
In Vitro and In Vivo Evaluation of Three Newly Isolated Bacteriophage Candidates, phiEF7H, phiEF14H1, phiEF19G, for Treatment of Enterococcus faecalis Endophthalmitis

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strain Reagents and Culture Media
2.2. Phage Isolation
2.3. Large-Scale Culture and Purification of Phages phiEF7H, phiEF14H1, and phiEF19G
2.4. Measurement of Phage Host Range
2.5. Electron Microscopy
2.6. Genome Sequencing and Analysis
2.7. Mouse Model of Endophthalmitis
2.7.1. Ethical Treatment of Animals
2.7.2. Mouse Model of Endophthalmitis
2.7.3. Measurement of Viable Bacteria in the Eye
2.7.4. Assay of Myeloperoxidase (MPO) Activity
2.8. Statistics
3. Results
3.1. In Vitro Effects of Phages in Bacteriolysis
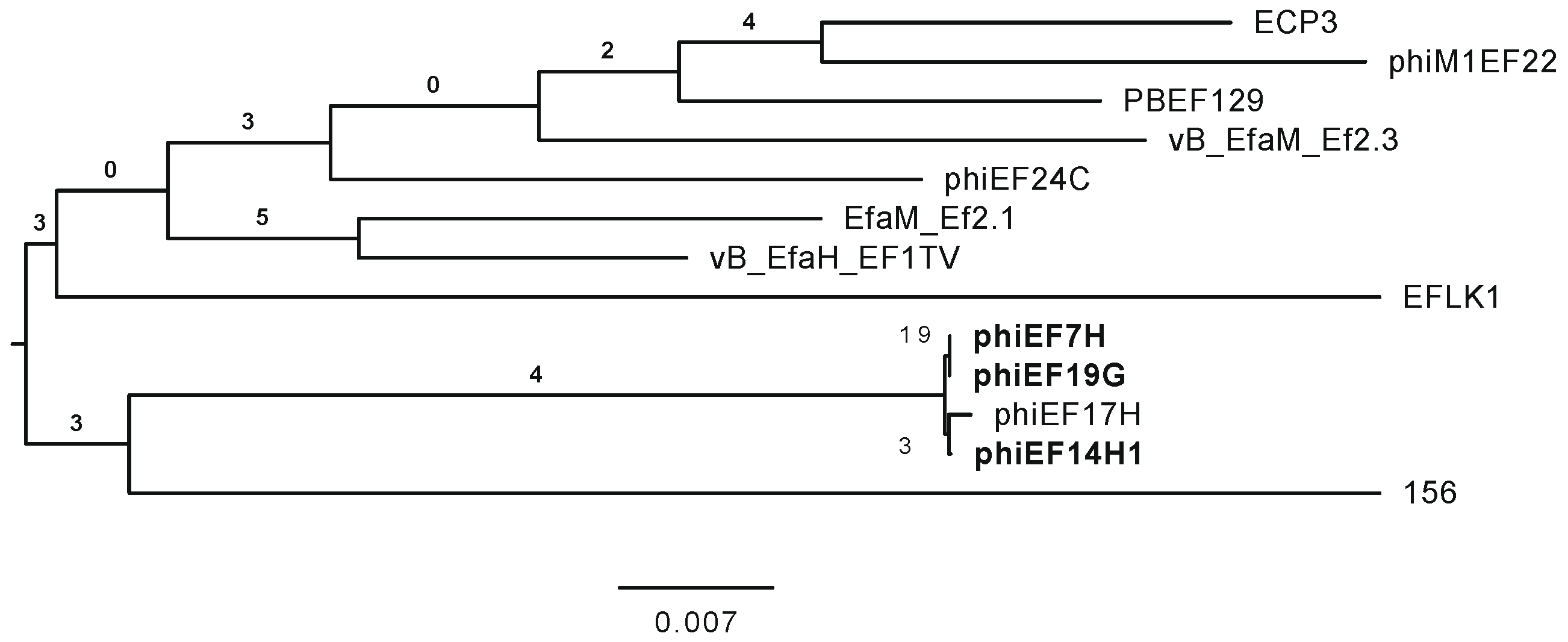
3.2. Characterization of Phages phiEF7H, phiEF14H1, and phiEF19G
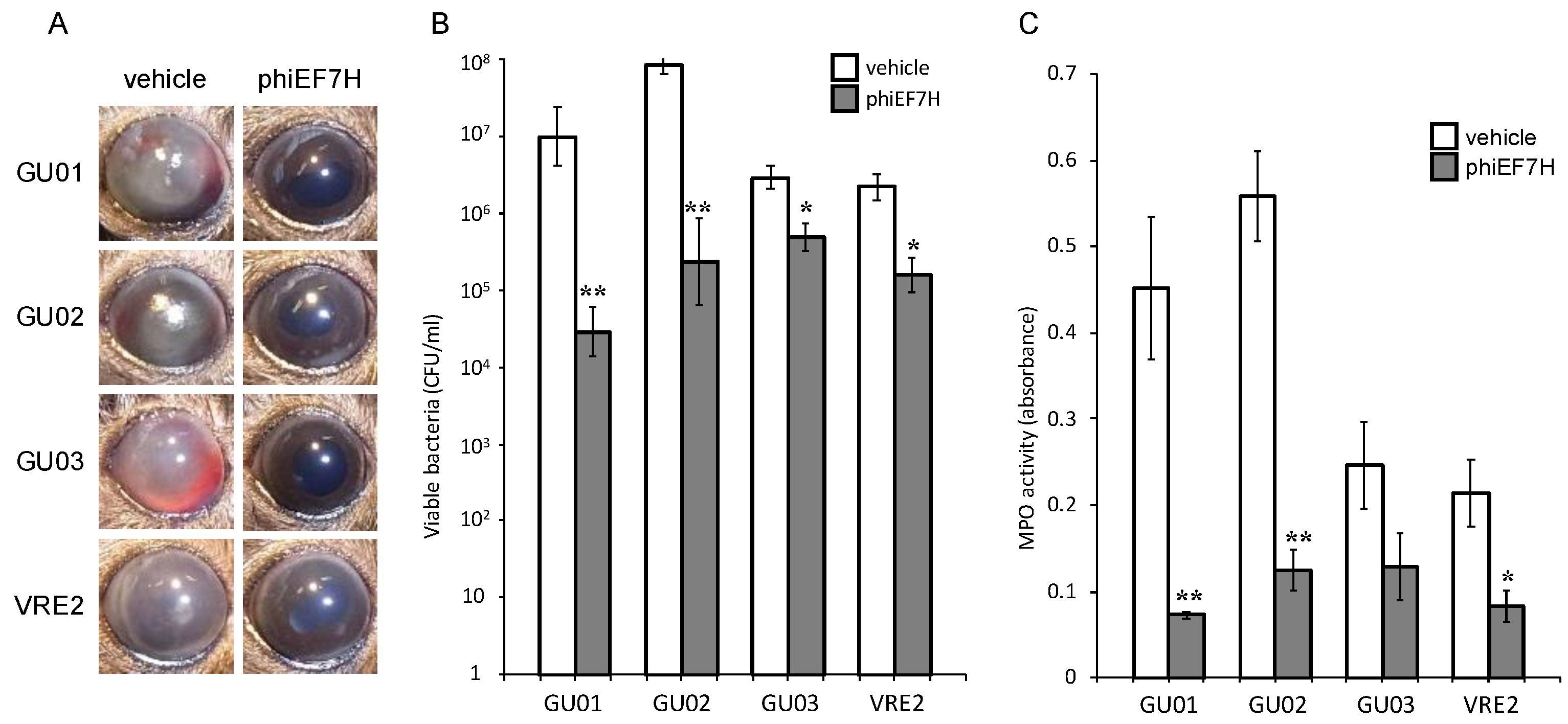
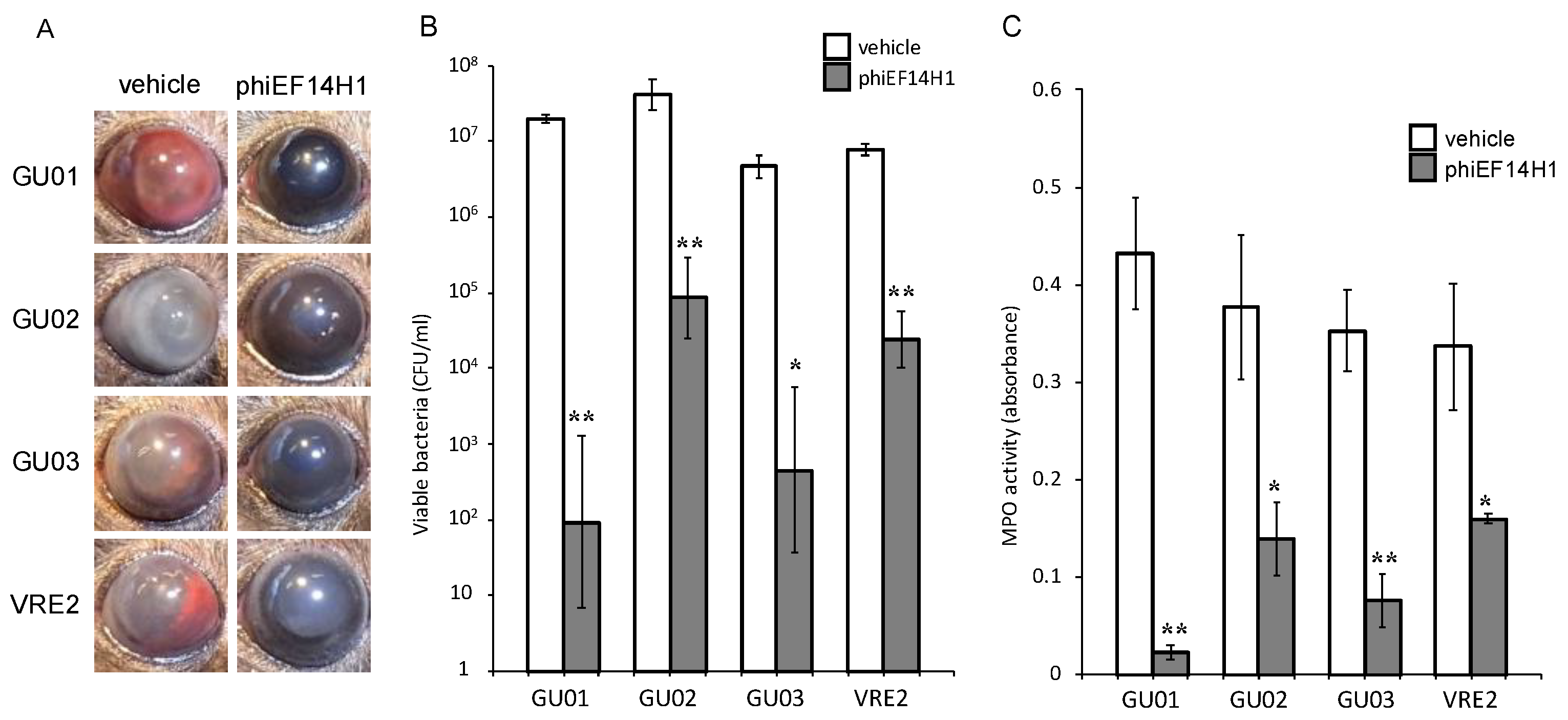
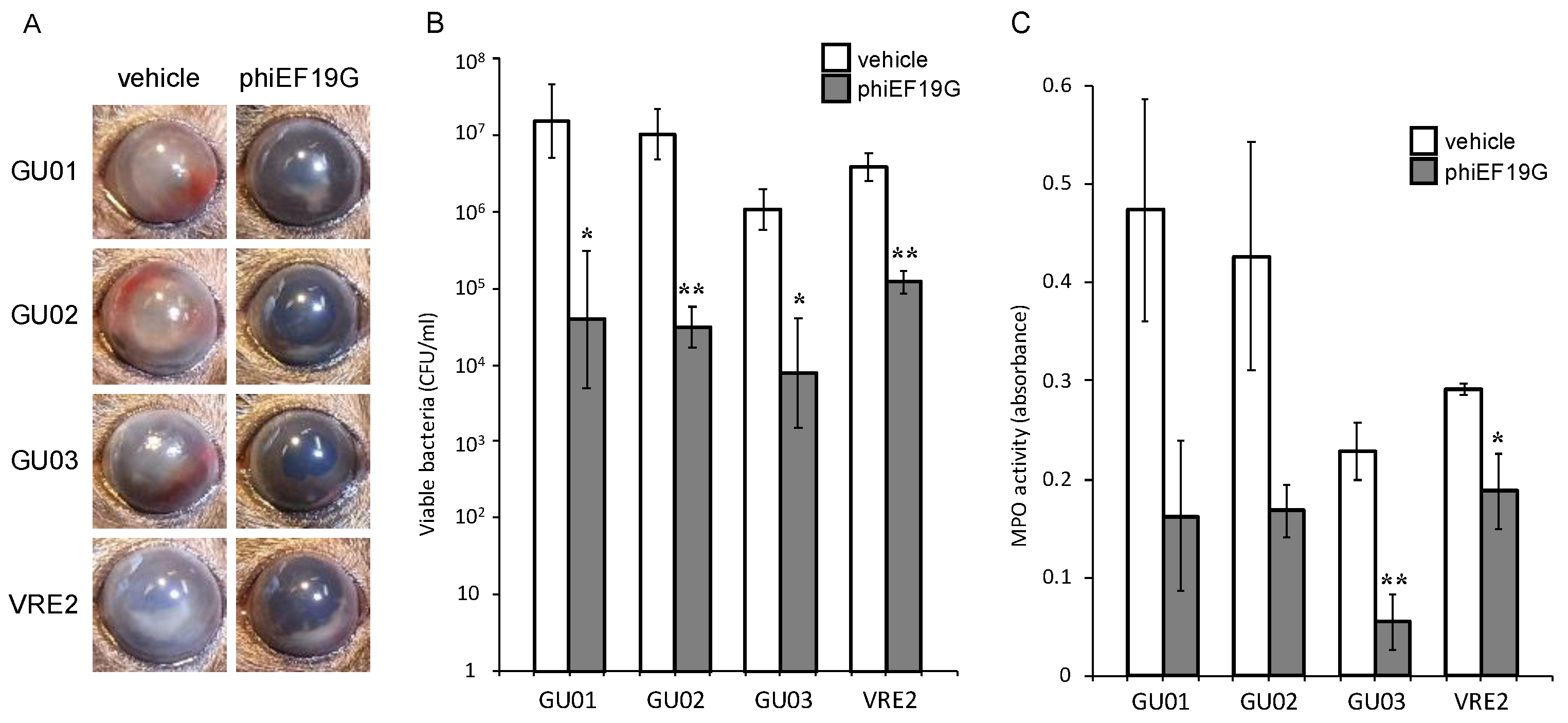
3.3. Effects of Intravitreous Phages on Bacterial Load in E. faecalis Endophthalmitis
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Matsuzaki, S.; Uchiyama, J.; Takemura-Uchiyama, I.; Daibata, M. Perspective: The age of the phage. Nature 2014, 509, S9. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, R.; Matsumoto, T.; Sano, G.; Ishii, Y.; Tateda, K.; Sumiyama, Y.; Uchiyama, J.; Sakurai, S.; Matsuzaki, S.; Imai, S.; et al. Efficacy of bacteriophage therapy against gut-derived sepsis caused by Pseudomonas aeruginosa in mice. Antimicrob. Agents Chemother. 2007, 51, 446–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takemura-Uchiyama, I.; Uchiyama, J.; Osanai, M.; Morimoto, N.; Asagiri, T.; Ujihara, T.; Daibata, M.; Sugiura, T.; Matsuzaki, S. Experimental phage therapy against lethal lung-derived septicemia caused by Staphylococcus aureus in mice. Microbes Infect. 2014, 16, 512–517. [Google Scholar] [CrossRef] [PubMed]
