Antioxidant Defenses in the Human Eye: A Focus on Metallothioneins
by
 , , ,
, , ,
Ana Álvarez-Barrios
1,2 ,
,
Lydia Álvarez
1 ,
,
Montserrat García
1,3,
Enol Artime
1,
Rosario Pereiro
1,2 and
Héctor González-Iglesias
1,3,* 1
Instituto Universitario Fernández-Vega (Fundación de Investigación Oftalmológica, Universidad de Oviedo), 33012 Oviedo, Spain
2
Department of Physical and Analytical Chemistry, Faculty of Chemistry, University of Oviedo, Julián Clavería, 8, 33006 Oviedo, Spain
3
Instituto Oftalmológico Fernández-Vega, Avda. Dres. Fernández-Vega, 34, 33012 Oviedo, Spain
*
Author to whom correspondence should be addressed.
Antioxidants 2021, 10(1), 89; https://doi.org/10.3390/antiox10010089
Submission received: 18 December 2020
/
Revised: 3 January 2021
/
Accepted: 4 January 2021
/
Published: 11 January 2021
(This article belongs to the Special Issue Antioxidants and Age-Related Ocular Diseases)
Abstract
:The human eye, the highly specialized organ of vision, is greatly influenced by oxidants of endogenous and exogenous origin. Oxidative stress affects all structures of the human eye with special emphasis on the ocular surface, the lens, the retina and its retinal pigment epithelium, which are considered natural barriers of antioxidant protection, contributing to the onset and/or progression of eye diseases. These ocular structures contain a complex antioxidant defense system slightly different along the eye depending on cell tissue. In addition to widely studied enzymatic antioxidants, including superoxide dismutase, glutathione peroxidase, catalase, peroxiredoxins and selenoproteins, inter alia, metallothioneins (MTs) are considered antioxidant proteins of growing interest with further cell-mediated functions. This family of cysteine rich and low molecular mass proteins captures and neutralizes free radicals in a redox-dependent mechanism involving zinc binding and release. The state of the art of MTs, including the isoforms classification, the main functions described to date, the Zn-MT redox cycle as antioxidant defense system, and the antioxidant activity of Zn-MTs in the ocular surface, lens, retina and its retinal pigment epithelium, dependent on the number of occupied zinc-binding sites, will be comprehensively reviewed.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
The eye is subjected to a highly oxidative environment due to its intense exposure to light, its robust metabolic activity, and its high oxygen tension. Considering that solar ultraviolet (UV) radiation is the major environmental inducer of oxidant reactive species formation, the eye has developed a complex protection system against oxidative damage organized in three levels: (1) Prevention of free radical yield by pigments capable of absorbing and filtering light; (2) detoxification of free radicals by enzymatic or non-enzymatic antioxidants; and (3) repairment systems of oxidized biomolecules [1]. In the following sections, the antioxidant enzymes that are part of the defense system of the human eye will be discussed, with a particular focus on the redox system zinc-metallothioneins.
1.1. The Human Eye
The human eye is a highly specialized organ of the visual system consisting of three primary layers surrounding the bulk of the ocular globe (Figure 1). The external part is the fibrous tunic, containing the sclera and cornea, the middle part is the uvea or vascular tunic composed of iris, ciliary body (CB), and choroid, and the inner part is the neural layer formed by the retina and its retinal pigment epithelium (RPE). These coats surround the lens and the transparent media, namely anterior chamber with aqueous humor and vitreous body [1]. Oxidative stress affects all the structures of the eye, specifically its antioxidant natural barriers formed by the ocular surface (i.e., cornea and the anterior part of the sclera), the lens and retina.
The cornea and sclera form a protective envelope that protects the ocular tissues and also provides structural support. The transparency of the cornea and its refractive power are among the most important properties, presenting a tough physical barrier to trauma and infection. The cornea has five different layers: The corneal epithelium, a stratified, squamous, non-keratinized epithelium; the anterior limiting membrane or Bowman’s layer, a modified acellular region of the stroma; the corneal stroma, a connective tissue composed of collagenous lamellae, keratocytes and modified fibroblasts; posterior limiting lamina or Descemet’s membrane; and corneal endothelium, a single layer of hexagonal cells. The lens, also responsible for light refraction and focus over the retina, possesses less refractive power than the cornea. This transparent body is composed of long-fibre like cells and epithelial cells highly organized and tightly packed, enclosed in an elastic collagenous capsule that confers its ability to change shape by the accommodation process. The retina is a light-sensitive tissue that lines the inner surface of the posterior segment of the eye, whose main function is to detect light, convert photochemical energy into neural signals, and transmit the signals to the visual cortex of the brain. The retina consists of an inner multilayer of neurosensory cells and an outer single neuroepithelium, the RPE. The neuroretina comprises the nerve fiber layer, the ganglion cell layer, the inner plexiform layer, the inner nuclear layer (INL), the outer plexiform layer and the outer nuclear layer (ONL), formed by different cell types including vascular endothelial, pericytes, glial, microglia, ganglion, horizontal, amacrine, bipolar, and photoreceptors. In addition, the RPE is a pigmented cell monolayer that nourishes and maintains the adhesion of the neurosensory retina and reduces light scattering within the eye [2].
1.2. Reactive Oxygen Species within the Eye
Oxidative stress occurs from formation of multiple reactive oxygen species (ROS), including superoxide, hydrogen peroxide, and hydroxyl radicals, which promote free radical production. These oxygen-containing free radicals are produced as natural subproducts of the normal metabolism of oxygen, and have important roles in cell signaling and homeostasis. An imbalance of ROS production and the capability of the eye to counteract these free radicals causes oxidative stress that may be amplified by a continuing cycle of metabolic stress leading to increased free radical production. It compromises the antioxidant capacity of free radical scavenger systems and triggers cell damage and death. This intracellular disequilibrium results in the onset and/or progression of ocular diseases, which is exacerbated during long-term oxidative stress exposure and ageing [3].
While exogenous ROS generators include UV light exposure, viral infection, chemical insults and drug intake, among others, endogenous ROS are mainly produced in the mitochondria due to cellular respiration [4]. UV light, one of the main factors of ROS production triggering oxidative damage, consists of radiation in different wavelengths: UVA (315–400 nm), UVB (280–315 nm) and UVC (100–280 nm). Whilst the cornea and lens absorb UVC radiation and most UVB, a small part of UVA radiation reaches the retina-RPE system (see Figure 1). This continuous oxidative damage plays a key role in the onset of age-related eye diseases affecting the cornea, lens and retina-RPE [5]. In the cornea, UV light producing ROS interferes with epithelial cells proliferation and reduces epithelial thickness, which may promote Fuchs’ endothelial dystrophy. This corneal disorder associated to oxidative stress leads to degeneration of endothelium causing stromal and epithelial edema resulting in visual acuity reduction. This corneal dystrophy is characterized by the loss of hexagonal shape and cell density of endothelial cells and formation of excrescences of Descemet’s membrane called guttae [6]. Besides, the lack of blood vessels in the lens exacerbates oxidative stress, increasing the risk of its opacification and resulting in cataract formation, which is one the most common causes of blindness worldwide [7,8,9]. Furthermore, the high oxygen consumption rate of retinal cells due to cellular respiration contributes synergistically with UV light and vascular flux defects to the pathogenesis of retinal diseases, including glaucoma, age-related macular degeneration (AMD), diabetic retinopathy and retinitis pigmentosa, among others [10]. During glaucoma, a neuropathy characterized by the abnormal increase of intraocular pressure that brings to degeneration of retinal ganglion cells (RGCs) causing irreversible blindness, a prolonged imbalance of oxidative species and antioxidants is considered a significant risk factor where high intraocular pressure stimulates in a feedback process the production of ROS, inducing RGCs to autophagy and apoptosis [11]. Moreover, the continuous exposure of the RPE to light energy, the oxygen-rich environment and the high metabolic activity of this neuroepithelial monolayer provide an ideal framework for the formation of ROS with the potential to damage proteins, DNA and lipids, resulting in AMD, a neurodegenerative disease, characterized by the irreversible damage in the macula and the formation of extracellular deposits between RPE and Bruch’s membrane (a modified connective tissue layer), called drusen [12]. Finally, the microvascular disease diabetic retinopathy is a complication of diabetes mellitus, caused by high mitochondrial ROS production, due to hyperglycemia. This results in capillaries cell damage and the release of plasma and erythrocytes in the retina causing edema and stimulating retinal angiogenesis [13].
2. Antioxidant Defense Systems in the Eye
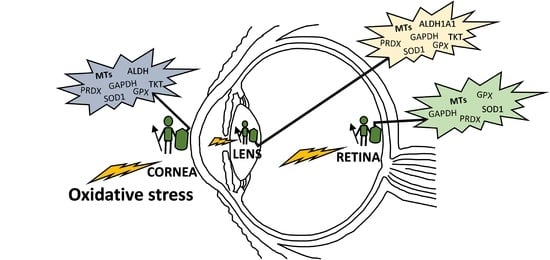
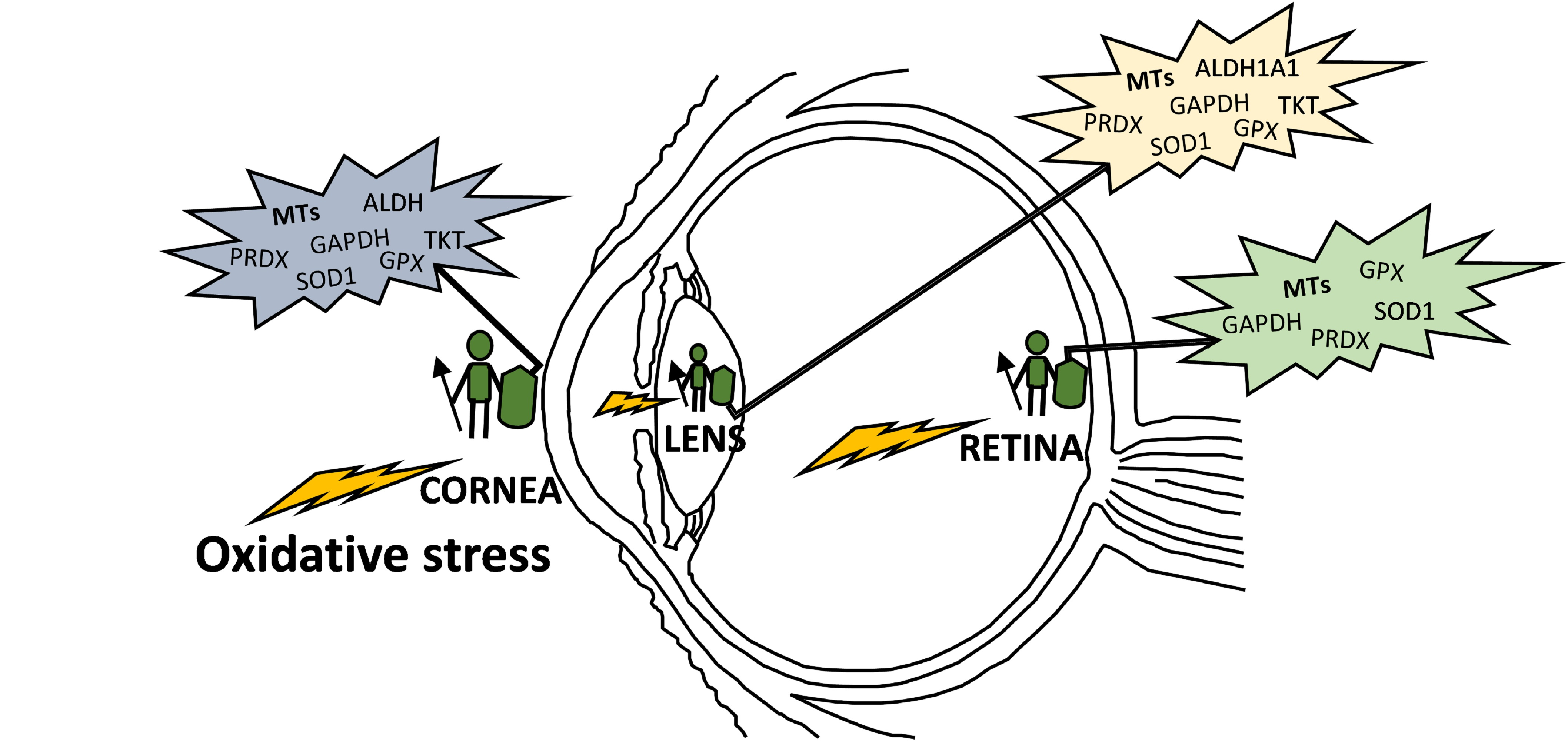
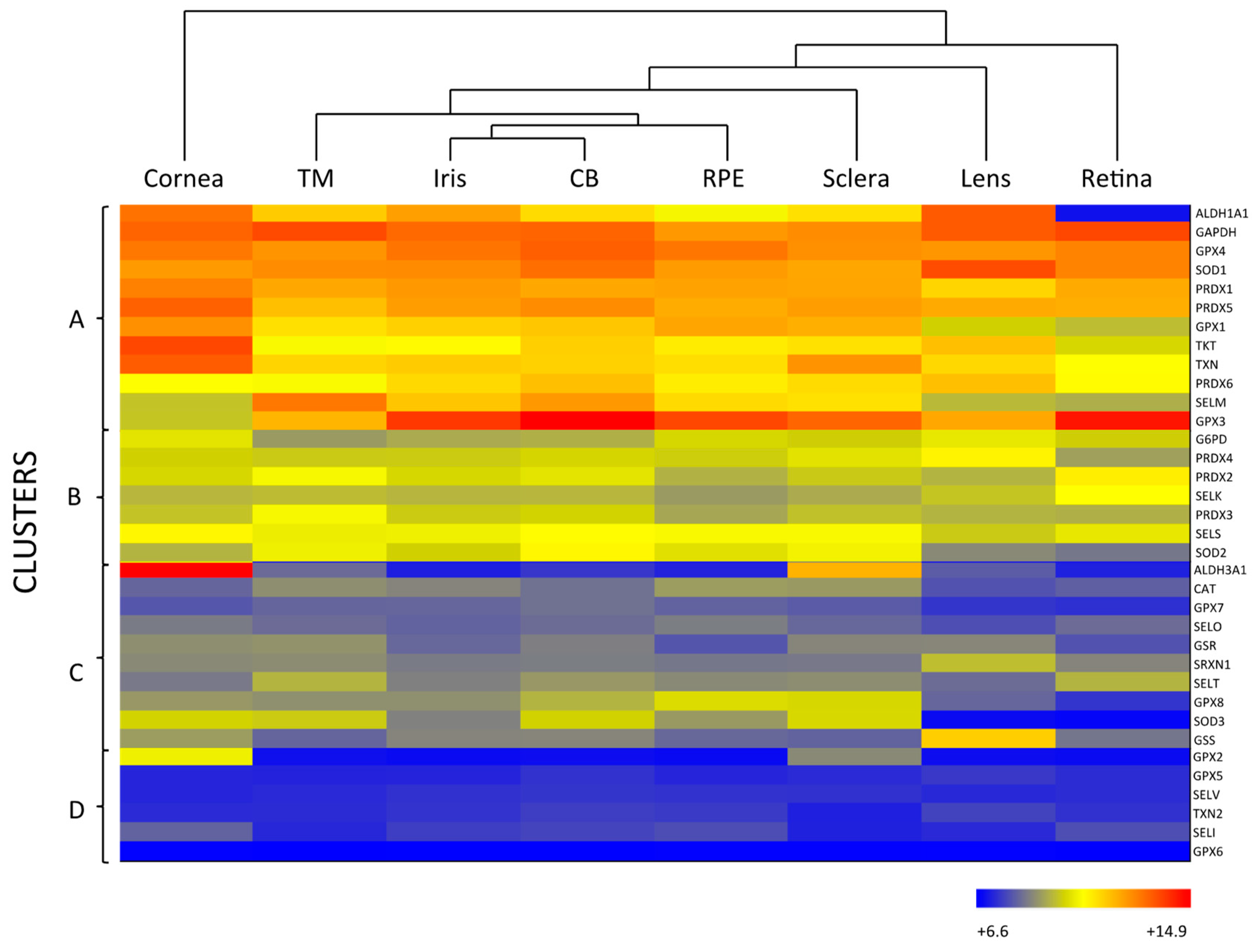
As mentioned above, the eye is constantly exposed to both exogenous and endogenous sources of oxidants, which put the cells under continuous pressure from oxidative stress. The eye relies on a complex antioxidant defense system to keep free radicals under control and maintain correct physiological functions [5]. Defense against oxidative stress is slightly different across the eye, matching with its structural complexity and accounting for the different sources of ROS threatening each region. For the purpose of this review, we have divided the eye into three natural barriers of defense: (1) The ocular surface, composed by the tear film, cornea, anterior sclera and aqueous humor; (2) the lens; and (3) the retina (neurosensory retina and RPE). Although it is out of the scope of this review, it should be noted the potential antioxidant role of the vitreous body, a body fluid of gel nature filling the posterior pole between the lens and the retina, filtering infrared light, has been recently reviewed [14]. Figure 2 shows the hierarchical ranking and clustering of enzymatic antioxidant genes present in the human eye, based on whole-genome expression microarray analysis by Alvarez et al., 2012 [15]. Four clusters could be distinguished (A, B, C and D), each containing genes expressed at levels ranging from highly abundant (i.e., cluster A) to moderate (i.e., cluster B and C), and almost absent (cluster D), which will be further discussed with respect to their ocular tissue antioxidant barrier function.
