Phototropin 1 and 2 Influence Photosynthesis, UV-C Induced Photooxidative Stress Responses, and Cell Death

 , , ,
, , ,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growth Conditions
2.2. Morphological and Physiological Traits Determination
2.3. Chlorophyll a Fluorescence Parameter Measurements
2.4. Photosynthetic Pigment Composition Analysis
2.5. Hydrogen Peroxide Levels Determination
2.6. Protein Extraction and Enzyme Activity Measurements
2.7. UV-C Treatment
2.8. Cell Death Analysis by Electrolyte Leakage and Evans Blue Staining
2.9. RNA Extraction and cDNA Synthesis
2.10. Relative Gene Expression Measurement by Real-Time qPCR
2.11. RNA Sequencing and RNA-Seq Analysis
3. Results
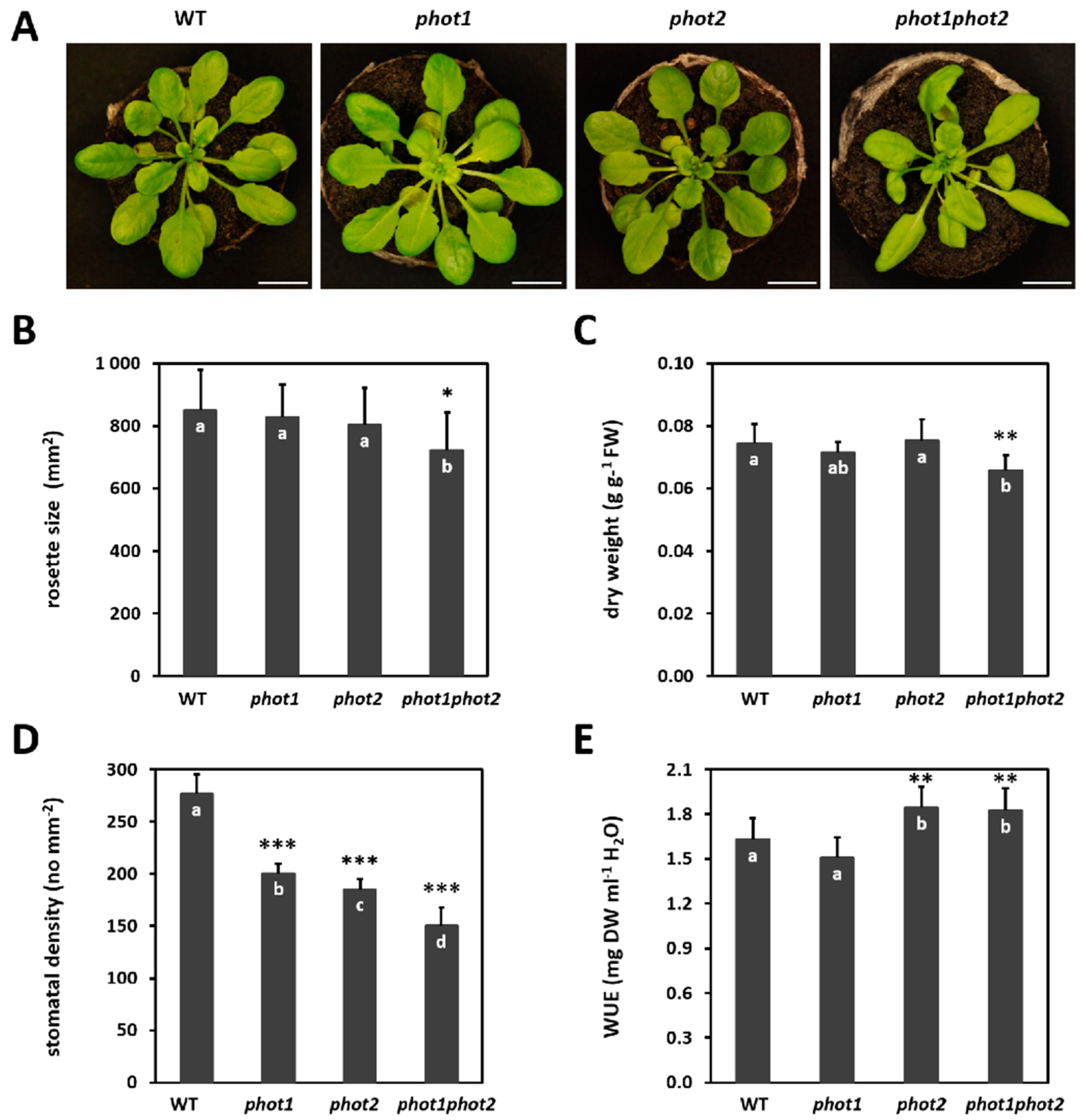
3.1. phot1 and phot2 Have an Impact on Rosette Size, Plant Dry Mass, Stomata Density, and Water Use Efficiency
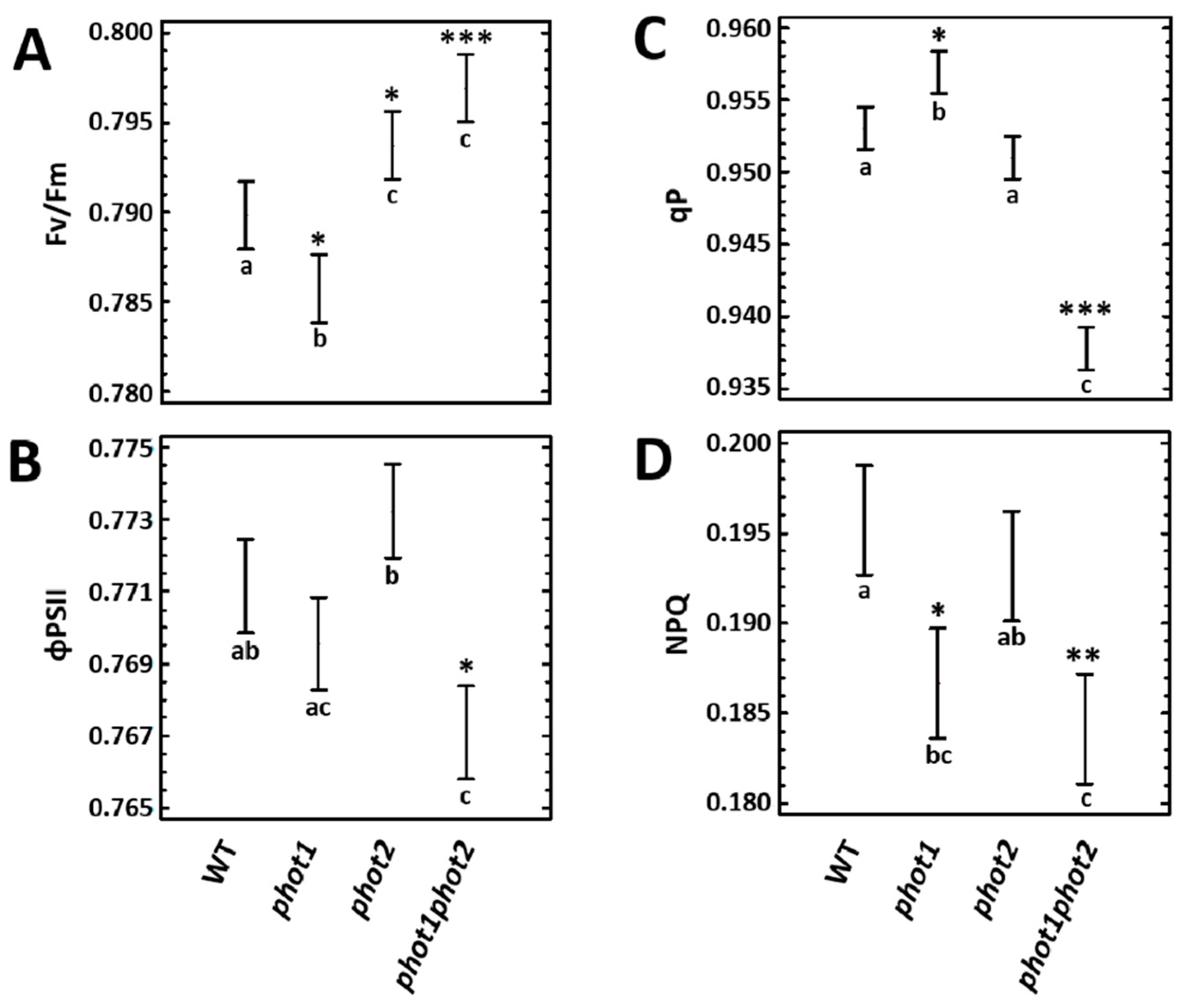
3.2. phot1 and phot2 Influence Photosynthetic Parameters and Pigment Composition
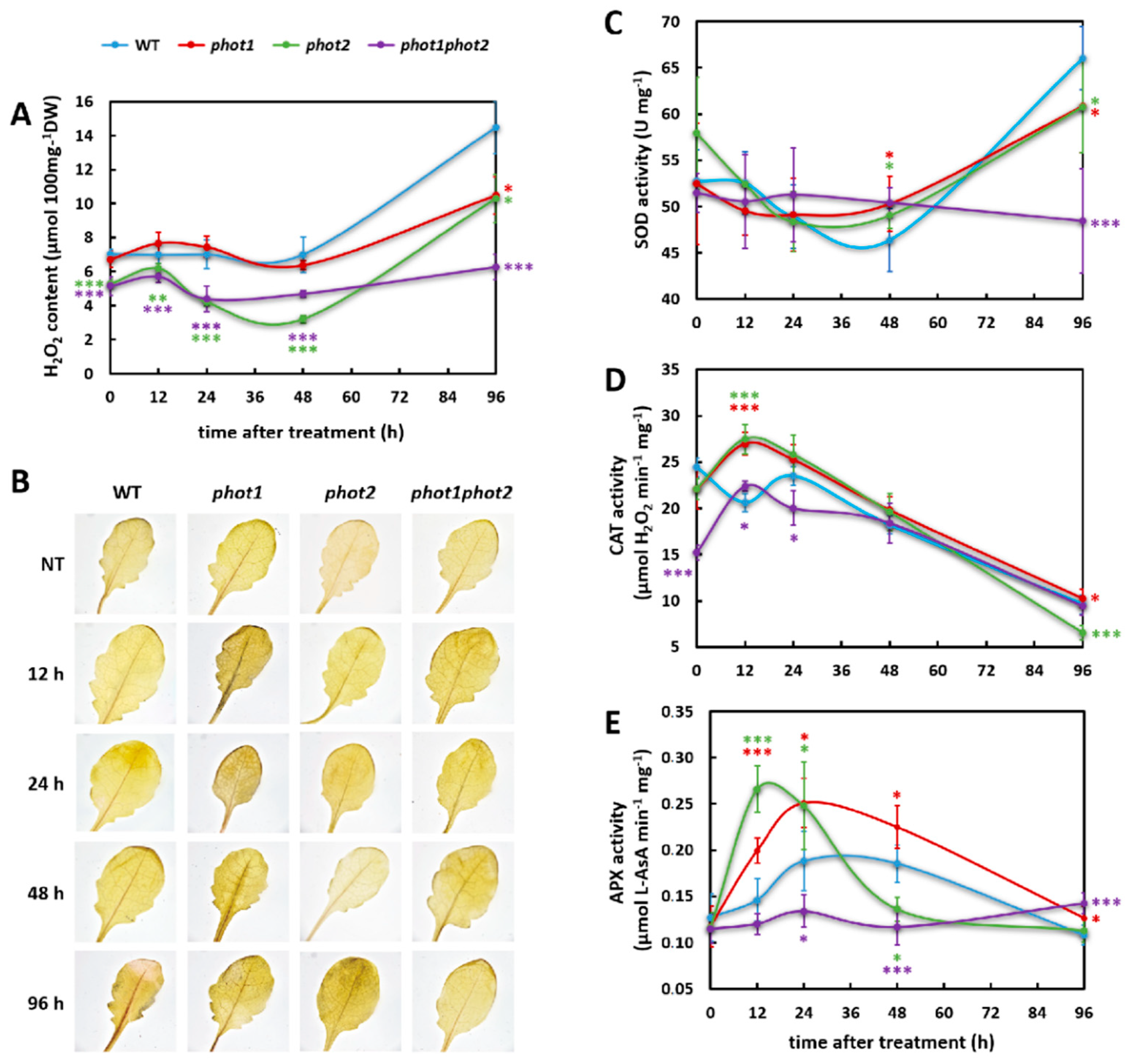
3.3. phot1 and phot2 Affect Foliar H2O2 Homeostasis in Non-Stress and Oxidative Stress Conditions
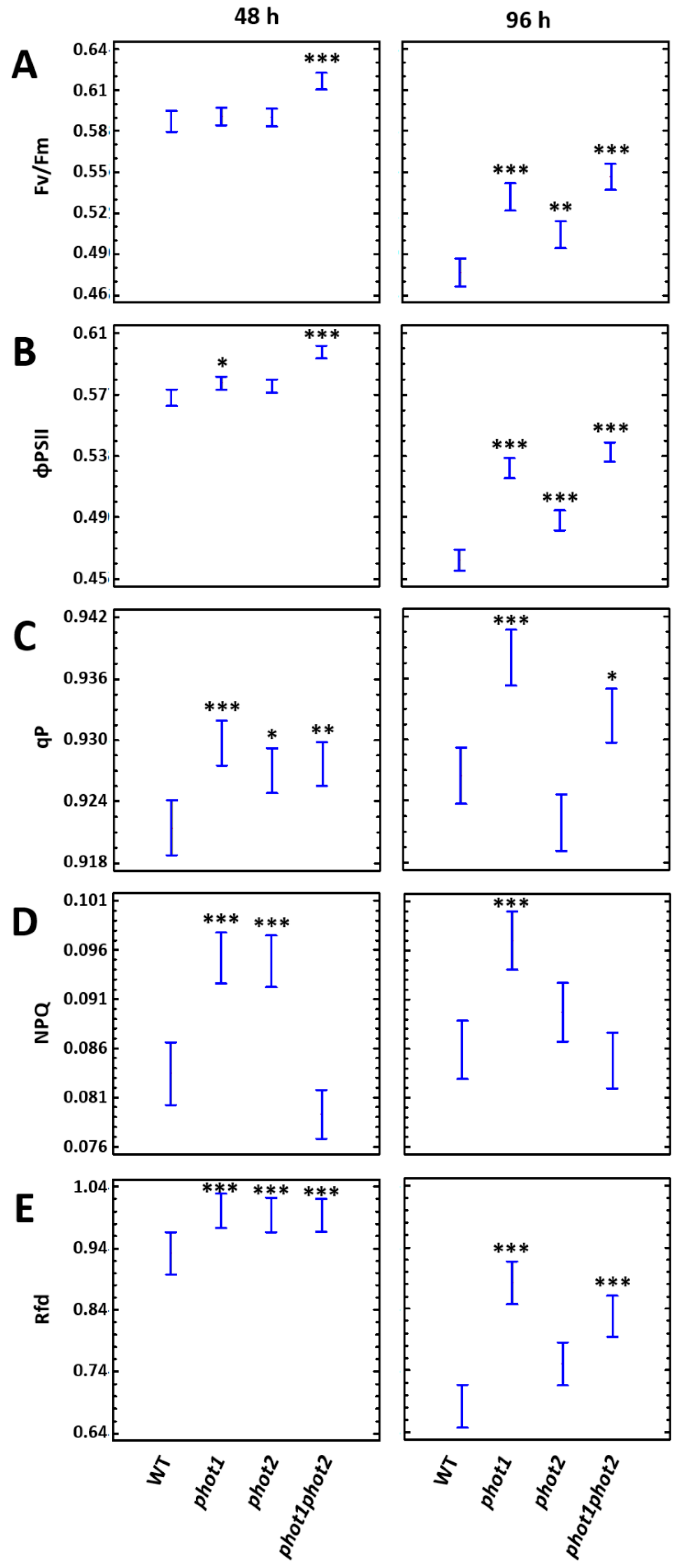
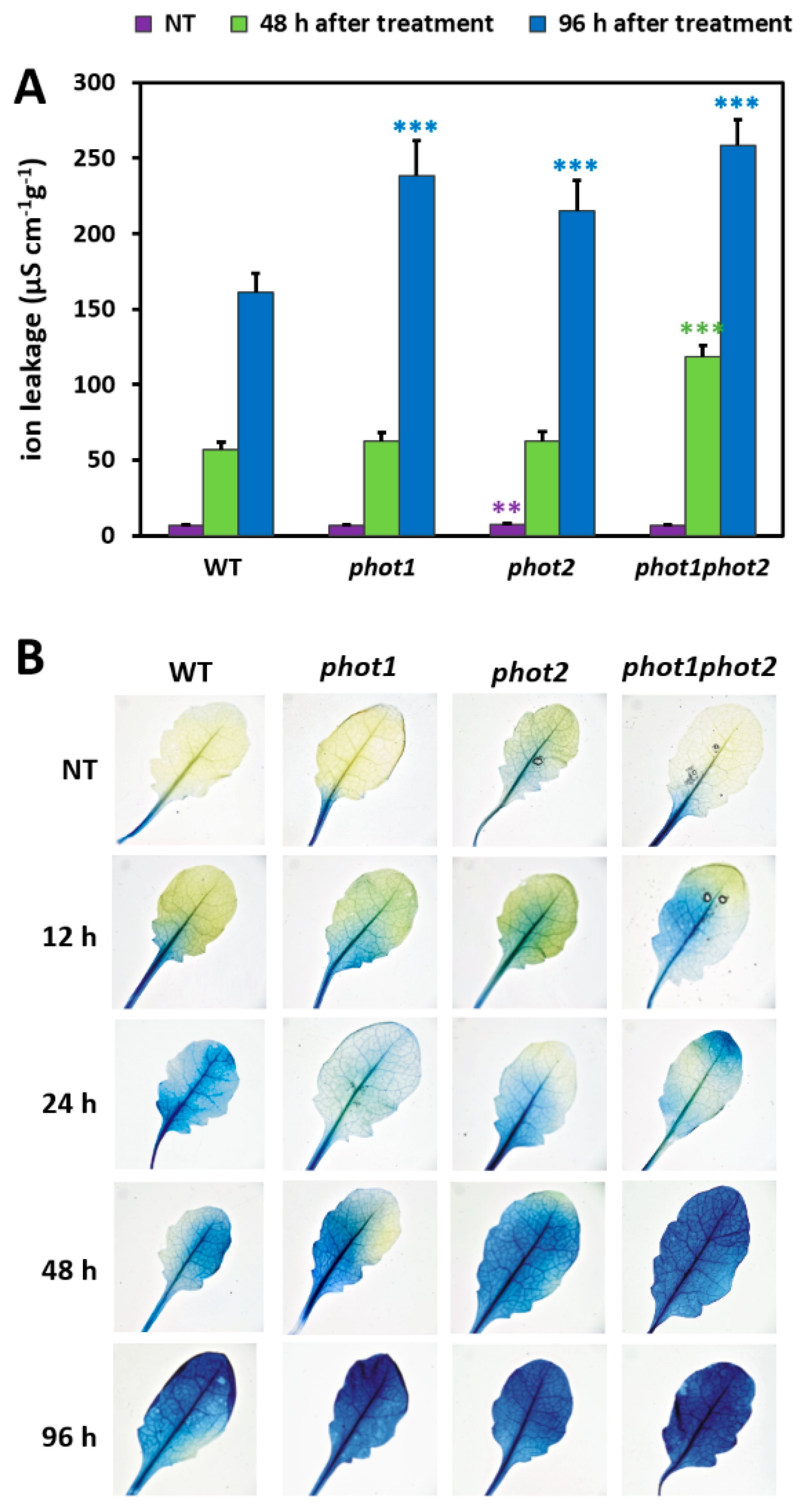
3.4. phot1 and phot2 Influence the Resistance Toward UV-C Damage
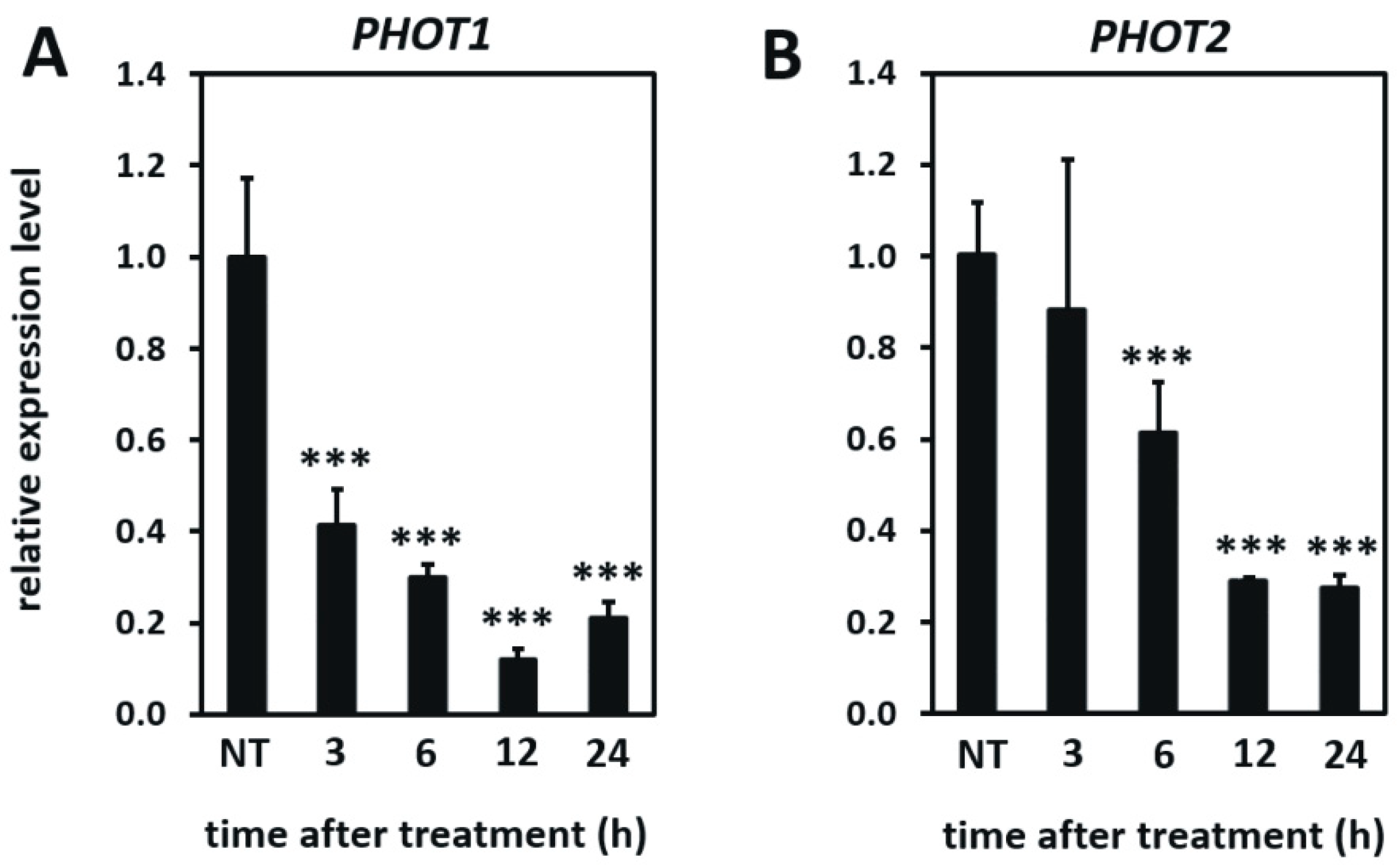
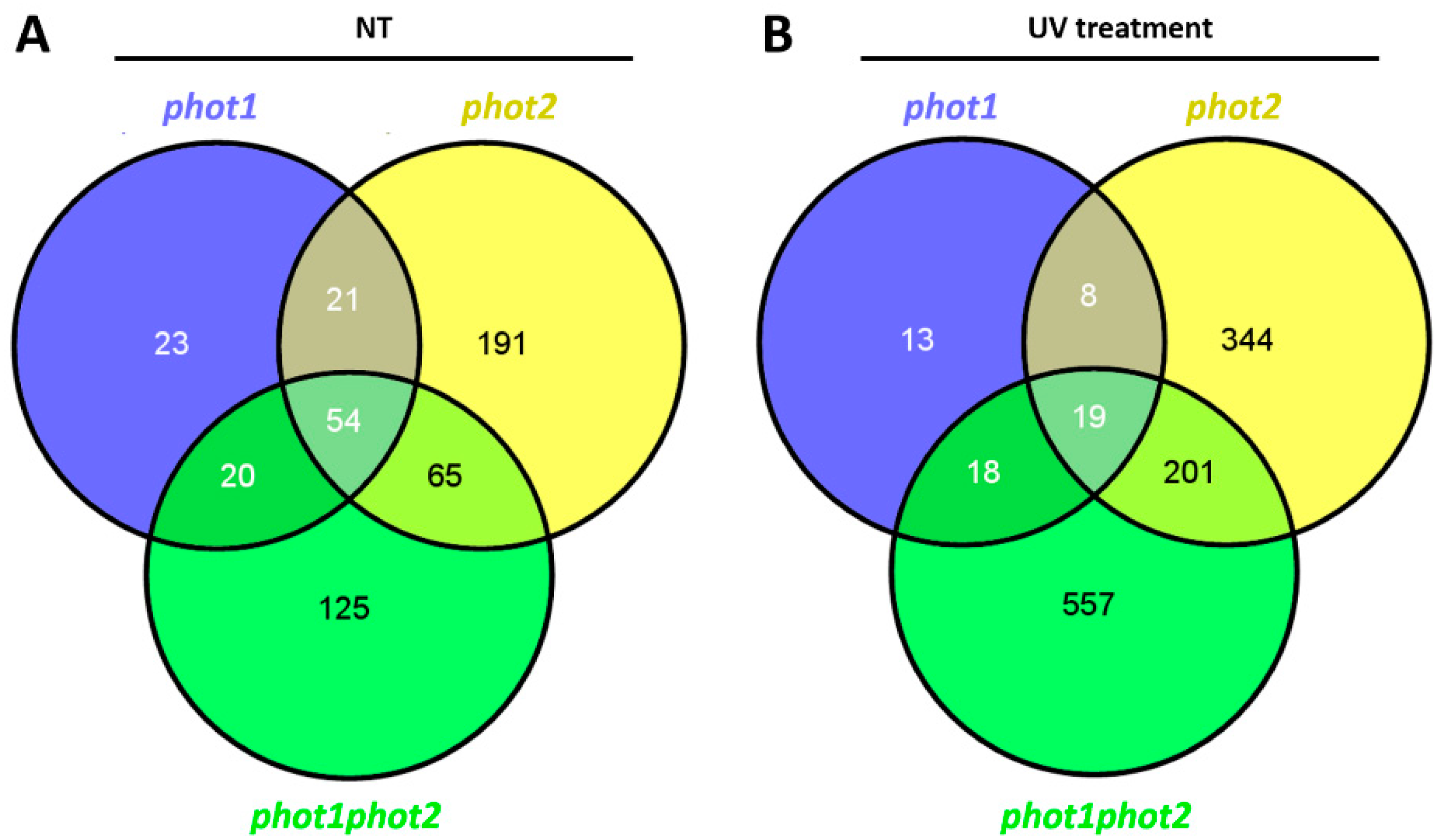
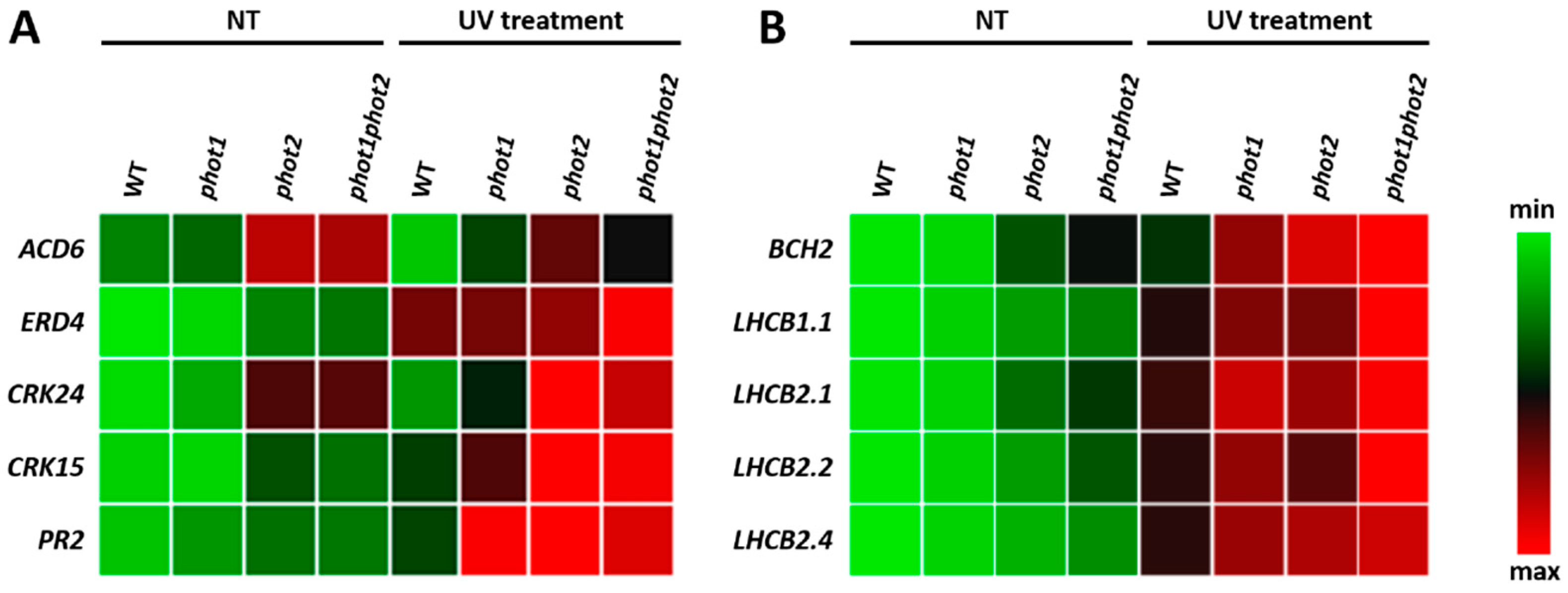
3.5. Differences in the Plant Transcriptome in Photo Mutants in Non-Stress and UV-C Stress Conditions
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ohnishi, N.; Allakhverdiev, S.I.; Takahashi, S.; Higashi, S.; Watanabe, M.; Nishiyama, Y.; Murata, N. Two-Step Mechanism of Photodamage to Photosystem II: Step 1 Occurs at the Oxygen-Evolving Complex and Step 2 Occurs at the Photochemical Reaction Center. Biochemistry 2005, 44, 8494–8499. [Google Scholar] [CrossRef]
- Wituszyńska, W.; Szechyńska-Hebda, M.; Sobczak, M.; Rusaczonek, A.; Kozłowska-Makulska, A.; Witoń, D.; Karpiński, S. Lesion Simulating Disease 1 and Enhanced Disease Susceptibility 1 Differentially Regulate UV-C-Induced Photooxidative Stress Signalling and Programmed Cell Death in Arabidopsis thaliana. Plant Cell Environ. 2015, 38, 315–330. [Google Scholar] [CrossRef] [PubMed]
- Czarnocka, W.; Rusaczonek, A.; Willems, P.; Sujkowska-Rybkowska, M.; Van Breusegem, F.; Karpiński, S. Novel Role of JAC1 in Influencing Photosynthesis, Stomatal Conductance, and Photooxidative Stress Signalling Pathway in Arabidopsis thaliana. Front. Plant Sci. 2020, 11, 1124. [Google Scholar] [CrossRef] [PubMed]
- Karpiński, S.; Szechyńska-Hebda, M.; Wituszyńska, W.; Burdiak, P. Light Acclimation, Retrograde Signalling, Cell Death and Immune Defences in Plants. Plant Cell Environ. 2013, 36, 736–744. [Google Scholar] [CrossRef]
- Czarnocka, W.; Karpiński, S. Friend or Foe? Reactive Oxygen Species Production, Scavenging and Signaling in Plant Response to Environmental Stresses. Free Radic. Biol. Med. 2018, 122, 4–20. [Google Scholar] [CrossRef] [PubMed]
- Demarsy, E.; Goldschmidt-Clermont, M.; Ulm, R. Coping with “Dark Sides of the Sun” through Photoreceptor Signaling. Trends Plant Sci. 2018, 23, 260–271. [Google Scholar] [CrossRef]
- Czarnocka, W.; Fichman, Y.; Bernacki, M.; Różańska, E.; Sańko-Sawczenko, I.; Mittler, R.; Karpiński, S. FMO1 Is Involved in Excess Light Stress-Induced Signal Transduction and Cell Death Signaling. Cells 2020, 9, 2163. [Google Scholar] [CrossRef]
- Gilroy, S.; Białasek, M.; Suzuki, N.; Górecka, M.; Devireddy, A.R.; Karpiński, S.; Mittler, R. ROS, Calcium, and Electric Signals: Key Mediators of Rapid Systemic Signaling in Plants. Plant Physiol. 2016, 171, 1606–1615. [Google Scholar] [CrossRef]
- Wituszyńska, W.; Karpiński, S. Programmed Cell Death as a Response to High Light, UV and Drought Stress in Plants. Abiotic Stress Plant Responses Appl. Agric. 2013. [Google Scholar] [CrossRef] [Green Version]