- Kishimoto, T.; Ishida, W.; Fukuda, K.; Nakajima, I.; Suzuki, T.; Uchiyama, J.; Matsuzaki, S.; Todokoro, D.; Daibata, M.; Fukushima, A. Therapeutic Effects of Intravitreously Administered Bacteriophage in a Mouse Model of Endophthalmitis Caused by Vancomycin-Sensitive or -Resistant Enterococcus faecalis. Antimicrob. Agents Chemother. 2019, 63. [Google Scholar] [CrossRef]
- West, E.S.; Behrens, A.; McDonnell, P.J.; Tielsch, J.M.; Schein, O.D. The incidence of endophthalmitis after cataract surgery among the U.S. Medicare population increased between 1994 and 2001. Ophthalmology 2005, 112, 1388–1394. [Google Scholar] [CrossRef]
- Pershing, S.; Lum, F.; Hsu, S.; Kelly, S.; Chiang, M.F.; Rich, W.L., 3rd; Parke, D.W., 2nd. Endophthalmitis after Cataract Surgery in the United States: A Report from the Intelligent Research in Sight Registry, 2013-2017. Ophthalmology 2020, 127, 151–158. [Google Scholar] [CrossRef] [Green Version]
- The Endophthalmitis Vitrectomy Study Group. Microbiologic factors and visual outcome in the endophthalmitis vitrectomy study. Am. J. Ophthalmol. 1996, 122, 830–846. [Google Scholar] [CrossRef]
- Todokoro, D.; Suzuki, T.; Kobayakawa, S.; Tomita, H.; Ohashi, Y.; Akiyama, H. Postoperative Enterococcus faecalis endophthalmitis: Virulence factors leading to poor visual outcome. Jpn. J. Ophthalmol. 2017, 61, 408–414. [Google Scholar] [CrossRef]
- Babalola, O.E. Intravitreal linezolid in the management of vancomycin-resistant enterococcal endophthalmitis. Am. J. Ophthalmol. Case Rep. 2020, 20, 100974. [Google Scholar] [CrossRef]
