plan comprising one or multiple fronds constructed of serial self-repeated, leaf-like branches, with primary branches lying at a high angle along a glide plane of symmetry running through the central axis of the frond [37,65]. Thus, in Charnia masoni, the symmetry of gliding reflection was combined with the scale (fractal) symmetry; there are also rare reports of bilateral symmetry at higher branching orders in some rangeomorphs, potentially revealing complexity in the axial patterning of these organisms [37]. It is supposed that rangeomorphs had multiple axial growth zones, which contradicts their bilaterian affiliations [37]. Dunn et al. [37] could not rule out a stem-metazoan affinity or a stem-poriferan affinity for rangeomorphs. Rangeomorph metazoans grew by the repetition and branching of self-similar units, creating the characteristic fractal body pattern and achieving maximum surface area by 3D space filling [150]. Such fractal branching organizations of rangeomorph fronds reveal a lost body plan [154].
Metameric organization probably arose early in metazoan history due to the rhythm of body growth or growth pulsation [37,141]. The gliding reflection symmetry conceivably indicates the sequential formation and growth of metameres on the right and left sides in the ontogeny of some Vendobionta as a genetically economic and simple way to increase body size, as well as a probable evolutionary transition from cyclomeria to bilaterality with the simultaneous formation of bilateral symmetry and segmentation [90,141,145]. In Proarticulata (Dickinsoniomorpha), the formation of half-segments (isomers) occurred terminally at the posterior end of the body, indicating a terminal location of the growth zone divided into two transverse rows [90,148]. The most parsimonious explanation for module addition in Dickinsoniomorpha is that they were added at the posterior end [150]; the well-studied Dickinsonia apparently possessed the main growth axis and a pre-terminal growth zone, which can be reconciled with the pre-terminal generative zone of segmented extant bilaterians [37]. If bilaterally symmetrical Dickinsonia and glide symmetrical Andiva and Yorgia are closely related, it is fair to assume they would possess a similarly positioned generative zone [37]. Thus, terminal addition as a morphological and developmental pattern, revealed in both fossil and livinganimals, was the basal condition in Bilateria as a critical component of development [65].
Tissue organization in combination with the anteroposterior and dorsoventral polarity of the Proarticulata body allows us to assign them to the Eumetazoa and Bilateria [148,149]. However, the nature of these fossils is difficult to establish. The interpretation of prints and reconstruction of Vendian animals are not generally accepted among scientists, and are subjects of endless discussion [149]. The Proarticulata resemble arthropods, but completely lack any appendages. In addition, unlike the arthropod segments, the isomers of the Proarticulata are arranged in an alternate order along the longitudinal body axis. This means that serially growing Yorgia and Andiva have the zigzagged median seam [11,146] instead of a rectilinear midline seam of true Bilateria. In many Proarticulata, the overall symmetry is not bilateral, but is approaching the spiral–biradial one; this type of symmetry is not typical for Eumetazoa and Bilateria [90,148,149], but bilaterally symmetric Dickinsonia costata had a midline running parallel to the long axis of the body [150], just as in Bilateria.
The alternating left and right segments and asymmetry of the anterior part, which is so obvious, for example, in Yorgia, resemble the arrangement of the muscle myomeres on the lateral side of the modern cephalochordate amphioxus [90]. In the amphioxus, the earliest chordate, considered a living fossil [104], a left/right asymmetry of most somatic pairs arises during somite formation, and this gliding somitic arrangement is maintained in adult animals [66,68]. Among chordates in vertebrates, the midline seam between left and right sides during somitogenesis and in adult animals is rectilinear, but the amphioxus Branchiostoma have a gliding left/right myomeric position that is morphologically to some degree similar to glide reflection symmetry (see also Section 4.5). Asymmetry may be the ancestral condition, predating the bilateria [25]. Forms of glide symmetry are known also in recent echinoids (e.g., between plates in the interambulacral zone) [23,37].
Ancient metazoans in the pre-Cambrian were probably already genetically and developmentally complex and possessed most of the modern gene families, genomic organization, and gene regulatory networks needed to produce a complex morphology [12,34,36,155]. It was proposed that the hypothetical last common ancestor of animals with bilateral symmetry included perpendicular Wnt and BMP signaling gradients as an ancestral property of Urbilateria [9]. Since the differences between the right and left body sides in bilaterally symmetrical animals, including the asymmetry of internal organs, are genetically determined by the Nodal signaling system, the Nodal signaling pathway was ancestral in segmentation. Notch signaling is a good candidate to drive this hypothetical transition from metamerism to a more advanced segmental organization (see [15,96]).
The emergence of new body plans and most of the main types of Bilateria, an increase in body size, and the complexity of their organization during the Cambrian explosion were probably predetermined by the formation of an integrated cluster of Hox genes and the transformation of genetic regulatory systems [36,48,64,66,72,79]. The Hox-based patterning system operating along the anterior–posterior axis of the Bilateria could have been involved in generating the main body axis in the last common ancestor of the Metazoa [8,52]. Before the appearance of the Hox cluster in Bilateria, other genetic mechanisms of growth and development control functioned in vendobionts; the unusual growth patterns among Vendian bilaterians may reveal unknown steps in the early evolution of the metazoan genome and may provide clues to the evolution of the Homeobox gene complex and Hox genes [90]. The serial homology of many Vendian animals suggests a compartmentalization of gene expression events commonly found in Hox genes [156].
7.2. Early Cambrian Helicoidal-Bodied Echinoderms
Another lost body plan evolved among early Cambrian echinoderms as a group of asymmetric spiral-bodied echinoderms, the oldest echinoderms with ambulacra represented by genera Helicoplacus and Helicocystis (see [11,23,136,157,158]). Helicoplacoids were spindle-shaped animals with a mouth sited laterally, an apical anus, and three ambulacral areas armored by rectangular platelets, which were arranged in spiral rows [11,136]. Smith and Zamora [157] described the spiral-plated Helicocystis moroccoensis that was the most primitive pentaradial echinoderm yet discovered, intermediate between helicoplacoids and the crown group of typical pentaradiate echinoderms. In H. moroccoensis, the spiral body part is composed of ambulacral and interambulacral rows of plates arranged helically, with five ambulacra spiralling clockwise from an apical peristome [157]. Helicocystis shows a novel and unexpected mix of echinoderm characters: it has a spiral body plating as a helicoplacoid but also a terminal mouth and five ambulacra, like crown-group echinoderms [157]. Helicocystis and helicoplacoids have a spirally plated theca, with sheets of polygonal interambulacral plates; however, the direction of spirality differs, being clockwise in Helicocystis and anticlockwise in Helicoplacus [157]. Thus, early fossil echinoderms show intermediate stages in the acquisition of both torsion and pentamery; helicoplacoids show evidence of coelomic torsion and at least three ambulacra [64]. Helicocystis had five ambulacra [157]. The echinoderm fossil record provides critical data on how this novel organization came about by revealing intermediate stages [157].
It has been proposed that the spiral cleavage of the egg and zygote can develop directly in a spiral way of growth at one pole of the embryo and subsequently in a growing adult organism [90], as in mollusks and other Spiralia [Section 5]. Among Deuterostomia, intrinsic chiral properties of the Xenopus egg cortex and the surprising chiral response of the cortex to drug treatment have been experimentally shown—an unusual torsion occurred between separating blastomeres, with the animal pole of each cell rotating several tens of degrees counterclockwise relative to the other, yielding a consistently chiral cleavage pattern [39] (see also Section 5).
7.3. Bilateral-to-Pentameral Transition in Echinodermata
All living echinoderms share a body plan that shows unique pentaradial symmetry, although the larvae remain bilaterally symmetric (see [23,37]). This ontogenetic transformation reflects an evolutionary history in which originally bilaterian ancestors evolved into pentaradiate forms; echinoderms show a dramatic torsion event, with the adult axes bearing no direct relationship with the clearly identifiable axes of the larva [23,59,157]. Echinoderm radial symmetry first appeared in the ambulacral system, when the ambulacral channel assumed the shape of a horseshoe; such hydrocoel determined the primary triradiate symmetry, which transformed into pentaradial symmetry as a result of a branching of two of the three primary radial channels [23]. The evolutionary transformation from a simple hydrocoel with one side channel to an elongated hydrocoel with two side channels, and finally to a nearly closed horseshoe-shaped hydrocoel with three radial channels, can be reconstructed based on the theca structure and the number of ambulacra in the evolutional row, including Helicoplacoidea [23]. In present day, echinoderm radial (usually pentaradial) symmetry with its multiple symmetry planes and a central rotation axis is combined with bilateral symmetry and metamerism. In adult living echinoderms, with unique pentaradial symmetry, metamerism can be observed in some body parts: the ambulacra and interambulacra of echinoids, the stems and arms of crinoids, the stems and brachioles of many blastozoans, the stems and arms of asteroids and ophiuroids, and the ambulacra of holothurians are all metameric [23].
8. Conclusions
Biological morphogenesis always has physical, geometrical, biomechanical, and constructive constraints [140]. The evolutionary success of living organisms depends on the phenotypic adaptation to the surrounding world, including its adequate reflection, from the determination of the basic axes of symmetry in plants and sedentary animals to the mapping of the spatial coordinates of the external world by the mammalian brain (see [45]).