- Pfündel, E.E.; Latouche, G.; Meister, A.; Cerovic, Z.G. Linking Chloroplast Relocation to Different Responses of Photosynthesis to Blue and Red Radiation in Low and High Light-Acclimated Leaves of Arabidopsis thaliana (L.). Photosynth. Res. 2018, 137, 105–128. [Google Scholar] [CrossRef]
- Suetsugu, N.; Wada, M. Chloroplast Photorelocation Movement Mediated by Phototropin Family Proteins in Green Plants. Biol. Chem. 2007, 388, 927–935. [Google Scholar] [CrossRef] [PubMed]
- Okajima, K. Molecular Mechanism of Phototropin Light Signaling. J. Plant Res. 2016, 129, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Majumdar, A.; Kar, R.K. Chloroplast Avoidance Movement: A Novel Paradigm of ROS Signalling. Photosynth. Res. 2020, 144, 109–121. [Google Scholar] [CrossRef] [PubMed]
- Sakai, T.; Kagawa, T.; Kasahara, M.; Swartz, T.E.; Christie, J.M.; Briggs, W.R.; Wada, M.; Okada, K. Arabidopsis Nph1 and Npl1: Blue Light Receptors That Mediate Both Phototropism and Chloroplast Relocation. Proc. Natl. Acad. Sci. USA 2001, 98, 6969–6974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kinoshita, T.; Doi, M.; Suetsugu, N.; Kagawa, T.; Wada, M.; Shimazaki, K. Phot1 and Phot2 Mediate Blue Light Regulation of Stomatal Opening. Nature 2001, 414, 656–660. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Robson, T.M.; Casal, J.J.; Shapiguzov, A.; Aphalo, P.J. Contributions of Cryptochromes and Phototropins to Stomatal Opening through the Day. Funct. Plant Biol. 2020, 47, 226–238. [Google Scholar] [CrossRef] [Green Version]
- Sakamoto, K.; Briggs, W.R. Cellular and Subcellular Localization of Phototropin 1. Plant Cell 2002, 14, 1723–1735. [Google Scholar] [CrossRef] [Green Version]
- Inoue, S.; Kinoshita, T.; Takemiya, A.; Doi, M.; Shimazaki, K. Leaf Positioning of Arabidopsis in Response to Blue Light. Mol. Plant 2008, 1, 15–26. [Google Scholar] [CrossRef]
- Sakai, T.; Wada, T.; Ishiguro, S.; Okada, K. RPT2. A Signal Transducer of the Phototropic Response in Arabidopsis. Plant Cell 2000, 12, 225–236. [Google Scholar] [CrossRef] [Green Version]
- Folta, K.M.; Lieg, E.J.; Durham, T.; Spalding, E.P. Primary Inhibition of Hypocotyl Growth and Phototropism Depend Differently on Phototropin-Mediated Increases in Cytoplasmic Calcium Induced by Blue Light. Plant Physiol. 2003, 133, 1464–1470. [Google Scholar] [CrossRef] [Green Version]
- Moni, A.; Lee, A.-Y.; Briggs, W.R.; Han, I.-S. The Blue Light Receptor Phototropin 1 Suppresses Lateral Root Growth by Controlling Cell Elongation. Plant Biol. Stuttg. Ger. 2015, 17, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Kagawa, T.; Sakai, T.; Suetsugu, N.; Oikawa, K.; Ishiguro, S.; Kato, T.; Tabata, S.; Okada, K.; Wada, M. Arabidopsis NPL1: A Phototropin Homolog Controlling the Chloroplast High-Light Avoidance Response. Science 2001, 291, 2138–2141. [Google Scholar] [CrossRef] [PubMed]
- Iwabuchi, K.; Sakai, T.; Takagi, S. Blue Light-Dependent Nuclear Positioning in Arabidopsis thaliana Leaf Cells. Plant Cell Physiol. 2007, 48, 1291–1298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kasahara, M.; Swartz, T.E.; Olney, M.A.; Onodera, A.; Mochizuki, N.; Fukuzawa, H.; Asamizu, E.; Tabata, S.; Kanegae, H.; Takano, M.; et al. Photochemical Properties of the Flavin Mononucleotide-Binding Domains of the Phototropins from Arabidopsis, Rice, and Chlamydomonas reinhardtii. Plant Physiol. 2002, 129, 762–773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanhaelewyn, L.; Schumacher, P.; Poelman, D.; Fankhauser, C.; Van Der Straeten, D.; Vandenbussche, F. Repressor of Ultraviolet-B Photomorphogenesis Function Allows Efficient Phototropin Mediated Ultraviolet-B Phototropism in Etiolated Seedlings. Plant Sci. Int. J. Exp. Plant Biol. 2016, 252, 215–221. [Google Scholar] [CrossRef] [Green Version]
