Specialist Birds Replace Generalists in Grassland Remnants as Land Use Change Intensifies
 Ingmar R. Staude1,2*
Ingmar R. Staude1,2*  Gerhard E. Overbeck3
Gerhard E. Overbeck3  Carla Suertegaray Fontana4
Carla Suertegaray Fontana4  Glayson A. Bencke5
Glayson A. Bencke5  Thaiane Weinert da Silva4 Anne Mimet6
Thaiane Weinert da Silva4 Anne Mimet6  Henrique M. Pereira1,2
Henrique M. Pereira1,2- 1German Centre for Integrative Biodiversity Research (iDiv) Halle-Jena-Leipzig, Leipzig, Germany
- 2Institute of Biology, Martin-Luther University Halle-Wittenberg, Halle, Germany
- 3Department of Botany, Universidade Federal do Rio Grande do Sul, Porto Alegre, Brazil
- 4Laboratório de Ornitologia, Museu de Ciências e Tecnologia, Programa de Pós-Graduação em Ecologia e Evolução da Biodiversidade, Pontifícia Universidade Católica do Rio Grande do Sul, PUCRS, Porto Alegre, Brazil
- 5Museu de Ciências Naturais, Fundação Zoobotânica do Rio Grande do Sul, Porto Alegre, Brazil
- 6Department of Ecology and Ecosystem Management, Technische Universität München, Freising, Germany
The ideal free distribution theory predicts that mobile species distribute themselves among habitat patches so as to optimize their fitness. Changes in land use alter the quality of habitat patches and thereby affect the distribution of species. Following the loss of native habitat, habitat specialists are expected to move to patches where native habitat still remains in order to survive. Competition for resources in habitat remnants should consequently increase. As generalists are able to use other habitats, generalists are expected to gradually disappear in remnants in order to avoid increasing competition with specialists. Here, we test these predictions by studying the response of habitat specialist and generalist birds to land-use change in Brazil's southern grasslands. Using a space-for-time substitution approach, we surveyed bird communities in native grassland sites (~4 ha) in 31 regions (10 × 10 km) with differing levels of conversion to agriculture (1–94%). We found a higher abundance of specialists in native grassland patches with increasing agricultural cover in the region, while the total number of individuals in remnants remained constant. At the same time, the share of generalists in total abundance and total species richness decreased. To gain insights into whether these patterns could be driven by shifts in competition, we tested whether generalists that continued to co-occur with specialists in remnants, had less dietary overlap with specialists. As a consequence of community composition in remnants, a higher proportion of generalists were omnivorous and the average generalist species fed less on seeds, whereas the average specialist species fed more on seeds when agricultural cover was high in the region. Our results, therefore, support predictions of the ideal free distribution theory. Specialists that are assumed to have a low survivorship outside of their specialized habitat, distribute to remnants of this habitat when it is converted elsewhere, while generalists, being able to survive in other habitats, disappear gradually in remnants. Such a process could partly explain the segregation of habitat specialist and generalist birds observed in many agricultural landscapes. Finally, our results suggest that native habitat remnants can be important temporary refugia for specialists.
Introduction
The population abundance of many bird species has declined at large spatial scales over the last few decades (Rosenberg et al., 2019). Grassland birds, in particular, face high threat levels as their habitats are rapidly being converted for human land uses (Schipper et al., 2016; Stanton et al., 2018; Correll et al., 2019). While abundant empirical evidence demonstrates that generalists dominate bird communities in agricultural habitats (Lockwood et al., 2000; Devictor et al., 2008), it is much less clear how bird communities change in grassland remnants when grassland is increasingly converted to agriculture in the region. Here we study these changes in consideration of the ideal free distribution (IFD) theory (Fretwell, 1970; Tregenza, 1995).
The IFD theory predicts that mobile species, such as birds, should move between patches as habitat quality changes, so as to optimize their fitness (Tregenza, 1995). As native grassland is converted to cultivated fields, opportunities for grassland specialists to survive and reproduce are decreasing. The IFD theory therefore predicts that specialists should move to patches in which native grassland remains (Tregenza, 1995). As a result of the arrival of new specialist individuals, competition for resources in these remnants is expected to increase. Grassland birds are assumedly better at exploiting the resources they specialized on than generalists that are able to use a broad range of habitats and resources (Poisot et al., 2011; Ponge, 2013; Reif et al., 2016). The IFD theory therefore predicts that, if resources are limited, habitat generalists should gradually disappear in remnants, as they are able to use other habitats that are more relaxed from competition with specialists (MacArthur and Pianka, 1966; Tregenza, 1995).