The antioxidant defense system of any cell is composed of a vast variety of compounds, including both enzymatic and non-enzymatic molecules. Non-enzymatic antioxidants are water-soluble (e.g., ascorbic acid and glutathione) or fat-soluble (e.g., carotenoids and vitamins A and E) molecules with relevant roles, not only in antioxidant protection, but also in UV-light absorption [16,17], by which other scientific papers are referenced [18,19,20]. In relation to enzymatic antioxidants, primary and secondary enzymatic defenses can be distinguished. Primary enzymatic antioxidants consist of enzymes that act at the root of the oxidative propagation chain, preventing the formation of new free radicals by removing precursors or inhibiting catalysts. These primary antioxidants include enzymes like superoxide dismutase (SOD), glutathione peroxidase (GPX), catalase (CAT) and peroxiredoxins (PRDXs). By comparison, secondary enzymatic antioxidants have preventive and supportive roles in antioxidant protection and may tackle different processes like metal deactivation, regeneration of primary antioxidants and maintenance of the cellular reducing power. Secondary antioxidants include metabolic enzymes like transketolase (TKT), glucose-6-phosphate dehydrogenase (G6PD) and glyceraldehyde-3-phosphate dehydrogenase (GAPDH); glutathione-redox cycle enzymes like glutathione reductase (GSR) and glutathione synthetase (GSS); selenoproteins (SELM, SELO, SELT, SELK, SELS, SELV and SELI) and thioredoxin-related enzymes like thioredoxin (TXN) and sulfiredoxin (SRXN) [21,22]. Both primary and secondary antioxidants are important constituents of the ocular surface, the lens, and the retina of the human eye.
2.1. Enzymatic Antioxidants of the Ocular Surface
As the outermost region of the eye, the ocular surface comprised of the cornea, sclera, tear film and aqueous humor, is directly exposed to environmental damaging agents. Considering that UV-light radiation is the major source of ROS among these environmental stressors, the ocular surface has a crucial function in UV-light filtering and absorption, protecting the inner ocular tissues from oxidative damage and constituting the first barrier of defense against oxidants [19,20]. However, UV-light absorption makes the cells of the cornea and anterior sclera themselves vulnerable to the effects of ROS, thus, requiring a robust antioxidant defense system optimized for both their function and protection [23].
2.1.1. Primary Antioxidant Enzymes
SOD catalyzes the dismutation of superoxide anion (O2·) to the less toxic free radical hydrogen peroxide (H2O2), being the only antioxidant, capable of reacting with O2·, and thus, often regarded as the most important primary antioxidant of the corneal antioxidant defense system [5]. Humans have three isoforms of SOD, namely SOD1, SOD2 and SOD3, that differ in their location (SOD1 is located in the cytosol, SOD2 in the mitochondria and SOD3 in the extracellular space) and cofactors (SOD1 and SOD3 use Cu and Zn, while SOD2 uses Mn) [16]. SOD isoforms also have different sensitivity to UV-light and oxidative stress, given that SOD1 is easily down-regulated by UVB, while SOD2 can be moderately induced under light-induced oxidative stress [24]. Due to their differences, some authors suggest that SOD isoforms perform different functions in the eye physiology. This hypothesis is supported by the fact that knockout animals show significantly different phenotypes depending on the isoform inactivated [19,25]. According to Figure 2, all three SOD isoforms are present in the ocular surface, being SOD1 the most expressed isoform in all the considered eye tissues, including the cornea. While SOD2 is a minor isoform of SOD in the human cornea, it is abundant in other mammalian ocular tissues and has gained interest because of its antioxidant function within the mitochondria, and its role in the prevention of apoptosis initiated by mitochondrial damage [26,27].
Although H2O2 is more stable than O2·, it can diffuse across hydrophobic membranes and generate other ROS such as OH- by reacting with metal ions. Therefore, H2O2 poses an important threat to cell function and numerous enzymes are committed to its detoxification, i.e., CAT, GPX and PRDX [28,29]. CAT is a heme-containing enzyme that catalyzes the dismutation of H2O2 into O2 and H2O using Mn as a cofactor. It also protects SOD from inactivation and may be directly involved in UVB-light filtering. CAT is ubiquitously expressed in mammalian cells, predominantly in peroxisomes [19,25], and the microarray analysis data depicted in Figure 2 suggest that CAT is expressed at low levels both in the cornea and sclera.
GPX catalyzes the same reaction as CAT, reducing H2O2 to H2O, but using reduced glutathione (GSH) as an electron donor. GPX activity depends on the regeneration of GSH, which is carried out by enzymes like GSR and GSS that produce GSH by reducing oxidized glutathione (glutathione disulfide, GSSG) or by de novo synthesizing it, respectively [5,19]. Eight isoforms of GPX are known in humans and five of them (GPX1–4 and GPX6) require Se as cofactor [25]. Like other selenoproteins, GPX family follows a hierarchical order for Se supply, being GPX1 and GPX4 the most preferentially isoforms [30]. Six isoforms of GPX are expressed in the ocular surface, with preferential presence of GPX1 and GPX4, in addition to GPX3 in the sclera (Figure 2). In comparing GPX and CAT expression within the eye, it seems that GPX rather than CAT may predominantly perform H2O2 decomposition in the cornea and sclera. Conversely, CAT may gain importance over the GSH redox system when H2O2 concentration increases [20,31].
PRDX is the third type of enzyme able to reduce and detoxify H2O2 using its redox active structural cysteines without the need of specific cofactors [14]. Six PRDX isoforms are expressed in humans, namely PRDX1-6, all localized both in the cornea and the sclera. PRDX may also have chaperone-like and redox-signaling properties, which link them to inflammation and aging processes [28,32].
2.1.2. Secondary Antioxidant Enzymes
A big part of the secondary antioxidant enzymes of the cornea belongs to a group of water-soluble proteins formerly called crystallins, which include very diverse proteins, like chaperones, metabolic enzymes, and members of the heat-shock family. Berzelius originally coined the name in 1830 to describe highly abundant proteins present in the bovine crystal-clear lens [33]. Later, it was found that crystallins are also present in the cornea and, less abundantly, in some outer-eye tissues [34]. In the eye, their main function is to provide the lens and cornea with the required transparency and refractive index to filter and focus the light adequately. These proteins are carefully packed to minimize variations in concentration and to create an appropriate concentration gradient across the eye [33]. Developmental modulators and environmental factors tightly regulate crystallins genes in order to create such concentration gradient [35]. During aging and/or post-translational modifications, crystallins are prone to aggregate, which disturbs their concentration pattern and the light focusing capacity of the eye, even possibly leading to cataracts exacerbated by oxidative stress [33,36,37]. Interestingly, crystallins in the eye also have additional functions involved in UV-light filtering, protein folding and ROS detoxification [24]. The multifunctionality of crystallins is caused by a phenomenon called “gene sharing”, which occurs when a gene codifies a protein that has remarkably different functions depending on its concentration and expression pattern [33]. This finding expanded the knowledge about crystallins by including enzymes that were long thought to only be involved in cellular processes, such as metabolism and stress response [35].
One of such called enzymatic crystallins is the aldehyde dehydrogenase (ALDH) family, which regulates the metabolism of aldehydes and has a function as crystallins, albeit only ALDH1A1 and ALDH3A1 are expressed in some tissues of the eye [38]. In the cornea, ALDH family is the most abundant secondary antioxidant, especially the isoform 3A1 (ALDH3A1), which is the most expressed corneal antioxidant enzyme (Figure 2) accounting for 5 to 50% of the total water-soluble protein fraction of the corneal epithelium in mammalian species [39]. ALDH superfamily catalyzes the NAD(P)+-dependent oxidation of a wide range of endogenous and exogenous toxic aldehydes and participates in the antioxidant defense by directly scavenging ROS and indirectly producing NADPH. Although, ALDH3A1 is considered a cytosolic protein, it has been also found in the cellular nucleus, suggesting that it may also protect DNA from oxidative damage [40]. While ALDH3A1 is almost exclusively expressed in the cornea and sclera, ALDH1A1 is present in all the eye tissues with the exception of the retina, being highly expressed in the cornea ahead of other abundant enzymes like SOD (Figure 2). These findings support the idea that the cornea requires a higher number of antioxidants than other parts of the eye to maintain its UV-light filtering function.
The enzymes TKT and GAPDH are found in great abundance in the cornea—hence, they are recognized as being crystallins—highlighting that TKT is the second most abundant secondary antioxidant enzyme just behind ALDH3A1. TKT metabolizes glycolytic products as part of the pentose phosphate pathway (PPP) and GAPDH is an enzyme involved in the glycolysis [41]. PPP and glycolysis are closely associated routes that generate ribose-5-phosphate, the essential building block for RNA synthesis, and ATP via glucose oxidation, respectively. Both pathways also generate NADPH, which is the main reducing agent of the cell, so enzymes like TKT and GAPDH serve an additional antioxidant role by promoting NADPH production [24].
It should be noted that the cornea counts with other crystallins that are non-enzymatic, such as chaperones, whose activity is fundamental to prevent protein aggregation and to maintain the transparency of the cornea [34]. In some instances, chaperones also prevent oxidative stress by conserving the structure of antioxidant enzymes, e.g., αB-crystallin reduces aggregation of mutant SOD-1 in familial amyotrophic lateral sclerosis and secures its antioxidant activity [42].
2.2. Enzymatic Antioxidants of the Lens
Together with the cornea, the lens is responsible for focusing the incident light into the retina, as well as filtering harmful UV radiation to protect the inner tissues, constituting a second barrier of antioxidant defense. The UV radiation that reaches the lens is greatly composed of UVA light, since the cornea absorbs UVC and most UVB but only a small fraction of UVA (see Figure 1). UVA radiation has been shown to be more effective at causing oxidative damage than any other UV radiation, putting the lens at a higher risk of oxidative stress induction since its absorption may cause damage to the cells [43,44]. Also, lens metabolism occurs mainly in the epithelium, while the synthesis of new proteins ceases with lens fiber cell formation, making the lens especially susceptible to the accumulation of oxidative damage, and accounts for its different antioxidant needs in comparison with the aforementioned cornea [28,45,46].
The lens contains high levels of GPX and SOD [1]. Superoxide dismutases are one of the main antioxidant enzymes of the ocular lens, where they show a clear distribution consisting of high levels of protein in the anterior layers of the lens (i.e., lens capsule and epithelial cells) and low levels in the posterior layers (i.e., cortex and nucleus composed of lens fibers) [47]. Although SOD2 and SOD3 may not be present at RNA level (Figure 2). The isoform SOD1 contributes about a 90% to the total SOD activity in the lens [48], whilst the overall SOD activity significantly decreases with age and is probably caused by the life-long accumulation of inhibitory modifications, as protein turnover is almost non-existent in the cortex and the nucleus. In contrast, the activity of SOD in the cornea does not decline to such extent during aging, reflecting the differences in protein synthesis and turnover between these eye tissues [47,49].