- Kaiserli, E.; Sullivan, S.; Jones, M.A.; Feeney, K.A.; Christie, J.M. Domain Swapping to Assess the Mechanistic Basis of Arabidopsis Phototropin 1 Receptor Kinase Activation and Endocytosis by Blue Light. Plant Cell 2009, 21, 3226–3244. [Google Scholar] [CrossRef] [Green Version]
- Freddolino, P.L.; Gardner, K.H.; Schulten, K. Signaling Mechanisms of LOV Domains: New Insights from Molecular Dynamics Studies. Photochem. Photobiol. Sci. Off. J. Eur. Photochem. Assoc. Eur. Soc. Photobiol. 2013, 12, 1158–1170. [Google Scholar] [CrossRef] [Green Version]
- Tokutomi, S.; Matsuoka, D.; Zikihara, K. Molecular Structure and Regulation of Phototropin Kinase by Blue Light. Biochim. Biophys. Acta 2008, 1784, 133–142. [Google Scholar] [CrossRef]
- Nakasako, M.; Iwata, T.; Matsuoka, D.; Tokutomi, S. Light-Induced Structural Changes of LOV Domain-Containing Polypeptides from Arabidopsis Phototropin 1 and 2 Studied by Small-Angle X-ray Scattering. Biochemistry 2004, 43, 14881–14890. [Google Scholar] [CrossRef]
- Inoue, S.-I.; Kinoshita, T.; Matsumoto, M.; Nakayama, K.I.; Doi, M.; Shimazaki, K.-I. Blue Light-Induced Autophosphorylation of Phototropin Is a Primary Step for Signaling. Proc. Natl. Acad. Sci. USA 2008, 105, 5626–5631. [Google Scholar] [CrossRef] [Green Version]
- Inoue, S.; Matsushita, T.; Tomokiyo, Y.; Matsumoto, M.; Nakayama, K.I.; Kinoshita, T.; Shimazaki, K. Functional Analyses of the Activation Loop of Phototropin2 in Arabidopsis. Plant Physiol. 2011, 156, 117–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ueno, K.; Kinoshita, T.; Inoue, S.-I.; Emi, T.; Shimazaki, K.-I. Biochemical Characterization of Plasma Membrane H+-ATPase Activation in Guard Cell Protoplasts of Arabidopsis thaliana in Response to Blue Light. Plant Cell Physiol. 2005, 46, 955–963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kozuka, T.; Kong, S.-G.; Doi, M.; Shimazaki, K.; Nagatani, A. Tissue-Autonomous Promotion of Palisade Cell Development by Phototropin 2 in Arabidopsis. Plant Cell 2011, 23, 3684–3695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Königer, M.; Jessen, B.; Yang, R.; Sittler, D.; Harris, G.C. Light, Genotype, and Abscisic Acid Affect Chloroplast Positioning in Guard Cells of Arabidopsis thaliana Leaves in Distinct Ways. Photosynth. Res. 2010, 105, 213–227. [Google Scholar] [CrossRef]
- Kong, S.-G.; Suzuki, T.; Tamura, K.; Mochizuki, N.; Hara-Nishimura, I.; Nagatani, A. Blue Light-Induced Association of Phototropin 2 with the Golgi Apparatus. Plant J. Cell Mol. Biol. 2006, 45, 994–1005. [Google Scholar] [CrossRef]
- Kong, S.-G.; Suetsugu, N.; Kikuchi, S.; Nakai, M.; Nagatani, A.; Wada, M. Both Phototropin 1 and 2 Localize on the Chloroplast Outer Membrane with Distinct Localization Activity. Plant Cell Physiol. 2013, 54, 80–92. [Google Scholar] [CrossRef] [Green Version]
- Wan, Y.-L.; Eisinger, W.; Ehrhardt, D.; Kubitscheck, U.; Baluska, F.; Briggs, W. The Subcellular Localization and Blue-Light-Induced Movement of Phototropin 1-GFP in Etiolated Seedlings of Arabidopsis thaliana. Mol. Plant 2008, 1, 103–117. [Google Scholar] [CrossRef]
- Roberts, D.; Pedmale, U.V.; Morrow, J.; Sachdev, S.; Lechner, E.; Tang, X.; Zheng, N.; Hannink, M.; Genschik, P.; Liscum, E. Modulation of Phototropic Responsiveness in Arabidopsis through Ubiquitination of Phototropin 1 by the CUL3-Ring E3 Ubiquitin Ligase CRL3(NPH3). Plant Cell 2011, 23, 3627–3640. [Google Scholar] [CrossRef] [Green Version]
- Aggarwal, C.; Banaś, A.K.; Kasprowicz-Maluśki, A.; Borghetti, C.; Labuz, J.; Dobrucki, J.; Gabryś, H. Blue-Light-Activated Phototropin2 Trafficking from the Cytoplasm to Golgi/Post-Golgi Vesicles. J. Exp. Bot. 2014, 65, 3263–3276. [Google Scholar] [CrossRef] [Green Version]
- Kadota, A.; Yamada, N.; Suetsugu, N.; Hirose, M.; Saito, C.; Shoda, K.; Ichikawa, S.; Kagawa, T.; Nakano, A.; Wada, M. Short Actin-Based Mechanism for Light-Directed Chloroplast Movement in Arabidopsis. Proc. Natl. Acad. Sci. USA 2009, 106, 13106–13111. [Google Scholar] [CrossRef] [Green Version]