Body axes form the basis of animal body plans, and bilateral symmetry is highly adaptive. The broad variety in animal forms appears to be founded on a few signaling pathways—Wnt, BMP, Notch, and Hedgehog—and on a unique mapping of the Hox cluster metagen into the anteroposterior axis in Bilateria (Section 2). The ancestral Wnt signaling pathway is a crucial program for animal development in the crosstalk with other gene regulatory cascades—not only patterning the primary axis, but also regulating morphogenetic cell movements, i.e., axial differences in cell fate, including the specification of cell lineages, cell proliferation, posterior growth elongation, and organogenesis. The growth of the metazoan body by the terminal addition of progenitor cells localized at the posterior growth zone was a cardinal evolutionary innovation. Since an adult animal body is shaped by the collective behavior of cells during embryonic development [43], we can understand the evolution of body plans using integrative approaches of cell, developmental, and evolutionary biology as well as paleontology. Evolutionary “tinkering” and “failed experiments” [151] resulted in “lost body plans” [154] with wonderful, unrepeated symmetry, such as the gliding reflection symmetry in some Vendian animals and the helicoidal body among early Cambrian echinoderms. Such paleontological data as well as laboratory experiments with a molecular toolkit of axial patterning [38,75] have led to a better understanding of the mechanisms that regulate the axial body plan and its transformations.
Since symmetry is a repetition of parts in different positions and orientations, biological symmetry and other variants of morphological and functional iterations are an effective way of morphogenesis; iterative, a recurrent algorithm is very often in development. The repeated events in the process of segmentation (translational symmetry) and in the course of the branching of biological fractal systems (scale symmetry), with repeated oscillating expression of the same genes or contours of the gene networks, represent an efficient tool of morphogenesis based on a compressed genetic program. Likewise, duplications of genes, gene clusters, whole genomes, and gene network modules based on translational symmetry are widely used in evolution as a simple method of amplification and the subsequent divergence of successful evolutionary innovations [22]. Fractal-like structures of an organism are a record of morphogenetic dynamics including elements of chaos. During branching, a cascade of bifurcations leads to irregular branching, resulting in the breaking of radial or bilateral symmetry of the organism.
Author Contributions
V.V.I.: contributions to the collection of literature data, interpretation of own and literature data, and writing the manuscript. N.V.K.: conceptual contributions, design of the work, illustrations, and help in writing and editing the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research (VVI) was partially funded by Russian Scientific Foundation, grant № 19-14-00346.
Institutional Review Board Statement
Not applicable in the review.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
This work has been supported by a grant, RSF № 19-14-00346, provided by the Russian Scientific Foundation.
Conflicts of Interest
The authors declare that there is no conflict of interest.
References
- Beklemishev, V.N. Principles of Comparative Anatomy of Invertebrates; Beklemishev, V.N., Ed.; Oliver & Boyd: Edinburgh, UK, 1969; Nauka: Moscow, Russia, 1964. (In Russian) [Google Scholar]
- Gerhart, J.C.; Kirschner, M.W. Cells, Embryos, and Evolution; Blackwell Science: Boston, MA, USA, 1997. [Google Scholar]
- Minelli, A. The Development of Animal Form. Ontogeny, Morphology, and Evolution; Cambridge University Press: Cambridge, UK, 2003. [Google Scholar]
- Minelli, A. EvoDevo and its significance for animal evolution and phylogeny. In Evolutionary Developmental Biology of Invertebrates; Wanninger, A., Ed.; Springer: Vienna, Austria, 2015; Volume 1, pp. 1–24. [Google Scholar]
- Ryan, J.F.; Baxevanis, A.D. Hox, Wnt, and the evolution of the primary body axis: Insights from the early-divergent phyla. Biol. Direct. 2007, 2, 37. [Google Scholar] [CrossRef] [Green Version]
- De Robertis, E.M. Evo-devo: Variations on ancestral themes. Cell 2008, 132, 185–195. [Google Scholar] [CrossRef] [Green Version]
- Meinhardt, H. The primary body axes of vertebrates: The generation of a near-Cartesian coordinate system and the role of Spemann-type organizer. Dev. Dyn. 2006, 235, 2907–2919. [Google Scholar] [CrossRef]
- Manuel, M. Early evolution of symmetry and polarity in metazoan body plans. Comptes Rendus Biol. 2009, 332, 184–209. [Google Scholar]
- Niehrs, C. On growth and form: A Cartesian coordinate system of Wnt and BMP signaling specifies bilaterian body axes. Development 2010, 137, 845–857. [Google Scholar] [CrossRef] [Green Version]
- Holló, G. A new paradigm for animal symmetry. Interface Focus 2015, 5, 20150032. [Google Scholar] [CrossRef] [Green Version]
- Seilacher, A.; Gishlick, A.D.S. Morphodynamics; CRC Press/Taylor & Francis Group: Boca Raton, FL, USA, 2015. [Google Scholar]
- Erwin, D.H. The origin of animal body plans: A view from fossil evidence and the regulatory genome. Development 2020, 147, dev182899. [Google Scholar] [CrossRef] [PubMed]
- Fields, C.; Levin, M. Does regeneration recapitulate phylogeny? Planaria as a model of body-axis specification in ancestral eumetazoa. Comm. Integr. Biol. 2020, 13, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Klingenberg, C.P. Analyzing fluctuating asymmetry with geometric morphometrics: Concepts, methods, and applications. Symmetry 2015, 7, 843–934. [Google Scholar]
- Mandelbrot, B.B. The Fractal Geometry of Nature; Freeman: New York, NY, USA, 1983. [Google Scholar]
- Stewart, I. Why Beauty Is Truth: The Story of Symmetry; Perseus: New York, NY, USA, 2007. [Google Scholar]
- Isaeva, V.V.; Kasyanov, N.V.; Presnov, E.V. Analysis situs of spatial-emporal architecture in biological morphogenesis. In Progress in Mathematical Biology Research; Kelly, J.T., Ed.; Nova Science Publishers: New York, NY, USA, 2008; pp. 141–189. [Google Scholar]
- Isaeva, V.V.; Kasyanov, N.V.; Presnov, E.V. Topological singularities and symmetry breaking in development. BioSystems 2012, 109, 280–298. [Google Scholar] [CrossRef] [PubMed]
- Graham, J.H.; Raz, S.; Hel-Or, H.; Nevo, E. Fluctuating asymmetry: Methods, theory, and applications. Symmetry 2010, 2, 466–540. [Google Scholar] [CrossRef] [Green Version]
- Beloussov, L.V. Self-organization, symmetry and morphomechanics in development of organisms. In Embryology–Updates and Highlights on Classic Topics; Pereira, L.A.V., Ed.; InTech: Rijeka, Croatia, 2012; pp. 189–210. [Google Scholar]
- Isaeva, V.V. Topological and fractal approaches to systematics and biodiversity. In Invertebrates: Classification, Evolution and Biodiversity; Riosmena-Rodrigez, R., Ed.; Nova Science Publishers: New York, NY, USA, 2013; pp. 225–242. [Google Scholar]
- Isaeva, V.V. Symmetry transformations in ontogeny and evolution. Paleontol. J. 2014, 48, 1127–1136. [Google Scholar] [CrossRef]
- Rozhnov, S.V. Symmetry of echinoderms: From initial bilaterally-asymmetric metamerism to pentaradiality. Paleontol. J. 2014, 6, 171–183. [Google Scholar] [CrossRef] [Green Version]
- Stern, C.D.; Charité, J.; Deschamps, J.; Duboule, D.; Durston, A.J.; Kmita, M.; Nicolas, J.-F.; Palmeirim, I.; Smith, J.C.; Wolpert, L. Head-tail patterning of the vertebrate embryo: One, two or many unresolved problems? Int. J. Dev. Biol. 2006, 50, 3–15. [Google Scholar] [CrossRef] [PubMed]
- Corballis, M.C. Bilaterally symmetrical: To be or not to be? Symmetry 2020, 12, 326. [Google Scholar] [CrossRef] [Green Version]
- Hirokawa, N.; Tanaka, Y.; Okada, Y. Left–right determination: Involvement of molecular motor KIF3, cilia, and nodal flow. Cold. Spring Harb. Perspect. Biol. 2009, 1, a000802. [Google Scholar] [CrossRef] [Green Version]
- Bouligand, Y. Morphological singulatities and macroevolution. Memor. Soc. Ital. Sci. Natur. Museo Civico Storia Natur. Milano 1996, 27, 89–94. [Google Scholar]
- Li, R.; Bowerman, B. Symmetry breaking in biology. Cold Spring Harb. Perspect. Biol. 2010, 2, a003475.