- Bains, H.S.; Weinberg, D.V.; Feder, R.S.; Noskin, G.A. Postoperative vancomycin-resistant Enterococcus faecium endophthalmitis. Arch. Ophthalmol. 2007, 125, 1292–1293. [Google Scholar] [CrossRef] [Green Version]
- Sharma, S.; Desai, R.U.; Pass, A.B.; Saffra, N.A. Vancomycin-resistant enterococcal endophthalmitis. Arch. Ophthalmol. 2010, 128, 794–795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukuda, K.; Ishida, W.; Uchiyama, J.; Rashel, M.; Kato, S.; Morita, T.; Muraoka, A.; Sumi, T.; Matsuzaki, S.; Daibata, M.; et al. Pseudomonas aeruginosa keratitis in mice: Effects of topical bacteriophage KPP12 administration. PLoS ONE 2012, 7, e47742. [Google Scholar] [CrossRef] [PubMed]
- Uchiyama, J.; Rashel, M.; Maeda, Y.; Takemura, I.; Sugihara, S.; Akechi, K.; Muraoka, A.; Wakiguchi, H.; Matsuzaki, S. Isolation and characterization of a novel Enterococcus faecalis bacteriophage phiEF24C as a therapeutic candidate. FEMS Microbiol. Lett. 2008, 278, 200–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oana, K.; Kawakami, Y.; Ohnishi, M.; Ishikawa, M.; Hirota, M.; Tozuka, M.; Atarashi, K.; Baba, K.; Fujiki, K.; Okazaki, M.; et al. Molecular and epidemiological study of the first outbreak of vanB type vancomycin-resistant Enterococcus faecalis in Japan. Jpn. J. Infect. Dis. 2001, 54, 17–22. [Google Scholar]
- Uchiyama, J.; Takemura, I.; Satoh, M.; Kato, S.; Ujihara, T.; Akechi, K.; Matsuzaki, S.; Daibata, M. Improved adsorption of an Enterococcus faecalis bacteriophage PhiEF24C with a spontaneous point mutation. PLoS ONE 2011, 6, e26648. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Silva, G.G.; Dutilh, B.E.; Matthews, T.D.; Elkins, K.; Schmieder, R.; Dinsdale, E.A.; Edwards, R.A. Combining de novo and reference-guided assembly with scaffold_builder. Source Code Biol. Med. 2013, 8, 23. [Google Scholar] [CrossRef] [Green Version]