- Gotoh, E.; Suetsugu, N.; Yamori, W.; Ishishita, K.; Kiyabu, R.; Fukuda, M.; Higa, T.; Shirouchi, B.; Wada, M. Chloroplast Accumulation Response Enhances Leaf Photosynthesis and Plant Biomass Production. Plant Physiol. 2018, 178, 1358–1369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kasahara, M.; Kagawa, T.; Oikawa, K.; Suetsugu, N.; Miyao, M.; Wada, M. Chloroplast Avoidance Movement Reduces Photodamage in Plants. Nature 2002, 420, 829–832. [Google Scholar] [CrossRef] [PubMed]
- Ichikawa, S.; Yamada, N.; Suetsugu, N.; Wada, M.; Kadota, A. Red Light, Phot1 and JAC1 Modulate Phot2-Dependent Reorganization of Chloroplast Actin Filaments and Chloroplast Avoidance Movement. Plant Cell Physiol. 2011, 52, 1422–1432. [Google Scholar] [CrossRef] [PubMed]
- Wada, M. Chloroplast and Nuclear Photorelocation Movements. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2016, 92, 387–411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wituszynska, W.; Karpiński, S. Determination of Water Use Efficiency for Arabidopsis thaliana. Bio-Protoc 2014, 4, e1041. [Google Scholar] [CrossRef]
- Wituszyńska, W.; Gałązka, K.; Rusaczonek, A.; Vanderauwera, S.; Van Breusegem, F.; Karpiński, S. Multivariable Environmental Conditions Promote Photosynthetic Adaptation Potential in Arabidopsis thaliana. J. Plant Physiol. 2013, 170, 548–559. [Google Scholar] [CrossRef] [PubMed]
- Baker, N.R. Chlorophyll Fluorescence: A Probe of Photosynthesis in Vivo. Annu. Rev. Plant Biol. 2008, 59, 89–113. [Google Scholar] [CrossRef] [Green Version]
- Rusaczonek, A.; Czarnocka, W.; Kacprzak, S.; Witoń, D.; Ślesak, I.; Szechyńska-Hebda, M.; Gawroński, P.; Karpiński, S. Role of Phytochromes A and B in the Regulation of Cell Death and Acclimatory Responses to UV Stress in Arabidopsis thaliana. J. Exp. Bot. 2015, 66, 6679–6695. [Google Scholar] [CrossRef] [Green Version]
- Orozco-Cardenas, M.; Ryan, C.A. Hydrogen Peroxide Is Generated Systemically in Plant Leaves by Wounding and Systemin via the Octadecanoid Pathway. Proc. Natl. Acad. Sci. USA 1999, 96, 6553–6557. [Google Scholar] [CrossRef] [Green Version]
- Mergemann, H.; Sauter, M. Ethylene Induces Epidermal Cell Death at the Site of Adventitious Root Emergence in Rice. Plant Physiol. 2000, 124, 609–614. [Google Scholar] [CrossRef] [Green Version]
- Hruz, T.; Laule, O.; Szabo, G.; Wessendorp, F.; Bleuler, S.; Oertle, L.; Widmayer, P.; Gruissem, W.; Zimmermann, P. Genevestigator v3: A Reference Expression Database for the Meta-Analysis of Transcriptomes. Adv. Bioinforma. 2008, 2008, 420747. [Google Scholar] [CrossRef] [PubMed]
- Ramakers, C.; Ruijter, J.M.; Deprez, R.H.L.; Moorman, A.F.M. Assumption-Free Analysis of Quantitative Real-Time Polymerase Chain Reaction (PCR) Data. Neurosci. Lett. 2003, 339, 62–66. [Google Scholar] [CrossRef]
- Pfaffl, M.W.; Horgan, G.W.; Dempfle, L. Relative Expression Software Tool (REST) for Group-Wise Comparison and Statistical Analysis of Relative Expression Results in Real-Time PCR. Nucleic Acids Res. 2002, 30, e36. [Google Scholar] [CrossRef] [PubMed]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast Universal RNA-Seq Aligner. Bioinforma. Oxf. Engl. 2013, 29, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.-Y.; Krishnakumar, V.; Chan, A.P.; Thibaud-Nissen, F.; Schobel, S.; Town, C.D. Araport11: A Complete Reannotation of the Arabidopsis thaliana Reference Genome. Plant J. Cell Mol. Biol. 2017, 89, 789–804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, Y.; Smyth, G.K.; Shi, W. FeatureCounts: An Efficient General Purpose Program for Assigning Sequence Reads to Genomic Features. Bioinforma. Oxf. Engl. 2014, 30, 923–930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. EdgeR: A Bioconductor Package for Differential Expression Analysis of Digital Gene Expression Data. Bioinform. Oxf. Engl. 2010, 26, 139–140. [Google Scholar] [CrossRef] [Green Version]