Of all generalist species, those generalists with resource use patterns most similar to specialists are expected to be affected most by increasing specialists (MacArthur and Pianka, 1966; MacArthur and Levins, 1967). This non-randomness should lead to an observable decrease in the share of generalists in total species richness in remnants as habitat conversion increases in the region. Moreover, generalists that still co-occur with specialists in remnants should show a reduced dietary overlap with specialists. Overall, these predictions of the IFD theory would be consistent with the pattern of spatial segregation of specialists and generalists that has been observed in mosaic landscapes of disturbed and stable habitats, where specialists have been found to dominate communities in more stable habitats and generalists to aggregate in more disturbed habitats (Julliard et al., 2006; Devictor et al., 2008).
Here we test whether such predictions based on the IFD theory are supported by empirical evidence from bird communities in Brazil's southern grasslands, which face a very recent and intensive conversion to agriculture (Overbeck et al., 2015; de Oliveira et al., 2017). We used a space-for-time substitution approach and surveyed local bird communities in grassland remnants of 31 regions with differing levels of agricultural cover. First, we test whether total species abundance and the abundance of specialists (i.e., grassland birds) in native grassland remnants changes with increasing agricultural cover in the region. Second, we test for specialist-generalist segregation by assessing whether the share of generalists (i.e., non-grassland birds) in total species richness is lower in remnants when regional agricultural cover is high. Third, we test whether generalist species that continue to co-occur with specialists in remnants, have a reduced dietary overlap with specialists.
Methods
Study Region
We studied bird communities within the South Brazilian grassland region in Rio Grande do Sul (RS), Brazil's southernmost state. Climate in RS is humid subtropical with warm summers and no pronounced dry seasons (Alvares et al., 2013). In the last four decades, ~50% of natural grasslands in the region have been converted to agriculture, i.e., mainly to soybeans and rice (Cordeiro et al., 2009; de Oliveira et al., 2017). This rapid conversion rate and lack of sufficient protection give these grasslands the highest Conservation Risk Index of all Brazilian biomes (Overbeck et al., 2015).
Study Sites and Bird Sampling
We identified suitable sampling regions by assessing the spatial distribution of land use/cover types in the entire territory of RS. For this, we georeferenced and visually interpreted Landsat 5 satellite images (from 2009) (Hasenack and Weber, 2010). We adopted a 10 × 10 km grid used by the Brazilian Ministry of the Environment for national forest inventories, and calculated the percentage of different land uses in each grid cell (henceforth referred to as region). We selected 31 regions (Supplementary Figure 1; Supplementary Table 1) to represent a gradient of regional agricultural cover (1–94% agricultural cover) and the distribution of native grasslands in RS.
In each region, we conducted bird surveys at three local sites within native grassland remnants. Local site selection followed judgement by botanists (presence of native grassland) and operational criteria (accessibility and permission). Each of the three local sites per region was surveyed once with one point count covering an area of ~ 4 ha (fixed radius of 112 m) for 15 min. This time of detection is sufficient, as it is common for grassland habitats to be sampled with only 5-min point counts (Ralph et al., 1995). We acknowledge, however, that by surveying only three points within remnants per region, we may have missed species and/or undercounted individuals. Mean minimum and maximum distance between the three local sites was 538 and 1,125 m, respectively, where the recommended minimum distance between point counts is 200 m to reduce the probability of the same bird being counted at different points (Sutherland et al., 2004).
All point counts were carried out by the same team of six experienced observers during the Austral spring to summer (i.e., during the breeding season of birds in southern Brazil; October–February). In order to standardize time and reduce temporal variation, all point counts were conducted under similar weather conditions either before 10:00 a.m. or after 16:00 p.m. when birds are most conspicuous (but note that during the breeding season grassland birds can be easily detected over the whole day). Observers recorded the presence of all birds seen and heard (see Supplementary Material for more details) and their abundance (i.e., the maximum number of individuals per species) at each local site. We did not correct for imperfect detection, as we were not able to assign distance bands to all individuals. Our counts therefore present relative and not absolute abundance measures. Sampling of the 31 regions took 3 years starting in 2011 until 2014.
Classification of Birds
Generalist vs. Specialist Species
Grassland bird species were characterized sensu Azpiroz et al. (2012). That is, species that are restricted to or make extensive use of grassland habitats—thus thought to be particularly sensitive to native grassland conversion—were classified as grassland specialists (51 of 106 species). The remaining birds were considered as non-grassland specialists and henceforth referred to as generalists (Supplementary Table 2, for all species and classification).
