Therapeutic Approaches for Metastases from Colorectal Cancer and Pancreatic Ductal Carcinoma

 ,
,
Abstract
:1. Introduction
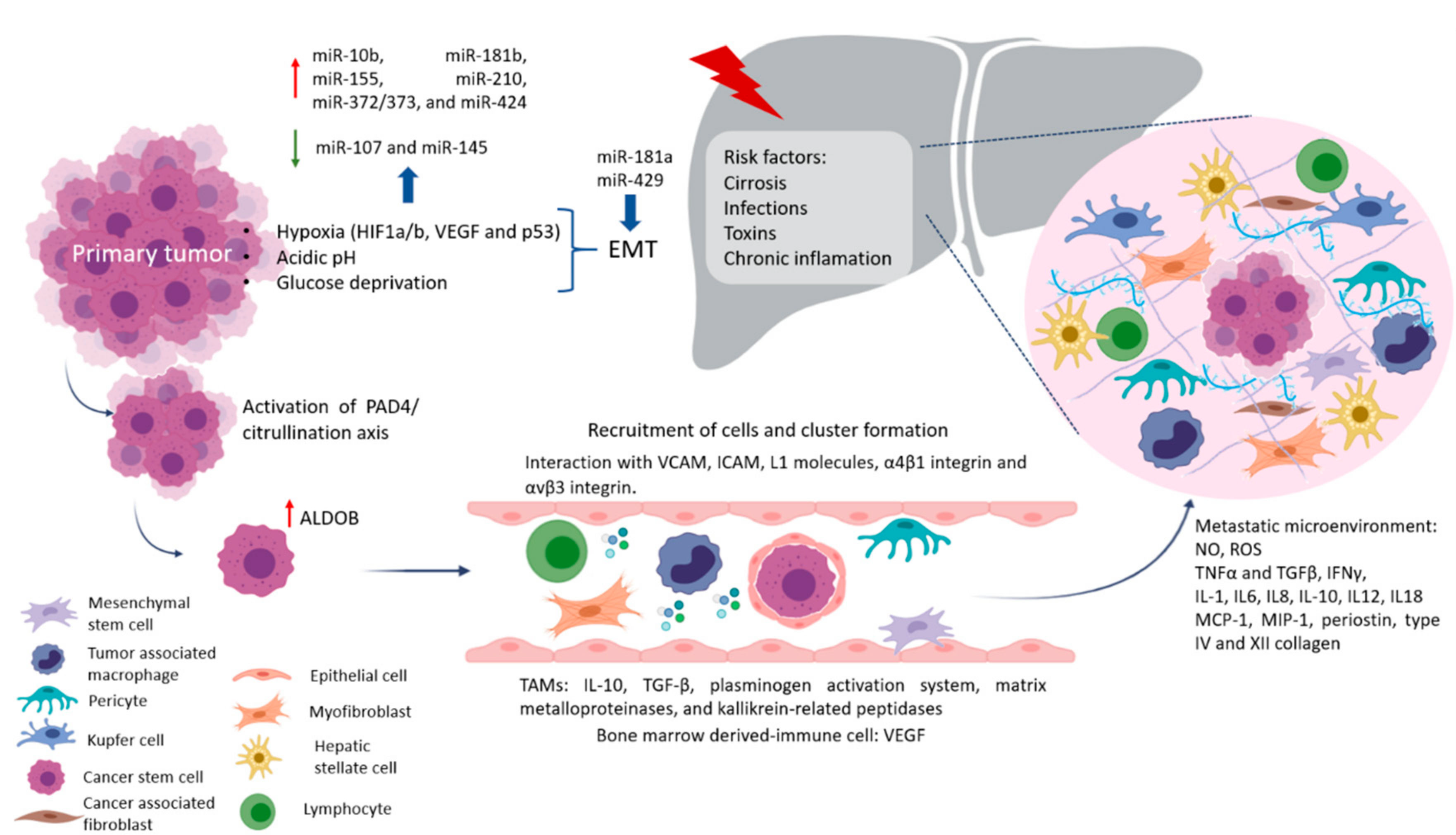
2. Why Liver Metastasis?
Factors Crucial for the Formation of Liver Metastasis
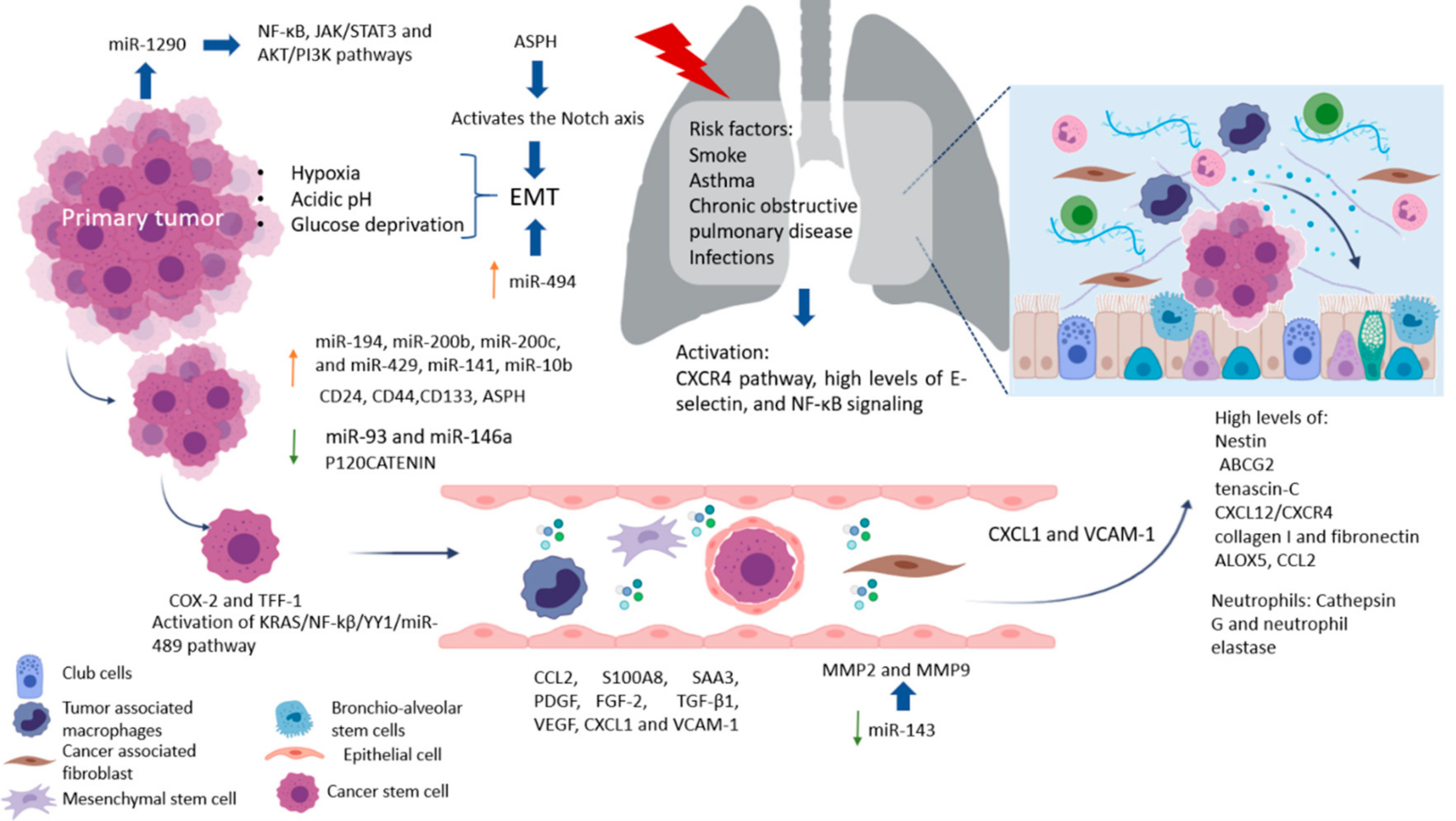
3. Lung Metastasis
Factors Crucial for the Formation of Lung Metastasis
4. Biomarkers in Metastatic Cancer
5. Novel Strategies for Liver and Lung Metastasis Treatment
5.1. Immunotherapy
5.2. Immune Checkpoint
5.3. Vaccines
5.4. Cell Therapy
5.5. Active Therapy
5.6. Passive Therapy
5.7. miRNA and siRNA
5.8. Gene Therapy
5.9. Nanomedicine
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Valastyan, S.; Weinberg, R.A. Tumor Metastasis: Molecular Insights and Evolving Paradigms. Cell 2011, 147, 275–292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quail, D.F.; Joyce, J.A. Microenvironmental regulation of tumor progression and metastasis. Nat. Med. 2013, 19, 1423–1437. [Google Scholar] [CrossRef] [PubMed]
- Kuşoğlu, A.; Avcı, Ç.B. Cancer stem cells: A brief review of the current status. Gene 2019, 681, 80–85. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Aznar, E.; Wiesmüller, L.; Sainz, J.B.; Hermann, P.C. EMT and Stemness—Key Players in Pancreatic Cancer Stem Cells. Cancers 2019, 11, 1136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsubakihara, Y.; Moustakas, A. Epithelial-Mesenchymal Transition and Metastasis under the Control of Transforming Growth Factor β. Int. J. Mol. Sci. 2018, 19, 3672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendoza, M.; Khanna, C. Revisiting the seed and soil in cancer metastasis. Int. J. Biochem. Cell Biol. 2009, 41, 1452–1462. [Google Scholar] [CrossRef] [PubMed]
- Codony-Servat, J.; Rosell, R. Cancer stem cells and immunoresistance: Clinical implications and solutions. Transl. Lung Cancer Res. 2015, 4, 689–703. [Google Scholar]
- Garza Treviño, E.N.; González, P.D.; Valencia Salgado, C.I.; Martinez Garza, A. Effects of pericytes and colon cancer stem cells in the tumor microenvironment. Cancer Cell Int. 2019, 19, 1–12. [Google Scholar] [CrossRef]
- Plaks, V.; Kong, N.; Werb, Z. The Cancer Stem Cell Niche: How Essential is the Niche in Regulating Stemness of Tumor Cells? Cell Stem Cell 2015, 16, 225–238. [Google Scholar] [CrossRef] [Green Version]
- Ren, J.; Ding, L.; Zhang, D.; Shi, G.; Xu, Q.; Shen, S.; Wang, Y.; Wang, T.; Hou, Y. Carcinoma-associated fibroblasts promote the stemness and chemoresistance of colorectal cancer by transferring exosomal lncRNA H19. Theranostics 2018, 8, 3932–3948. [Google Scholar] [CrossRef]
- Hida, K.; Maishi, N.; Annan, D.A.; Hida, Y. Contribution of Tumor Endothelial Cells in Cancer Progression. Int. J. Mol. Sci. 2018, 19, 1272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salazar, N.; Zabel, B.A. Support of Tumor Endothelial Cells by Chemokine Receptors. Front. Immunol. 2019, 10, 147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, P.P. Aneuploid Circulating Tumor-Derived Endothelial Cell (CTEC): A Novel Versatile Player in Tumor Neovascularization and Cancer Metastasis. Cells 2020, 9, 1539. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.F.; Yang, Q.; Zhang, Y.; Zhao, T.J.; Liu, X.M.; Zhong, J.; Ma, J.; Chen, Y.X.; Zhao, C.; Li, J.X. Plumbagin restrains hepatocellular carcinoma angiogenesis by suppressing the migration and invasion of tumor-derived vascular endothelial cells. Oncotarget 2017, 8, 15230–15241. [Google Scholar] [CrossRef] [Green Version]
- Cima, I.; Kong, S.L.; Sengupta, D.; Tan, I.B.; Phyo, W.M.; Lee, D.; Hu, M.; Iliescu, C.; Alexander, I.; Goh, W.L.; et al. Tumor-derived circulating endothelial cell clusters in colorectal cancer. Sci. Transl. Med. 2016, 8, 345ra89. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Zhang, X.; Liu, Y.; Zhang, T.; Wang, Z.; Gu, M.; Li, Y.; Wang, D.D.; Li, W.; Lin, P.P. PD-L1+ aneuploid circulating tumor endothelial cells (CTECs) exhibit resistance to the checkpoint blockade immunotherapy in advanced NSCLC patients. Cancer Lett. 2020, 469, 355–366. [Google Scholar] [CrossRef]
- Stella, G.M.; Kolling, S.; Benvenuti, S.; Bortolotto, C. Lung-seeking metastases. Cancers 2019, 11, 1010. [Google Scholar] [CrossRef] [Green Version]
- Jahanafrooz, Z.; Hashemzaei, M. Colon cancer therapy by focusing on colon cancer stem cells and their tumor microenvironment. J. Cell. Phys. 2019, 235, 4153–4166. [Google Scholar] [CrossRef]
- Kulke, M.H. Metastatic Pancreatic Cancer. Curr. Treat. Options Oncol. 2002, 3, 449–457. [Google Scholar] [CrossRef]
- Oweira, H.; Petrausch, U.; Helbling, D.; Schmidt, J.; Mannhart, M.; Mehrabi, A.; Schöb, O.; Giryes, A.; Decker, M.; Abdel-Rahman, O. Prognostic value of site-specific metastases in pancreatic adenocarcinoma: A Surveillance Epidemiology and End Results database analysis. World J. Gastroenterol. 2017, 23, 1872–1880. [Google Scholar] [CrossRef]
- Bellon, E.; Gebauer, F.; Tachezy, M.; Izbicki, J.R.; Bockhorn, M. Pancreatic cancer and liver metastases: State of the art. Update Surg. 2016, 68, 247–251. [Google Scholar] [CrossRef] [PubMed]
- Deeb, A.; Haque, S.-U.; Olowokure, O. Pulmonary metastases in pancreatic cancer, is there a survival influence? J. Gastrointest. Oncol. 2015, 6, E48–E51. [Google Scholar] [PubMed]
- Trefts, E.; Gannon, M.; Wasserman, D.H. The Liver. Curr Biol. 2017, 27, R1147–R1151. [Google Scholar] [CrossRef] [PubMed]
- Zarour, L.R.; Anand, S.; Billingsley, K.G.; Bisson, W.H.; Cercek, A.; Clarke, M.F.; Coussens, L.M.; Gast, C.E.; Geltzeiler, C.B.; Hansen, L.; et al. Colorectal Cancer Liver Metastasis: Evolving Paradigms and Future Directions. Cell. Mol. Gastroenterol. Hepatol. 2017, 3, 163–173. [Google Scholar] [CrossRef] [Green Version]