Low CAT RNA-expression levels are observed in the whole lens (Figure 2). Some studies show that CAT is more concentrated in the periphery of the lens epithelium, where cells may require a stronger defense system [50]. Recent studies have been centered on the signaling function of H2O2 in the cell, which has been shown to occur by transient bursts of oxidants that inactivate both CAT and PRDXs [51,52]. However, it is most likely that daily H2O2 decomposition in the lens is carried out by PRDXs and GPX, according to their raised abundance in the tissue, and that CAT is more active against chronic exposure to H2O2 similarly to its function in the cornea [18,28]. All PRDX isoforms are, to a greater or lesser extent, expressed in the lens, as well as in the rest of the ocular tissues, suggesting a distinct and specific role in eye physiology for each of them (Figure 2) [53].
The glutathione redox system is especially important in the lens, since its high protein content calls for a strict redox control on thiol groups to avoid protein aggregation and this system can fulfill such function [54]. According to Figure 2, among the eight isoforms of human GPX, the lens mostly expresses GPX1, GPX3 and GPX4. GPX1 is the main intracellular isoform of GPX and its deficiency has been linked with membrane damage in the lenticular nucleus. Neither the epithelium nor the cortex of the lens is significantly affected by GPX1 loss, probably due to the abundance of other compensating antioxidants [55]. In another study, GPX3 is secreted and appears in the extracellular space, while GPX4, a mediator in the non-apoptotic and iron-dependent programmed cell death pathway that produces a rapid release of ROS (i.e., ferroptosis) is coupled with cell membranes [56].
As discussed in the previous section, crystallins are extremely important to the lens due to their dual role in antioxidant protection and light refraction. Among the enzymatic crystallins, members of the ALDH superfamily are the most abundant in the lens, specifically the isoform ALDH1A1, just behind SOD1 levels [38]. In contrast to the cornea, ALDH activity in the lens is carried out entirely by ALDH1A1 and not ALDH3A1. Although, in some species, ALDH1A1 has been described in the cornea at even greater levels than ALDH3A1, the later has never been reported to have significant abundance in the lens [44,57]. This suggests that ALDH1A1 has an important function both in the cornea and the lens, which could be related to the detoxification of reactive aldehydes. Meanwhile, ALDH3A1 may have a bigger role in UV radiation filtering than aldehyde detoxification, since by filtering light in the cornea it also indirectly protects the underlying lens [16]. ALDH1A1 has a heterogeneous distribution throughout the lens, being more concentrated in the epithelial cells and the cortex than in the nucleus, presumably because of its bigger exposure to free radicals coming from the aqueous humor [57].
Finally, metabolic enzymes, involved in NADPH regeneration, like GAPDH and TKT, are also an important part of the lens antioxidant defense system. Differential expression of TKT in the lens and cornea (Figure 2) may be the result of different gene regulatory mechanisms. It is suggested that crystallin gene expression in the lens is mostly regulated by tissue- and developmental-specific transcription factors, while environmental agents may play a more determining role in the expression profile of the cornea [58].
2.3. Enzymatic Antioxidants of the Retina
The retina is the innermost part of the eye and has a crucial role in vision, being the structure that transforms light energy into electrical outputs that are transmitted to the brain. Due to its function in vision, the retina has the highest oxygen consumption rate per kg of the body, and thus, is particularly exposed to ROS of endogenous origin [20]. In addition, photoreceptors contain high concentrations of polyunsaturated lipids that are easily oxidized and further increase the susceptibility of the retina to oxidative stress. Apart from endogenous sources, ROS in the retina can also arise from short-wavelength light that is not filtered by the first and second barriers of defense, specifically UVA and blue visible light. The RPE decreases light scatter and protects against oxidative stress generated by photooxidation. However, light-induced oxidative stress risk increases with age due to antioxidant deficiency and the accumulation of oxidative agents [59], contributing to the onset of age-related eye diseases affecting the retina. The macular region, located near the center of the retina-RPE, is particularly susceptible to light damage, and contains yellow pigments (carotenoids) that reduce glare from short wavelength blue light. These carotenoids, referred to xanthophyll macular pigments, also have an antioxidant role particularly at low levels of oxygenation. In addition, the pigmented RPE contains high amounts of melanin, which in the reduced form is a very effective free radical scavenger [1].
Primary enzymes like SOD, GPX, CAT and PRDXs are the main components of the enzymatic antioxidant defense system of the retina-RPE. The CAT concentration in the retina increases upon light exposure, but its levels do not change during prolonged exposure times. In such cases, SOD and GPX may take over CAT for antioxidant protection. In fact, CAT activity has been found to decrease with age and during age-related pathologies like AMD, while SOD does not show a clear correlation with aging [20]. However, the SOD distribution across the retina seems to change during aging [60], and recently, it was reported that SOD2 isoform experiences an age-related increase in the neural retina and more prominently in the macular region. Although, it should be noted that SOD2 levels in the retina are low in comparison to SOD1 and other antioxidant enzymes [59].
Interestingly, GPX isoform composition in the retina differs from that of the cornea and lens, being GPX3 the most abundant isoform followed by GPX4. Moreover, the GPX3 is the enzymatic antioxidant with the highest expression in the retina (Figure 2). GPX3 is present in the extracellular space, where it may protect the cell surface and basal membranes [61]. It has been reported that GPX3 is present in high levels in the aqueous humor and ciliary body of human and bovine eyes, but its presence and role in the retina has not been deeply studied [62,63]. It should be noted that GPX1 is abundantly expressed in the retina of the immature eye, likely acting as an early defense mechanism [64]. During aging, a similar pattern of expression to SOD has been described for GPX, with up-regulated levels in the photoreceptor outer segments, down-regulated levels on the peripheral retina and increased concentration upon light exposure [65]. In a recent work, the enzyme glyoxalase 1 prevented induced oxidative stress damage through the detoxification of methylglyoxalm, whose excess inactivates GPx and SOD, which may be involved in the etiopathogenesis of retinitis pigmentosa [66].
The PRDX family also contributes to the detoxification of H2O2 in the retina, although it does not show the same relevance as SOD and GPX enzymes. PRDX may complement the SOD/GPX H2O2-detoxification system by acting as redox sensors in the retina, since under oxidative stress they can become hyperoxidized and prompt redox signaling cascades, as well as function as protein chaperones. PRDX show a distinct distribution in the neural retina depending on the cellular type, where PRDX1 is mainly associated with cells in the inner nuclear layer and cone photoreceptors and PRDX5 is ubiquitously expressed in the mitochondria and sometimes found in peroxisomes, the cytoplasm and the nucleus [67].
Other antioxidants like GAPDH and some selenoproteins work in a supportive role to SOD, GPX, PRDX and CAT enzymes. Specifically, GAPDH is the secondary antioxidant most expressed in the retina due to its role in glycolysis and NADPH generation and contributes to the maintenance of the cellular reducing power under oxidative stress. Interestingly up to twenty crystallins have been described in the retina, most of them sparsely expressed and carrying out different functions than those they have in the cornea and lens. It is plausible that some crystallins interact with the cytoskeleton of retinal cells to maintain its structural integrity [68]. The overexpression of crystallins has been found to protect the RPE from oxidative stress, since some of them act as heat-shock proteins and metabolic enzymes involved in NADPH generation [69].
3. Metallothionein Antioxidant System
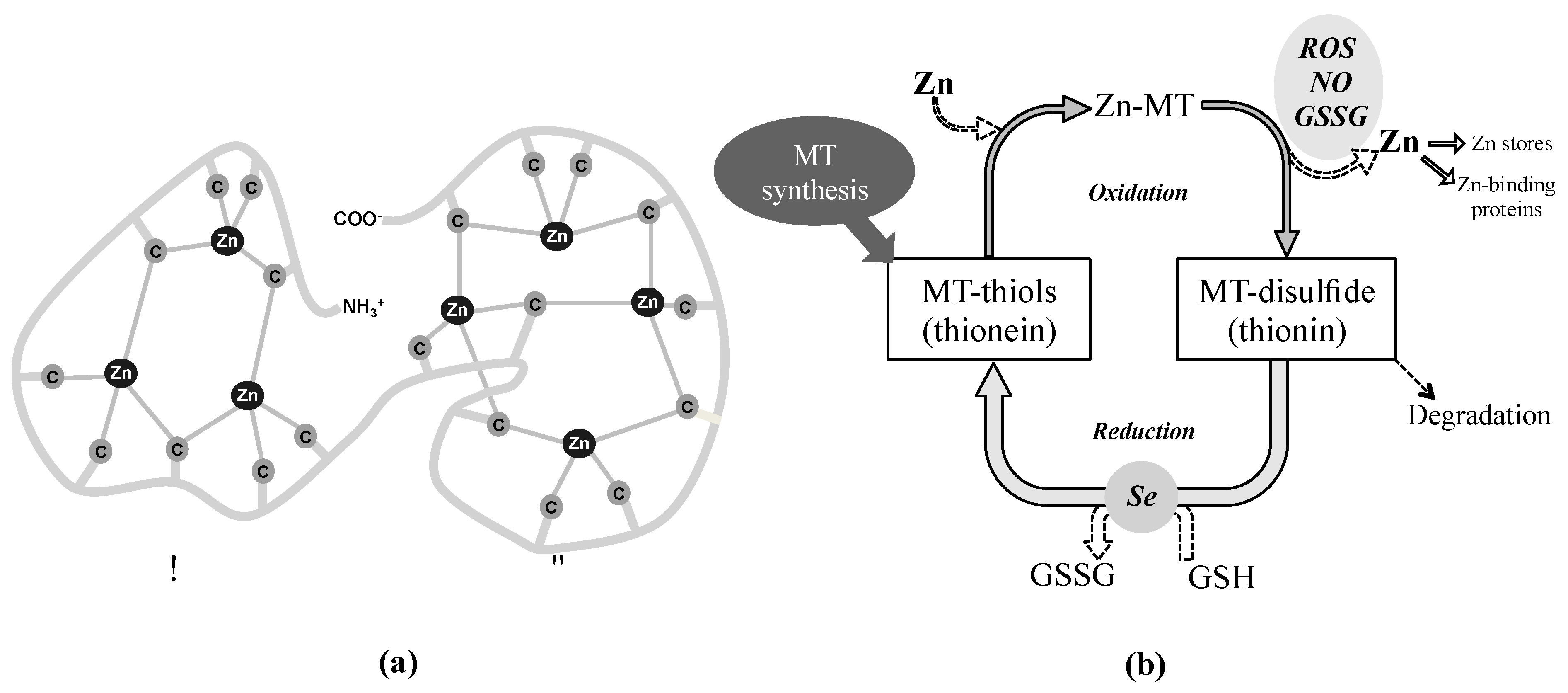
One of the antioxidant systems of growing interest in the eye is constituted by the metallothioneins (MTs) that capture and neutralize free radicals through their cysteine sulfur ligands serving as zinc-ion donors in a redox-dependent fashion. Although it is not classically considered an enzymatic antioxidant system per se, its redox activity relies on the redox-dependent zinc release. The family of MTs is a group of cysteine rich and metal-binding low molecular mass proteins (6–7 kDa) present in all eukaryotes and many prokaryotes [70]. MT molecule is composed of two structural domains with a high content of sulfur and metals, forming metal-thiolate clusters through the covalent binding of metal atoms with the sulfhydryl cysteine residues. As shown Figure 3 (panel a), the N-terminal part of the peptide (β-domain) has three binding sites for divalent ions and the C-terminal part (α-domain) can bind up four divalent metal ions [71]. These proteins bind mainly Zn, Cu, Cd and Hg, while in mammals, under physiological conditions, MTs mostly contain zinc divalent ions, which can be displaced by copper or cadmium if they are in excess [72,73]. The coexistence of stable, partially metalated MTs along with the apo form of the protein and the fully-metalated protein indicate a non-cooperative metalation mechanism [74,75,76].
3.1. General Properties
3.1.1. Classification
In humans, the MT family consists of four groups designated as MT1 to MT4, containing eleven isoforms and subisoforms and codified by 11 active genes located in a cluster on chromosome 16 (MT1A, MT1B, MT1E, MT1F, MT1G, MT1H, MT1M, MT1X, MT2A, MT3 and MT4) [77], although there are five additional genes (pseudogenes), not expressed in humans [78]. All MTs share a high degree of homology at the nucleotide and amino acid levels and retain 20 invariant metal-binding cysteinyl residues [79]. While MT1 and MT2 are present in almost all tissues, MT4 is most abundant in certain stratified tissues [80] and the expression of MT3 has been exclusively related to brain. Although this latter one is also present in heart, kidneys and reproductive organs [81,82] being even a ubiquitously expressed gene [78]. As discussed below, in the human eye, MT1 and MT2 isoforms are expressed to a greater or lesser extent in all of the ocular tissues, while MT3 expression is restricted mainly to the retina and MT4 transcript is not detected [15].
3.1.2. Functions
MTs, considered multipurpose proteins, are involved in a great diversity of processes, including cellular zinc homeostasis, metal detoxification, defense against oxidative damage through free radical scavenging and neuroprotection [83].
Zinc is an integral part of up to 10% of human proteins [84], being its homeostasis essential for proper cell function and metabolism. Cellular zinc homeostasis is dependent on proteins involved in the influx and efflux of zinc from cells (zinc transporters) and storage within cells (zinc-binding proteins). MTs also function as the main intracellular zinc reservoir as one MT molecule can bind up to seven zinc ions under physiological conditions. Zinc binding and release to MTs is mainly conditioned by the cellular redox state, where oxidation of cysteine residues of MTs facilitates the zinc transfer from these proteins to a wide variety of metalloproteins and transcription factors, regulating numerous cellular processes [85,86,87,88]. In addition to zinc ions, which are considered the main binding partner of apo-protein, MTs bind with high affinity other metal or heavy metal ions (e.g., Cu, Cd, Hg, Pb, Pt, etc.), exerting cellular protection against their toxicity [89,90,91,92,93,94].