Trophic Niches
Foraging attributes were obtained for 96 out of the 106 recorded species (91% of species) from the Elton traits database (Wilman et al., 2014). We focused on two foraging attributes to test for reduced trophic niche overlap between generalists and specialists: seed use and omnivory (Supplementary Table 2). We a priori expected grassland specialists to be superior competitors for seeds, one of the main resources of grasslands. As a response variable we therefore used the percentage of seeds in a species' diet (column “Diet-Seed” in Elton trait database; esapubs.org/archive/ecol/E095/178/metadata.php) averaged across all generalist species (and separately across all specialist species) per site. We expected a decreasing average seed consumption by generalists and an increasing average seed consumption by specialists in remnants, as agricultural cover increases in the region. Because omnivores with broad trophic niches can opportunistically forage on any available food item as per capita resource availability declines, we expected an increasing share of co-occurring generalist species to be omnivorous, as agricultural cover increases in the region. In the Elton trait database, species are defined as omnivores if they have a score of <=50% in all four diet categories (i.e., Plant and Seeds; Fruits and Nectar, Invertebrates; Vertebrates, Fish and Carrion; see column “Diet-5Cat” on esapubs.org/archive/ecol/E095/178/metadata.php). We calculated the proportion of generalist species (and separately specialist species) that are omnivores in each community.
Statistical Analysis
The focal explanatory environmental variable in all following models is regional agricultural cover (%), henceforth referred to as percent agriculture. Response variables are the average of the respective community parameter across the three local sites per region in all subsequent analyses. We model the average because the allocation of the three local sites per region was not standardized across regions due to the aforementioned operational criteria for site selection (i.e., limited accessibility and owner permission requirements prohibited entry to space sites equally across regions).
Change in Total and Specialist Abundance
We used Poisson regression models to predict total abundance and specialist abundance with percent agriculture in the region, including an observation-level random effect to account for over-/underdispersion (Harrison, 2014). Models were fitted using the function “glmer” from the LME4 package (Bates et al., 2014). Two species, Vanellus chilensis and Myiopsitta monachus, were observed in large flocks of 200 and 94 individuals, respectively. This led to outliers in total and specialist abundance (Supplementary Figures 2A,B). We thus leveled those counts to the mean number of individuals across all other sites these species occupied, rounded to the nearest integer (cf. Julliard et al., 2006). Results were robust when the unlevelled data were analyzed (Supplementary Figure 2C).
Spatial Segregation of Specialists and Generalists
To test whether changes in land use induce a spatial segregation of specialists from generalists, we tested (1) whether percent agriculture influences total richness, and (2) whether percent agriculture predicts the share of generalists in total species richness. Proportions of generalists were calculated for each local site individually and then averaged for each region. Averaged proportions were regressed on percent agriculture with a beta regression model using the BETAREG package (Zeileis et al., 2010). The relationship between total species richness and percent agriculture was assessed using the same model as for abundance.
Trophic Niche Overlap Between Generalists and Specialists
First, we modeled the percent use of seeds averaged across all generalist species (and separately across all specialist species) with percent agriculture as predictor. Second, we regressed the proportion of generalist and specialist species with an omnivorous diet on percent agriculture. Again, we calculated these response variables as the average of the three local sites. These values were regressed again on percent agriculture using a beta regression model. To accommodate 0/1 values, percent use of seeds was transformed to the open unit interval (0, 1) using a continuity correction (y*(n − 1) + 0.5)/n, where n is the sample size (Smithson and Verkuilen, 2006; Zeileis et al., 2010).
Influence of Native Grassland in the Immediate Surroundings
We tested whether the amount of native grassland in the immediate surroundings of the three local sites has an effect on community parameters. Within a 1 km radius of the three local sites' centroid, native grassland area was on average 2,500 ha (ranging from 540–3,100 ha). We added native grassland area within this radius to all the above models as an explanatory variable (Pearson correlation of native grassland area and agricultural cover in the region was ρ = −0.35). We used the asymptotic likelihood ratio test, as implemented by the function “LRTEST” from the LTEST package (Hothorn et al., 2019) to test whether adding this variable improved the statistical fit of the model.
Model Validation and Spatial Autocorrelation
Generalized linear mixed effect models were validated with DHARMA scaled residual plots (Hartig, 2017) and beta regression models with BETAREG diagnostic plots (Zeileis et al., 2010). To test for spatial autocorrelation we performed the Moran's I test on all models.