- Carneiro, C.; Brito, J.; Bilreiro, C.; Barros, M.; Bahia, C.; Santiago, I.; Caseiro-Alves, F. All about portal vein: A pictorial display to anatomy, variants and physiopathology. Insights Imaging 2019, 10, 38. [Google Scholar] [CrossRef] [Green Version]
- Riihimäki, M.; Hemminki, A.; Sundquist, J.; Hemminki, K. Patterns of metastasis in colon and rectal cancer. Sci. Rep. 2016, 6, 29765. [Google Scholar] [CrossRef] [Green Version]
- Rossetto, A.; De Re, V.; Steffan, A.; Ravaioli, M.; Miolo, G.; Leone, P.; Racanelli, V.; Uzzau, A.; Baccarani, U.; Cescon, M. Carcinogenesis and Metastasis in Liver: Cell Physiological Basis. Cancers 2019, 11, 1731. [Google Scholar] [CrossRef] [Green Version]
- Bendas, G.; Borsig, L. Cancer Cell Adhesion and Metastasis: Selectins, Integrins, and the Inhibitory Potential of Heparins. Int. J. Cell Biol. 2012, 2012, 1–10. [Google Scholar] [CrossRef]
- Yuzhalin, A.E.; Gordon-Weeks, A.; Tognoli, M.L.; Jones, K.; Markelc, B.; Konietzny, R.; Fischer, R.; Muth, A.; O’Neill, E.; Thompson, P.R.; et al. Colorectal cancer liver metastatic growth depends on PAD4-driven citrullination of the extracellular matrix. Nat. Commun. 2018, 9, 1–15. [Google Scholar] [CrossRef]
- Gonzalez-Villarreal, C.A.; Quiroz-Reyes, A.G.; Islas, J.F.; Garza-Treviño, E.N. Colorectal Cancer Stem Cells in the Progression to Liver Metastasis. Front. Oncol. 2020, 10, 1–17. [Google Scholar] [CrossRef]
- Winkler, J.; Abisoye-Ogunniyan, A.; Metcalf, K.J.; Werb, Z. Concepts of extracellular matrix remodelling in tumour progression and metastasis. Nat. Commun. 2020, 11, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, H.; Hagerling, C.; Werb, Z. Roles of the immune system in cancer: From tumor initiation to metastatic progression. Genes Dev. 2018, 32, 1267–1284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cortese, N.; Soldani, C.; Franceschini, B.; Barbagallo, M.; Marchesi, F.; Torzilli, G.; Donadon, M. Macrophages in Colorectal Cancer Liver Metastases. Cancers 2019, 11, 633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goh, C.; Narayanan, S.; Hahn, Y.S. Myeloid-derived suppressor cells: The dark knight or the joker in viral infections? Immunol. Rev. 2013, 255, 210–221. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.-Q.; Hu, X.-W.; Liu, Y.-L.; Ye, Z.-J.; Gui, Y.-H.; Zhou, D.-L.; Qi, C.-L.; He, X.-D.; Wang, H.; Wang, L.-J. CD11b deficiency suppresses intestinal tumor growth by reducing myeloid cell recruitment. Sci. Rep. 2015, 5, 15948. [Google Scholar] [CrossRef]
- Zhou, S.-N.; Pan, W.-T.; Pan, M.-X.; Luo, Q.-Y.; Zhang, L.; Lin, J.-Z.; Zhao, Y.-J.; Yan, X.-L.; Yuan, L.-P.; Zhang, Y.-X.; et al. Comparison of Immune Microenvironment Between Colon and Liver Metastatic Tissue in Colon Cancer Patients with Liver Metastasis. Dig. Dis. Sci. 2020, 1–9. [Google Scholar] [CrossRef]
- Islas, J.F.; Moreno-Cuevas, J.E. A MicroRNA Perspective on Cardiovascular Development and Diseases: An Update. Int. J. Mol. Sci. 2018, 19, 2075. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, L.S.; Fregeac, J.; Bole-Feysot, C.; Cagnard, N.; Iyer, A.; Anink, J.J.; Aronica, E.; Alibeu, O.; Nitschke, P.; Colleaux, L. Role of miR-146a in neural stem cell differentiation and neural lineage determination: Relevance for neurodevelopmental disorders. Mol. Autism 2018, 9, 1–12. [Google Scholar] [CrossRef]
- Vila-Navarro, E.; Fernandez-Castañer, E.; Rovira-Rigau, M.; Raimondi, G.; Vila-Casadesus, M.; Lozano, J.J.; Soubeyran, P.; Iovanna, J.; Castells, A.; Fillat, C.; et al. MiR-93 is related to poor prognosis in pancreatic cancer and promotes tumor progression by targeting microtubule dynamics. Oncog. 2020, 9, 1–14. [Google Scholar] [CrossRef]
- Fabian, M.R.; Mathonnet, G.; Sundermeier, T.; Mathys, H.; Zipprich, J.T.; Svitkin, Y.V.; Rivas, F.; Jinek, M.; Wohlschlegel, J.; Doudna, J.A.; et al. Mammalian miRNA RISC Recruits CAF1 and PABP to Affect PABP-Dependent Deadenylation. Mol. Cell 2009, 35, 868–880. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Lei, C.; He, Q.; Pan, Z.; Xiao, D.; Tao, Y. Nuclear functions of mammalian MicroRNAs in gene regulation, immunity and cancer. Mol. Cancer 2018, 17, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun Lee, Y.; Dutta, A. MicroRNAs in cancer. Annu. Rev. Pathol. 2009, 4, 199–227. [Google Scholar] [CrossRef]
- Muz, B.; De La Puente, P.; Azab, F.; Azab, A.K. The role of hypoxia in cancer progression, angiogenesis, metastasis, and resistance to therapy. Hypoxia 2015, 3, 83–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shibue, T.; Weinberg, R.A. EMT, CSCs, and drug resistance: The mechanistic link and clinical implications. Nat. Rev. Clin. Oncol. 2017, 14, 611–629. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.; Zuo, J. Emerging roles of miR-210 and other non-coding RNAs in the hypoxic response Non-coding RNAs—Critical Regulators of Gene Function. Acta Biochim. Biophys. Sin. 2014, 46, 220–232. [Google Scholar] [CrossRef] [Green Version]
- Cho, H.-S.; Han, T.-S.; Hur, K.; Ban, H.S. The Roles of Hypoxia-Inducible Factors and Non-Coding RNAs in Gastrointestinal Cancer. Genes 2019, 10, 1008. [Google Scholar] [CrossRef] [Green Version]
- Al Tameemi, W.; Dale, T.P.; Al-Jumaily, R.M.K.; Forsyth, N. Hypoxia-Modified Cancer Cell Metabolism. Front. Cell Dev. Biol. 2019, 7, 4. [Google Scholar] [CrossRef] [Green Version]
- Soon, P.; Kiaris, H. MicroRNAs in the tumour microenvironment: Big role for small players. Endocr. Relat. Cancer 2013, 20, R257–R267. [Google Scholar] [CrossRef]
- Graham, R.M.; Frazier, D.P.; Thompson, J.W.; Haliko, S.; Li, H.; Wasserlauf, B.J.; Spiga, M.-G.; Bishopric, N.H.; Webster, K.A. A unique pathway of cardiac myocyte death caused by hypoxia-acidosis. J. Exp. Biol. 2004, 207, 3189–3200. [Google Scholar] [CrossRef] [Green Version]