One of the main interesting functions of MTs lies in their ability to protect cells and tissues against oxidative stress from multiples sources. MTs act as a potent antioxidant scavenging of free radicals, protecting the cells against the toxic effects from ROS and reactive nitrogen species (RNS) or electrophiles [95,96]. Under oxidative stress conditions, zinc is released from MTs and their free sulfhydryl groups may act as effective scavengers of free radicals. The antioxidant role of MTs has received broad experimental support both through in vitro and in vivo studies, using wild type and genetically modified animal models [97,98]. Furthermore, MTs can protect the cell by sequestering copper or iron, preventing their participation in redox reactions, as the Fenton’s one, and consequently avoiding the release of free radicals [99,100]. Alternatively, MTs may function as an indirect antioxidant by providing metal cofactors for antioxidant enzymes such as SOD, among others [101].
Concerning their neuroprotective role, the importance of MTs in the central nervous system (CNS) underlies in the brain high oxygen consumption rates and its elevated zinc levels, which makes it susceptible to oxidative stress and must tightly regulate the zinc homeostasis, respectively. Within the CNS, MT1/2 isoforms are primarily expressed in astrocytes and to a much lower extent in neurons. Moreover, the cells of microglia and oligodendrocytes, lacking expression of MTs under normal conditions, show up-regulation of these proteins in response to brain injury [102,103,104,105]. In relation to the MT3, while there are contradictory reports about its expression, it is generally accepted that is mainly localized in neural cells, especially those with high concentrations of zinc. Intriguingly, this MT3 protein was identified for the first time in patients with Alzheimer’s disease as a neuronal growth-inhibitory factor [82]. Its growth-inhibitory activity seems to be a specific functional characteristic of this MT3 isoform [104,106,107,108,109]. In contrast to MT1/2, which are up-regulated even against small damages, the response of MT3 to damage does not provide the same pattern in response to brain injury according to in vivo studies [110]. Since the neural retina is considered part of the CNS, the MTs perform within the eye complementary functions to those previously described. Neuroprotection mediated by MTs can be partially attributed to intracellular free radical scavenging and to zinc and copper regulation in injured cells [111,112]. Furthermore, this redox-induced zinc ions release activates multiple zinc-dependent transcription factors and proteins involved in MT-triggered neuroprotective processes [98,112,113,114]. There are also evidences of the extracellular role of MTs, released from astrocytes, in neuronal survival as well as enhancing regenerative growth in the injured brain [110,115,116]. The damage response mediated by extracellular MTs involves the membrane receptor protein megalin (LRP2), which belongs to the family of low-density lipoprotein receptor-related proteins. LRP2 binds and internalizes MTs in the neuronal cytoplasm, which activates the signal transduction pathways that support neurite outgrowth and survival [116,117,118,119,120].
Finally, the MT isoforms play an anti-apoptotic protective role [121,122]. Given that the regulation of the intracellular zinc concentration is essential for cell survival, a decrease of intracellular zinc levels or an increase in cytosolic free zinc can lead to apoptosis [123,124,125]. MTs supply zinc or other metals to target molecules, including enzymes, transcription factor or tumor suppressor gene products [126,127,128,129,130]. Furthermore, MTs interact with some proteins involved in apoptosis, and the specific isoforms MT1 and MT2 regulate the level, activity and cellular location of NF-κB, a transcription factor that is involved in the regulation of cell death [131,132,133]. This makes MTs to play an important role in the process of cancer initiation and progression as well as in the response of patients to specific treatment [134,135].
3.1.3. The Zn-MT Redox Cycle as an Antioxidant Defense Mechanism
Under oxidative stress conditions, MTs act as effective scavengers of free radicals through the Zn-MT redox system. This redox cycle, simplified in Figure 3 (panel b), has been previously described in numerous publications to date [71,87,98,136,137,138,139]. In brief, when the cellular environment becomes oxidized, zinc bound to MT (Zn-MT) is released through oxidation of the thiolate cluster with the consequent formation of MT-disulfide (thionin) [140]. This process is enhanced in the presence of free radicals such as nitric oxide, ROS and GSSG. The released zinc can be stored (zincosomes), transferred to other zinc-binding proteins or interact with transcription factors, including the metal regulatory transcription factor 1 (MTF-1), which induces MTs expression in the cellular nucleus [74]. MT-disulfide is unstable and may be degraded. However, when the environment becomes reduced due to, for example, an increase in the glutathione/glutathione disulfide ratio (GSH/GSSG), MT disulfide is reduced to MT-thiol (thionein), process that may be facilitated by a selenium-derived catalyst [141]. Later, in the presence of zinc, the Zn–MT system will be then quickly reconstituted to its reduced form [142].
3.1.4. MTs Regulation
Despite the widely conserved sequence of MTs, the RNA expression level of each MT isoform is unique mainly due to differences in the number, location, and orientation of the regulatory sequences of their promoters [143]. Furthermore, human MT isoforms are regulated in a cell type–specific manner, which may allow each isoform to have a specific role [144,145]. The basal activity of human MTs is mainly regulated by several general transcription factors, including the transcription factor II D (TFIID) complex, which consists of the TATA-binding protein (TBP) and TBP-associated factors, and the transcription factor Sp1, which binds to multiple GC boxes [143,146,147]. In addition, while MT1 and MT2 expression can be induced by a wide variety of stimuli, including metal ions, inflammatory mediators (such as cytokines), growth factors, hormones, oxidative stress, irradiation, and other stressors, MT3 and MT4 are constitutive expressed tissue-specific isoforms that do not respond to metal induction [72,148,149]. Induced expression of the MT1 and MT2 genes occurs primarily at the level of transcription through regulatory sequences in the 5′-promoter region of MTs genes [110,150,151]. These cis-acting DNA elements that can bind diverse transcription factor include:
- Article I.
- Metal response elements (MRE): The MTF-1 transcription factor is a central regulator of the metal inducible expression levels of MT1 and MT2. The binding of zinc to MTF-1 enables its union to MREs in the promoter region, which initiates gene transcription [146,147]. Heavy metal ions like Zn, Cu, Cd or Hg, as well as hypoxia, oxidative stress, glucocorticoids, nitric oxide and high temperature induce the transcriptional activity of MTF-1 [152,153,154,155].
- Article II.
- Article III.
- Article IV.
- STAT-binding sites: Elements activated by signal transducers and activators of transcription (STAT) proteins mediate transcriptional responses to cytokines [162]. Pro-inflammatory cytokines secreted by the activated macrophages during acute inflammation induce the expression of MTs [163]. STAT proteins convert the cytokine signal into gene expression programs regulating the inflammatory response in different cell types [164,165] and, in a cooperative mechanism of negative feedback, MTs inhibit the release of pro-inflammatory cytokines [166,167].
- Article V.
On the other hand, some transcription factors and DNA binding proteins have been reported as down-regulators of MTs expression, including transcription factor PU.1 [170,171], the 120-kDa zinc finger protein (PZ120) [172], the CCAAT/enhancer-binding protein alpha (C/EBP α) [173], and the Ku protein [174], among others. Moreover, epigenetic modifications can also modulate MTs transcription, such as the methylated and un-methylated MT1 promoters that are differentially regulated by DNA methyltransferase and methyl-CpG binding proteins [175]. Also, acetylation of histone binding sites results in chromatin unfolding and transcription induction [176]. Finally, the levels of MTs are influenced by the intracellular protein degradation, in which MTs turnover depends on cellular conditions. Oxidized and/or metalated proteins are more resistant to degradation that the native ones, directly depending on the metal and occupied binding sites [177,178].
3.2. Oxidative Stress, Age-Related Diseases and MTs in the Human Eye
As previously mentioned, the excess of ROS and subsequent oxidative stress causes damage to DNA, lipids and proteins inhibiting their biological functions [179,180,181,182,183], which affects the eye natural barriers against oxidative damage resulting in the pathogenesis of numerous age-related ocular diseases including cataracts [184,185,186], photokeratoconjunctivitis and photokeratitis [187,188,189,190], dry-eye disorders [191,192,193,194,195,196,197,198], diabetic retinopathy [199,200,201,202,203,204,205,206,207,208,209], retinal vascular diseases [210,211,212], retinal dystrophies [213], glaucoma [214,215,216,217,218,219,220,221,222,223] and AMD [224,225,226,227,228,229,230,231,232]. Under normal physiological conditions, oxidative stress is reduced by the presence of antioxidants and cellular repair systems, being the ocular tissues enriched in MTs that undoubtedly contribute to mitigate the negative effects of oxidants by capturing and neutralizing free radicals through their cysteine sulfur ligands [79,138,233,234].
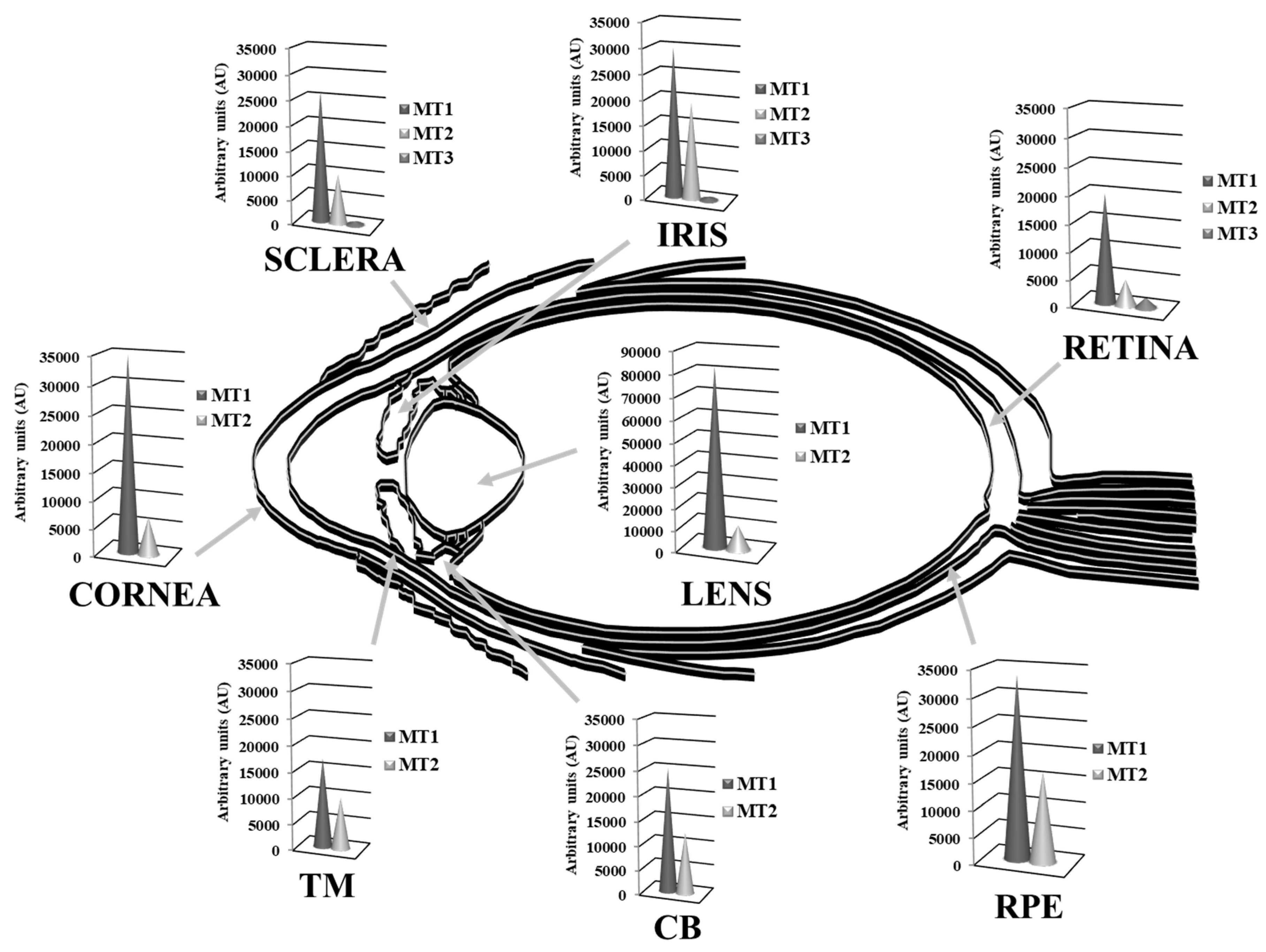
Multiple MT isoforms are highly and diversely expressed in the human ocular tissues [15,138]. Alvarez et al., 2012 [15] analyzed the gene expression profile of MTs isoforms in normal human eye donors (cadavers) by microarray techniques and compared their abundance between tissues, showing in Figure 4 the MT isoforms expression levels within eye structures, clustered in MT1, MT2 and MT3 subgroups. While MT1 and MT2 groups are abundantly expressed, exceedingly in the cornea, lens and retina-RPE tissues, MT3 is barely distributed in the neurosensory retina. The MT1A, MT2A and MT1X sub-isoforms are highly present in all ocular tissues, while MT1E, MT1F, MT1M and MT1G are expressed in much lower levels, with the exception of MT1G that is abundantly distributed in lens followed by the RPE. Conversely, MT1H and MT3 isoforms are expressed in low levels, but restricted preferentially to the lens and the retina, respectively. Besides, according to this study, MT1 and MT2 protein isoforms are highly expressed in the epithelial and endothelial corneal cells and confined in the cytoplasm.