- Thom, R. Topological models in biology. Topology 1969, 8, 313–335. [Google Scholar] [CrossRef] [Green Version]
- Thom, R. Structural Stability and Morphogenesis: An Outline of a General Theory of Models; Addison-Wesley Publishing Co: New York, NY, USA, 1997. [Google Scholar]
- Bérczi, S.; Lukács, B. Symmetry versus katachi in animal organisms. Forma 2001, 16, 91–99. [Google Scholar]
- Larroux, C.; Fahey, B.; Degnan, S.M.; Adamski, M.; Rokhsar, D.S.; Degnan, B.M. The NK homeobox gene cluster predates the origin of Hox genes. Curr. Biol. 2007, 17, 706–710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilbert, S.F. Developmental Biology, 10th ed.; Sinauer Ass. Inc.: Sunderland, MA, USA, 2010. [Google Scholar]
- Holstein, T.W. The evolution of the Wnt pathway. Cold Spring Harb. Perspect. Biol. 2012, 4, a007922. [Google Scholar] [CrossRef] [PubMed]
- Simakov, O.; Kawashima, T. Independent evolution of genomic characters during major metazoan transitions. Dev. Biol. 2017, 427, 179–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srivastava, M. A Comparative genomics perspective on the origin of multicellularity and early animal evolution. In Evolutionary Transitions to Multicellular Life. Principles and Mechanisms; Ruiz-Trillo, I., Nedelcu, A.M., Eds.; Springer Science+Business Media: Dordrecht, The Netherlands, 2015; pp. 269–300. [Google Scholar]
- Dunn, F.S.; Liu, A.G.; Donoghue, P.C.J. Ediacaran developmental biology. Biol. Rev. 2017, 1. [Google Scholar] [CrossRef] [Green Version]
- Su, Y.-H.; Chen, Y.-C.; Ting, H.-C.; Fan, T.-P.; Lin, C.-Y.; Wang, K.-T.; Yu, J.-K. BMP controls dorsoventral and neural patterning in indirect-developing hemichordates providing insight into a possible origin of chordates. Proc. Natl. Acad. Sci. USA 2019, 116, 12925–12932. [Google Scholar] [CrossRef] [Green Version]
- Danilchik, M.V.; Brown, E.E.; Riegert, K. Intrinsic chiral properties of the Xenopus egg cortex: An early indicator of left–right asymmetry? Development 2006, 133, 4517–4526. [Google Scholar] [CrossRef] [Green Version]
- Davidson, E.H. The Regulatory Genome: Gene Regulatory Networks in Development and Evolution; Academic Press: San Diego, CA, USA, 2006. [Google Scholar]
- Rogers, K.W.; Schier, A.F. Morphogen gradients: From generation to interpretation. Ann. Rev. Cell Dev. Biol. 2011, 27, 377–407. [Google Scholar] [CrossRef] [Green Version]
- Bajard, L.; Morelli, L.G.; Ares, S.; Pécréaux, J.; Jülicher, F.; Oates, A.C. Wnt-regulated dynamics of positional information in zebrafish somitogenesis. Development 2014, 141, 1381–1391. [Google Scholar] [CrossRef] [Green Version]
- Lyons, D.C.; Martindale, M.Q.; Srivastava, M. The cell’s view of animal body-plan evolution. Integr. Comp. Biol. 2014, 54, 658–666. [Google Scholar] [CrossRef] [Green Version]
- Dunn, C.W.; Ryan, J.F. The evolution of animal genomes. Curr. Opin. Genet. Dev. 2015, 54, 25–32. [Google Scholar] [CrossRef]
- Isaeva, V.V. Genomic-morphogenetic correlations and evolutionary trajectories of Bilateria. Paleontol. J. 2018, 52, 1655–1662. [Google Scholar] [CrossRef]
- Isaeva, V.V.; Rozhnov, S.V. Evolutionary transformations of metazoan body plan: Genomic-morphogenetic correlations. Paleontol. J. 2021, 55, in press. [Google Scholar]
- Erwin, D.H. Early origin of the bilaterian developmental toolkit. Phil. Trans. R. Soc. 2009, 364, 2253–2261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erwin, D.H.; Valentine, J.W. The Cambrian Explosion: The Construction of Animal Biodiversity; Roberts & Co.: Greenwood, IN, USA, 2013. [Google Scholar]
- Lang, D.; Rensing, S.A. The evolution of transcriptional regulation in the viridiplantae and its correlation with morphological complexity. In Evolutionary Transitions to Multicellular Life. Principles and Mechanisms; Ruiz-Trillo, I., Nedelcu, A.M., Eds.; Springer Science+Business Media: Dordrecht, The Netherlands, 2015; pp. 301–334. [Google Scholar]
- Technau, U.; Genikhovich, G.; Kraus, J.E.M. Cnidaria. In Evolutionary Developmental Biology of Invertebrates; Wanninger, A., Ed.; Introduction, Non-Bilateria, Acoelomorpha, Xenoturbellida, Chaetognatha; Springer: Vienna, Austria, 2015; Volume 1, pp. 115–163. [Google Scholar]
- Loh, K.M.; van Amerongen, R.; Nusse, R. Generating cellular diversity and spatial form: Wnt signaling and the evolution of multicellular animals. Dev. Cell 2016, 38, 643–655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DuBuc, T.Q.; Bobkov, Y.; Ryan, J.; Martindale, M.Q. The radial expression of dorsal–ventral patterning genes in placozoans, Trichoplax adhaerens, argues for an oral-aboral axis. bioRxiv 2018. [Google Scholar] [CrossRef]
- Lee, P.N.; Pang, K.; Matus, D.Q.; Martindale, M.Q. A Wnt of things to come: Evolution of Wnt signaling and polarity in cnidarians. Sem. Cell Dev. Biol. 2006, 17, 157–167. [Google Scholar] [CrossRef]
- Hogvall, M.; Vellutini, B.C.; Martín-Durán, J.M.; Hejnol, A.; Budd, G.E.; Janssen, R. Embryonic expression of priapulid Wnt genes. Dev. Genes Evol. 2019, 229, 125–135. [Google Scholar] [CrossRef] [Green Version]
- Petersen, C.P.; Reddien, P.W. Wnt signaling and the polarity of the primary body axis. Cell 2009, 139, 1056–1068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chipman, A.D. Hexapoda: Comparative aspects of early development. In Evolutionary Developmental Biology of Invertebrates; Wanninger, A., Ed.; Ecdysozoa III: Hexapoda; Springer: Vienna, Austria, 2015; Volume 5, pp. 92–109. [Google Scholar]
- Constantinou, S.J.; Duan, N.; Nagy, L.M.; Chipman, A.D.; Williams, T.A. Elongation during segmentation shows axial variability, low mitotic rates, and synchronized cell cycle domains in the crustacean, Thamnocephalus platyurus. EvoDevo 2020, 11, 1. [Google Scholar] [CrossRef] [Green Version]
- Neijts, R.; Simmini, S.; Giuliani, F.; van Rooijen, C.; Deschamps, J. Region-specific regulation of posterior axial elongation during vertebrate embryogenesis. Dev. Dyn. 2014, 243, 88–98. [Google Scholar] [CrossRef]
- Angerer, L.M.; Yaguchi, S.; Angerer, R.C.; Burke, R.D. The evolution of nervous system patterning: Insights from sea urchin development. Development 2011, 138, 3613–3623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holló, G.; Novák, M. The manoeuvrability hypothesis to explain the maintenance of bilateral symmetry in animal evolution. Biol. Direct. 2012, 7, 22. Available online: http://www.biology-direct.com/content/7/1/22 (accessed on 10 January 2021).