- Robinson, M.D.; Oshlack, A. A Scaling Normalization Method for Differential Expression Analysis of RNA-Seq Data. Genome Biol. 2010, 11, R25. [Google Scholar] [CrossRef] [Green Version]
- McCarthy, D.J.; Chen, Y.; Smyth, G.K. Differential Expression Analysis of Multifactor RNA-Seq Experiments with Respect to Biological Variation. Nucleic Acids Res. 2012, 40, 4288–4297. [Google Scholar] [CrossRef] [Green Version]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Krishnakumar, V.; Hanlon, M.R.; Contrino, S.; Ferlanti, E.S.; Karamycheva, S.; Kim, M.; Rosen, B.D.; Cheng, C.-Y.; Moreira, W.; Mock, S.A.; et al. Araport: The Arabidopsis Information Portal. Nucleic Acids Res. 2015, 43, D1003–D1009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thimm, O.; Bläsing, O.; Gibon, Y.; Nagel, A.; Meyer, S.; Krüger, P.; Selbig, J.; Müller, L.A.; Rhee, S.Y.; Stitt, M. Mapman: A User-Driven Tool to Display Genomics Data Sets onto Diagrams of Metabolic Pathways and Other Biological Processes. Plant J. Cell Mol. Biol. 2004, 37, 914–939. [Google Scholar] [CrossRef]
- Górecka, M.; Lewandowska, M.; Dąbrowska-Bronk, J.; Białasek, M.; Barczak-Brzyżek, A.; Kulasek, M.; Mielecki, J.; Kozłowska-Makulska, A.; Gawroński, P.; Karpiński, S. Photosystem II 22kDa Protein Level—A Prerequisite for Excess Light-Inducible Memory, Cross-Tolerance to UV-C and Regulation of Electrical Signalling. Plant Cell Environ. 2020, 43, 649–661. [Google Scholar] [CrossRef]
- Haitz, M.; Lichtenthaler, H.K. The measurement of Rfd-values as plant vitality indices with the portable field chlorophyll fluorometer and the pam-fluorometer. In Applications of Chlorophyll Fluorescence in Photosynthesis Research, Stress Physiology, Hydrobiology and Remote Sensing: An Introduction to the Various Fields of Applications of the In Vivo Chlorophyll Fluorescence also Including the Proceedings of the First International Chlorophyll Fluorescence Symposium Held in the Physikzentrum, Bad Honnef, F.R.G., 6–8 June 1998; Lichtenthaler, H.K., Ed.; Springer: Dordrecht, The Netherlands, 1988; pp. 249–254. ISBN 978-94-009-2823-7. [Google Scholar]
- Pillitteri, L.J.; Dong, J. Stomatal Development in Arabidopsis. Arab. Book Am. Soc. Plant Biol. 2013, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Shrestha, J.; Tateda, C.; Greenberg, J.T. Salicylic Acid Signaling Controls the Maturation and Localization of the Arabidopsis Defense Protein Accelerated Cell Death6. Mol. Plant 2014, 7, 1365–1383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wrzaczek, M.; Brosché, M.; Salojärvi, J.; Kangasjärvi, S.; Idänheimo, N.; Mersmann, S.; Robatzek, S.; Karpiński, S.; Karpińska, B.; Kangasjärvi, J. Transcriptional Regulation of the CRK/DUF26 Group of Receptor-like Protein Kinases by Ozone and Plant Hormones in Arabidopsis. BMC Plant Biol. 2010, 10, 95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bourdais, G.; Burdiak, P.; Gauthier, A.; Nitsch, L.; Salojärvi, J.; Rayapuram, C.; Idänheimo, N.; Hunter, K.; Kimura, S.; Merilo, E.; et al. Large-Scale Phenomics Identifies Primary and Fine-Tuning Roles for CRKs in Responses Related to Oxidative Stress. PLoS Genet. 2015, 11, e1005373. [Google Scholar] [CrossRef] [Green Version]
- van Loon, L.C.; Rep, M.; Pieterse, C.M.J. Significance of Inducible Defense-Related Proteins in Infected Plants. Annu. Rev. Phytopathol. 2006, 44, 135–162. [Google Scholar] [CrossRef] [Green Version]
- Kandasamy, M.K.; Deal, R.B.; McKinney, E.C.; Meagher, R.B. Plant Actin-Related Proteins. Trends Plant Sci. 2004, 9, 196–202. [Google Scholar] [CrossRef]
- Sun, Z.; Qi, X.; Wang, Z.; Li, P.; Wu, C.; Zhang, H.; Zhao, Y. Overexpression of TsGOLS2, a Galactinol Synthase, in Arabidopsis thaliana Enhances Tolerance to High Salinity and Osmotic Stresses. Plant Physiol. Biochem. 2013, 69, 82–89. [Google Scholar] [CrossRef]