Using micro-dissected human lens epithelial cells and fibers, Oppermann et al. 2001 [235] analyzed the MT expression at mRNA level by reverse transcription polymerase chain reaction (RT-PCR) and at a protein level by immunoblotting, observing different spatial expression patterns. Specifically, MT1E, MT1F, MT1G, and MT1H isoforms were detected in both lens epithelium and fiber cells, while MT1E and MT1F isoforms were expressed at slightly higher levels in the epithelium than in the lens fibers. By contrast, MT2A was only detected in lens epithelium, although low amounts of MT2A were detected in fibers from individual lenses presenting very high amounts of RNA. Due to the low amplification specificity, spatial analysis was not performed on isoform MT1L. At protein level, a predominance of MT protein in the lens epithelium relative to the fibers was observed. These reported differences in the spatial expression patterns between MT1 and MT2 may indicate distinct roles for these isoforms in the human lens. According to Tate et al. 2002 [236], within the RPE and choroids, the MT isoforms were detected by in situ hybridization, and to a lesser extent, in the sclera. The RT-PCR analysis corroborated the in situ hybridization findings and demonstrated that MTs were mainly distributed in the RPE and the photoreceptor layer of the retina. Moreover, the high metabolic oxygen flux and high phagocytic activity of the RPE correlate with the high expression levels of MT isoforms and their protective function against apoptosis and oxidative stress [15,138].
Interestingly, MT3 isoform is preferentially distributed in the cytoplasm of the RGCs and the retinal nerve fiber layer (NFL), while is also localized in the stromal cells cytoplasm of the iris [138]. The cell-restricted and isoform-specific MT3 detection in RGCs may reflect biochemical and/or physiological differences with other MT isoforms. The neuronal RGCs are the first retinal cells to undergo cell death in age-related eye diseases such as glaucoma, and they are highly sensitive to damage during oxidative stress caused by intense visible and UV light exposure [217].
Therefore, the higher levels of expression and diversity of MT isoforms in human cornea, lens and retina-RPE suggest a MTs role in cellular defense against oxidative stress in these ocular tissues that represent natural barriers to external insults (i.e., UV light) with physiological roles against age-related eye diseases [237]. The high content of sulfhydryl groups allows MTs to scavenge superoxide anion and hydroxyl radical with an affinity more than 300-fold higher than that of reduced GSH [238]. The structure of MTs provides a chemical environment by which the cysteine sulfur ligands can be oxidized and reduced concomitantly with the release and binding of zinc (see Figure 3) [138,142], conferring the antioxidant activity on the Zn-MT complex [87,138]. The MT affinity for zinc ions varies depending on the number of elements bound per protein: the first four zinc ions bind tightly (log K = 11.8), the next two bind with intermediate strength (log K = 10), and the last one bind relatively weak (log K = 7.7) [71,77]. These different stability constants imply that zinc release is thermodynamically favorable in saturated MTs, which entails that Zn7-MT species present greater antioxidant capacity than Znx-MT species (where x ranges from 0 to 6 zinc atoms), and so on [15,138]. Hence, the antioxidant activity and the capacity of MTs and the Zn-MT redox system of free radical scavenging might vary depending on the occupation rate of metals per protein molecule within each ocular protection barrier.
Therefore, the higher levels of expression and diversity of MT isoforms in human cornea, lens and retina-RPE suggest a MTs role in cellular defense against oxidative stress in these ocular tissues that represent natural barriers to external insults (i.e., UV light) with physiological roles against age-related eye diseases [237]. The high content of sulfhydryl groups allows MTs to scavenge superoxide anion and hydroxyl radical with an affinity more than 300-fold higher than that of reduced GSH [238]. The structure of MTs provides a chemical environment by which the cysteine sulfur ligands can be oxidized and reduced concomitantly with the release and binding of zinc (see Figure 3) [138,142], conferring the antioxidant activity on the Zn-MT complex [87,138]. The MT affinity for zinc ions varies depending on the number of elements bound per protein: the first four zinc ions bind tightly (log K = 11.8), the next two bind with intermediate strength (log K = 10), and the last one bind relatively weak (log K = 7.7) [71,77]. These different stability constants imply that zinc release is thermodynamically favorable in saturated MTs, which entails that Zn7-MT species present greater antioxidant capacity than Znx-MT species (where x ranges from 0 to 6 zinc atoms), and so on [15,138]. Hence, the antioxidant activity and the capacity of MTs and the Zn-MT redox system of free radical scavenging might vary depending on the occupation rate of metals per protein molecule within each ocular protection barrier.
3.2.1. Antioxidant Activity of MTs in the Ocular Surface
The production of oxidative stress might be induced on the ocular surface as a result of prolonged exposure to atmospheric oxygen and UV radiation, in addition to deficient support of antioxidants due to the instability in tear film, thereby, contributing to the onset of several eye diseases related with corneal defects. Tear fluid includes in its composition several antioxidants such as ascorbic acid, lactoferrin, uric acid, cysteine and MTs [239,240]. MTs are expressed in stem cells, located in the basal epithelium at the corneoscleral limbus [241,242], which are responsible of the maintenance of a healthy corneal epithelium. It has also been reported the MT expression in human corneas by laser capture microdissection [243] microarrays techniques [15] and RNA sequencing [244], where MT1A, MT2A and MT1X sub-isoforms were highly expressed [15]. In particular, immunohistochemistry analysis reflected that MT1/2 isoforms were confined in the cytoplasm of the corneal epithelium and endothelium [138]. Gottsch et al., 2003 [245] compared the gene expression profile in normal human corneal endothelium with corneal endothelium affected with Fuchs’ dystrophy using serial analysis of gene expression. These Fuchs’ endothelial cells exhibited a decrease in the abundance of transcripts related to antioxidants and proteins, conferring protection against toxic stress and an increase of MT2A mRNA, which may represent tissue injury repair in Fuchs’ dystrophy and/or tissue protection against oxidative stress. Conversely, De Roo et al., 2017 [246] also studied the transcriptome of corneal endothelial cells affected by Fuch’s dystrophy using microarrays and found that MT2A was down-regulated. These differences could be caused by different stages of the disease. They suggest that the oxidant-antioxidant imbalance in Fuchs’ endothelial cells could be a consequence of the down-regulation of antioxidants, including MTs, without compensatory up-regulation of catalase and glutathione-dependent antioxidants [247].
Recently, Kaluzhny et al., 2020 [248] evaluated the regulation of genes related to oxidative stress response in a reconstructed three-dimensional (3D) human corneal epithelial tissue, observing that only UVB led to significant up-regulation of MT3 while hydrogen peroxide, nitrogen-mustard and desiccating stress conditions have not exert any observable effect. Interestingly, mice injected with diquat capable to induce oxidative stress led to up-regulation of MT1 isoform [249], which may imply gene specific regulation mediated by oxidative stress. This cell- or tissue-specific antioxidant function of MTs was further discussed by a seminal work of Álvarez et al., 2012 [15]. Using an in vitro model of human corneal epithelial cells, the stoichiometry of the Zn-MT redox system was studied, showing that under steady-state conditions native MTs contained six zinc and one copper ions (Zn6Cu1-MT). Moreover, when zinc or pro-inflammatory cytokines were added separately, it favored the formation of Zn7-MT species, which may mean it interacts more effectively with ROS to decrease potential oxidative damage. Altogether, the number of zinc ions per MT molecule may predict the cell or tissue ability to overcome oxidative stress insults in the ocular surface of the human eye, with potential implication in the onset or progression of age-related ocular dystrophies.
3.2.2. Antioxidant Activity of MTs in the Lens
The major threat to lens transparency is oxidative damage, especially from photooxidation origin, which accumulates over the lifetime of an individual [250] and is involved in the pathogenesis of cataracts [184]. The lens responds to oxidative stress by increasing the production of glutathione, SOD and CAT, according to classical literature [251,252]. However, during ageing the efficacy of the lens antioxidant defenses declines, with a rate of 12% per decade, contributing to the development of lens diseases [253]. Kantorow et al. 1998 [254] analyzed and compared, by differential display RT-PCR, the RNAs obtained from human lens epithelial cells of cataractous and control subjects, showing higher levels of MT2A associated with age related cataract. Studies by Whitson et al., 2017 [255] have shown that MT2A and MT1 are up-regulated in a GSH-depleted mouse model of cataracts, suggesting that they have a counteracting protective role. These findings support the hypothesis that the up-regulation of MT2A in the cataractous lenses represents a specific protective response of this tissue against oxidative or other stress events related with the onset of cataracts [254,256]. On the other hand, Hawse et al., 2003 [257] compared the global gene expression profiles between pooled age-matched human lens epithelium isolated from cataract and clear lenses (control subjects) by oligonucleotide microarray hybridization. The expression levels of a subset of genes altered were further evaluated by semi-quantitative RT-PCR, identifying significant decreases in genes associated with oxidative stress such as the MT1 isoforms in the cataractous human lens epithelium. All these findings suggested an age- and oxidative stress-related decrease in the antioxidant activity of MTs in the human lens.
In vitro studies reported that over-expression of MT2A in human lens epithelial cells results in protection against cadmium and tertiary butyl hydroperoxide-induced oxidative stress [258]. Later, using the lens epithelial cell line alphaTN4-1, the protective effects of MT1 and MT2 against UV-induced oxidative stress have been quantitatively determined [259]. Recent studies carried out by González-Iglesias et al., 2017 [260] reported that MT1 and MT2 isoforms are preferential located in the human lens epithelium by immunohistochemistry analysis. A human lens epithelial cell line was used as in vitro model to study the Zn-MT regulation and stoichiometry, under the native state and following zinc and pro-inflammatory interleukin-1α (IL1α) supplementation. Their findings suggested that, while Zn3Cu0.2MT species are present under control conditions, zinc or pro-inflammatory cytokines changed their elemental composition of MTs to Zn7Cu0.1MT or Zn7Cu0.2MT, indicating that the new MTs are saturated by zinc ions, rendering the cell more resistant to oxidative insults. It must be stressed that MT proteins in the lens are in different redox state, compared to the cornea, being the latter with the highest antioxidant power, which may be related to the roles of cornea and lens as the first, and second natural barriers combating free radicals, respectively [15,260].
3.2.3. Antioxidant Activity of MTs in the Retina-RPE
The neurosensory retina and its RPE are particularly susceptible to oxidative stress because of their high rate of oxygen consumption, their exposure to visible and UV light and their high proportion of polyunsaturated fatty acids [261,262,263,264]. ROS are involved in several retinal pathologies, and therefore conscious strategies to enhance the production of antioxidant enzymes, including MTs, have been addressed to reduce oxidative stress or to promote cytoprotective-signaling pathways [213]. Seminal studies demonstrated that MT expression decreases in the retina-RPE with aging and oxidative stress, which results in release of zinc within RPE, most dramatically in the macular region [265]. Various evidence implicates the imbalance of the MT antioxidant defense system with the development of AMD as a result of RPE degeneration and photoreceptor loss [266]. Tate et al., 1993 [265] reported that MTs decline as a function of age in the macular-RPE of patients with AMD, with proportionately greater loss in the macula than in the periphery. Moreover, while the antioxidant CAT and the metal zinc also decrease in aged human retinal pigment epithelial cells with signs of AMD, like MTs, SOD seems to increase their total levels [267,268,269]. Recently, Wi et al., 2020 [270] observed higher expression of MT3, MT1G, MT2A and GPX3 in Müller cells of the periphery of aged retinas compared with those cells of the foveal region using single-cell RNA sequencing, stating that the MT expression pattern may underlie foveal to peripheral retinal ageing gradient [271]. These observations triggered plenty of research both in animal models and in vitro cell cultures. Studies carried out on retinal and RPE homogenates from an early onset macular degeneration monkey model, detected a decrease in the activities of CAT and GPX antioxidants, lowered zinc concentration and reduced MT gene expression in affected retinas compared with control individuals [272]. Later, Miceli et al., 1999 [273] found that a zinc deficiency diet reduced the concentration of MT in the retina and the RPE, and increased the levels of retina lipid peroxidation in rats, where a crosstalk between oxidative stress and MT expression exists since MT transcription is up-regulated by oxidative stress [158].
Light-induced damage has been widely used as a model system to study retinal degeneration [274], whereby prolonged and/or raised light exposure causes retinal oxidative stress [20,43,275,276,277,278,279]. In this sense, Chen et al., 2004 [280] analyzed the gene expression profile following light-induced retinal damage in a mouse model. The microarray analysis of RNA isolated from mice retinas subjected to acute photic injury identified 70 genes up-regulated (≥2-fold) when compared with control subjects, determining the overexpression of MT1 and MT2 isoforms subsequently confirmed by qPCR. Similarly, the simultaneous analysis of MT at protein and RNA levels by immunoblot, immunohistochemistry and qPCR in light-induced oxidative damage mouse retinas confirmed the increased expression of the antioxidant MTs, and suggested that their up-regulation is an important acute retinal response to photo-oxidative stress [281]. The up-regulation of MT2A after light exposure was also found by Natoli et al., 2010 [282] in a rat model of retinal degeneration using microarrays. Later, Tsuruma et al., 2012 [283] evaluated the specific roles of MT1, MT2 and MT3 isoforms in light-induced retinal degeneration using MT1/2 deficient [284,285,286], MT3–deficient and wild-type (WT) mice. The expression levels of all MT isoforms were significantly up-regulated in the murine retinas after light exposure. The functional role of MTs in light-induced retinal dysfunction was investigated in the MT1/2- and MT3–deficient mice by electroretinogram recording, which a-waves and b-waves reflected the function of photoreceptors and Müller cells, respectively. Interestingly both reductions in the a- and b-wave amplitudes were significantly greater in the MT3–deficient mice than in the WT following light exposure. Moreover, the ONL was significantly thinner in the MT3-deficient mice compared to the light exposed WT, while no changes in the retinal function or ONL thickness were observed in the MT1/2–deficient mice. These observations were confirmed using MT3 knockdown by siRNA in a mouse transformed photoreceptor-cell line (661W) subjected to light-induced damage, observing amplified cell damage and increased production of ROS in response to light exposure [283]. Altogether, these findings suggested that MT3 might protect more effectively against light-induced retinal damage compared to MT1 and MT2, probably mediated by its antioxidant properties in addition to other major biological roles in cellular processes and diseases [287].