- He, S.; del Viso, F.C.-Y.; Ikmi, A.; Kroesen, A.E.; Gibson, M.C. An axial Hox code controls tissue segmentation and body patterning in Nematostella vectensis. Science 2018, 361, 1377–1380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holland, L.Z. Tunicates. Curr. Biol. 2016, 26, 146–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stolfi, A.; Brown, F.D. Tunicata. In Evolutionary Developmental Biology of Invertebrates; Wanninger, A., Ed.; Springer: Vienna, Austria, 2015; Volume 6, pp. 135–204. [Google Scholar]
- Peterson, K.J.; Eernisse, D.J. The phylogeny, evolutionary developmental biology, and paleobiology of the Deuterostomia: 25 years of new techniques, new discoveries, and new ideas. Org. Divers. Evol. 2016, 16, 401–418. [Google Scholar] [CrossRef]
- Jacobs, D.K.; Hughes, N.C.; Fitz-Gibbon, S.T.; Winchell, C.J. Terminal addition, the Cambrian radiation and the Phanerozoic evolution of bilaterian form. Evol. Dev. 2005, 7, 498–514. [Google Scholar] [CrossRef]
- Holland, P.W.H. Did homeobox gene duplications contribute to the Cambrian explosion? Zool. Lett. 2015, 1, 1. [Google Scholar] [CrossRef] [Green Version]
- Schier, A.F. Nodal morphogens. Cold Spring Harb. Perspect. Biol. 2009, 1, a003459. [Google Scholar] [CrossRef] [Green Version]
- Bertrand, S.; Aldea, D.; Oulion, S.; Subirana, L.; de Lera, A.R.; Somorjai, I.; Escriva, H. Evolution of the role of RA and FGF signals in the control of somitogenesis in chordates. PLoS ONE 2015, 10, e0136587. [Google Scholar] [CrossRef] [Green Version]
- Shylo, N.A.; Ramrattan, D.; Weatherbee, S.D. Presence of midline cilia supersedes the expression of Lefty1 in forming the midline barrier during the establishment of left–right asymmetry. bioRxiv 2018. [Google Scholar] [CrossRef] [Green Version]
- Grande, C.; Patel, N.H. Lophotrochozoa get into the game: The nodal pathway and left/right asymmetry in Bilateria. Cold Spring Harb. Symp. Quant. Biol. 2009, 74, 281–287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soukup, V. Left–right asymmetry specification in amphioxus: Review and prospects. Int. J. Dev. Biol. 2017, 61, 611–620. [Google Scholar] [CrossRef] [PubMed]
- Ferrier, D.E.K. The origin of the Hox/ParaHox genes, the Ghost Locus hypothesis and the complexity of the first animal. Brief. Funct. Genom. 2016, 15, 333–341. [Google Scholar] [CrossRef] [Green Version]
- Lane, N. The Vital Question: Energy, Evolution, and the Origin of Complex Life; Profile Books: London, UK, 2015. [Google Scholar]
- Koonin, E.V. The Logic of Chance: The Nature and Origin of Biological Evolution; Pearson Education Inc.: Upper Saddle River, NJ, USA, 2012. [Google Scholar]
- Iglesias, M.; Gomez-Skarmeta, J.L.; Saló, E.; Adell, T. Silencing of Smed-beta-catenin1 generates radial-like hypercephalized planarians. Development 2008, 135, 1215–1221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baguña, J.; Martinez, P.; Paps, J.; Riutort, M. Back in time: A new systematic proposal for the Bilateria. Philos. Trans. R. Soc. B Biol. Sci. 2008, 363, 1481–1491. [Google Scholar] [CrossRef] [Green Version]
- Duboule, D. The rise and fall of Hox gene clusters. Development 2007, 134, 2549–2560. [Google Scholar] [CrossRef] [Green Version]
- Darbelay, F.; Duboule, D. Topological domains, metagenes, and the emergence of pleiotropic regulations at Hox loci. Curr. Top. Dev. Biol. 2016, 116, 299–314. [Google Scholar]
- Halanych, K.M. How our view of animal phylogeny was reshaped by molecular approaches: Lessons learned. Org. Divers. Evol. 2016, 16, 319–328. [Google Scholar] [CrossRef]
- Duboule, D. Temporal colinearity and the phylotypic progression: A basis for the stability of a vertebrate Bauplan and the evolution of morphologies through heterochrony. Development 1994, 1994, 135–142. [Google Scholar]
- Tschopp, P.; Duboule, D. A Genetic approach to the transcriptional regulation of Hox gene clusters. Annu. Rev. Genet. 2011, 45, 145–166. [Google Scholar] [CrossRef]
- Sánchez-Herrero, E. Hox targets and cellular functions. Scientifica 2013, 738257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, T.-M.; Kanda, M.; Satoh, N.; Furuya, H. The phylogenetic position of dicyemid mesozoans offers insights into spiralian evolution. Zool. Lett. 2017, 3, 6. [Google Scholar] [CrossRef] [Green Version]
- Zverkov, O.A.; Mikhailov, K.V.; Isaev, S.V.; Rusin, L.Y.; Popova, O.V.; Logacheva, M.D.; Penin, A.A.; Moroz, L.L.; Panchin, Y.V.; Lyubetsky, V.A.; et al. Dicyemida and Orthonectida: Two stories of body plan simplification. Front. Genet. 2019, 10, 443. [Google Scholar] [CrossRef] [PubMed]
- Høeg, J.T.; Deutsch, J.; Chan, B.K.K.; Le Semmler, H. “Crustacea”: Cirripedia. In Evolutionary Developmental Biology of Invertebrates; Wanninger, A., Ed.; Ecdysozoa II: “Crustacea”; Springer: Vienna, Austria, 2015; Volume 4, pp. 153–181. [Google Scholar]
- Shukalyuk, A.I.; Isaeva, V.V. Molecular and subcellular gametogenic machinery of stem and germline cells across Metazoa. In Current Frontiers and Perspectives in Cell Biology; Najman, S., Ed.; InTech: Rieka, Croatia, 2012; pp. 279–314. [Google Scholar]
- Shukalyuk, A.I.; Isaeva, V.V. Molecular signature and sub-cellular machinery of metazoan gametogenic stem cells. In Recent Advances in Germ Cells Research; Perrotte, A., Ed.; Nova Science Publishers Inc.: New York, NY, USA, 2013; pp. 1–40. [Google Scholar]
- Mullins, R.D. Cytoskeletal mechanisms for breaking cell symmetry. Cold Spring Harb. Perspect. Biol. 2010, 2, a003392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacob, F. Evolution and tinkering. Science 1977, 196, 1161–1166. [Google Scholar] [CrossRef] [Green Version]
- Fedonkin, M.A. The origin of the Metazoa in the light of the Proterozoic fossil records. Palentol. Res. 2003, 7, 9–41. [Google Scholar] [CrossRef] [Green Version]
- Nakamoto, A.; Hester, S.D.; Constantinou, S.J.; Blaine, W.G.; Tewksbury, A.B.; Matei, M.T.; Nagy, L.M.; Williams, T.A. Changing cell behaviours during beetle embryogenesis correlates with slowing of segmentation. Nat. Commun. 2015, 6, 6635. [Google Scholar] [CrossRef] [Green Version]
- Maroto, M.; Bone, R.A.; Dale, J.K. Somitogenesis. Development 2012, 139, 2453–2456. [Google Scholar] [CrossRef] [Green Version]
- Oates, A.C.; Morelli, L.G.; Ares, S. Patterning embryos with oscillations: Structure, function and dynamics of the vertebrate segmentation clock. Development 2012, 139, 625–639. [Google Scholar] [CrossRef] [Green Version]
- Aulehla, A.; Pourquié, O. Signaling gradients during paraxial mesoderm development. Cold Spring Harb. Perspect. Biol. 2010, 2, a000869. [Google Scholar] [CrossRef] [Green Version]
- Yin, J.; Saunders, T.E. Shh induces symmetry breaking in the presomitic mesoderm by inducing tissue shear and orientated cell rearrangements. bioRxiv 2019. [Google Scholar] [CrossRef]
- Pueyo, J.I.; Lanfear, R.; Couso, J.P. Ancestral Notch-mediated segmentation revealed in the cockroach Periplaneta americana. Proc. Natl. Acad. Sci. USA 2008, 105, 16614–16619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarrazin, A.F.; Peel, A.D.; Averof, M. A segmentation clock with two-segment periodicity in insects. Science 2012, 336, 338–341. [Google Scholar] [CrossRef] [PubMed]
- Jockusch, E.L.; Smith, F.W. Hexapoda: Comparative aspects of later embryogenesis and metamorphosis. In Evolutionary Developmental Biology of Invertebrates; Wanninger, A., Ed.; Ecdysozoa III: Hexapoda; Springer: Vienna, Austria, 2015; Volume 5, pp. 111–208. [Google Scholar]
- Zakany, J.; Kmita, M.; Alarcon, P.; De La Pompa, J.L.; Duboule, D. Localized and transient transcription of Hox genes suggests a link between patterning and the segmentation clock. Cell 2001, 106, 207–217. [Google Scholar] [CrossRef] [Green Version]
- Holland, L.Z. Cephalochordata. In Evolutionary Developmental Biology of Invertebrates; Wanninger, A., Ed.; Deuterostomia; Springer: Vienna, Austria, 2015; Volume 6, pp. 91–134. [Google Scholar]
- Martin, B.L.; Kimelman, D. Wnt signaling and the evolution of embryonic posterior development. Curr. Biol. 2009, 19, R215–R219. [Google Scholar] [CrossRef] [Green Version]
- Byrne, M.; Martinez, P.; Morris, V. Evolution of a pentameral body plan was not linked to translocation of anterior Hox genes: The echinoderm HOX cluster revisited. Evol. Dev. 2016. [Google Scholar] [CrossRef]
- Cavodeassi, F.; ·Creuzet, S.; Etchevers, H.C. The hedgehog pathway and ocular developmental anomalies. Hum. Genet. 2019, 138, 917–936. [Google Scholar] [CrossRef] [Green Version]
- Abramyan, J. Hedgehog signaling and embryonic craniofacial disorders. J. Dev. Biol. 2019, 7, 9. [Google Scholar] [CrossRef] [Green Version]
- Ertzer, R.; Müller, F.; Hadzhiev, Y.; Vinothkumar, S. Cooperation of sonic hedgehog enhancers in midline expression. Dev. Biol. 2006, 301, 578–589. [Google Scholar] [CrossRef]
- Lumsden, A.; Graham, A. A forward role for Hedgehog. Curr. Biol. 1995, 5, 1347–1350. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, T.; Kinsman, S.; Makris, N.; Grant, E.; Haselgrove, C.; McInerney, S.; Kennedy, D.N.; Takahashi, T.; Fredrickson, K.; Mori, S.; et al. Semilobar holoprosencephaly with midline ‘seam’: A topologic and morphogenetic model based upon MRI analysis. Cereb. Cortex 2003, 13, 1299–1312. [Google Scholar] [CrossRef] [PubMed]