Results
Change in Total and Specialist Abundance
We found that total species abundance in grassland remnants did not change clearly with agricultural cover in the region (χ2 = 0.65, p = 0.42). However, analyzing grassland specialists separately from generalists showed a statistically clear increase in specialist abundance with increasing regional land use (χ2 = 4.4, p = 0.036; Figure 1; Supplementary Table 3). Correspondingly, the share of generalists in total abundance declined with increasing agriculture in the region (χ2 = 4.58, p = 0.032; Supplementary Figure 3).
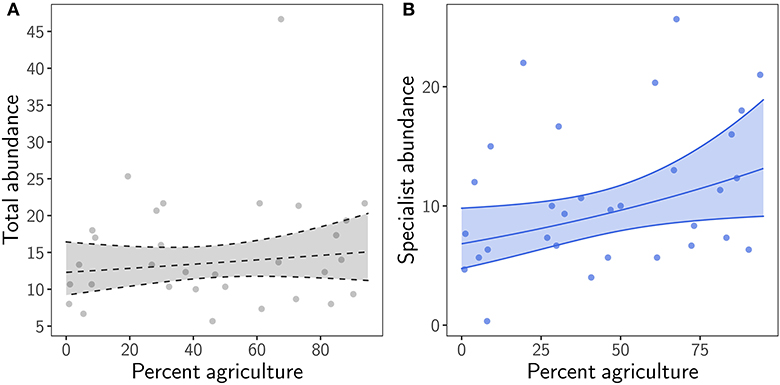
Figure 1. Relationship between percent agriculture in the region and (A) total abundance (i.e., the combined number of individuals of specialists and generalists), and (B) specialist abundance in grassland remnants. Transparent ribbon represents the 95% confidence interval around the mean. Ribbon and regression line are dashed when p ≥ 0.05.
Spatial Segregation of Specialists and Generalists
While total species richness in remnants remained constant (χ2 = 0.9, p = 0.342), the contribution of generalist species to total species richness decreased by more than 50% with increasing agricultural cover in the region (χ2 = 5, p = 0.025; Figure 2; Supplementary Table 3).
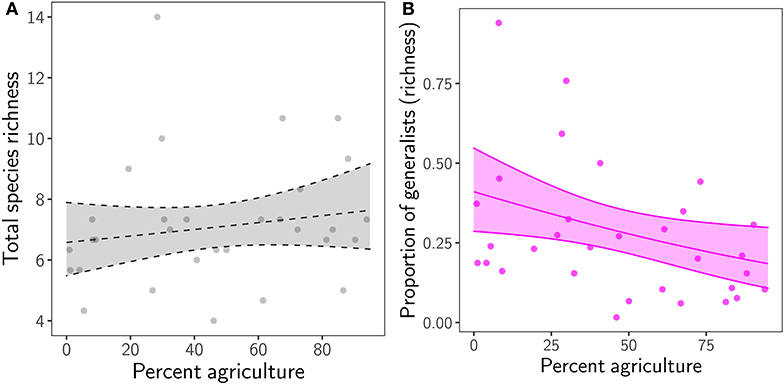
Figure 2. Relationship between percent agriculture in the region and (A) total species richness (i.e., specialists and generalists), and (B) the proportion of generalist species (to total species richness) in grassland remnants. Transparent ribbon represents the 95% confidence interval around the mean. Ribbon and regression line are dashed when p ≥ 0.05.
Trophic Niche Overlap Between Generalists and Specialists
As a consequence of species composition in remnants, the percent use of seeds averaged across generalist species strongly decreased from above 30 to <10% with more agriculture in the region (χ2 = 7.63, p = 0.005) (Figure 3A; Supplementary Table 4). In contrast, we found a marginally significant increase in the average percent use of seeds by specialists with more agriculture in the region (χ2 = 3.67, p = 0.055) (Figure 3B). The proportion of generalist species with an omnivorous diet doubled (χ2 = 4.17, p = 0.041), while the proportion of omnivorous specialist species remained constant with increasing regional agriculture (χ2 = 0, p = 0.86) (Figures 3C,D; Supplementary Table 4).