MT1/2-deficient mice were also used to study the functional roles of MTs in retinal damage induced by intravitreal injection of N-methyl-D-aspartate (NMDA), together with the evaluation of the protective role of zinc and vitamin D3 [288]. NMDA induces retinal cell damage by activation of NMDA receptor, which leads to intracellular Ca2+ influx, increases ROS that affect cell viability [289,290] and depletes the intracellular GSH that may decrease the intracellular capacity against ROS [291,292]. Whilst intravitreal NMDA increased the MT immunostaining in the inner retina of WT mice (NFL, INL and some of the RGCs), no MT labeling was detected in the retina of MT1/2-deficient mice. Conversely, at mRNA level, MT2 increased but MT1 and MT3 decreased following NMDA injection in the WT mice. The cell loss in RGC layer after NMDA injection was significantly higher in MT1/2-deficient mice compared to the WT, but no differences were observed in the inner plexiform layer thickness in both studied animal models. Moreover, intravitreous injection of zinc or vitamin D3 24-h after NMDA injury increased inner retinal MT immunoreactivity, in line with previous reports [159,293], and significantly attenuated NMDA-induced RGC layer loss in WT mice, although zinc pretreatment did not protect against RGCs loss in MT1/2-deficient mice. These findings indicated that MT up-regulation, specifically the MT2 isoform, might play an important neuroprotective role against retinal oxidative damage.
The use of cell models for in vitro studies as simplified systems has also contributed to elucidate the molecular mechanism by which MTs protect against oxidative stress in the retina. Tate et al., 1999 [294] used an in vitro model of zinc deficiency through the primary culture of RPE cells in low zinc medium to identify a correlation between the concentration of MTs and the activity of antioxidant enzymes with the oxidative damage of reactive intermediates. As reported, while in low zinc cultured cells (0.55 µM zinc), MTs and CAT content decreased following oxidative stress, the cultured cells in adequate zinc levels (14 µM zinc) increased both MTs and CAT levels after oxidative stress damage. Thereby, zinc may stabilize both proteins by protecting their functional thiol groups from oxidative damage [295], along with the induction of MTs synthesis [296,297], which contain a high number of zinc-thiolate clusters able to scavenge free radicals [136]. In line with this, Tate et al., 2006 [298] tested the effect of different zinc physicochemical formulations (zinc acetate, zinc chloride, zinc cysteine and zinc sulfate) on the concentration of antioxidant proteins in a primary culture of human RPE cells. Increased CAT and GPX activities and glutathione and MT cellular content were observed after zinc supplementation with all tested formulations, highlighting greatest increase using zinc cysteine. In addition, the cytotoxic effects of H2O2 and t-butyl hydroperoxide were reduced after pretreatment with these zinc formulations, concluding the strong antioxidant protective capacity of MT proteins through the Zn-MT redox cycle. Even more, the MT antioxidant role was evidenced by the former studies of Lu et al., 2002 [243] showing that elevation of MT levels by transfection of RPE cells (D407 spontaneously immortalized cell line) protected against toxic levels of cadmium [299], heme- and iron-induced oxidation and UV light-induced apoptosis. In this regard, the combination of N-retinylidene-N-retinylethanolamine (A2E) and blue light permitted to study causative genes of retinitis pigmentosa using primary cultures of RPE cells under oxidative conditions, showing altered expression of aryl hydrocarbon receptor (AHR) and retinal outer segment membrane protein 1 (ROM1), but also other genes, including MTs [300]. Similarly, the effect of A2E-induced oxidative stress on human RPE cells determined relevant differences in gene expression and splicing events contributing to elucidate molecular mechanisms linking oxidative stress and retinal dystrophies [301].
Commercial ARPE-19 cells, a spontaneously immortalized cell line of human RPE have been widely used as in vitro model to study the cellular effects of oxidative stress. Several works have reported that ARPE-19 cells tend to be extremely resistant to oxidative stress [302,303,304,305]. Even though their capacity for H2O2 scavenging is not superior to other cell models, ARPE-19 cells are able to withstand a single initial dose (i.e., bolus addition) of 15 mM H2O2 before lysosomal rupture and subsequent induction of apoptosis occur [305]. Karlsson et al., 2013 [306] depicted that ARPE-19 cells contain very high basal levels of MTs, ferritin and heat shock protein 70 (HSP70), which can be easily up-regulated following specific stimulation with zinc, iron, or heat, respectively. These steady state high protein levels, specifically those of MTs, may be related with their high resistance to oxidative stress. Later studies of the same laboratory explored whether the up-regulation of the synthesis of MTs, ferritin and HSP70 could further increase the tolerance of ARPE-19 facing oxidative stress [307], while attenuation of MTs levels would affect cell susceptibility to free radicals. Therefore, decreasing MT levels using siRNA made cells more susceptible to oxidative stress, being statistically significant after 10 mM H2O2 following the conditions assayed. Although, treatments with 100 µM ZnSO4, 500 µM FeCl3 or 43 °C heat shock exposure increased the levels of MTs, ferritin or HSP70, respectively, this up-regulation did not improve the tolerance of ARPE-19 cells to H2O2. It must be stressed that these studies were carried out with 60 h-old cells at a considerably higher density, likely affecting their sensitivity along with an already reached maximum protection. Moreover, same treatments using different cell lines from non-ocular tissues with much lower basal levels of these proteins provided increased resistance to H2O2 stress and prevention of lysosomal membrane permeabilization [308,309,310,311,312]. In connection with the above, Choudhary et al., 2005 [313] reported that ARPE-19 treated with a thionein hexapeptide H-Lys-Cys-Thr-Cys-Cys-Ala-OH, which is a fragment of mouse liver MT1 [314,315] significantly protected against H2O2 and 4-hydroxynonenal (HNE)-induced mitochondrial depolarization and ameliorated H2O2 and HNE-induced apoptosis. Additional evidence about the antioxidant properties of MTs were collected in the recent work of Wang et al., 2020 [316] on ARPE-19 cells exposed to sodium iodate-induced oxidative damage. In this study, cells transfected with overexpression plasmids of MT2A and MT1G genes showed a higher cell survival rate than control cells after exposure to sodium iodate (10 µM for 18 h). Conversely, silencing of the same genes with siRNA was associated with an accelerated cell death after treatment. The same study also demonstrated that the small molecule D609 acts as an effective antioxidant in RPE cells partially due to the induction of MTs expression.
The in vitro production of free radicals using H2O2 provides high variability of induced oxidative stress due to the Fenton reaction, where iron catalyzes the conversion of H2O2 into hydroxyl radical intracellular turning out into a non-specific and unstable mechanism difficult to reproduce [317]. In this way, additional stressors have been widely used to elicit free radicals in vitro and test the antioxidant mechanism of MTs in the RPE. Recently, Ahmed et al., 2016 [318] studied the mitochondrial oxidative stress in ARPE-19 cells treated with paraquat and in the presence or absence of the neuroprotective xaliproden. Paraquat is an herbicide structurally similar to 1-methyl-4-phenylpyridinium ion (MPP+), which is converted within the cell into a free radical that interacts with molecular oxygen to form superoxide [319]. Using these reagents, the authors observed by qPCR an induction in the transcription of MT1 in the presence of paraquat (5.5-fold) or xaliproden (3.3-fold) and a synergistic effect when combined (10-fold increase). Albeit the induction of the antioxidant pathways was not sufficient to prevent the toxic oxidant effect on ARPE-19 cells, the increased stimulation of MT1 and other antioxidant genes by the treatment combined (paraquat and xaliproden) resulted in an increased survival of the cells.
Similarly, ARPE-19 cells treated with 8-hydroxy-2-(di-n-propylamino)-tetralin (8-OH-DPAT) in the presence of paraquat led to a significant increase of MT1 expression along with other antioxidant enzymes, including heme oxygenase 1 (HO1), NAD(P)H dehydrogenase quinone 1 (NqO1), CAT and SOD1 [320]. The 8-OH-DPAT, a 5-hydroxy-tryptophan 1a (5HT1a) receptor agonist with neuroprotective effects [321], increased the MT synthesis in a primary culture of astrocytes [322] suggesting that 8-OH-DPAT may protect ARPE-19 cells by the specific induction of MT1, HO1 and NqO1 [320]. Moreover, differentiated ARPE-19 cells treated with paraquat, 8-OH-DPAT or a combination of both (paraquat and 8-OH-DPAT) showed a potent induction of MT1 (27-fold), HO1 (2-fold) and NqO1 (45-fold) expression [320], but with the absence of SOD1 and SOD2 overexpression. In line with this, the authors carried out in vivo studies in the retinas of mice treated with 8-OH-DPAT, showing a significant increase in the expression of MT1 (2-fold), HO-1 (4-fold), NqO1 (1.7-fold) CAT (17-fold) and SOD1 (1.6-fold), while SOD2 remained constant, concluding that 8-OH-DPAT specifically induces antioxidants (e.g., MTs), protecting the RPE and the neural retina from mitochondrial oxidative stress.
The protective role of MTs against oxidative species, and their inherent mechanisms, were further explored by Rodríguez-Menéndez et al., 2018 [323,324] using an in vitro model of immortalized human RPE cells (HRPEsv). Oxidative stress was induced in HRPEsv cells using 2,2′-azobis(2-methylpropionamidine) dihydrochloride (AAPH) and quantified by fluorescent probes, the MTs were induced through zinc supplementation and the Zn-MT system was quantitatively determined by mass spectrometry using enriched isotopes and chemometrics-based calculations. Zinc at different concentrations (25, 50 or 100 μM ZnSO4) specifically induced Zn-MT synthesis (1.6-, 3.6- and 11.9-fold, respectively), while pre-treated cells with zinc followed by AAPH oxidative stressor significantly reduced the ROS levels when compared to AAPH-treated cells. In these studies, the authors simultaneously obtained the zinc to MT ratio to determine whether oxidative stress may affect the stoichiometry and therefore their antioxidant power. In these HRPEsv cultured cells, under steady state conditions a Zn1.4Cu0.11MT stoichiometry was obtained, while under 100 μM zinc pre-treatment and APPH treatment the MT binding sites were saturated by zinc, i.e., Zn7.4Cu0.03MT. Moreover, AAPH treatment decreased MT levels (0.4-fold) but increased the number of zinc atoms bound to MTs (Zn2.07Cu0.17MT), which may be related with the antioxidant capacity of the Zn-MT system. The authors hypothesized that saturated MTs (Zn7-MT) may have the greater antioxidant capacity in HRPEsv cells, since when an excess of zinc is added the MTs interact more effectively with ROS [234,260]. It was therefore demonstrated that the Zn-MT overexpression significantly attenuated the oxidative stress induced by free radicals, whence the stoichiometry of Zn-MT plays an important role in oxidative stress response regulating cellular metal homeostasis and transcription factors.
4. Conclusions
Oxidative stress affects all structures of the human eye, specifically the ocular surface, the lens and the retina, which are considered antioxidant natural barriers against external insults. The imbalance between ROS production and the neutralizing ability of the antioxidant defense results in an excessive oxidative milieu that contributes to the onset or progression of age-related eye diseases. Oxidative stress has been implicated in the pathogenesis of dry-eye, cataracts, diabetic retinopathy, retinal vascular occlusions, retinal dystrophies, glaucoma and AMD diseases, inter alia. The underlying ocular antioxidant defense is a complex system responsible of free radicals scavenging and is composed of a vast variety of compounds, including both, enzymatic and non-enzymatic molecules. The main enzymatic antioxidants constituent the natural barriers of the human eye, i.e., cornea, lens and retina, include SOD, GPX, CAT and PRDX as primary antioxidants and TKT, G6PD, GSS and selenoproteins as secondary antioxidants. These antioxidants are diversely expressed along eye tissues and contribute to endogenous and exogenous free radical scavenging.
Even if MTs are not broadly considered enzymatic antioxidants, their redox activity dependent on zinc release warrants reappraisal consideration. This low molecular mass family of proteins neutralizes free radicals through their cysteine rich sulfur ligands and maintains cellular zinc homeostasis. The biological and biochemical significance of the multiple MT isoforms within the eye is not totally understood. The lack of specific antibodies for MT1 isoforms makes it difficult to assess their potential cell-specific distribution throughout the eye. Nowadays, novel therapeutic approaches against oxidative stress involved in eye diseases explore the enhancement of antioxidant enzymes production, including MTs, to reduce free radicals or to promote cytoprotection. Furthermore, considering that MTs expression decreases with age and oxidative stress, boosting the MT system may contribute to slow down or stop eye tissues degeneration and disease progression.
In this sense, the stoichiometry of Zn-MTs is of fundamental importance to oxidative stress response and to transduce oxidative signals into zinc signals. The MTs stoichiometry has been studied in three in vitro models of eye cells, obtaining Zn6MT species in corneal epithelial cells, Zn3MT species in the lens epithelial cells, and Zn1.5MT in RPE cells under control conditions. The observed different Zn-MT redox state may be related to the roles of the cornea as the initial barrier protecting the inner ocular tissues against external insults including UV light, the lens as second barrier responsible of the refraction of light, and the RPE as third barrier reducing light scatter. Therefore, the MT antioxidant capacity seems different within each tissue, being probably higher in the cornea, compared to the lens and to the RPE, which is related to the requirement of a more effective antioxidant system in the outer layers exposed to a higher oxidative milieu. As a perspective, the Zn-MT redox system might be used as a therapeutic target to combat oxidative stress in the ocular tissues for the treatment of age- and oxidative-stress-related eye diseases, through its reactivation by MT synthesis induction, thionein system regeneration, or both. Moreover, its combination with the activation of additional enzymatic antioxidants within the eye may contribute to diminish free radicals and enhance population healthy ageing.