- Sharma, D.; Yadav, J.; Gard, E. Cyclopia syndrome. BMJ Case Rep. 2014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Fernàndez, J.; Benito-Gutiérrez, E. It’s a long way from amphioxus: Descendants of the earliest chordate. BioEssays 2009, 31, 665–675. [Google Scholar] [CrossRef] [PubMed]
- Wanninger, A.; Wollesen, T. Mollusca. In Evolutionary Developmental Biology of Invertebrates; Wanninger, A., Ed.; Springer: Vienna, Austria, 2015; Volume 2, pp. 103–153. [Google Scholar]
- Vopalensky, P.; Tosches, M.A.; Achim, K.; Handberg-Thorsager, M.; Arendt, D. From spiral cleavage to bilateral symmetry: The developmental cell lineage of the annelid brain. BMC Biol. 2019, 17, 81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuroda, R.; Endo, B.; Masanri, A.; Shimuzu, M. Chiral blastomere arrangement dictates zygotic left–right asymmetry pathway in snails. Nature 2009, 462, 790–794. [Google Scholar] [CrossRef] [PubMed]
- Morgan, T.H. Experimental Embryology; Columbia Univ. Press: New York, NY, USA, 1927. [Google Scholar]
- Murray, J.; Clarke, B. The inheritance of polymorphic shell characters in Partula (Gastropoda). Genetics 1966, 54, 1261–1277. [Google Scholar] [CrossRef]
- Tee, Y.H.; Shemesh, T.; Thiagarajan, V.; Hariadi, R.F.; Anderson, K.L.; Page, C.; Volkmann, N.; Hanein, D.; Sivaramakrishnan, S.; Kozlov, M.M.; et al. Cellular chirality arising from the self-organization of the actin cytoskeleton. Nat. Cell Biol. 2015, 17, 445–457. [Google Scholar] [CrossRef]
- Mandelbrot, B.B. Fractals: Form, Chance, and Dimension; W. H. Freeman: San Francisco, CA, USA, 1977. [Google Scholar]
- Weibel, E.R. Fractal geometry–a design principle for living organisms. Am. J. Physiol. 1991, 261, 361–369. [Google Scholar] [CrossRef]
- Peitgen, H.-O.; Jürgens, H.; Saupe, D. Fractals for the Classroom; Spriger: Berlin, Germany, 1992. [Google Scholar]
- Davies, J.A. Why a book on branching, and why now? Afterword. In Branching Morphogenesis; Davies, J.A., Ed.; Springer: Boston, MA, USA, 2006. [Google Scholar]
- Høeg, J.T.; Lützen, J. Life cycle and reproduction in the Cirripedia Rhizocephala. Oceanogr. Mar. Biol. Ann. Rev. 1995, 33, 427–485. [Google Scholar]
- Glenner, H.; Lützen, J.; Takahashi, T. Molecular and morphological evidence for a monophyletic clade of asexually reproducing Rhizocephala: Polyascus, new genus (Cirripedia). J. Crustacean Res. 2003, 23, 548–557. [Google Scholar] [CrossRef] [Green Version]
- Isaeva, V.V. Fractal and chaotic patterns in animal morphogenesis. Transact. Zool. Inst. RAN 2009, 1, 199–218. (In Russian) [Google Scholar]
- Isaeva, V.V.; Akhmadieva, A.V.; Shukalyuk, A.I. The hidden coloniality at the parasitic stage in Peltogaster reticulatus (Crustacea: Rhizocephala). J. Mar. Biol. Assoc. UK 2012, 92, 457–462. [Google Scholar] [CrossRef]
- Metzger, R.J.; Klein, O.D.; Martin, G.R.; Krasnow, M.A. The branching programme of mouse lung development. Nature 2008, 453, 745–750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hannezo, E.; Scheele, C.L.G.J.; Moad, M.; Drogo, N.; Heer, R.; Sampogna, R.V.; van Rheenen, J.; Simons, B.D. A unifying theory of branching morphogenesis. Cell 2017, 171, 242–255. [Google Scholar] [CrossRef] [Green Version]
- Damiani, G. Evolutionary meaning, functions and morphogenesis of branching structures in biology. In Fractals in Biology and Medicine; Nonnenmacher, T.F., Losa, G.A., Weibel, E.R., Eds.; Birkhäuser Verlag: Basel, Switzerland, 1994; pp. 104–115. [Google Scholar]
- Fleury, V.; Watanabe, T.; Nguyen, T.-H.; Unbekandt, M.; Warburton, D.; Dejmek, M.; Nguyen, M.B.; Lindner, A.; Schwartz, L. Physical mechanisms of branching morphogenesis in animals: From viscous fingering to cartilage rings. In Branching Morphogenesis; Davies, J.A., Ed.; Eurekah.com and Springer Science+Business Media: New York, NY, USA, 2006; pp. 202–234. [Google Scholar]
- Warburton, D.; El-Hashash, A.; Carraro, G.; Tiozzo, C.; Sala, F.; Rogers, O.; De Langhe, S.; Kemp, P.J.; Riccardi, D.; Torday, J.; et al. Lung organogenesis. Curr. Top. Dev. Biol. 2010, 90, 73–158. [Google Scholar] [PubMed]
- Chuang, P.; McMahon, A. Branching morphogenesis of the lung: New molecular insights into an old problem. Trends Cell. Biol. 2003, 13, 86–91. [Google Scholar] [CrossRef]
- Menshykau, D.; Kraemer, C.; Iber, D. Branch mode selection during early lung development. PLoS Comput. Biol. 2012, 8, e1002377. [Google Scholar] [CrossRef] [Green Version]
- Chernyshev, A.V.; Isaeva, V.V. Formation of the chaotic patterns of gastro-vascular systems of the jellyfish Aurelia aurita in ontogeny. Russ. J. Mar. Biol. 2002, 28, 377–381. [Google Scholar]
- Isaeva, V.V.; Karetin, Y.A.; Chernyshev, A.V.; Shkuratov, D.Y. Fractals and Chaos in Biological Morphogenesis; Dalnauka: Vladivostok, Russia, 2004. (In Russian) [Google Scholar]
- Isaeva, V.V.; Pushchina, E.V.; Karetin, Y.A. Changes in morphometric parameters and fractal dimensions of spinal marrow neurons in ontogeny of the cherry salmon Oncorhynchus masou. Russ. J. Mar. Biol. 2006, 32, 106–114. [Google Scholar] [CrossRef]
- Savriama, Y.; Vitulo, M.; Gerber, S.; Debat, V.; Fusco, G. Modularity and developmental stability in segmented animals: Variation in translational asymmetry in geophilomorph centipedes. Dev. Genes Evol. 2016. [Google Scholar] [CrossRef]
- Rasskin-Gutman, D.; Izpisua-Belmonte, J.C. Theoretical morphology of developmental asymmetries. BioEssays 2004, 26, 405–412. [Google Scholar] [CrossRef] [PubMed]
- Benton, M.J.; Harper, D.A.T. Introduction to Paleobiology and the Fossil Records; Wiley-Blackwell: Oxford, UK, 2009. [Google Scholar]
- Gould, S.J. Wonderful Life. The Burgess and the Nature of History; Norton Co: New York, NY, USA; London, UK, 1989. [Google Scholar]
- Smith, M. Fossil focus–Hallucigenia and the evolution of animal body plans. Palaeontology 2017, 5, 1–9, Online. [Google Scholar]
- Budd, G.E.; Jackson, I.S.C. Ecological innovations in the Cambrian and the origins of the crown group phyla. Philos. Trans. R. Soc. B Biol. Sci. 2016, 371, 20150287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gould, S.J. The Structure of Evolutionary Theory; Harvard University Press: Cambridge, MA, USA; London, UK, 2002. [Google Scholar]
- Fedonkin, M.A. Metameric features in the Vendian metazoans. Ital. J. Zool. 1998, 65, 11–27. [Google Scholar] [CrossRef]
- Davidson, E.H.; Erwin, D.H. An integrated view of precambrian eumetazoan evolution. Cold Spring Harb. Symp. Quant. Biol. 2009, 74, 65–80. [Google Scholar] [CrossRef] [Green Version]
- Evans, S.D.; Hughes, I.V.; Gehling, J.G.; Droser, M.L. Discovery of the oldest bilaterian from the Ediacaran of South Australia. Proc. Natl. Acad. Sci. USA 2020, 117, 7845–7850. [Google Scholar] [CrossRef]
- Giribet, G. New animal phylogeny: Future challenges for animal phylogeny in the age of phylogenomics. Org. Divers. Evol. 2016, 16, 419–426. [Google Scholar] [CrossRef]
- Xiao, S.; Laflamme, M. On the eve of animal radiation: Phylogeny, ecology and evolution of the Ediacara biota. Trends Ecol. Evol. 2008, 24, 31–40. [Google Scholar] [CrossRef]
- Seilacher, A. The nature of vendobionts. In The Rise and Fall of the Ediacaran Biota; Vickers-Rich, P., Komarower, P., Eds.; Geological Society: London, UK, 2007; Volume 286, pp. 387–397. [Google Scholar]
- Fedonkin, M.A. Precambrian metazoans: The problems of preservation, systematics and evolution. Philos. Trans. R. Soc. Lond. B 1985, 311, 27–45. [Google Scholar]
- Ivantsov, A.Y. The most probable Eumetazoa among late Precambrian macrofossils. Invertebr. Zool. 2017, 14, 127–133. [Google Scholar] [CrossRef]
- Ivantsov, A.Y.; Zakrevskaya, M.A.; Nagovitsyn, A.L. Morphology of integuments of the Precambrian animals, Proarticulata. Invertebr. Zool. 2019, 16, 19–26. [Google Scholar] [CrossRef]
- Evans, S.D.; Droser, M.L.; Gehling, J.G. Highly regulated growth and development of the Ediacara macrofossil Dickinsonia costata. PLoS ONE 2017, 12, e0176874. [Google Scholar] [CrossRef] [PubMed]
- Muscente, A.D.; Boag, T.H.; Bykova, N.; Schiffbauer, J.D. Environmental disturbance, resource availability, and biologic turnover at the dawn of animal life. Earth-Sci. Rev. 2018, 177, 248–264. [Google Scholar] [CrossRef]
- Antcliffe, J.B.; Brasier, M.D. Towards a morphospace for the Ediacara biota. In The Rise and Fall of the Ediacaran Biota; Vickers-Rich, P., Komarower, P., Eds.; Geological Society: London, UK, 2007; Volume 286, pp. 377–386. [Google Scholar]
- Tojo, B.; Saito, R.; Kawakami, S.; Ohno, T. Theoretical morphology of quilt structures in Ediacaran fossils. In The Rise and Fall of the Ediacaran Biota; Vickers-Rich, P., Komarower, P., Eds.; Geological Society: London, UK, 2007; Volume 286, pp. 399–404. [Google Scholar]
- Hoyal Cuthill, J.F.; Conway Morris, S. Fractal branching organizations of Ediacaran rangeomorph fronds reveal a lost Proterozoic body plan. Proc. Natl. Acad. Sci. USA 2014, 111, 13122–13126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivantsov, A.Y.; Leonov, M.V. The Imprints of Vendian Animals–Unique Paleontological Objects of the Arkhangelsk Region; Arkhangelsk, Russia. 2008. Available online: https://www.researchgate.net/publication/260191573 (accessed on 10 January 2021). (In Russian).