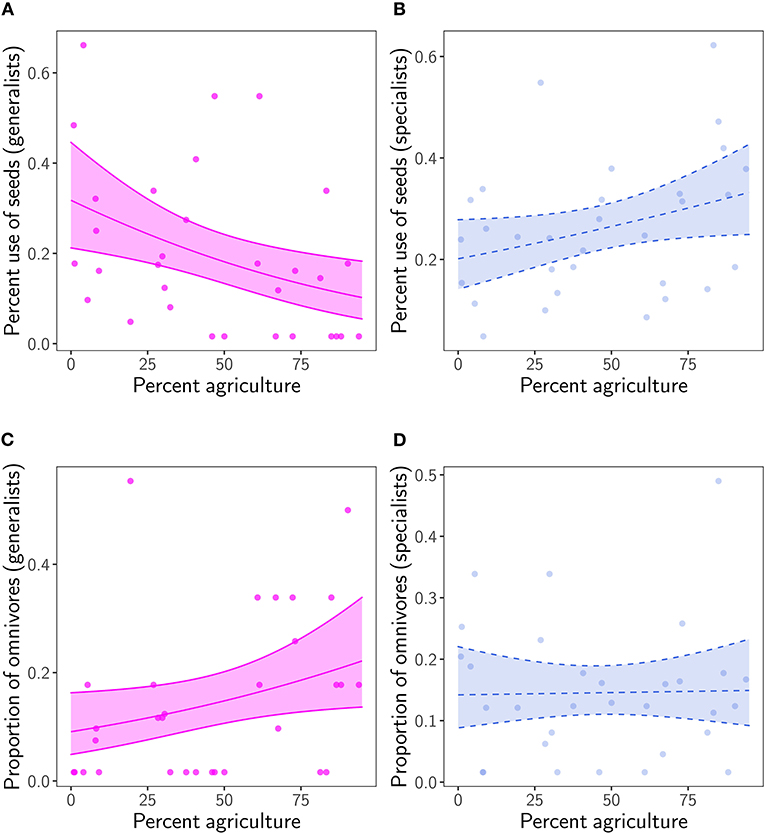
Figure 3. Relationship between percent agriculture in the region and percent use of seeds averaged (A) across generalist species and (B) specialist species, and with an omnivorous diet in (C) generalists and (D) specialists. Transparent ribbon represents the 95% confidence interval around the mean. Ribbon and regression line are dashed when p ≥ 0.05.
None of the models improved significantly when native grassland amount in the surrounding landscape was added as a second explanatory variable (Supplementary Table 5). Residuals did not show spatial autocorrelation in any model (Supplementary Figure 2).
Discussion
Using data from bird communities in native grassland remnants, we studied the response of grassland specialist and generalist birds to the increasing conversion of native grasslands to agriculture. While total species abundance and richness was constant, specialist abundance increased and the contribution of generalist species to total species abundance and richness decreased in grasslands remnants with increasing agriculture in the region. Furthermore, generalist species that continued to co-occur with specialists had a smaller dietary overlap with specialists. Together, these results support the predictions of the ideal free distribution theory: specialists that are assumed to have a low survivorship outside of their preferred habitat move away upon habitat conversion to patches where this habitat remains, while habitat generalists that are able to use other habitats, gradually disappear from these remnants.
While our results are in support of the IFD theory, our study has several limitations. We did not directly study temporal changes and instead base our inference on space-for-time substitution. Thus, we do not unequivocally show that bird communities in grassland remnants are indeed changing in response to regional land-use intensification. Moreover, we were not able to measure what individual birds were eating and whether birds changed their individual diets. We could therefore not unequivocally demonstrate that competition is driving the patterns found in this study (Dhondt, 2011). In addition, biological communities respond slower than the rate at which humans change the environment (Damgaard, 2019). Especially in our study area, land use change is very recent and rapid (Cordeiro et al., 2009), such that the patterns we found here may be transient and not stable in the long-term.
Within these limitations, we found that local species richness and total abundance of birds did not decrease in grassland remnants with increasing regional land-use change. It is likely that communities in grassland remnants show simply a lagged response (Tilman et al., 1994; Kuussaari et al., 2009). That is, species loss in intensive agriculture (Hendershot et al., 2020) and widespread negative population trends of birds (Rosenberg et al., 2019) are likely to translate to declines in richness and abundance in remnants too. However, our results suggest that at least temporarily, native habitat remnants could play an important role in sustaining the populations of some species. We found that the abundance of grassland specialists is increasing in remnants when natural habitat is increasingly converted in the region. Thus, the preservation of native habitat remnants could be an important conservation tool for sustaining populations of species that are facing loss of their specialized habitat.