- Tanizawa, Y.; Fujisawa, T.; Nakamura, Y. DFAST: A flexible prokaryotic genome annotation pipeline for faster genome publication. Bioinformatics 2018, 34, 1037–1039. [Google Scholar] [CrossRef] [Green Version]
- Meier-Kolthoff, J.P.; Goker, M. VICTOR: Genome-based phylogeny and classification of prokaryotic viruses. Bioinformatics 2017, 33, 3396–3404. [Google Scholar] [CrossRef] [Green Version]
- Uchiyama, J.; Matsui, H.; Murakami, H.; Kato, S.I.; Watanabe, N.; Nasukawa, T.; Mizukami, K.; Ogata, M.; Sakaguchi, M.; Matsuzaki, S.; et al. Potential Application of Bacteriophages in Enrichment Culture for Improved Prenatal Streptococcus agalactiae Screening. Viruses 2018, 10, 552. [Google Scholar] [CrossRef] [Green Version]
- Kang, H.; Kim, S.; Kim, J. Isolation and characterization of an Enterococcus faecalis bacteriophage. Korean J. Microbiol. 2015, 51, 194–198. [Google Scholar] [CrossRef] [Green Version]
- D’Andrea, M.M.; Frezza, D.; Romano, E.; Marmo, P.; Henrici De Angelis, L.; Perini, N.; Thaller, M.C.; Di Lallo, G. The lytic bacteriophage vB_EfaH_EF1TV, a new member of the Herelleviridae family, disrupts biofilm produced by Enterococcus faecalis clinical strains. J. Glob. Antimicrob. Resist. 2020, 21, 68–75. [Google Scholar] [CrossRef] [PubMed]
- Del Rio, B.; Sanchez-Llana, E.; Redruello, B.; Magadan, A.H.; Fernandez, M.; Martin, M.C.; Ladero, V.; Alvarez, M.A. Enterococcus faecalis Bacteriophage 156 Is an Effective Biotechnological Tool for Reducing the Presence of Tyramine and Putrescine in an Experimental Cheese Model. Front. Microbiol. 2019, 10, 566. [Google Scholar] [CrossRef] [PubMed]