- Valentine, J.W. Seeing ghosts: Neoproterozoic bilaterian body plans. In The Rise and Fall of the Ediacaran Biota; Vickers-Rich, P., Komarover, P., Eds.; Geological Society: London, UK, 2007; Volume 286, pp. 369–375. [Google Scholar]
- Smith, A.B.; Zamora, S. Cambrian spiral-plated echinoderms from Gondwana reveal the earliest pentaradial body plan. Proc R Soc B 2013, 280, 20131197. [Google Scholar] [CrossRef] [Green Version]
- Topper, T.P.; Guo, J.; Clausen, S.; Skovsted, C.B.; Zhang, Z. A stem group echinoderm from the basal Cambrian of China and the origins of Ambulacraria. Nat. Commun. 2019. [Google Scholar] [CrossRef]
Figure 1.
A scheme of body symmetry transformation in planarians. (a) An intact planaria; (b) Two-headed bipolar form; (c) “Hypercephalized”, radialized form ((b,c): β-catenin knockdown with RNAi). Adapted from [75].
Figure 1.
A scheme of body symmetry transformation in planarians. (a) An intact planaria; (b) Two-headed bipolar form; (c) “Hypercephalized”, radialized form ((b,c): β-catenin knockdown with RNAi). Adapted from [75].

Figure 2.
The correlation of the direction of blastomere displacement during the third division of the cleavage and shell twisting in the pond snail Lymnaea. Adapted from [113].
Figure 2.
The correlation of the direction of blastomere displacement during the third division of the cleavage and shell twisting in the pond snail Lymnaea. Adapted from [113].

Figure 3.