- Vasilaki, E.; Morikawa, M.; Koinuma, D.; Mizutani, A.; Hirano, Y.; Ehata, S.; Sundqvist, A.; Kawasaki, N.; Cedervall, J.; Olsson, A.-K.; et al. Ras and TGF-β signaling enhance cancer progression by promoting the ΔNp63 transcriptional program. Sci. Signal. 2016, 9, ra84. [Google Scholar] [CrossRef]
- Tsilimigras, D.I.; Ntanasis-Stathopoulos, I.; Bagante, F.; Moris, D.; Cloyd, J.M.; Spartalis, E.; Pawlik, T.M. Clinical significance and prognostic relevance of KRAS, BRAF, PI3K and TP53 genetic mutation analysis for resectable and unresectable colorectal liver metastases: A systematic review of the current evidence. Surg. Oncol. 2018, 27, 280–288. [Google Scholar] [CrossRef] [PubMed]
- AshokKumar, P.; Divya, T.; Kumar, K.; Dineshbabu, V.; Velavan, B.; Sudhandiran, G. Colorectal carcinogenesis: Insights into the cell death and signal transduction pathways: A review. World J. Gastrointest. Oncol. 2018, 10, 244–259. [Google Scholar] [CrossRef] [PubMed]
- Chae, Y.C. Cancer stem cell metabolism: Target for cancer therapy. BMB Rep. 2018, 51, 319–326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Francesco, E.M.; Sotgia, F.; Lisanti, M.P. Cancer stem cells (CSCs): Metabolic strategies for their identification and eradication. Biochem. J. 2018, 475, 1611–1634. [Google Scholar] [CrossRef] [Green Version]
- Bu, P.; Chen, K.-Y.; Xiang, K.; Johnson, C.; Crown, S.B.; Rakhilin, N.; Ai, Y.; Wang, L.; Xi, R.; Astapova, I.; et al. Aldolase B-Mediated Fructose Metabolism Drives Metabolic Reprogramming of Colon Cancer Liver Metastasis. Cell Metab. 2018, 27, 1249–1262. [Google Scholar] [CrossRef] [Green Version]
- Paschos, K.A. Natural history of hepatic metastases from colorectal cancer—pathobiological pathways with clinical significance. World J. Gastroenterol. 2014, 20, 3719–3737. [Google Scholar] [CrossRef]
- Lin, Q.; Ren, L.; Jian, M.; Xu, P.; Li, J.; Zheng, P.; Feng, Q.; Yang, L.; Ji, M.; Wei, Y.; et al. The mechanism of the premetastatic niche facilitating colorectal cancer liver metastasis generated from myeloid-derived suppressor cells induced by the S1PR1–STAT3 signaling pathway. Cell Death Dis. 2019, 10, 1–18. [Google Scholar] [CrossRef]
- Nielsen, S.R.; Quaranta, V.; Linford, A.; Emeagi, P.; Rainer, C.; Santos, A.; Ireland, L.; Sakai, T.; Sakai, K.; Kim, Y.-S.; et al. Macrophage-secreted granulin supports pancreatic cancer metastasis by inducing liver fibrosis. Nat. Cell Biol. 2016, 18, 549–560. [Google Scholar] [CrossRef] [Green Version]
- Hu, C.-T.; Guo, L.-L.; Feng, N.; Zhang, L.; Zhou, N.; Ma, L.-L.; Shen, L.; Tong, G.-H.; Yan, Q.-W.; Zhu, S.-J.; et al. MIF, secreted by human hepatic sinusoidal endothelial cells, promotes chemotaxis and outgrowth of colorectal cancer in liver prometastasis. Oncotarget 2017, 6, 22410–22423. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Liu, R.; Yang, F.; Cheng, R.; Chen, X.; Cui, S.; Gu, Y.; Sun, W.; You, C.; Liu, Z.; et al. miR-19a promotes colorectal cancer proliferation and migration by targeting TIA1. Mol. Cancer 2017, 16, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Cellura, D.; Pickard, K.; Quaratino, S.; Parker, H.; Strefford, J.; Thomas, G.; Mitter, R.; Mirnezami, A.; Peake, N. miR-19–Mediated Inhibition of Transglutaminase-2 Leads to Enhanced Invasion and Metastasis in Colorectal Cancer. Mol. Cancer Res. 2015, 13, 1095–1105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jablonska, E.; Gorniak, P.; Prusisz, W.; Kiliszek, P.; Szydlowski, M.; Sewastianik, T.; Bialopiotrowicz, E.; Polak, A.; Prochorec-Sobieszek, M.; Szumera-Cieckiewicz, A.; et al. MiR-155 Amplifies AKT and NFkB Signaling By Targeting Multiple Regulators of BCR Signal in DLBCL. Blood 2015, 126, 2455. [Google Scholar] [CrossRef]
- Ling, X.; Yao, D.; Kang, L.; Zhou, J.; Zhou, Y.; Dong, H. Involment of RAS/ERK1/2 signaling and MEF2C in miR-155-3p inhibition-triggered cardiomyocyte differentiation of embryonic stem cell. Oncotarget 2017, 8, 84403–84416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, F.; Song, D.; Wu, Y.; Liu, X.; Zhu, J.; Tang, Y. MiR-155 inhibits proliferation and invasion by directly targeting PDCD4 in non-small cell lung cancer. Thorac. Cancer 2017, 8, 613–619. [Google Scholar] [CrossRef] [Green Version]
- Balacescu, O.; Sur, D.; Cainap, C.; Vișan, S.; Cruceriu, D.; Manzat-Saplacan, R.; Muresan, M.-S.; Balacescu, L.; Lisencu, C.; Irimie, A. The Impact of miRNA in Colorectal Cancer Progression and Its Liver Metastases. Int. J. Mol. Sci. 2018, 19, 3711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, S.; Tan, X.; Zhongcheng, H.; Chen, Z.; Lin, P.; Fu, W. microNA biomarkers in colorectal cancer liver metastasis. J. Cancer 2018, 9, 3867–3873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maru, Y. The lung metastatic niche. J. Mol. Med. 2015, 93, 1185–1192. [Google Scholar] [CrossRef]
- El Rayes, T.; Catena, R.; Lee, S.; Stawowczyk, M.; Joshi, N.; Fischbach, C.; Powell, C.A.; Dannenberg, A.J.; Altorki, N.K.; Gao, D.; et al. Lung inflammation promotes metastasis through neutrophil protease-mediated degradation of Tsp-1. Proc. Natl. Acad. Sci. USA 2015, 112, 16000–16005. [Google Scholar] [CrossRef] [Green Version]
- Font-Clos, F.; Zapperi, S.; La Porta, C.A.M. Blood Flow Contributions to Cancer Metastasis. iScience 2020, 23, 101073. [Google Scholar] [CrossRef]
- Shin, S.H.; Kim, H.J.; Hwang, D.W.; Lee, J.H.; Song, K.B.; Jun, E.; Shim, I.K.; Hong, S.-M.; Kim, H.J.; Park, K.M.; et al. The DPC4/SMAD4 genetic status determines recurrence patterns and treatment outcomes in resected pancreatic ductal adenocarcinoma: A prospective cohort study. Oncotarget 2017, 8, 17945–17959. [Google Scholar] [CrossRef] [Green Version]