- Song, C.; Chung, W.S.; Lim, C.O. Overexpression of Heat Shock Factor Gene HsfA3 Increases Galactinol Levels and Oxidative Stress Tolerance in Arabidopsis. Mol. Cells 2016, 39, 477–483. [Google Scholar] [CrossRef]
- Selvaraj, M.G.; Ishizaki, T.; Valencia, M.; Ogawa, S.; Dedicova, B.; Ogata, T.; Yoshiwara, K.; Maruyama, K.; Kusano, M.; Saito, K.; et al. Overexpression of an Arabidopsis thaliana Galactinol Synthase Gene Improves Drought Tolerance in Transgenic Rice and Increased Grain Yield in the Field. Plant Biotechnol. J. 2017, 15, 1465–1477. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.; Gan, S. AtNAP, a NAC Family Transcription Factor, Has an Important Role in Leaf Senescence. Plant J. Cell Mol. Biol. 2006, 46, 601–612. [Google Scholar] [CrossRef]
- Seok, H.-Y.; Woo, D.-H.; Nguyen, L.V.; Tran, H.T.; Tarte, V.N.; Mehdi, S.M.M.; Lee, S.-Y.; Moon, Y.-H. Arabidopsis AtNAP Functions as a Negative Regulator via Repression of AREB1 in Salt Stress Response. Planta 2017, 245, 329–341. [Google Scholar] [CrossRef] [PubMed]
- Lei, W.; Li, Y.; Yao, X.; Qiao, K.; Wei, L.; Liu, B.; Zhang, D.; Lin, H. NAP Is Involved in GA-Mediated Chlorophyll Degradation and Leaf Senescence by Interacting with DELLAs in Arabidopsis. Plant Cell Rep. 2020, 39, 75–87. [Google Scholar] [CrossRef]
- Kim, S.J.; Kim, W.T. Suppression of Arabidopsis RING E3 Ubiquitin Ligase AtATL78 Increases Tolerance to Cold Stress and Decreases Tolerance to Drought Stress. FEBS Lett. 2013, 587, 2584–2590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suh, J.Y.; Kim, S.J.; Oh, T.R.; Cho, S.K.; Yang, S.W.; Kim, W.T. Arabidopsis Tóxicos En Levadura 78 (AtATL78) Mediates ABA-Dependent ROS Signaling in Response to Drought Stress. Biochem. Biophys. Res. Commun. 2016, 469, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, P.; Nöthen, J.; von Braun, S.S.; Bohnsack, M.T.; Mirus, O.; Schleiff, E. Transitions of Gene Expression Induced by Short-Term Blue Light. Plant Biol. Stuttg. Ger. 2011, 13, 349–361. [Google Scholar] [CrossRef] [PubMed]
- Zybailov, B.; Rutschow, H.; Friso, G.; Rudella, A.; Emanuelsson, O.; Sun, Q.; van Wijk, K.J. Sorting Signals, N-Terminal Modifications and Abundance of the Chloroplast Proteome. PLoS ONE 2008, 3, e1994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Gao, J.; Yao, L.; Ren, G.; Zhu, X.; Gao, S.; Qiu, K.; Zhou, X.; Kuai, B. The Role of ANAC072 in the Regulation of Chlorophyll Degradation during Age- and Dark-Induced Leaf Senescence. Plant Cell Rep. 2016, 35, 1729–1741. [Google Scholar] [CrossRef] [PubMed]
- Kamranfar, I.; Xue, G.-P.; Tohge, T.; Sedaghatmehr, M.; Fernie, A.R.; Balazadeh, S.; Mueller-Roeber, B. Transcription Factor RD26 Is a Key Regulator of Metabolic Reprogramming during Dark-Induced Senescence. New Phytol. 2018, 218, 1543–1557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benschop, J.J.; Mohammed, S.; O’Flaherty, M.; Heck, A.J.R.; Slijper, M.; Menke, F.L.H. Quantitative Phosphoproteomics of Early Elicitor Signaling in Arabidopsis. Mol. Cell. Proteomics 2007, 6, 1198–1214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luhua, S.; Ciftci-Yilmaz, S.; Harper, J.; Cushman, J.; Mittler, R. Enhanced Tolerance to Oxidative Stress in Transgenic Arabidopsis Plants Expressing Proteins of Unknown Function. Plant Physiol. 2008, 148, 280–292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bechtold, U.; Richard, O.; Zamboni, A.; Gapper, C.; Geisler, M.; Pogson, B.; Karpinski, S.; Mullineaux, P.M. Impact of Chloroplastic- and Extracellular-Sourced ROS on High Light-Responsive Gene Expression in Arabidopsis. J. Exp. Bot. 2008, 59, 121–133. [Google Scholar] [CrossRef]
- Takemiya, A.; Inoue, S.-I.; Doi, M.; Kinoshita, T.; Shimazaki, K.-I. Phototropins Promote Plant Growth in Response to Blue Light in Low Light Environments. Plant Cell 2005, 17, 1120–1127. [Google Scholar] [CrossRef] [Green Version]