- Khalifa, L.; Coppenhagen-Glazer, S.; Shlezinger, M.; Kott-Gutkowski, M.; Adini, O.; Beyth, N.; Hazan, R. Complete Genome Sequence of Enterococcus Bacteriophage EFLK1. Genome Announc. 2015, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cabrera-Aguas, M.; Khoo, P.; George, C.R.R.; Lahra, M.M.; Watson, S.L. Antimicrobial resistance trends in bacterial keratitis over 5 years in Sydney, Australia. Clin. Exp. Ophthalmol. 2020, 48, 183–191. [Google Scholar] [CrossRef]
- Parchand, S.M.; Agrawal, D.; Chatterjee, S.; Gangwe, A.; Mishra, M.; Agrawal, D. Post-cataract surgery cluster endophthalmitis due to multidrug-resistant Pseudomonas aeruginosa: A retrospective cohort study of six clusters. Indian J. Ophthalmol. 2020, 68, 1424–1431. [Google Scholar] [CrossRef]
- Khan, M.; Stapleton, F.; Summers, S.; Rice, S.A.; Willcox, M.D.P. Antibiotic Resistance Characteristics of Pseudomonas aeruginosa Isolated from Keratitis in Australia and India. Antibiotics 2020, 9, 600. [Google Scholar] [CrossRef]
- Alexandrakis, G.; Alfonso, E.C.; Miller, D. Shifting trends in bacterial keratitis in south Florida and emerging resistance to fluoroquinolones. Ophthalmology 2000, 107, 1497–1502. [Google Scholar] [CrossRef]
- Ranjith, K.; SaiAbhilash, C.R.; Sai Prashanthi, G.; Padakandla, S.R.; Sharma, S.; Shivaji, S. Phylogenetic Grouping of Human Ocular Escherichia coli Based on Whole-Genome Sequence Analysis. Microorganisms 2020, 8, 422. [Google Scholar] [CrossRef] [Green Version]
- Pinna, A.; Donadu, M.G.; Usai, D.; Dore, S.; Boscia, F.; Zanetti, S. In Vitro Antimicrobial Activity of a New Ophthalmic Solution Containing Hexamidine Diisethionate 0.05% (Keratosept). Cornea 2020, 39, 1415–1418. [Google Scholar] [CrossRef]