Author Contributions
Writing—original draft preparation, A.Á.-B., L.Á., M.G. and E.A.; literature search: A.Á.-B., L.Á., M.G. and E.A.; writing—review and editing, R.P. and H.G.-I. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by IDE/2019/000289 project of “Instituto de Desarrollo Económico del Principado de Asturias” (IDEPA) of Gobierno del Principado de Asturias and FEDER, and by CTQ2016-79015-R project of Agencia Estatal de Investigación (Spain) and FEDER, The Instituto Oftalmológico Fernández-Vega and Fundación de Investigación Oftalmológica acknowledge financial support from the Fundación Rafael del Pino (http://www.frdelpino.es) from which H.G.-I. is “Catedrático Rafael del Pino”.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
Abbreviations
| 5HT1a | 5-hydroxy-tryptophan 1a |
| 8-OH-DPAT | 8-hydroxy-2-(di-n-propylamino)-tetralin |
| A2E | N-retinylidene-N-retinylethanolamine |
| AAPH | 2,2′-azobis(2-methylpropionamidine) dihydrochloride |
| AHR | Aryl hydrocarbon receptor |
| ALDH | Aldehyde dehydrogenase |
| AMD | Age-related macular degeneration |
| AP-1 | Activator proteins 1 |
| AP-2 | Activator proteins 2 |
| ARE | Antioxidant response element |
| AU | Arbitrary units |
| C/EBPα | CCAAT/enhancer-binding protein alpha |
| CAT | Catalase |
| CB | Ciliary body |
| CNS | Central nervous system |
| G6PD | Glucose-6-phosphate dehydrogenase |
| GADPH | Glyceraldehyde-3-phosphate dehydrogenase |
| GPX | Glutathione peroxidase |
| GREs | Glucocorticoid response elements |
| GSH | Glutathione |
| GSR | Glutathione reductase |
| GSS | Glutathione synthetase |
| GSSG | Glutathione disulfide |
| HNE | 4-hydroxynonenal |
| HO1 | Heme oxygenase 1 |
| HRPEsv | Human RPE cells |
| HSP70 | Heat shock protein 70 |
| IL1α | Interleukin-1α |
| INL | Inner nuclear layer |
| LRP-2 | Membrane receptor protein megalin |
| MPP+ | 1-methyl-4-phenylpyridinium ion |
| MRE | Metal response elements |
| MTF-1 | Metal regulatory transcription factor 1 |
| MTs | Metallothioneins |
| NF-1 | Nuclear factor 1 |
| NFL | Nerve fiber layer |
| NMDA | N-methyl-D-aspartate |
| NqO1 | NAD(P)H dehydrogenase quinone 1 |
| O2· | Superoxide anion |
| ONL | Outer nuclear layer |
| PPP | Pentose phosphate pathway |
| PRDXs | Peroxiredoxins |
| PZ120 | 120-kilodalton zinc finger protein |
| RGC | Retinal ganglion cell |
| RNS | Reactive nitrogen species |
| ROM1 | Retinal outer segment membrane protein 1 |
| ROS | Reactive oxygen species |
| RPE | Retinal pigment epithelium |
| RT-PCR | Reverse transcription polymerase chain reaction |
| SEL | Selenoproteins |
| SOD | Superoxide dismutase |
| SRXN | Sulfiredoxin |
| STAT | Signal transducers and activators of transcription |
| TBP | TATA-binding protein |
| TFIID | Transcription factor II D |
| TKT | Transketolase |
| TM | Trabecular meshwork |
| TXN | Thioredoxin |
| UV | Ultraviolet |
| USF1 | Upstream stimulatory factor 1 |
| WT | Wild-type |
References
- Forrester, J.V.; Dick, A.D.; McMenamin, P.G.; Roberts, F. The Eye, Basic Sciences in Practice, 3rd ed.; Saunders Elsevier: Edinburgh, UK, 2008; Volume 540, ISBN 978-0702028410. [Google Scholar]
- Levin, L.; Nilsson, S.; Ver Hoeve, J.; Wu, S.; Kaufman, P.; Alm, A. Adler’s Physiology of the Eye, 11th ed.; Saunders Elsevier: St. Louis/Mosby, MO, USA, 2011; ISBN 9780323057141. [Google Scholar]
- Sies, H. Oxidative stress: A concept in redox biology and medicine. Redox Biol. 2015, 4, 180–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ung, L.; Pattamatta, U.; Carnt, N.; Wilkinson-Berka, J.L.; Liew, G.; White, A.J.R. Oxidative stress and reactive oxygen species: A review of their role in ocular disease. Clin. Sci. 2017, 131, 2865–2883. [Google Scholar] [CrossRef] [PubMed]
- Cabrera, M.P.; Chihuailaf, R.H. Antioxidants and the integrity of ocular tissues. Vet. Med. Int. 2011, 905153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wojcik, K.A.; Kaminska, A.; Blasiak, J.; Szaflik, J.; Szaflik, J.P. Oxidative stress in the pathogenesis of keratoconus and Fuchs endothelial corneal dystrophy. Int. J. Mol. Sci. 2013, 14, 19294–19308. [Google Scholar] [CrossRef] [Green Version]
- Vinson, J.A. Oxidative stress in cataracts. Pathophysiology 2006, 13, 151–162. [Google Scholar] [CrossRef]
- Beebe, D.C.; Holekamp, N.M.; Shui, Y.B. Oxidative damage and the prevention of age-related cataracts. Ophthalmic Res. 2010, 44, 155–165. [Google Scholar] [CrossRef] [Green Version]
- Varma, S.D.; Chand, D.; Sharma, Y.R.; Kuck, J.F., Jr.; Richards, R.D. Oxidative stress on lens and cataract formation: Role of light and oxygen. Curr. Eye Res. 1984, 3, 35–57. [Google Scholar] [CrossRef]
- Nita, M.; Grzybowski, A. The Role of the Reactive Oxygen Species and Oxidative Stress in the Pathomechanism of the Age-Related Ocular Diseases and Other Pathologies of the Anterior and Posterior Eye Segments in Adults. Oxid. Med. Cell. Longev. 2016, 16, 3164734. [Google Scholar] [CrossRef] [Green Version]
- Tanito, M.; Kaidzu, S.; Takai, Y.; Ohira, A. Association between systemic oxidative stress and visual field damage in open-angle glaucoma. Sci. Rep. 2016, 6, 25792. [Google Scholar] [CrossRef] [Green Version]
- Beatty, S.; Koh, H.H.; Phil, M.; Henson, D.; Boulton, M. The Role of Oxidative Stress in the Pathogenesis of Age-Related Macular Degeneration. Surv. Ophthalmol. 2000, 45, 115–134. [Google Scholar] [CrossRef] [Green Version]
- Kowluru, R.A.; Kowluru, A.; Mishra, M.; Kumar, B. Oxidative stress and epigenetic modifications in the pathogenesis of diabetic retinopathy. Prog. Retin. Eye Res. 2015, 48, 40–61. [Google Scholar] [CrossRef] [PubMed]
- Ankamah, E.; Sebag, J.; Ng, E.; Nolan, J.M. Vitreous Antioxidants, Degeneration, and Vitreo-Retinopathy: Exploring the Links. Antioxidants 2020, 9, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alvarez, L.; Gonzalez-Iglesias, H.; Garcia, M.; Ghosh, S.; Sanz-Medel, A.; Coca-Prados, M. The stoichiometric transition from Zn6Cu1-metallothionein to Zn7-metallothionein underlies the up-regulation of metallothionein (MT) expression: Quantitative analysis of MT-metal load in eye cells. J. Biol. Chem. 2012, 287, 28456–28469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchitti, S.A.; Chen, Y.; Thompson, D.C.; Vasiliou, V. Ultraviolet radiation: Cellular antioxidant response and the role of ocular aldehyde dehydrogenase enzymes. Eye Contact Lens 2011, 37, 206–213. [Google Scholar] [CrossRef] [Green Version]
- Zoidis, E.; Seremelis, I.; Kontopoulos, N.; Danezis, G.P. Selenium-Dependent Antioxidant Enzymes: Actions and Properties of Selenoproteins. Antioxidants 2018, 7, 66. [Google Scholar] [CrossRef] [Green Version]
- Brennan, L.A.; McGreal, R.S.; Kantorow, M. Oxidative stress defense and repair systems of the ocular lens. Front. Biosci. 2012, 4, 141–155. [Google Scholar] [CrossRef]
- Chen, Y.; Mehta, G.; Vasiliou, V. Antioxidant defenses in the ocular surface. Ocul. Surf. 2009, 7, 176–185. [Google Scholar] [CrossRef] [Green Version]
- Saccà, S.C.; Roszkowska, A.M.; Izzotti, A. Environmental light and endogenous antioxidants as the main determinants of non-cancer ocular diseases. Mutat. Res. 2013, 752, 153–171. [Google Scholar] [CrossRef]
- Carocho, M.; Ferreira, I.C. A review on antioxidants, prooxidants and related controversy: Natural and synthetic compounds, screening and analysis methodologies and future perspectives. Food Chem. Toxicol. 2013, 51, 15–25. [Google Scholar] [CrossRef]
- Laher, I. Systems Biology of Free Radicals and Antioxidants; Springer: Heidelberg/Berlin, Germany, 2014; Chapter LXI; p. 4178. ISBN 978-3-642-30017-2. [Google Scholar]
- Hammond, B.R.; Johnson, B.A.; George, E.R. Oxidative photodegradation of ocular tissues: Beneficial effects of filtering and exogenous antioxidants. Exp. Eye Res. 2014, 129, 135–150. [Google Scholar] [CrossRef] [Green Version]
- Lassen, N.; Black, W.J.; Estey, T.; Vasiliou, V. The role of corneal crystallins in the cellular defense mechanisms against oxidative stress. Semin. Cell. Dev. Biol. 2008, 19, 100–112. [Google Scholar] [CrossRef] [PubMed]
- Lei, X.G.; Zhu, J.H.; Cheng, W.H.; Bao, Y.; Ho, Y.S.; Reddi, A.R.; Holmgren, A.; Arnér, E.S. Paradoxical Roles of Antioxidant Enzymes: Basic Mechanisms and Health Implications. Physiol. Rev. 2016, 96, 307–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siu, A.W.; Maldonado, M.; Sanchez-Hidalgo, M.; Tan, D.X.; Reiter, R.J. Protective effects of melatonin in experimental free radical-related ocular diseases. J. Pineal Res. 2006, 40, 101–109. [Google Scholar] [CrossRef]
- Brennan, L.A.; Kantorow, M. Mitochondrial function and redox control in the aging eye: Role of MsrA and other repair systems in cataract and macular degenerations. Exp. Eye. Res. 2009, 88, 195–203. [Google Scholar] [CrossRef] [Green Version]
- Babizhayev, M.A.; Yegorov, Y.E. Reactive Oxygen Species and the Aging Eye: Specific Role of Metabolically Active Mitochondria in Maintaining Lens Function and in the Initiation of the Oxidation-Induced Maturity Onset Cataract—A Novel Platform of Mitochondria-Targeted Antioxidants With Broad Therapeutic Potential for Redox Regulation and Detoxification of Oxidants in Eye Diseases. Am. J. Ther. 2016, 23, e98–e117. [Google Scholar] [CrossRef] [PubMed]
- Ohia, S.E.; Opere, C.A.; Leday, A.M. Pharmacological consequences of oxidative stress in ocular tissues. Mutat. Res. 2005, 579, 22–36. [Google Scholar] [CrossRef] [PubMed]
- Brigelius-Flohé, R. Glutathione peroxidases and redox-regulated transcription factors. Biol. Chem. 2006, 387, 1329–1335. [Google Scholar] [CrossRef]
- Ma, W.; Kleiman, N.J.; Sun, F.; Li, D.; Spector, A. Peroxide toxicity in conditioned lens epithelial cells—Evaluation of multi-defense systems. Exp. Eye Res. 2003, 77, 711–720. [Google Scholar] [CrossRef]
- Detienne, G.; De Haes, W.; Mergan, L.; Edwards, S.L.; Temmerman, L.; Van Bael, S. beyond ROS clearance: Peroxiredoxins in stress signaling and aging. Ageing Res. Rev. 2018, 44, 33–48. [Google Scholar] [CrossRef]
- Piatigorsky, J. Gene Sharing and Evolution the Diversity of Protein Functions; Harvard University Press: Cambridge, UK, 2007; Chapter XV; p. 320. ISBN 978-0674023413. [Google Scholar]
- Cooper, D.L.; Isola, N.R.; Stevenson, K.; Baptist, E.W. Members of the ALDH gene family are lens and corneal crystallins. Adv. Exp. Med. Biol. 1993, 328, 169–179. [Google Scholar] [CrossRef]
- Sax, C.M.; Kays, W.T.; Salamon, C.; Chervenak, M.M.; Xu, Y.S.; Piatigorsky, J. Transketolase gene expression in the cornea is influenced by environmental factors and developmentally controlled events. Cornea 2000, 19, 833–841. [Google Scholar] [CrossRef] [PubMed]
- Lampi, K.J.; Wilmarth, P.A.; Murray, M.R.; David, L.L. Lens β-crystallins: The role of deamidation and related modifications in aging and cataract. Prog. Biophys. Mol. Biol. 2014, 115, 21–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anbarasu, K.; Sivakumar, J. Multidimensional significance of crystallin protein-protein interactions and their implications in various human diseases. Biochim. Biophys. Acta 2016, 1860, 222–233. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Thompson, D.C.; Koppaka, V.; Jester, J.V.; Vasiliou, V. Ocular aldehyde dehydrogenases: Protection against ultraviolet damage and maintenance of transparency for vision. Prog. Retin. Eye Res. 2013, 33, 28–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Estey, T.; Piatigorsky, J.; Lassen, N.; Vasiliou, V. ALDH3A1: A corneal crystallin with diverse functions. Exp. Eye Res. 2007, 84, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Voulgaridou, G.P.; Tsochantaridis, I.; Tolkas, C.; Franco, R.; Giatromanolaki, A.; Panayiotidis, M.I.; Pappa, A. Aldehyde dehydrogenase 3A1 confers oxidative stress resistance accompanied by altered DNA damage response in human corneal epithelial cells. Free Radic. Biol. Med. 2020, 150, 66–74. [Google Scholar] [CrossRef]