The stability of total species abundance in remnants could furthermore indicate a carrying capacity and that resources are limited in remnants. The arrival of new specialist individuals is therefore likely to move the remnant beyond carrying capacity and increase the competition for resources. Since specialists are assumedly superior competitors for the resources of their specialized habitat (Reif et al., 2016), their arrival may act to replace generalists. Indeed, we found that the relative abundance and species richness of generalists has declined in remnants. Moreover, we found that within the generalist sub-assemblage, species with similar resource use pattern to specialists gradually disappeared. We therefore hypothesize that competition could indeed play a role in explaining such a replacement pattern.
The replacement of generalists by specialists, which we infer here, contrasts with biotic homogenization, the process by which rare, specialist species are widely replaced by common, generalist species owing to human activities (Lockwood et al., 2000; Mimet et al., 2019). However, given the lack of temporal data in this study, future research on mobile organisms is needed to determine whether this is a more general pattern found in matrices of agriculture and habitat remnants. For example, land-use history can influence processes underlying community change (Isbell et al., 2019; Mimet et al., 2019). In our study region, habitat remnants are still close to a pristine state (i.e., no changes in land-use occurred in the recent past) and thus more likely to support specialists. These conditions may be different in agricultural landscapes of, for example, Europe where land use change has a much longer history (Goldewijk, 2001).
Nonetheless, also in European agricultural landscapes where bird communities should have stabilized in response to land-use history, generalists seem to spatially segregate from specialists in mosaics of perturbed and more stable sites (Julliard et al., 2006; Devictor et al., 2008). While generalists dominate communities in human-modified, disturbed sites, specialists dominate in stable, more natural sites (Devictor et al., 2008). Although the mechanism behind this spatial segregation is still largely unclear, our results suggest this segregation is consistent with predictions of the IFD theory. Both specialists and generalists seek to escape declines in resources; specialists likely in response to native habitat conversion and generalists likely in response to consequent increases in specialists in habitat remnants.
We draw two conclusions from our study. First, our results support predictions of the IFD theory. Specialized birds that are assumed to have little prospect of surviving outside of their specialized habitat, increased in remnants of this habitat when it was converted elsewhere. In accordance with our hypothesis that this may increase competition in remnants, generalists with similar resource use patterns to specialists gradually disappeared in remnants. The IFD theory could therefore help explain aspects of bird community change, such as specialist-generalist segregation, in response to land-use change. Second, our results indicate that remnants of native grassland embedded in regions devoted primarily to human activities may buy time for the conservation of grassland birds (Silva et al., 2015). Considering that ongoing agricultural intensification may eventually significantly reduce local bird diversity (Hendershot et al., 2020) preserving remnants of native habitat merits more consideration in strategies to conserve biodiversity.
Data Availability Statement
Rmarkdown file with R code for all statistical analyses and data visualization, as well as the data to reproduce the findings of this study are available on figshare at: https://doi.org/10.6084/m9.figshare.13318211.v1.
Author Contributions
IS and HP conceived the study. CF, GB, and TS organized and carried out fieldwork. IS analyzed data with input from AM and wrote the manuscript with substantial input from all authors. All authors contributed to the article and approved the submitted version.
Funding
We acknowledge funding by CNPq/FAPERGS (grants 563271/2010-8 and 11/2185-0, respectively) within the SISBIOTA program. IS, AM, and HP acknowledge funding of iDiv via the German Research Foundation (DFG FZT 118). GO and CF acknowledge CNPq grants (310345/2018-9 and 457475/2012-9, 309438/2016-0, respectively).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank Valério D. Pillar, UFRGS, for project coordination and Heinrich Hasenack and Eduardo Vélez-Martin, UFRGS, for remote sensing data and site selection. We wish to thank Diogenes Machado, Maurício Silveira, Gabriel Larré, Maurício Bettio, Felipe Zilio, Eduardo Chiarani, Daniele Cantelli, and Tiago Steffen for help in the field work and/or data digitalization, and all land users for the permission to work on their land.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2020.597542/full#supplementary-material
References
Alvares, C. A., Stape, J. L., Sentelhas, P. C., de Moraes, G., Leonardo, J., and Sparovek, G. (2013). Köppen's climate classification map for Brazil. Meteorol. Zeitschrift 22, 711–728. doi: 10.1127/0941-2948/2013/0507
Azpiroz, A. B., Isacch, J. P., Dias, R. A., Di Giacomo, A. S., Fontana, C. S., and Palarea, C. M. (2012). Ecology and conservation of grassland birds in southeastern South America: a review. J. F. Ornithol. 83, 217–246. doi: 10.1111/j.1557-9263.2012.00372.x
Bates, D., Mächler, M., Bolker, B., and Walker, S. (2014). Fitting linear mixed-effects models using lme4. arXiv Preprint arXiv1406.5823. doi: 10.18637/jss.v067.i01
Cordeiro, J. L. P., Hasenack, H., et al. (2009). Cobertura Vegetal Atual do Rio Grande do Sul. Campos Sulinos: Conservação e uso Sustentável da Biodiversidade. Brasilia: Ministério do Meio Ambiente, 285–299.