- Yang, G.; Sau, C.; Lai, W.; Cichon, J.; Li, W. miR-93-directed down-regulation of DAB2 defines a novel oncogenic pathway in lung cancer. Oncogene 2014, 33, 4307–4315. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Zhang, S.; Guo, C.; Li, J.; Sang, W. Downregulation of miR-200c protects cardiomyocytes from hypoxia-induced apoptosis by targeting GATA-4. Int. J. Mol. Med. 2017, 39, 1589–1596. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, S.K.; Arora, S.; Singh, S.; Bhardwaj, A.; Averett, C.; Singh, A.P. MicroRNAs in pancreatic malignancy: Progress and promises. Cancer Lett. 2014, 347, 167–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Zhu, Y.; Hu, L.; Yan, F.; Chen, J. miR-494 induces EndMT and promotes the development of HCC (Hepatocellular Carcinoma) by targeting SIRT3/TGF-β/SMAD signaling pathway. Sci. Rep. 2019, 9, 7213. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Bai, Z.; Song, J.; Yang, Y.; Wang, J.; Han, W.; Zhang, J.; Meng, H.; Ma, X.; Yang, Y.; et al. Differential expression of serum miR-126, miR-141 and miR-21 as novel biomarkers for early detection of liver metastasis in colorectal cancer. Chin. J. Cancer Res. 2014, 26, 95–103. [Google Scholar]
- Lee, S.Y.; Ju, M.K.; Jeon, H.M.; Lee, Y.J.; Kim, C.H.; Park, H.G.; Han, S.I.; Kang, H.S. Oncogenic Metabolism Acts as a Prerequisite Step for Induction of Cancer Metastasis and Cancer Stem Cell Phenotype. Oxidative Med. Cell. Longev. 2018, 2018, 1–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Ren, D.; Wu, X.; Lin, X.; Ye, L.; Lin, C.; Wu, S.; Zhu, J.; Peng, X.; Song, L. miR-1266 Contributes to Pancreatic Cancer Progression and Chemoresistance by the STAT3 and NF-κB Signaling Pathways. Mol. Ther. Nucleic Acids 2018, 11, 142–158. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Dong, Q.; Li, J.; Zhang, K.; Qin, J.; Zhao, J.; Sun, Q.; Wang, Z.; Wartmann, T.; Jauch, K.W.; et al. Targeting cancer stem cells and their niche: Perspectives for future therapeutic targets and strategies. Semin. Cancer Biol. 2018, 53, 139–155. [Google Scholar] [CrossRef]
- Sheedy, P.; Medarova, Z. The fundamental role of miR-10b in metastatic cancer. Am. J. Cancer Res. 2018, 8, 1674–1688. [Google Scholar]
- Vychytilova-Faltejskova, P.; Pesta, M.; Radova, L.; Liska, V.; Daum, O.; Kala, Z.; Svoboda, M.; Kiss, I.; Capoor, M.N. Genome-wide microRNA Expression Profiling in Primary Tumors and Matched Liver Metastasis of Patients with Colorectal Cancer. Cancer Genom. Proteom. 2016, 13, 311–316. [Google Scholar]
- Abdelmaksoud-Dammak, R.; Chamtouri, N.; Triki, M.; Saadallah-Kallel, A.; Ayadi, W.; Charfi, S.; Khabir, A.; Ayadi, L.; Sallemi-Boudawara, T.; Mokdad-Gargouri, R. Overexpression of miR-10b in colorectal cancer patients: Correlation with TWIST-1 and E-cadherin expression. Tumor Biol. 2017, 39, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, M.A.; Zubair, H.; Srivastava, S.K.; Singh, S.; Singh, A.P. Insights into the Role of microRNAs in Pancreatic Cancer Pathogenesis: Potential for Diagnosis, Prognosis, and Therapy. In microRNA: Cancer; Springer: Cham, Germany, 2015; pp. 71–87. [Google Scholar] [CrossRef] [Green Version]
- Fang, C.; Dai, C.-Y.; Mei, Z.; Jiang, M.-J.; Gu, D.-N.; Huang, Q.; Tian, L. microRNA-193a stimulates pancreatic cancer cell repopulation and metastasis through modulating TGF-β2/TGF-βRIII signalings. J. Exp. Clin. Cancer Res. 2018, 37, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Liu, P.; Wu, H.; Cui, P.; Li, Y.; Liu, Y.; Liu, Z.; Gou, S. MicroRNA-323-3p inhibits cell invasion and metastasis in pancreatic ductal adenocarcinoma via direct supression of SMAD2 and SMAD3. Oncotarget 2016, 7, 14912–14924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lanfranca, M.P.; Thompson, J.K.; Bednar, F.; Halbrook, C.; Lyssiotis, C.; Levi, B.; Frankel, T.L. Metabolism and Epigenetics of Pancreatic Cancer Stem Cells. Semin. Cancer Biol. 2019, 57, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, Y.; Yoshimura, H.; Ueda, J.; Naito, Z.; Korc, M.; Ishiwata, T. Nestin delineates pancreatic cancer stem cells in metastatic foci of NOD/Shi-scid IL2Rγ(null) (NOG) mice. Am. J. Pathol. 2014, 184, 674–685. [Google Scholar] [CrossRef] [Green Version]
- Reichert, M.; Bakir, B.; Moreira, L.; Pitarresi, J.R.; Feldmann, K.; Simon, L.; Suzuki, K.; Maddipati, R.; Rhim, A.D.; Schlitter, A.M.; et al. Regulation of Epithelial Plasticity Determines Metastatic Organotropism in Pancreatic Cancer. Dev. Cell 2018, 45, 696–711.e8. [Google Scholar] [CrossRef] [Green Version]
- Ogawa, K.; Lin, Q.; Li, L.; Bai, X.; Chen, X.; Chen, H.; Kong, R.; Wang, Y.; Zhu, H.; He, F.; et al. Prometastatic secretome trafficking via exosomes initiates pancreatic cancer pulmonary metastasis. Cancer Lett. 2020, 481, 63–75. [Google Scholar] [CrossRef]
- Apte, M.V.; Pothula, S.P.; Wilson, J.S.; Apte, M.V. Pancreatic cancer and its stroma: A conspiracy theory. World J. Gastroenterol. 2014, 20, 11216–11229. [Google Scholar] [CrossRef]
- Yuan, P.; He, X.H.; Rong, Y.F.; Cao, J.; Li, Y.; Hu, Y.P.; Liu, Y.; Li, D.; Lou, W.; Liu, M.F. KRAS/NF-kb/YY1/miR489 Signaling Axis Controls Pancreatic Cancer Metastasis. Mol. Cell. Pathobiol. 2016, 28, 100–111. [Google Scholar]
- Ta, N.; Huang, X.; Zheng, K.; Zhang, Y.; Gao, Y.; Deng, L.; Zhang, B.; Jiang, H.; Zheng, J. miRNA-1290 Promotes Aggressiveness in Pancreatic Ductal Adenocarcinoma by Targeting IKK1. Cell. Physiol. Biochem. 2018, 51, 711–728. [Google Scholar] [CrossRef]