- Daza, J.H.U.; Righetto, G.M.; Chaud, M.V.; da Conceicao Amaro Martins, V.; Lopes Baratella da Cunha Camargo, I.; Maria de Guzzi Plepis, A. PVA/anionic collagen membranes as drug carriers of ciprofloxacin hydrochloride with sustained antibacterial activity and potential use in the treatment of ulcerative keratitis. J. Biomater. Appl. 2020, 35, 301–312. [Google Scholar] [CrossRef] [PubMed]
- Salama, A.H.; AbouSamra, M.M.; Awad, G.E.A.; Mansy, S.S. Promising bioadhesive ofloxacin-loaded polymeric nanoparticles for the treatment of ocular inflammation: Formulation and in vivo evaluation. Drug Deliv. Transl. Res. 2020. [Google Scholar] [CrossRef] [PubMed]
- Fadlallah, A.; Chelala, E.; Legeais, J.M. Corneal Infection Therapy with Topical Bacteriophage Administration. Open Ophthalmol. J. 2015, 9, 167–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Witkin, A.J.; Shah, A.R.; Engstrom, R.E.; Kron-Gray, M.M.; Baumal, C.R.; Johnson, M.W.; Witkin, D.I.; Leung, J.; Albini, T.A.; Moshfeghi, A.A.; et al. Postoperative hemorrhagic occlusive retinal vasculitis: Expanding the clinical spectrum and possible association with vancomycin. Ophthalmology 2015, 122, 1438–1451. [Google Scholar] [CrossRef] [PubMed]
- Todorich, B.; Faia, L.J.; Thanos, A.; Amin, M.; Folberg, R.; Wolfe, J.D.; Todorich, K.M.; Raphtis, E.; Ruby, A.J.; Williams, G.A.; et al. Vancomycin-associated hemorrhagic occlusive retinal vasculitis: A clinical-pthophysiological analysis. Am. J. Ophthalmol. 2018, 188, 131–140. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phage Name | Length (bp) | Accession No. | Isolation | REF. |
|---|---|---|---|---|
| phiEF7H | 143,399 | LC596377 | Japan | This study |