Correll, M. D., Strasser, E. H., Green, A. W., and Panjabi, A. O. (2019). Quantifying specialist avifaunal decline in grassland birds of the northern great plains. Ecosphere 10:e02523. doi: 10.1002/ecs2.2523
Damgaard, C. (2019). A critique of the space-for-time substitution practice in community ecology. Trends Ecol. Evol. 34, 416–421. doi: 10.1016/j.tree.2019.01.013
de Oliveira, T. E., de Freitas, D. S., Gianezini, M., Ruviaro, C. F., Zago, D., Mércio, T. Z., et al. (2017). Agricultural land use change in the Brazilian pampa biome: the reduction of natural grasslands. Land Use Policy 63, 394–400. doi: 10.1016/j.landusepol.2017.02.010
Devictor, V., Julliard, R., and Jiguet, F. (2008). Distribution of specialist and generalist species along spatial gradients of habitat disturbance and fragmentation. Oikos 117, 507–514. doi: 10.1111/j.0030-1299.2008.16215.x
Dhondt, A. A. (2011). Interspecific Competition in Birds. Oxford; New York, NY: Oxford University Press.
Fretwell, S. D. (1970). On territorial behavior and other factors influencing habitat distribution in birds. Acta Biotheor. 19, 45–52. doi: 10.1007/BF01601955
Goldewijk, K. K. (2001). Estimating global land use change over the past 300 years: the HYDE database. Glob. Biogeochem. Cycles 15, 417–433. doi: 10.1029/1999GB001232
Harrison, X. A. (2014). Using observation-level random effects to model overdispersion in count data in ecology and evolution. PeerJ 2:e616. doi: 10.7717/peerj.616
Hartig, F. (2017). DHARMa: Residual Diagnostics for Hierarchical (Multi-Level/Mixed) Regression Models. R Package version 0.15.
Hasenack, H., and Weber, E. (2010). Base cartográfica vetorial contínua do Rio Grande do Sul-escala 1:50.000. Porto Alegre: UFRGS Centro de Ecologia 1.
Hendershot, J. N., Smith, J. R., Anderson, C. B., Letten, A. D., Frishkoff, L. O., Zook, J. R., et al. (2020). Intensive farming drives long-term shifts in avian community composition. Nature 579, 393–396. doi: 10.1038/s41586-020-2090-6
Hothorn, T., Zeileis, A., Farebrother, R. W., Cummins, C., Millo, G., Mitchell, D., et al. (2019). Package ‘lmtest’.
Isbell, F., Tilman, D., Reich, P. B., and Clark, A. T. (2019). Deficits of biodiversity and productivity linger a century after agricultural abandonment. Nat. Ecol. Evol. 3, 1533–1538. doi: 10.1038/s41559-019-1012-1
Julliard, R., Clavel, J., Devictor, V., Jiguet, F., and Couvet, D. (2006). Spatial segregation of specialists and generalists in bird communities. Ecol. Lett. 9, 1237–1244. doi: 10.1111/j.1461-0248.2006.00977.x
Kuussaari, M., Bommarco, R., Heikkinen, R. K., Helm, A., Krauss, J., Lindborg, R., et al. (2009). Extinction debt: a challenge for biodiversity conservation. Trends Ecol. Evol. 24, 564–571. doi: 10.1016/j.tree.2009.04.011
Lockwood, J. L., Brooks, T. M., and Mckinney, M. L. (2000). Taxonomic homogenization of the global avifauna. Anim. Conserv. Forum. 1, 27–35. doi: 10.1111/j.1469-1795.2000.tb00084.x
MacArthur, R., and Levins, R. (1967). The limiting similarity, convergence, and divergence of coexisting species. Am. Nat. 101, 377–385. doi: 10.1086/282505
MacArthur, R. H., and Pianka, E. R. (1966). On optimal use of a patchy environment. Am. Nat. 100, 603–609. doi: 10.1086/282454
Mimet, A., Buitenwerf, R., Sandel, B., Svenning, J.-C., and Normand, S. (2019). Recent global changes have decoupled species richness from specialization patterns in North American birds. Glob. Ecol. Biogeogr. 28, 1621–1635. doi: 10.1111/geb.12978
Overbeck, G. E., Vélez-Martin, E., Scarano, F. R., Lewinsohn, T. M., Fonseca, C. R., Meyer, S. T., et al. (2015). Conservation in Brazil needs to include non-forest ecosystems. Divers. Distrib. 21, 1455–1460. doi: 10.1111/ddi.12380
Poisot, T., Bever, J. D., Nemri, A., Thrall, P. H., and Hochberg, M. E. (2011). A conceptual framework for the evolution of ecological specialisation. Ecol. Lett. 14, 841–851. doi: 10.1111/j.1461-0248.2011.01645.x
Ponge, J.-F. (2013). Disturbances, organisms and ecosystems: a global change perspective. Ecol. Evol. 3, 1113–1124. doi: 10.1002/ece3.505
Ralph, J. C., Sauer, J. R., and Droege, S. (1995). Monitoring Bird Populations by Point Counts. Gen. Tech. Rep. PSW-GTR-149. Albany, CA: US Department of Agriculture; Forest Service; Pacific Southwest Research Station, 149.