- López-Juez, E.; Bowyer, J.R.; Sakai, T. Distinct Leaf Developmental and Gene Expression Responses to Light Quantity Depend on Blue-Photoreceptor or Plastid-Derived Signals, and Can Occur in the Absence of Phototropins. Planta 2007, 227, 113–123. [Google Scholar] [CrossRef]
- Sullivan, S.; Thomson, C.E.; Lamont, D.J.; Jones, M.A.; Christie, J.M. In Vivo Phosphorylation Site Mapping and Functional Characterization of Arabidopsis Phototropin 1. Mol. Plant 2008, 1, 178–194. [Google Scholar] [CrossRef]
- Harada, A.; Takemiya, A.; Inoue, S.; Sakai, T.; Shimazaki, K. Role of RPT2 in Leaf Positioning and Flattening and a Possible Inhibition of Phot2 Signaling by Phot1. Plant Cell Physiol. 2013, 54, 36–47. [Google Scholar] [CrossRef]
- Hart, J.E.; Sullivan, S.; Hermanowicz, P.; Petersen, J.; Diaz-Ramos, L.A.; Hoey, D.J.; Łabuz, J.; Christie, J.M. Engineering the Phototropin Photocycle Improves Photoreceptor Performance and Plant Biomass Production. Proc. Natl. Acad. Sci. USA 2019, 116, 12550–12557. [Google Scholar] [CrossRef] [Green Version]
- Shimazaki, K.; Doi, M.; Assmann, S.M.; Kinoshita, T. Light Regulation of Stomatal Movement. Annu. Rev. Plant Biol. 2007, 58, 219–247. [Google Scholar] [CrossRef] [Green Version]
- Takemiya, A.; Doi, A.; Yoshida, S.; Okajima, K.; Tokutomi, S.; Shimazaki, K.-I. Reconstitution of an Initial Step of Phototropin Signaling in Stomatal Guard Cells. Plant Cell Physiol. 2016, 57, 152–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boccalandro, H.E.; Giordano, C.V.; Ploschuk, E.L.; Piccoli, P.N.; Bottini, R.; Casal, J.J. Phototropins but Not Cryptochromes Mediate the Blue Light-Specific Promotion of Stomatal Conductance, While Both Enhance Photosynthesis and Transpiration under Full Sunlight. Plant Physiol. 2012, 158, 1475–1484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kostaki, K.-I.; Coupel-Ledru, A.; Bonnell, V.C.; Gustavsson, M.; Sun, P.; McLaughlin, F.J.; Fraser, D.P.; McLachlan, D.H.; Hetherington, A.M.; Dodd, A.N.; et al. Guard Cells Integrate Light and Temperature Signals to Control Stomatal Aperture. Plant Physiol. 2020, 182, 1404–1419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tuberosa, R. Molecular approaches to unravel the genetic basis of water use efficiency. In Water Use Efficiency in Plant Biology; Bacon, M.A., Ed.; Blackwell Publishing: Oxford, UK, 2004. [Google Scholar]
- Sztatelman, O.; Waloszek, A.; Banaś, A.K.; Gabryś, H. Photoprotective Function of Chloroplast Avoidance Movement: In Vivo Chlorophyll Fluorescence Study. J. Plant Physiol. 2010, 167, 709–716. [Google Scholar] [CrossRef] [PubMed]
- Davis, P.A.; Hangarter, R.P. Chloroplast Movement Provides Photoprotection to Plants by Redistributing PSII Damage within Leaves. Photosynth. Res. 2012, 112, 153–161. [Google Scholar] [CrossRef]
- Weston, E.; Thorogood, K.; Vinti, G.; López-Juez, E. Light Quantity Controls Leaf-Cell and Chloroplast Development in Arabidopsis thaliana Wild Type and Blue-Light-Perception Mutants. Planta 2000, 211, 807–815. [Google Scholar] [CrossRef]
- Terashima, I.; Hikosaka, K. Comparative Ecophysiology of Leaf and Canopy Photosynthesis. Plant Cell Environ. 1995, 18, 1111–1128. [Google Scholar] [CrossRef]
- Kitajima, K.; Hogan, K.P. Increases of Chlorophyll a/b Ratios during Acclimation of Tropical Woody Seedlings to Nitrogen Limitation and High Light. Plant Cell Environ. 2003, 26, 857–865. [Google Scholar] [CrossRef]
- Howard, M.M.; Bae, A.; Königer, M. The Importance of Chloroplast Movement, Nonphotochemical Quenching, and Electron Transport Rates in Light Acclimation and Tolerance to High Light in Arabidopsis thaliana. Am. J. Bot. 2019, 106, 1444–1453. [Google Scholar] [CrossRef]