Schematic image of the fractal colonial organization of the rhizocephalan barnacle Polyascus polygenea inside the host crab. Budding stolons with primordial externae and trophic “roots”. From [122].
Figure 3.
Schematic image of the fractal colonial organization of the rhizocephalan barnacle Polyascus polygenea inside the host crab. Budding stolons with primordial externae and trophic “roots”. From [122].

Figure 4.
Ontogenetic changes of morphology and fractal dimension value of the same class of spinal neurons in the cherry salmon Oncorhynchus masou after the first and second year of fish life. Df: fractal dimension. Adapted from [133].
Figure 4.
Ontogenetic changes of morphology and fractal dimension value of the same class of spinal neurons in the cherry salmon Oncorhynchus masou after the first and second year of fish life. Df: fractal dimension. Adapted from [133].

Figure 5.
The formation of the gastro-vascular system in young Aurelia aurita during early ontogenesis. (a) the youngest medusa, 6 mm in diameter; (b) a medusa, 8 mm; (c) a later stage, 20 mm in diameter. Adapted from [131].
Figure 5.
The formation of the gastro-vascular system in young Aurelia aurita during early ontogenesis. (a) the youngest medusa, 6 mm in diameter; (b) a medusa, 8 mm; (c) a later stage, 20 mm in diameter. Adapted from [131].

Figure 6.
The natural branching pattern of four channels in an individual Aurelia aurita (left) and its presentation as standard fractal trees (right). Numbers of sequential bifurcations are indicated. Adapted from [132].
Figure 6.