Reif, J., Hovrák, D., Krištin, A., Kopsová, L., and Devictor, V. (2016). Linking habitat specialization with species' traits in European birds. Oikos 125, 405–413. doi: 10.1111/oik.02276
Rosenberg, K. V., Dokter, A. M., Blancher, P. J., Sauer, J. R., Smith, A. C., Smith, P. A., et al. (2019). Decline of the north american avifauna. Science 366, 120–124. doi: 10.1126/science.aaw1313
Schipper, A. M., Belmaker, J., de Miranda, M. D., Navarro, L. M., Böhning-Gaese, K., Costello, M. J., et al. (2016). Contrasting changes in the abundance and diversity of North American bird assemblages from 1971 to 2010. Glob. Change Biol. 22, 3948–3959. doi: 10.1111/gcb.13292
Silva, T. W., Dotta, G., and Fontana, C. S. (2015). Structure of avian assemblages in grasslands associated with cattle ranching and soybean agriculture in the Uruguayan savanna ecoregion of Brazil and Uruguay. Condor Ornithol. Appl. 117, 53–63. doi: 10.1650/CONDOR-14-85.1
Smithson, M., and Verkuilen, J. (2006). A better lemon squeezer? Maximum-likelihood regression with beta-distributed dependent variables. Psychol. Methods 11:54. doi: 10.1037/1082-989X.11.1.54
Stanton, R. L., Morrissey, C. A., and Clark, R. G. (2018). Analysis of trends and agricultural drivers of farmland bird declines in North America: a review. Agric. Ecosyst. Environ. 254, 244–254. doi: 10.1016/j.agee.2017.11.028
Sutherland, W. J., Newton, I., and Green, R. (2004). Bird ecology and Conservation: A Handbook of Techniques. Oxford: Oxford University Press.
Tilman, D., May, R. M., Lehman, C. L., and Nowak, M. A. (1994). Habitat destruction and the extinction debt. Nature 371, 65–66. doi: 10.1038/371065a0
Tregenza, T. (1995). “Building on the ideal free distribution,” in Advances in Ecological Research (Elsevier), 253–307.
Wilman, H., Belmaker, J., Simpson, J., de la Rosa, C., Rivadeneira, M. M., and Jetz, W. (2014). EltonTraits 1.0: species-level foraging attributes of the world's birds and mammals: ecological archives E095-178. Ecology 95:2027. doi: 10.1890/13-1917.1
Keywords: ideal free distribution, trophic niche, habitat loss, biodiversity change, Brazil
Citation: Staude IR, Overbeck GE, Fontana CS, Bencke GA, Silva TWd, Mimet A and Pereira HM (2021) Specialist Birds Replace Generalists in Grassland Remnants as Land Use Change Intensifies. Front. Ecol. Evol. 8:597542. doi: 10.3389/fevo.2020.597542
Received: 21 August 2020; Accepted: 07 December 2020;
Published: 14 January 2021.
Edited by:
John Maxwell Halley, University of Ioannina, GreeceReviewed by:
Gabor L. Lovei, Aarhus University, DenmarkNicole Michel, National Audubon Society, United States
Copyright © 2021 Staude, Overbeck, Fontana, Bencke, Silva, Mimet and Pereira. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ingmar R. Staude, ingmar.staude@idiv.de
orcid.org/0000-0003-2306-8780