- Petrov, V.; Hille, J.; Mueller-Roeber, B.; Gechev, T.S. ROS-Mediated Abiotic Stress-Induced Programmed Cell Death in Plants. Front. Plant Sci. 2015, 6, 69. [Google Scholar] [CrossRef] [Green Version]
- Foyer, C.H. Reactive Oxygen Species, Oxidative Signaling and the Regulation of Photosynthesis. Environ. Exp. Bot. 2018, 154, 134–142. [Google Scholar] [CrossRef] [PubMed]
- Wituszynska, W.; Slesak, I.; Vanderauwera, S.; Szechynska-Hebda, M.; Kornas, A.; Van Der Kelen, K.; Mühlenbock, P.; Karpinska, B.; Mackowski, S.; Van Breusegem, F.; et al. Lesion Simulating Disease1, Enhanced Disease Susceptibility1, and Phytoalexin Deficient4 Conditionally Regulate Cellular Signaling Homeostasis, Photosynthesis, Water Use Efficiency, and Seed Yield in Arabidopsis. Plant Physiol. 2013, 161, 1795–1805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Czarnocka, W.; Van Der Kelen, K.; Willems, P.; Szechyńska-Hebda, M.; Shahnejat-Bushehri, S.; Balazadeh, S.; Rusaczonek, A.; Mueller-Roeber, B.; Van Breusegem, F.; Karpiński, S. The Dual Role of Lesion Simulating Disease 1 as a Condition-Dependent Scaffold Protein and Transcription Regulator. Plant Cell Environ. 2017, 40, 2644–2662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Łabuz, J.; Sztatelman, O.; Banaś, A.K.; Gabryś, H. The Expression of Phototropins in Arabidopsis Leaves: Developmental and Light Regulation. J. Exp. Bot. 2012, 63, 1763–1771. [Google Scholar] [CrossRef] [Green Version]
- Galen, C.; Rabenold, J.J.; Liscum, E. Functional Ecology of a Blue Light Photoreceptor: Effects of Phototropin-1 on Root Growth Enhance Drought Tolerance in Arabidopsis thaliana. New Phytol. 2007, 173, 91–99. [Google Scholar] [CrossRef]
- Königer, M.; Delamaide, J.A.; Marlow, E.D.; Harris, G.C. Arabidopsis thaliana Leaves with Altered Chloroplast Numbers and Chloroplast Movement Exhibit Impaired Adjustments to Both Low and High Light. J. Exp. Bot. 2008, 59, 2285–2297. [Google Scholar] [CrossRef]
- Preuten, T.; Blackwood, L.; Christie, J.M.; Fankhauser, C. Lipid Anchoring of Arabidopsis Phototropin 1 to Assess the Functional Significance of Receptor Internalization: Should I Stay or Should I Go? New Phytol. 2015, 206, 1038–1050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fankhauser, C.; Christie, J.M. Plant Phototropic Growth. Curr. Biol. 2015, 25, R384–R389. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| WT | phot1 | phot2 | phot1phot2 | |
|---|---|---|---|---|
| Total Chlorophyll | 28680 ± 2440 | 29378 ± 2175 | 25300 ± 1876 ** | 26628 ± 2298 * |
| Chlorophyll a | 20739 ± 1752 | 21496 ± 1599 | 18469 ± 1382 ** | 19626 ± 1693 * |
| Chlorophyll b | 7941 ± 688 | 7882 ± 583 | 6830 ± 508 *** | 7002 ± 605 ** |
| Chlorophyll a/b | 2.61 ± 0.01 | 2.73 ± 0.04 *** | 2.70 ± 0.06 *** | 2.80 ± 0.004 *** |
| Lutein | 20575 ± 1592 | 19440 ± 1445 * | 17740 ± 1397 *** | 18028 ± 1413 *** |
| (A/2 + Z)/(V + A + Z) | 0.081 ± 0.002 | 0.057 ± 0.005 *** | 0.071 ± 0.002 *** | 0.074 ± 0.004 *** |
| β-Carotene | 5515 ± 480 | 5353 ± 396 | 5176 ± 455 | 4635 ± 408 *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rusaczonek, A.; Czarnocka, W.; Willems, P.; Sujkowska-Rybkowska, M.; Van Breusegem, F.; Karpiński, S. Phototropin 1 and 2 Influence Photosynthesis, UV-C Induced Photooxidative Stress Responses, and Cell Death. Cells 2021, 10, 200. https://doi.org/10.3390/cells10020200
Rusaczonek A, Czarnocka W, Willems P, Sujkowska-Rybkowska M, Van Breusegem F, Karpiński S. Phototropin 1 and 2 Influence Photosynthesis, UV-C Induced Photooxidative Stress Responses, and Cell Death. Cells. 2021; 10(2):200. https://doi.org/10.3390/cells10020200
Chicago/Turabian StyleRusaczonek, Anna, Weronika Czarnocka, Patrick Willems, Marzena Sujkowska-Rybkowska, Frank Van Breusegem, and Stanisław Karpiński. 2021. "Phototropin 1 and 2 Influence Photosynthesis, UV-C Induced Photooxidative Stress Responses, and Cell Death" Cells 10, no. 2: 200. https://doi.org/10.3390/cells10020200