The natural branching pattern of four channels in an individual Aurelia aurita (left) and its presentation as standard fractal trees (right). Numbers of sequential bifurcations are indicated. Adapted from [132].

Figure 7.
Variability of the branching pattern of larval tracheal gills in a mayfly Siphlonurus immanis. Fractal patterns in the upper and lower petals from a double larval gill pair. From [132].
Figure 7.
Variability of the branching pattern of larval tracheal gills in a mayfly Siphlonurus immanis. Fractal patterns in the upper and lower petals from a double larval gill pair. From [132].


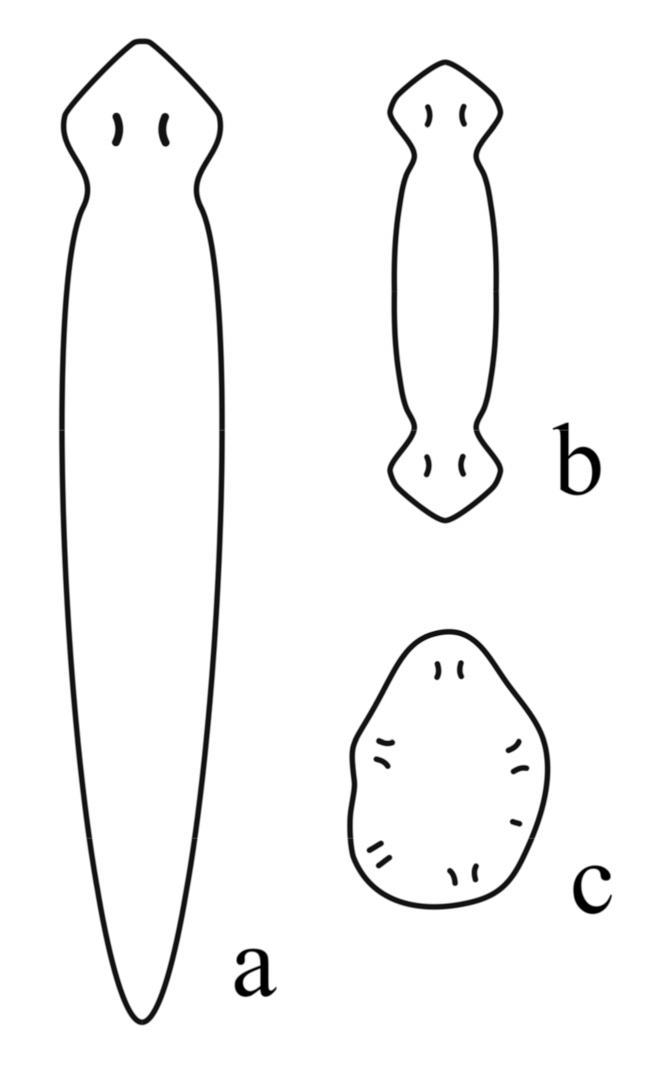{kind=link}
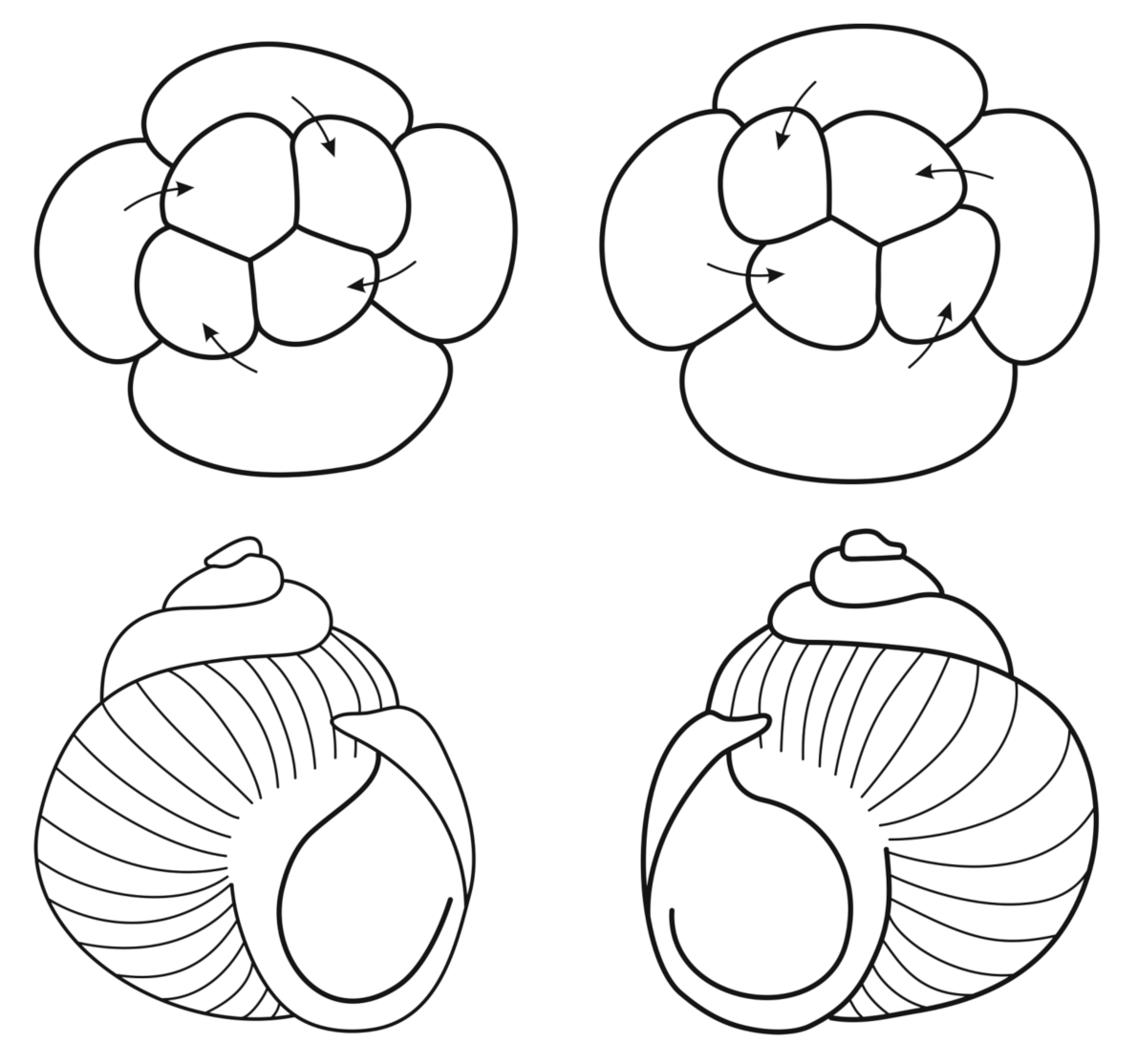{kind=link}
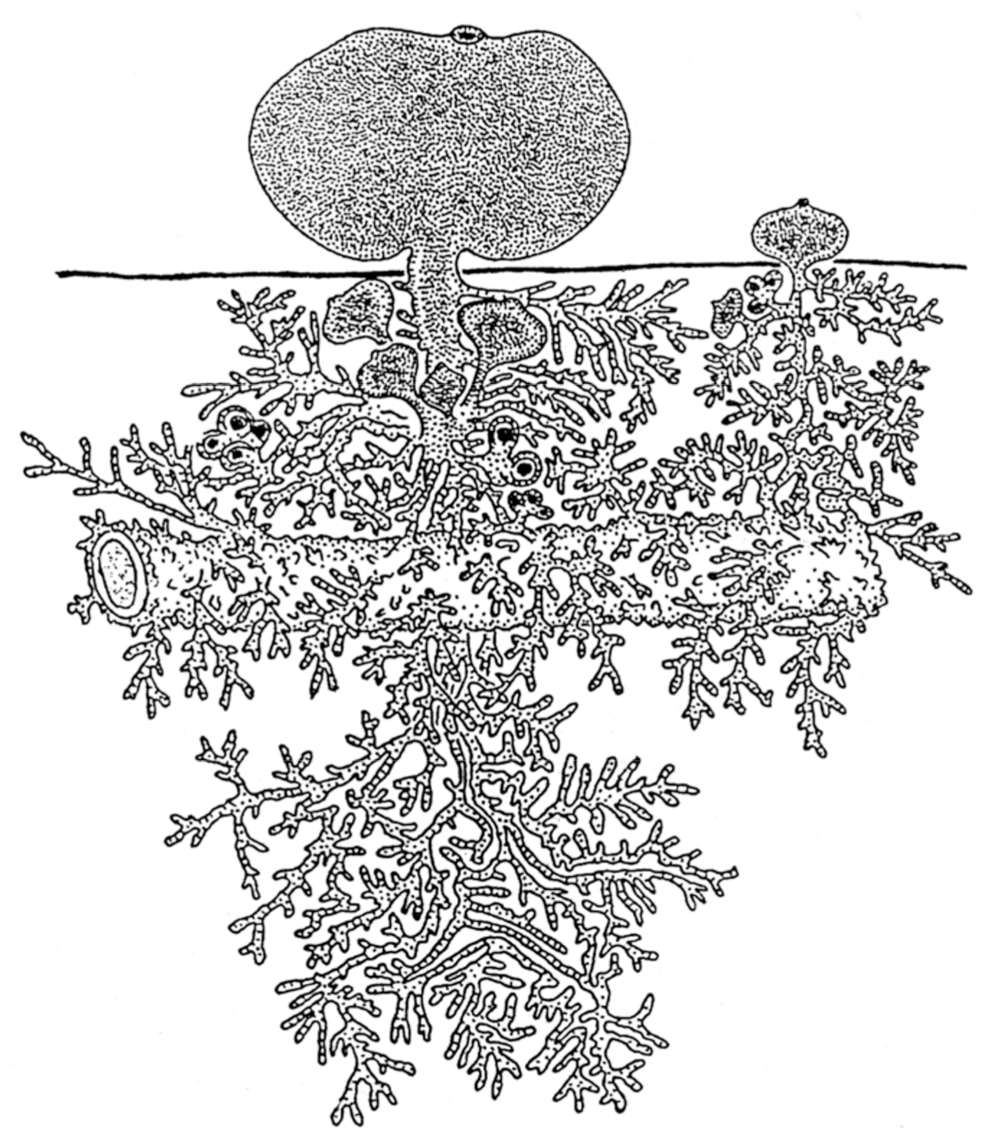{kind=link}
{kind=link}
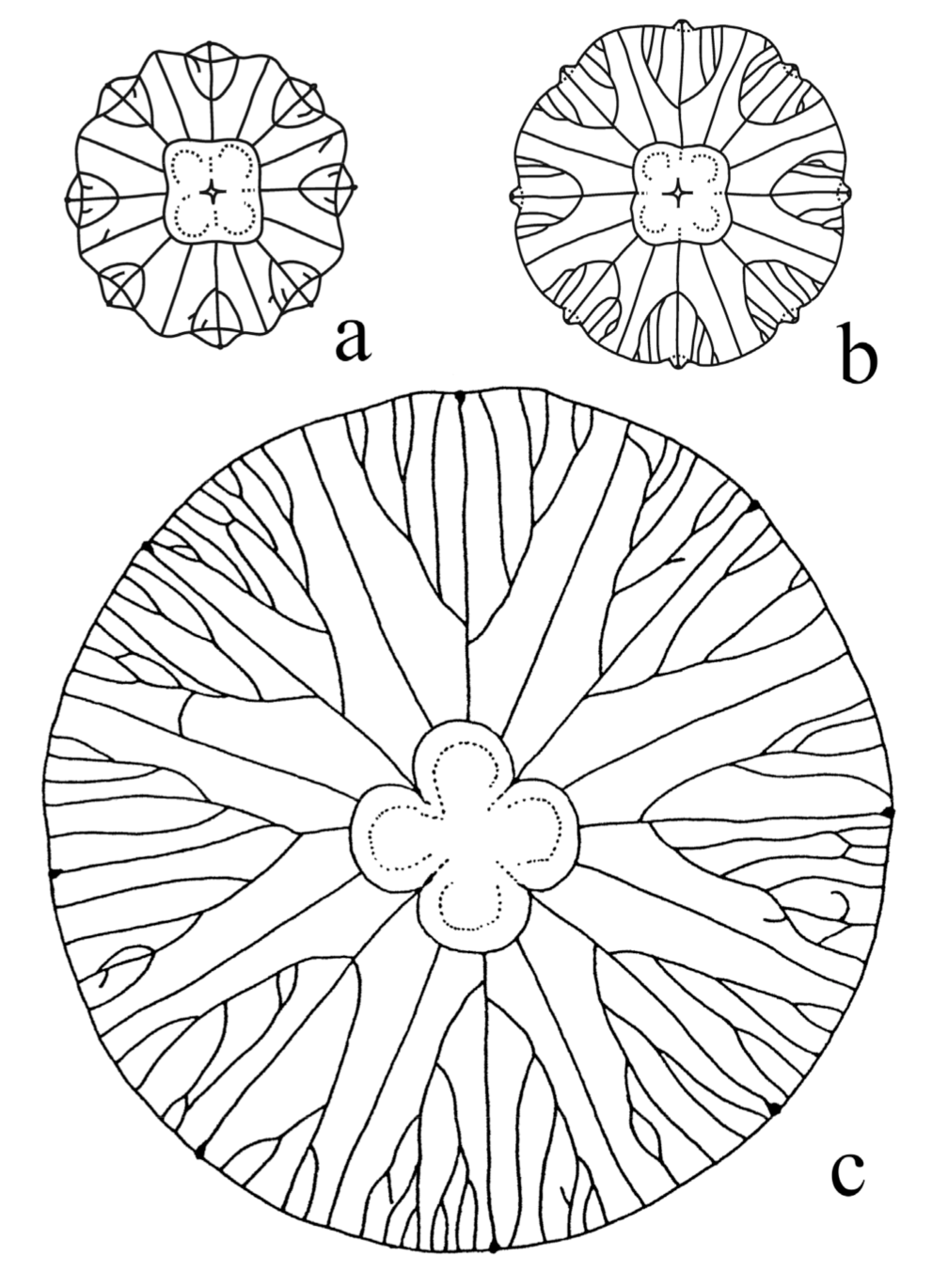{kind=link}
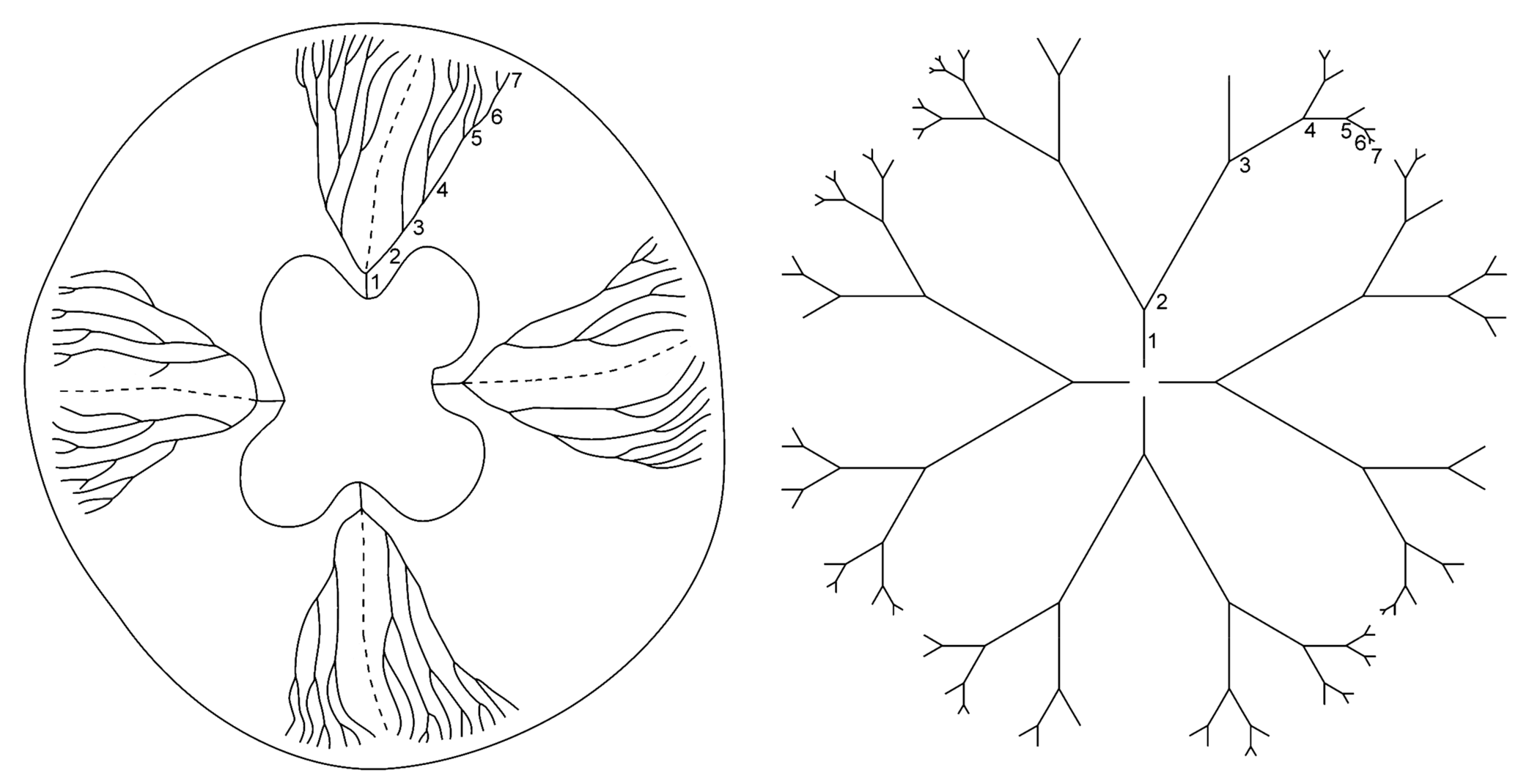{kind=link}
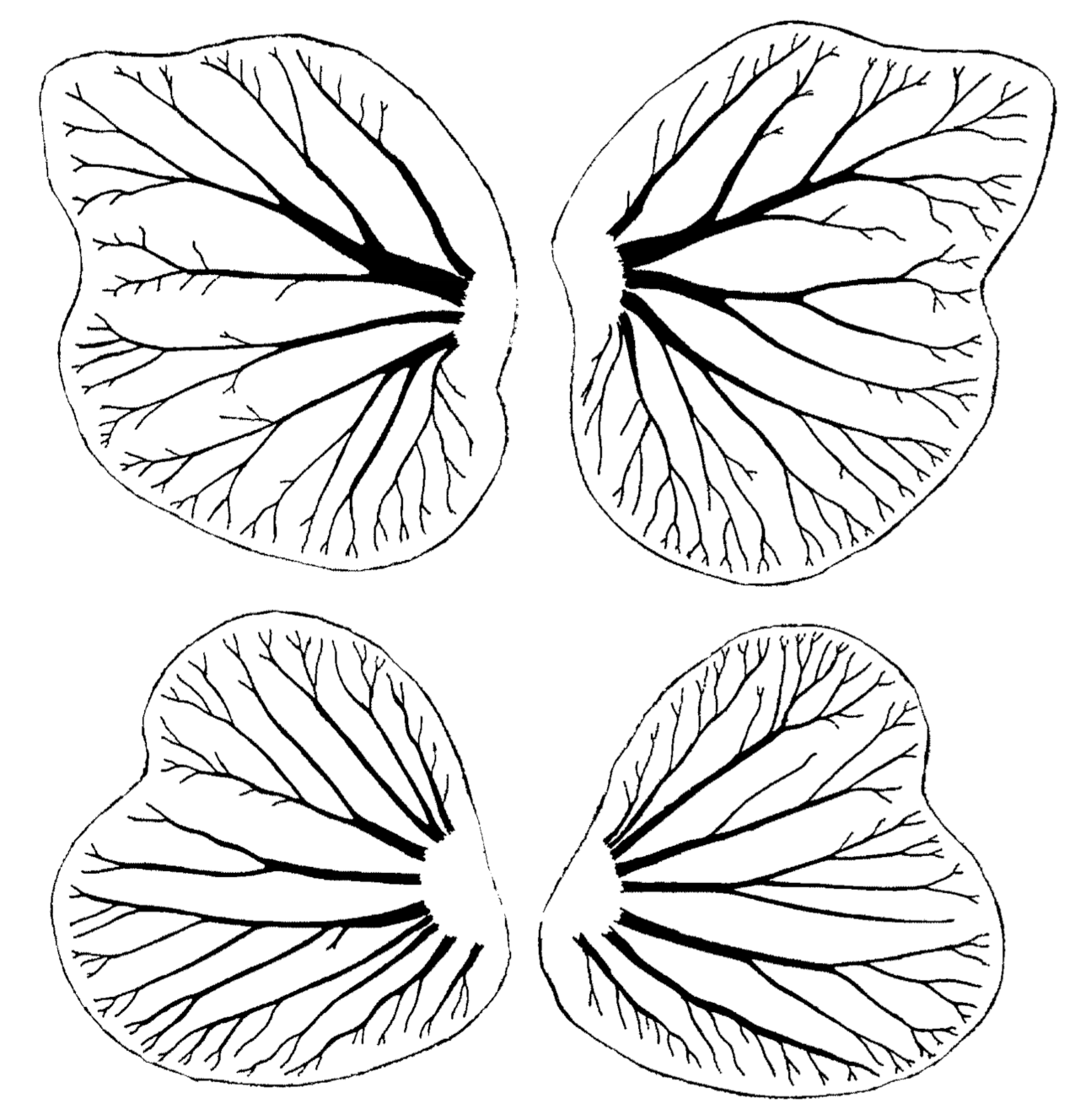{kind=link}
{kind=link}
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Isaeva, V.V.; Kasyanov, N.V. Symmetry Transformations in Metazoan Evolution and Development. Symmetry 2021, 13, 160. https://doi.org/10.3390/sym13020160
AMA Style
Isaeva VV, Kasyanov NV. Symmetry Transformations in Metazoan Evolution and Development. Symmetry. 2021; 13(2):160. https://doi.org/10.3390/sym13020160
Chicago/Turabian StyleIsaeva, Valeria V., and Nickolay V. Kasyanov. 2021. "Symmetry Transformations in Metazoan Evolution and Development" Symmetry 13, no. 2: 160. https://doi.org/10.3390/sym13020160
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.