- Ma, Y.; Yu, S.; Zhao, W.; Lu, Z.; Chen, J. miR-27a regulates the growth, colony formation and migration of pancreatic cancer cells by targeting Sprouty2. Cancer Lett. 2010, 298, 150–158. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Cao, Z.; Yang, G.; You, L.; Zhang, T.; Zhao, Y.-P. MicroRNA-27a (miR-27a) in Solid Tumors: A Review Based on Mechanisms and Clinical Observations. Front. Oncol. 2019, 9, 893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamada, S.; Masamune, A.; Miura, S.; Satoh, K.; Shimosegawa, T. MiR-365 induces gemcitabine resistance in pancreatic cancer cells by targeting the adaptor protein SHC1 and pro-apoptotic regulator BAX. Cell. Signal. 2014, 26, 179–185. [Google Scholar] [CrossRef] [PubMed]
- Xiao, X.; Yang, D.; Gong, X.; Mo, D.; Pan, S.; Xu, J. miR-1290 promotes lung adenocarcinoma cell proliferation and invasion by targeting SOCS4. Oncotarget 2018, 9, 11977–11988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, J.; Yang, L.; Wu, Y.-N.; Xu, J. Serum miR-1290 and miR-1246 as Potential Diagnostic Biomarkers of Human Pancreatic Cancer. J. Cancer 2020, 11, 1325–1333. [Google Scholar] [CrossRef]
- Hidalgo-Sastre, A.; Lubeseder-Martellato, C.; Engleitner, T.; Steiger, K.; Zhong, S.; Desztics, J.; Öllinger, R.; Rad, R.; Schmid, R.M.; Hermeking, H.; et al. Mir34a constrains pancreatic carcinogenesis. Sci. Rep. 2020, 10, 9654. [Google Scholar] [CrossRef]
- Zhou, R.; Yuan, P.; Wang, Y.; Hunsberger, J.G.; Elkahloun, A.G.; Wei, Y.; Damschroder-Williams, P.; Du, J.; Chen, G.; Manji, H.K. Evidence for Selective microRNAs and Their Effectors as Common Long-Term Targets for the Actions of Mood Stabilizers. Neuropsychopharmacology 2009, 34, 1395–1405. [Google Scholar] [CrossRef]
- Chao, C.; Lee, C.; Chang, T.; Chen, P.; Liu, J. CXCL1/CXCR2 Paracrine Axis Contributes to Lung Metastasis in Osteosarcoma. Cancers 2020, 12, 459. [Google Scholar] [CrossRef] [Green Version]
- Stewart, C.L.; Warner, S.; Ito, K.; Raoof, M.; Wu, G.X.; Lu, W.P.; Kessler, J.; Kim, J.Y.; Fong, Y. Cytoreduction for Colorectal Metastases: Liver, Lung, Peritoneum, Lymph Nodes, Bone, Brain. When Does it Palliate, Prolong Survival, and Potentially Cure. Curr Probl Surg. 2018, 55, 330–379. [Google Scholar] [CrossRef]
- Pfannschmidt, J.; Muley, T.; Hoffmann, H.; Dienemann, H. Prognostic factors and survival after complete resection of pulmonary metastases from colorectal carcinoma: Experiences in 167 patients. J. Thorac. Cardiovasc. Surg. 2003, 126, 732–739. [Google Scholar] [CrossRef] [Green Version]
- Garza Treviño, E.N.; Delgado-Gonzalez, P.; Valencia Salgado, C.I.; Ortega Garcia, J.L. Pericytes Relationship with Cancer Stem Cells in the Colon. Curr. Tissue Microenviron. Rep. 2020, 1, 187–198. [Google Scholar] [CrossRef]
- Amin, D.N.; Hida, K.; Bielenberg, D.R.; Klagsbrun, M. Tumor Endothelial Cells Express Epidermal Growth Factor Receptor (EGFR) but not ErbB3 and Are Responsive to EGF and to EGFR Kinase Inhibitors. Cancer Res. 2006, 66, 2173–2180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.F.; Wu, K.J. Endothelial transdifferentiation of tumor cells triggered by the Twist1-Jagged1-KLF4 axis: Relationship between cancer stemness and angiogenesis. Stem Cells Int. 2016, 2016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schizas, D.; Charalampakis, N.; Kole, C.; Economopoulou, P.; Koustas, E.; Gkotsis, E.; Ziogas, D.C.; Psyrri, A.; Karamouzis, M.V. Immunotherapy for pancreatic cancer: A 2020 update. Cancer Treat. Rev. 2020, 86, 102016. [Google Scholar] [CrossRef] [PubMed]
- Sahin, I.H.; Elias, H.; Chou, J.F.; Capanu, M.; O’Reilly, E.M. Pancreatic adenocarcinoma: Insights into patterns of recurrence and disease behavior. BMC Cancer 2018, 18, 769. [Google Scholar] [CrossRef]
- Liu, J.; Xi, L.; Yu, Y.; Chen, N.; Wang, S.; Sun, P.; Liu, A.; Zhang, X.; Gong, W.; Zhang, X.; et al. Thermal ablation for colorectal pulmonary metastases: A meta-analysis. Int. J. Clin. Exp. Med. 2018, 11, 11521–11534. [Google Scholar]
- Puijk, R.S.; COLLISION Trial Group; Ruarus, A.H.; Vroomen, L.; Van Tilborg, A.A.J.M.; Scheffer, H.J.; Nielsen, K.; De Jong, M.C.; De Vries, J.J.J.; Zonderhuis, B.M.; et al. Colorectal liver metastases: Surgery versus thermal ablation (COLLISION)—A phase III single-blind prospective randomized controlled trial. BMC Cancer 2018, 18, 821. [Google Scholar] [CrossRef]
- Gurusamy, K.S.; Corrigan, N.; Croft, J.; Twiddy, M.; Morris, S.; Woodward, N.; Bandula, S.; Hochhauser, D.; Napp, V.; Pullan, A.; et al. Liver resection surgery versus thermal ablation for colorectal LiVer MetAstases (LAVA): Study protocol for a randomised controlled trial. Trials 2018, 19, 105. [Google Scholar] [CrossRef] [Green Version]
- Dizon, D.S.; Schwartz, J.; Kemeny, N. Regional Chemotherapy: A Focus on Hepatic Artery Infusion for Colorectal Cancer Liver Metastases. Surg. Oncol. Clin. N. Am. 2008, 17, 759–771. [Google Scholar] [CrossRef]
- Ammori, J.B.; Kemeny, N.E. Regional Hepatic Chemotherapies in Treatment of Colorectal Cancer Metastases to the Liver. Semin. Oncol. 2010, 37, 139–148. [Google Scholar] [CrossRef]
- Welsh, J.S.; Kennedy, A.S.; Thomadsen, B. Selective internal radiation therapy (SIRT) for liver metastases secondary to colorectal adenocarcinoma. Int. J. Radiat. Oncol. 2006, 66, S62–S73. [Google Scholar] [CrossRef] [PubMed]
- Robin, T.P.; Raben, D.; Schefter, T. A Contemporary Update on the Role of Stereotactic Body Radiation Therapy (SBRT) for Liver Metastases in the Evolving Landscape of Oligometastatic Disease Management. Semin. Radiat. Oncol. 2018, 28, 288–294. [Google Scholar] [CrossRef] [PubMed]
- Riley, R.S.; June, C.H.; Langer, R.; Mitchell, M.J. Delivery technologies for cancer immunotherapy. Nat. Rev. Drug Discov. 2019, 18, 175–196. [Google Scholar] [CrossRef] [PubMed]
- Greenwald, R.J.; Freeman, G.J.; Sharpe, A.H. THE B7 FAMILY REVISITED. Annu. Rev. Immunol. 2005, 23, 515–548. [Google Scholar] [CrossRef]