| phiEF14H1 | 143,280 | LC596378 | Japan | This study |
| phiEF19G | 143,400 | LC596379 | Japan | This study |
| phiEF17H | 143,638 | AP018714.1 | Japan | [20] |
| ECP3 | 145,518 | KJ801817.1 | Korea | [21] |
| PBEF129 | 144,230 | MN854830.1 | Korea | Unpublished |
| vB_EfaH_EF1TV | 143,507 | MK268686.1 | Italy | [22] |
| phiM1EF22 | 143,046 | AP018715.1 | Japan | [20] |
| vB_EfaM_Ef2.3 | 147,289 | MK721192.1 | USA | Unpublished |
| vB_EfaM_Ef2.1 | 140,938 | MK693030.1 | USA | Unpublished |
| phiEF24C | 142,072 | AP009390.1 | Japan | [13] |
| 156 | 141,133 | LR031359.1 | Spain | [23] |
| EFLK1 | 130,952 | NC_029026.1 | Israel | [24] |
| Host | Isolated Source | Phage | |||
|---|---|---|---|---|---|
| phiEF7H | phiEF14H1 | phiEF19G | phiEF24C | ||
| EF1 | Vaginal discharge | + | + | + | + |
| EF2 | Vaginal discharge | + | + | + | + |
| EF3 | Sputum | + | + | + | + |
| EF4 | Pharyngis | + | + | + | LFW |
| EF5 | Skin | + | + | + | + |
| EF6 | Urine | + | + | + | + |
| EF7 | Eye discharge | + | + | + | + |
| EF8 | Other | + | + | + | + |
| EF9 | Urine | - | - | - | + |
| EF10 | Other | + | + | + | - |
| EF11 | Urine | + | + | + | LFW |
| EF12 | Urine | + | + | + | - |
| EF13 | Pharyngis | - | - | - | - |
| EF14 | Vaginal discharge | + | + | + | + |
| EF15 | Pus | + | + | + | - |
| EF16 | Vaginal discharge | + | + | + | + |
| EF17 | Vaginal discharge | + | + | + | + |
| EF18 | Vaginal discharge | + | + | + | + |
| EF19 | Vaginal discharge | + | + | + | + |
| EF20 | Pus | LFW | - | - | + |
| EF21 | Pus | + | + | + | + |
| EF22 | Mouthwash | + | + | + | + |
| EF23 | Sputum | + | + | + | + |
| EF24 | Vaginal discharge | + | + | + | + |
| EF25 | Other | + | + | + | + |
| EF26 | Vaginal discharge | + | + | + | + |
| EF27 | Eye discharge | LFW | - | - | + |