- Nolte, M.A.; Van Olffen, R.W.; Van Gisbergen, K.P.J.M.; Van Lier, R.A.W. Timing and tuning of CD27-CD70 interactions: The impact of signal strength in setting the balance between adaptive responses and immunopathology. Immunol. Rev. 2009, 229, 216–231. [Google Scholar] [CrossRef]
- Liu, L.; Yin, B.; Yi, Z.; Liu, X.; Hu, Z.; Gao, W.; Yu, H.; Li, Q. Breast cancer stem cells characterized by CD70 expression preferentially metastasize to the lungs. Breast Cancer 2018, 25, 706–716. [Google Scholar] [CrossRef]
- Zumwalt, T.J.; Goel, A. Immunotherapy of Metastatic Colorectal Cancer: Prevailing Challenges and New Perspectives. Curr. Color. Cancer Rep. 2015, 11, 125–140. [Google Scholar] [CrossRef]
- De Vos, L.; Grünwald, I.; Bawden, E.G.; Dietrich, J.; Scheckenbach, K.; Wiek, C.; Zarbl, R.; Bootz, F.; Landsberg, J.; Dietrich, D. The landscape of CD28, CD80, CD86, CTLA4, and ICOS DNA methylation in head and neck squamous cell carcinomas. Epigenetics 2020, 15, 1195–1212. [Google Scholar] [CrossRef]
- Dong, P.; Xiong, Y.; Yue, J.; Hanley, S.J.B.; Watari, H. B7H3 As a Promoter of Metastasis and Promising Therapeutic Target. Front. Oncol. 2018, 8, 264. [Google Scholar] [CrossRef] [Green Version]
- Piao, L.; Yang, Z.; Jin, J.; Ni, W.; Qi, W.; Xuan, Y. B7H4 is associated with stemness and cancer progression in esophageal squamous cell carcinoma. Hum. Pathol. 2018, 80, 152–162. [Google Scholar] [CrossRef]
- Chen, Q.; Wang, J.; Chen, W.; Zhang, Q.; Wei, T.; Zhou, Y.; Xu, X.; Bai, X.; Liang, T. B7-H5/CD28H is a co-stimulatory pathway and correlates with improved prognosis in pancreatic ductal adenocarcinoma. Cancer Sci. 2019, 110, 530–539. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, M.; Herishanu, Y.; Katz, B.-Z.; Dezorella, N.; Sun, C.; Kay, S.; Polliack, A.; Avivi, I.; Wiestner, A.; Perry, C. Lymphocyte activation gene 3: A novel therapeutic target in chronic lymphocytic leukemia. Haematology 2017, 102, 874–882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Chen, Y.; Liu, X.; Zhang, J.; He, X.; Teng, G.; Yu, D. Tim3/Gal9 interactions between T cells and monocytes result in an immunosuppressive feedback loop that inhibits Th1 responses in osteosarcoma patients. Int. Immunopharmacol. 2017, 44, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Deuss, F.A.; Gully, B.S.; Rossjohn, J.; Berry, R. Recognition of nectin-2 by the natural killer cell receptor T cell immunoglobulin and ITIM domain (TIGIT). J. Biol. Chem. 2017, 292, 11413–11422. [Google Scholar] [CrossRef] [Green Version]
- Reha, J.; Katz, S.C. Regional immunotherapy for liver and peritoneal metastases. J. Surg. Oncol. 2017, 116, 46–54. [Google Scholar] [CrossRef]
- Goodwin, T.J.; Shen, L.; Hu, M.; Li, J.; Feng, R.; Dorosheva, O.; Liu, R.; Huang, L. Liver Specific Gene Immunotherapies Resolve Immune Suppressive Ectopic Lymphoid Structures of Liver Metastases and Prolong Survival. Biomatherials 2017, 141, 260–271. [Google Scholar] [CrossRef]
- Choi, Y.J.; Park, S.-J.; Park, Y.-S.; Park, H.S.; Yang, K.; Heo, K. EpCAM peptide-primed dendritic cell vaccination confers significant anti-tumor immunity in hepatocellular carcinoma cells. PLoS ONE 2018, 13, e0190638. [Google Scholar] [CrossRef] [Green Version]
- Ko, A.H. Progress in the Treatment of Metastatic Pancreatic Cancer and the Search for Next Opportunities. J. Clin. Oncol. 2015, 33, 1779–1786. [Google Scholar] [CrossRef]
- Strebhardt, K.; Ullrich, A. Targeting polo-like kinase 1 for cancer therapy. Nat. Rev. Cancer 2006, 6, 321–330. [Google Scholar] [CrossRef]
- Burdelya, L.G.; Brackett, C.M.; Kojouharov, B.; Gitlin, I.I.; Leonova, K.I.; Gleiberman, A.S.; Aygun-Sunar, S.; Veith, J.; Johnson, C.; Haderski, G.J.; et al. Central role of liver in anticancer and radioprotective activities of Toll-like receptor 5 agonist. Proc. Natl. Acad. Sci. USA 2013, 110, E1857–E1866. [Google Scholar] [CrossRef] [Green Version]
- Tallerico, R.; Todaro, M.; Di Franco, S.; Maccalli, C.; Garofalo, C.; Sottile, R.; Palmieri, C.; Tirinato, L.; Pangigadde, P.; La Rocca, R.; et al. Human NK Cells Selective Targeting of Colon Cancer–Initiating Cells: A Role for Natural Cytotoxicity Receptors and MHC Class I Molecules. J. Immunol. 2013, 190, 2381–2390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, C.; Han, M.; Heinrich, B.; Fu, Q.; Zhang, Q.; Sandhu, M.; Agdashian, D.; Terabe, M.; Berzofsky, J.A.; Fako, V.; et al. Gut microbiome–mediated bile acid metabolism regulates liver cancer via NKT cells. Science 2018, 360, eaan5931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghatalia, P.; Nagarathinam, R.; Cooper, H.; Geynisman, D.M.; El-Deiry, W.S. Mismatch repair deficient metastatic colon cancer and urothelial cancer: A case report of sequential immune checkpoint therapy. Cancer Biol. Ther. 2017, 18, 651–654. [Google Scholar] [CrossRef] [PubMed]
- Toyoshima, Y.; Kitamura, H.; Xiang, H.; Ohno, Y.; Homma, S.; Kawamura, H.; Takahashi, N.; Kamiyama, T.; Tanino, M.; Taketomi, A. IL6 Modulates the Immune Status of the Tumor Microenvironment to Facilitate Metastatic Colonization of Colorectal Cancer Cells. Cancer Immunol. Res. 2019, 7, 1944–1957. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; Wei, W.; Zhao, Q. B7-H3, a checkpoint molecule, as a target for cancer immunotherapy. Int. J. Biol. Sci. 2020, 16, 1767–1773. [Google Scholar] [CrossRef] [Green Version]
- Bagheri, V.; Abbaszadegan, M.R.; Memar, B.; Motie, M.R.; Asadi, M.; Mahmoudian, R.A.; Gholamin, M. Induction of T cell-mediated immune response by dendritic cells pulsed with mRNA of sphere-forming cells isolated from patients with gastric cancer. Life Sci. 2019, 219, 136–143. [Google Scholar] [CrossRef]
- Wefers, C.; Schreibelt, G.; Massuger, L.F.A.G.; De Vries, I.J.M.; Torensma, R. Immune Curbing of Cancer Stem Cells by CTLs Directed to NANOG. Front. Immunol. 2018, 9, 1412. [Google Scholar] [CrossRef]
- Hwang, J.-E.; Shim, H.-J.; Park, Y.-K.; Cho, S.-H.; Bae, W.-K.; Kim, D.-E.; Kim, K.-K.; Chung, I.-J. Intravenous KITENIN shRNA Injection Suppresses Hepatic Metastasis and Recurrence of Colon Cancer in an Orthotopic Mouse Model. J. Korean Med. Sci. 2011, 26, 1439–1445. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Lin, C.; Liao, G.; Liu, S.; Ding, J.; Tang, F.; Wang, Z.; Liang, X.; Li, B.; Wei, Y.; et al. MicroRNA-506 suppresses tumor proliferation and metastasis in colon cancer by directly targeting the oncogene EZH2. Oncotarget 2015, 6, 32586–32601. [Google Scholar] [CrossRef] [Green Version]
- Al-Haidari, A.; Algaber, A.; Madhi, R.; Syk, I.; Thorlacius, H. MiR-155-5p controls colon cancer cell migration via post-transcriptional regulation of Human Antigen R (HuR). Cancer Lett. 2018, 421, 145–151. [Google Scholar] [CrossRef]
- Zhu, W.; Luo, X.; Fu, H.; Liu, L.; Sun, P.; Wang, Z. MiR-3653 inhibits the metastasis and epithelial-mesenchymal transition of colon cancer by targeting Zeb2. Pathol. Res. Pract. 2019, 215, 152577. [Google Scholar] [CrossRef] [PubMed]
- Kamimura, K.; Yokoo, T.; Abe, H.; Terai, S. Gene Therapy for Liver Cancers: Current Status from Basic to Clinics. Cancers 2019, 11, 1865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chawla, S.P.; Bruckner, H.; Morse, M.A.; Assudani, N.; Hall, F.L.; Gordon, E.M. A Phase I-II Study Using Rexin-G Tumor-Targeted Retrovector Encoding a Dominant-Negative Cyclin G1 Inhibitor for Advanced Pancreatic Cancer. Mol. Ther. Oncolytics 2019, 12, 56–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, Q.; Guo, S.; Qian, Z.; Chen, X. Development of individualized anti-metastasis strategies by engineering nanomedicines. Chem. Soc. Rev. 2015, 44, 6258–6286. [Google Scholar] [CrossRef] [Green Version]
- Arshad, U.; Sutton, P.A.; Ashford, M.B.; Treacher, K.E.; Liptrott, N.J.; Rannard, S.P.; Goldring, C.E.; Owen, A. Critical considerations for targeting colorectal liver metastases with nanotechnology. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnol. 2019, 12, e1588. [Google Scholar] [CrossRef] [Green Version]
- Rychahou, P.; Haque, F.; Shu, Y.; Zaytseva, Y.; Weiss, H.L.; Lee, E.Y.; Mustain, W.; Valentino, J.; Guo, P.; Evers, B.M. Delivery of RNA Nanoparticles into Colorectal Cancer Metastases Following Systemic Administration. ACS Nano 2015, 9, 1108–1116. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.; Chen, L.; Liu, J.; Dai, B.; Xu, G.; Shen, G.; Luo, Q.; Zhang, Z. Immune modulation of liver sinusoidal endothelial cells by melittin nanoparticles suppresses liver metastasis. Nat. Commun. 2019, 10, 1–14. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Biomarker | Characteristic | Reference |
|---|---|---|
| Serum amyloid A (SAA) | -Main acute phase proteins expressed in the liver. -Circulating levels correlate with cancer progression and poor survival. -SAA3 in PDAC is associated with fewer differentiated tumors, greater migration, and the number of CD133+ CSCs. | [101] |
| Thrombospondin-2 | -Detection of liver metastases. -Monitor therapy response. | [101] |
| DPC4 | -Prognostic marker in PDAC and CRC. -Promotes EMT. -DPC4 loss is associated with metastatic disease. | [101] |
| ASPH | -Prognostic factor of PDAC. -Upregulated in metastatic PDAC. | [101] |
| Oncostatin M (OSM) | -Elevated in the serum of PC patients. -Associated with highly aggressive metastatic cancers, increased risk of tumor recurrence, and poor prognosis. | [101] |
| Non-alcoholic fatty liver disease fibrosis score (NFS) | -Includes age, diabetes/hyperglycemia, BMI, platelet count, albumin, and AST/ALT ratio. -Predicts survival in liver metastases. | [101] |
| Carcinoembrionary antigen (CEA) | -Predicts survival in liver metastasis during treatment. | [101] |
| Treatment | Advantages | Limitations | Reference |
|---|---|---|---|
| Surgical resection | -Gold standard for liver metastases. -5-year survival of 33% in CRC liver metastases. -5-year survival is 27% in PDAC liver metastases. -Surgical mortality rate is <5%. The 5-year survival of 43% (21–62%) in PC lung metastases. -Better outcome in PDAC (40%) lung metastases. | -Limited liver resection. -Relapse occurred in 75% of patients in the first 2 years after surgery. -Liver resection is not available for all patients in PC. -5-year survival of 19.8% in PDAC lung multiple metastases. | [21,100,101,106] |
| Thermal ablation | -5-year overall survival of 19.9–70% in CRC lung metastases and 25–55% in liver metastases. -Minimal invasiveness, safety, equivalent local control, and survival to lung resection. | -More suitable for small-volume tumors (diameter ≤ 3 cm). -Aiming at a tumor-free margin of > 10 mm. -Recurrence of 46% after thermal ablation. | [107,108,109] |
| Regional hepatic chemotherapy | -Overall survival of 6 to 12 months. -Alternative for patients unable to hepatic resection or those with poor prognostic features. -Allows chemotherapy administration for a longer time. -10–20% of unresectable patients become resectable with chemotherapy | -70% of patients have residual metastases. -Pump placement by surgery. | [100,110,111] |
| Radiation | -Reduction in CEA. -Tumor volumes decrease in liver metastases. -Option for unresectable disease and for medically inoperable patients. -Survival is 70%, 46%, and 46% at 6, 12, and 18 months using selective internal radiation therapy (SIRT). -Local controlled disease is >70% using stereotactic body radiation therapy (SBRT). | -Requires chemotherapy for better response. -Tumor diameter of less than 6 cm. | [112,113] |