| EF28 | Vaginal discharge | + | + | + | + |
| EF29 | Pus | + | + | + | + |
| EF30 | Vaginal discharge | + | + | + | - |
| GU01 | Endopthalmitis | + | + | + | + |
| GU02 | Endopthalmitis | + | + | + | LFW |
| GU03 | Endopthalmitis | + | + | + | LFW |
| GU04 | Endopthalmitis | + | + | + | - |
| GU05 | Endopthalmitis | + | + | + | - |
| VRE1 | HGH | + | + | + | - |
| VRE2 | HGH | + | + | + | + |
| VRE3 | HGH | + | + | + | + |
| VRE4 | HGH | + | + | + | + |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kishimoto, T.; Ishida, W.; Nasukawa, T.; Ujihara, T.; Nakajima, I.; Suzuki, T.; Uchiyama, J.; Todokoro, D.; Daibata, M.; Fukushima, A.; et al. In Vitro and In Vivo Evaluation of Three Newly Isolated Bacteriophage Candidates, phiEF7H, phiEF14H1, phiEF19G, for Treatment of Enterococcus faecalis Endophthalmitis. Microorganisms 2021, 9, 212. https://doi.org/10.3390/microorganisms9020212
Kishimoto T, Ishida W, Nasukawa T, Ujihara T, Nakajima I, Suzuki T, Uchiyama J, Todokoro D, Daibata M, Fukushima A, et al. In Vitro and In Vivo Evaluation of Three Newly Isolated Bacteriophage Candidates, phiEF7H, phiEF14H1, phiEF19G, for Treatment of Enterococcus faecalis Endophthalmitis. Microorganisms. 2021; 9(2):212. https://doi.org/10.3390/microorganisms9020212
Chicago/Turabian StyleKishimoto, Tatsuma, Waka Ishida, Tadahiro Nasukawa, Takako Ujihara, Isana Nakajima, Takashi Suzuki, Jumpei Uchiyama, Daisuke Todokoro, Masanori Daibata, Atsuki Fukushima, and et al. 2021. "In Vitro and In Vivo Evaluation of Three Newly Isolated Bacteriophage Candidates, phiEF7H, phiEF14H1, phiEF19G, for Treatment of Enterococcus faecalis Endophthalmitis" Microorganisms 9, no. 2: 212. https://doi.org/10.3390/microorganisms9020212