| Molecule | Characteristics | Reference |
|---|---|---|
| ICOS (CD28)/ICOSL | -ICOS: ICOSL pathway provides key positive second signals that promote T cell activation, differentiation and effector responses, and T cell-dependent B cell responses. | [115] |
| CD27/CD70 | -Ligation of CD27 by CD70 induces strong ubiquitination of TRAF and the activation of both canonical and non-canonical nuclear factor-kB (NF-kB) pathways. -Reduced regulatory CD4 T-cell numbers -CD70 is expressed in most primary human breast carcinomas and that its expression selectively correlates with lung metastasis. | [116,117] |
| GITR/GITRL | GITR enhances T-cell proliferation and that the absence of GITR is protective in several inflammatory disease models, which is attributed to an impaired effector T-cell function of T cells | [116] |
| PD-1/PDL-1 (PD-L2) | Members of CD28 and B7 families Induce tolerance by regulation of T-cell | [118] |
| BTLA/HVEM | BTLA may specifically downregulate Th1-mediated inflammatory responses Exerts inhibitory effects on B and T lymphocytes | [115] |
| CTLA4/CD80 (CD86) | An inhibitory, co-stimulatory molecule which interferes with the process of T cell activation Natural ligands of CTLA-4 are CD80 (B7-1) and CD86 (B7-2), which are both expressed by antigen-presenting cells (APCs). | [119] |
| B7H3 | Tumor cell immune evasion promotes angiogenesis by upregulating VEGFA | [120] |
| B7H4 | molecular biomarker associated with tumor progression and prognosis | [121] |
| B7H5/HVEM | Sub-expression of B7H5 is correlated with metastases and poor prognosis | [122] |
| LAG3/MHC II | Negatively regulates T-cell function, contributing to tumor escape. | [123] |
| TIM3/GAL9 | Associated with immunosuppression and worse clinical outcome in multiple cancers. | [124] |
| TIGIT/Nectin-2 | Inhibition of NK cell activation. | [125] |
| Indoleamine 2,3-dioxygenase (IDO) | Tolerance and suppressing T cell responses to MHC mismatched allografts, tumors, and self-antigens. | [115] |
| Strategy | Drug | Advances | Clinical Trials | Reference |
|---|---|---|---|---|
| Anti-PD-L1 | Nivolumab Pembrolizumab | -Nivolumab has an objective response rate of 31% in metastatic CRC. -Pembrolizumab has a response rate of 40%. | NCT03307603 | [118,134] |
| NCT03832621 | ||||
| NCT04030260 | ||||
| NCT04575922 | ||||
| NCT02834052 | ||||
| NCT03265080 | ||||
| Atezolizumab Durvalumab Avelumab | -Suppress metastatic colonization on CRC cells. -Reduction of CEA levels. | NCT03721653 | [118,134,135] | |
| NCT03256344 | ||||
| NCT03555149 | ||||
| NCT03193190 | ||||
| NCT03435107 | ||||
| NCT02734160 | ||||
| NCT03563144 | ||||
| GM-CSF Vaccine | GVAX | -Enhance T cell proliferation and secretion of IFN γ and IL-2. -Overall survival of 2.3 and 4.3 months. -Less toxic effects. | NCT01417000 | [118,126] |
| NCT00727441 | ||||
| NCT02004262 | ||||
| Anti-B7H3 | Monoclonal antibody (3E8) and bevacizumab MGC018 DS-7300a | -Reduce CSC number. -Inhibits tumor growth and angiogenesis. -Depletion of cancer-associated fibroblasts. | NCT03729596 | [120,136] |
| NCT04145622 | ||||
| Dendritic Cells (DC) Vaccines | DC loaded with lysates of CSC Peptides: CD90, CD54, CD44, EpCAM, Panc-1, ALDH, mRNA | -Induction of MHC expression, cytokine production (IFN γ), lymphocyte infiltration, proliferation T, and B lymphocytes. | NCT02615574 | [128,130,137] |
| NCT02503150 | ||||
| NCT00176761 | ||||
| NCT00558051 | ||||
| NCT00868114 | ||||
| NCT01410968 | ||||
| Mesothelin Vaccine | Live-attenuated Listeria monocytogenes vaccine | -Activates mesothelin T cells in PC. | NCT03122106 | [129] |
| NCT03956056 | ||||
| DC Expressing CSC Transcription Factors | NANOG, OKT4a, SOX2, c-MYC, KLF4 | -Cellular immunological memory. | NCT00103142 | [138] |
| Natural Killer Cells | Recognition of receptors in CSC: NKp30, NKp44 | -Targeting of CSC. | NCT03008499 | [131] |
| CAR-T Cells | Chimeric antigen receptors for specific proteins Anti-mesothelin Anti-CEA | -Increase in cell death. -Serum CEA decrease. | NCT01897415 | [129] |
| NCT01583686 | ||||
| NCT02416466 |
| miRNA | Disease | Characteristics | |
|---|---|---|---|
| miR-93 | Pancreatic adenocarcinoma and lung metastases | Regulates microtubule dynamics by controlling YES1, CRMP2, and MAPRE1 expression. | [39,71] |
| miR-146a | Pancreatic adenocarcinoma and lung metastases | Diminishes in metastasis. | [72,73] |
| miR-200 family members: miR-194, miR-200b, miR-200c, and miR-429 | Pancreatic adenocarcinoma | EMT facilitating invasion, targeting PTEN, EP300, and TGF-b/SMAD pathways. | [72,73,74] |
| miR-141 and miR-429 | Pancreatic adenocarcinoma | Inhibit tumor development. | [75,76,77,78] |
| miR-10b | Pancreatic adenocarcinoma | Increases invasiveness in metastasis by the effect of EGF and TGF-β signaling. | [82,83,84] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Quiroz-Reyes, A.G.; Islas, J.F.; Delgado-Gonzalez, P.; Franco-Villarreal, H.; Garza-Treviño, E.N. Therapeutic Approaches for Metastases from Colorectal Cancer and Pancreatic Ductal Carcinoma. Pharmaceutics 2021, 13, 103. https://doi.org/10.3390/pharmaceutics13010103
Quiroz-Reyes AG, Islas JF, Delgado-Gonzalez P, Franco-Villarreal H, Garza-Treviño EN. Therapeutic Approaches for Metastases from Colorectal Cancer and Pancreatic Ductal Carcinoma. Pharmaceutics. 2021; 13(1):103. https://doi.org/10.3390/pharmaceutics13010103
Chicago/Turabian StyleQuiroz-Reyes, Adriana G., Jose F. Islas, Paulina Delgado-Gonzalez, Hector Franco-Villarreal, and Elsa N. Garza-Treviño. 2021. "Therapeutic Approaches for Metastases from Colorectal Cancer and Pancreatic Ductal Carcinoma" Pharmaceutics 13, no. 1: 103. https://doi.org/10.3390/pharmaceutics13010103