- Xu, I.M.; Lai, R.K.; Lin, S.H.; Tse, A.P.; Chiu, D.K.; Koh, H.Y.; Law, C.T.; Wong, C.M.; Cai, Z.; Wong, C.C.; et al. Transketolase counteracts oxidative stress to drive cancer development. Proc. Natl. Acad. Sci. USA 2016, 113, E725–E734. [Google Scholar] [CrossRef] [Green Version]
- Romi, F.; Helgeland, G.; Gilhus, N.E. Heat-shock proteins in clinical neurology. Eur. Neurol. 2011, 66, 65–69. [Google Scholar] [CrossRef]
- Ivanov, I.V.; Mappes, T.; Schaupp, P.; Lappe, C.; Wahl, S. Ultraviolet radiation oxidative stress affects eye health. J. Biophotonics 2018, 11, e201700377. [Google Scholar] [CrossRef] [Green Version]
- Stagos, D.; Chen, Y.; Cantore, M.; Jester, J.V.; Vasiliou, V. Corneal aldehyde dehydrogenases: Multiple functions and novel nuclear localization. Brain Res. Bull. 2010, 81, 211–218. [Google Scholar] [CrossRef] [Green Version]
- Piatigorsky, J. Lens and cornea: The “refracton hypothesis”. Semin. Cell. Dev. Biol. 2008, 19, 69–70. [Google Scholar] [CrossRef] [PubMed]
- Berthoud, V.M.; Beyer, E.C. Oxidative stress, lens gap junctions, and cataracts. Antioxid. Redox Signal. 2009, 11, 339–353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crouch, R.K.; Patrick, J.; Goosey, J.; Coles, W.H. The effect of age on corneal and lens superoxide dismutase. Curr. Eye Res. 1984, 3, 1119–1123. [Google Scholar] [CrossRef] [PubMed]
- Rajkumar, S.; Vasavada, A.R.; Praveen, M.R.; Ananthan, R.; Reddy, G.B.; Tripathi, H.; Ganatra, D.A.; Arora, A.I.; Patel, A.R. Exploration of molecular factors impairing superoxide dismutase isoforms activity in human senile cataractous lenses. Investig. Ophthalmol. Vis. Sci. 2013, 54, 6224–6233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joyce, N.C.; Harris, D.L.; Zhu, C.C. Age-related gene response of human corneal endothelium to oxidative stress and DNA damage. Investig. Ophthalmol. Vis. Sci. 2011, 52, 1641–1649. [Google Scholar] [CrossRef] [Green Version]
- Reddan, J.R.; Steiger, C.A.; Dziedzic, D.C.; Gordon, S.R. Regional differences in the distribution of catalase in the epithelium of the ocular lens. Cell. Mol. Biol. 1996, 42, 209–219. [Google Scholar]
- Sepasi Tehrani, H.; Moosavi-Movahedi, A.A. Catalase and its mysteries. Prog. Biophys. Mol. Biol. 2018, 140, 5–12. [Google Scholar] [CrossRef]
- Wahlig, S.; Lovatt, M.; Mehta, J.S. Functional role of peroxiredoxin 6 in the eye. Free Radic. Biol. Med. 2018, 126, 210–220. [Google Scholar] [CrossRef]
- Shibata, S.; Shibata, N.; Shibata, T.; Sasaki, H.; Singh, D.P.; Kubo, E. The role of Prdx6 in the protection of cells of the crystalline lens from oxidative stress induced by UV exposure. Jpn. J. Ophthalmol. 2016, 60, 408–418. [Google Scholar] [CrossRef] [Green Version]
- Ganea, E.; Harding, J.J. Glutathione-related enzymes and the eye. Curr. Eye Res. 2006, 31, 1–11. [Google Scholar] [CrossRef]
- Reddy, V.N.; Giblin, F.J.; Lin, L.R.; Dang, L.; Unakar, N.J.; Musch, D.C.; Boyle, D.L.; Takemoto, L.J.; Ho, Y.S.; Knoernschild, T.; et al. Glutathione peroxidase-1 deficiency leads to increased nuclear light scattering, membrane damage, and cataract formation in gene-knockout mice. Investig. Ophthalmol. Vis. Sci. 2001, 42, 3247–3255. [Google Scholar]
- Cardoso, B.R.; Hare, D.J.; Bush, A.I.; Roberts, B.R. Glutathione peroxidase 4: A new player in neurodegeneration? Mol. Psychiatry 2017, 22, 328–335. [Google Scholar] [CrossRef] [Green Version]
- Choudhary, S.; Xiao, T.; Vergara, L.A.; Srivastava, S.; Nees, D.; Piatigorsky, J.; Ansari, N.H. Role of aldehyde dehydrogenase isozymes in the defense of rat lens and human lens epithelial cells against oxidative stress. Investig. Ophthalmol. Vis. Sci. 2005, 46, 259–267. [Google Scholar] [CrossRef] [Green Version]
- Sax, C.M.; Salamon, C.; Kays, W.T.; Guo, J.; Yu, F.X.; Cuthbertson, R.A.; Piatigorsky, J. Transketolase is a major protein in the mouse cornea. J. Biol. Chem. 1996, 271, 33568–33574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rath, P.C. Models, Molecules and Mechanisms in Biogerontology: Physiological Abnormalities, Diseases and Interventions; Springer: Singapore, 2019; Chapter XL; p. 436. [Google Scholar] [CrossRef]
- Cejková, J.; Vejrazka, M.; Pláteník, J.; Stípek, S. Age-related changes in superoxide dismutase, glutathione peroxidase, catalase and xanthine oxidoreductase/xanthine oxidase activities in the rabbit cornea. Exp. Gerontol. 2004, 39, 1537–1543. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.; Worley, B.L.; Phaëton, R.; Hempel, N. Extracellular Glutathione Peroxidase GPx3 and Its Role in Cancer. Cancers 2020, 12, 2197. [Google Scholar] [CrossRef] [PubMed]
- Martín-Alonso, J.M.; Ghosh, S.; Coca-Prados, M. Cloning of the bovine plasma selenium-dependent glutathione peroxidase (GP) cDNA from the ocular ciliary epithelium: Expression of the plasma and cellular forms within the mammalian eye. J. Biochem. 1993, 114, 284–291. [Google Scholar] [CrossRef] [PubMed]
- Yildirim, Z.; Ucgun, N.I.; Yildirim, F. The role of oxidative stress and antioxidants in the pathogenesis of age-related macular degeneration. Clinics 2011, 66, 743–746. [Google Scholar] [CrossRef]
- Tan, S.M.; Stefanovic, N.; Tan, G.; Wilkinson-Berka, J.L.; de Haan, J.B. Lack of the antioxidant glutathione peroxidase-1 (GPx1) exacerbates retinopathy of prematurity in mice. Investig. Ophthalmol. Vis. Sci. 2013, 54, 555–562. [Google Scholar] [CrossRef] [Green Version]
- Tokarz, P.; Kaarniranta, K.; Blasiak, J. Role of antioxidant enzymes and small molecular weight antioxidants in the pathogenesis of age-related macular degeneration (AMD). Biogerontology 2013, 14, 461–482. [Google Scholar] [CrossRef] [Green Version]
- Donato, L.; Scimone, C.; Alibrandi, S.; Nicocia, G.; Rinaldi, C.; Sidoti, A.; D’Angelo, R. Discovery of GLO1 New Related Genes and Pathways by RNA-Seq on A2E-Stressed Retinal Epithelial Cells Could Improve Knowledge on Retinitis Pigmentosa. Antioxidants 2020, 9, 416. [Google Scholar] [CrossRef] [PubMed]
- Chidlow, G.; Wood, J.P.; Knoops, B.; Casson, R.J. Expression and distribution of peroxiredoxins in the retina and optic nerve. Brain. Struct. Funct. 2016, 221, 3903–3925. [Google Scholar] [CrossRef] [Green Version]
- Andley, U.P. Crystallins in the eye: Function and pathology. Prog. Retin. Eye Res. 2007, 26, 78–98. [Google Scholar] [CrossRef]
- Kannan, R.; Sreekumar, P.G.; Hinton, D.R. Alpha crystallins in the retinal pigment epithelium and implications for the pathogenesis and treatment of age-related macular degeneration. Biochim. Biophys. Acta. 2016, 1860, 258–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vasák, M.; Hasler, D.W. Metallothioneins: New functional and structural insights. Curr. Opin. Chem. Biol. 2000, 4, 177–183. [Google Scholar] [CrossRef]
- Bell, S.G.; Vallee, B.L. The metallothionein/thionein System: An Oxidoreductive Metabolic Zinc Link. ChemBioChem 2009, 10, 55–62. [Google Scholar] [CrossRef]
- Kägi, J.H. Overview of Metallothionein. Methods Enzymol. 1991, 205, 613–626. [Google Scholar] [CrossRef]
- Shaw, C.F.; Savas, M.M.; Petering, D.H. Ligand substitution and sulfhydryl reactivity of metallothionein. Methods Enzymol. 1991, 205, 401–414. [Google Scholar] [CrossRef]
- Sutherland, D.E.K.; Summers, K.L.; Stillman, M.J. Noncooperative Metalation of Metallothionein 1a and Its Isolated Domains with Zinc. Biochemistry 2012, 51, 6690–6700. [Google Scholar] [CrossRef]
- Ngu, T.T.; Stillman, M.J. Metal-binding mechanisms in metallothioneins. Dalton Trans. 2009, 5425–5433. [Google Scholar] [CrossRef]
- Krężel, A.; Maret, W. The Functions of Metamorphic Metallothioneins in Zinc and Copper Metabolism. Int. J. Mol. Sci. 2017, 18, 1237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Maret, W. Human metallothionein metallomics. J. Anal. At. Spectrom. 2008, 23, 1055–1062. [Google Scholar] [CrossRef]
- Moleirinho, A.; Carneiro, J.; Matthiesen, R.; Silva, R.M.; Amorim, A.; Azevedo, L. Gains, losses and changes of function after gene duplication: Study of the metallothionein family. PLoS ONE 2011, 6, e18487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vasákm, M. Advances in metallothionein structure and functions. J. Trace Elem. Med. Biol. 2005, 19, 13–17. [Google Scholar] [CrossRef] [PubMed]
- Quaife, C.J.; Findley, S.D.; Erickson, J.C.; Froelick, G.J.; Kelly, E.J.; Zambrowicz, B.P.; Palmiter, R.D. Induction of a new metallothionein isoform (Mt-Iv) occurs during differentiation of stratified squamous epithelia. Biochemistry 1994, 33, 7250–7259. [Google Scholar] [CrossRef]
- Moffatt, P.; Seguin, C. Expression of the gene encoding metallothionein-3 in organs of the reproductive system. DNA Cell. Biol. 1998, 17, 501–510. [Google Scholar] [CrossRef]
- Uchida, Y.; Takio, K.; Titani, K.; Ihara, Y.; Tomonaga, M. The growth inhibitory factor that is deficient in the Alzheimers-disease brain is a 68-amino acid metallothionein-like protein. Neuron 1991, 7, 337–347. [Google Scholar] [CrossRef]
- Bremner, I. Nutritional and physiological significance of metallothionein. Experientia Suppl. 1987, 52, 81–107. [Google Scholar] [CrossRef]
- Maret, W.; Li, Y. Coordination dynamics of zinc in proteins. Chem. Rev. 2009, 109, 4682–4707. [Google Scholar] [CrossRef]
- Zeng, J.; Vallee, B.L.; Kagi, J.H. Zn transfer from transcription factor IIIA fingers to thionein clusters. Proc. Natl. Acad. Sci. USA 1991, 88, 9984–9988. [Google Scholar] [CrossRef] [Green Version]
- Maret, W. Metallothionein/disulfide interactions, oxidative stress, and the mobilization of cellular zinc. Neurochem. Int. 1995, 27, 111–117. [Google Scholar] [CrossRef]
- Kang, Y.J. Metallothionein redox cycle and function. Exp. Biol. Med. 2006, 231, 1459–1467. [Google Scholar] [CrossRef] [PubMed]
- Formigari, A.; Irato, P.; Santon, A. Zinc, antioxidant systems and metallothionein in metal mediated-apoptosis: Biochemical and cytochemical aspects. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2007, 146, 443–459. [Google Scholar] [CrossRef] [PubMed]
- Templeton, D.M.; Cherian, M.G. Toxicological significance of metallothionein. Methods Enzymol. 1991, 205, 11–24. [Google Scholar] [CrossRef] [PubMed]
- Klaassen, C.D.; Liu, J.; Diwan, B.A. Metallothionein protection of cadmium toxicity. Toxicol. Appl. Pharmacol. 2009, 238, 215–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nordberg, M.; Nordberg, G.F. Toxicological aspects of metallothionein. Cell. Mol. Biol. 2000, 46, 451–463. [Google Scholar] [PubMed]
- Ngu, T.T.; Stillman, M.J. Arsenic binding to human metallothionein. J. Am. Chem. Soc. 2006, 128, 12473–12483. [Google Scholar] [CrossRef]
- Gumulec, J.; Masarik, M.; Krizkova, S.; Adam, V.; Hubalek, J.; Hrabeta, J.; Eckschlager, T.; Stiborova, M.; Kizek, R. Insight to physiology and pathology of zinc(II) ions and their actions in breast and prostate carcinoma. Curr. Med. Chem. 2011, 18, 5041–5051. [Google Scholar] [CrossRef] [Green Version]
- Babula, P.; Masarik, M.; Adam, V.; Eckschlager, T.; Stiborova, M.; Trnkova, L.; Skutkova, H.; Provaznik, I.; Hubalek, J.; Kizek, R. Mammalian metallothioneins: Properties and functions. Metallomics 2012, 4, 739–750. [Google Scholar] [CrossRef]
- Maret, W.; Vallee, B.L. Thiolate ligands in metallothionein confer redox activity on zinc clusters. Proc. Natl. Acad. Sci. USA 1998, 95, 3478–3482. [Google Scholar] [CrossRef] [Green Version]
- Maret, W. The function of zinc metallothionein: A link between cellular zinc and redox state. J. Nutr. 2000, 130 (Suppl. 5), 1455S–1458S. [Google Scholar] [CrossRef]
- Chiaverini, N.; De Ley, M. Protective effect of metallothionein on oxidative stress-induced DNA damage. Free Radic. Res. 2010, 44, 605–613. [Google Scholar] [CrossRef]
- Ruttkay-Nedecky, B.; Nejdl, L.; Gumulec, J.; Zitka, O.; Masarik, M.; Eckschlager, T.; Stiborova, M.; Adam, V.; Kizek, R. The role of metallothionein in oxidative stress. Int. J. Mol. Sci. 2013, 14, 6044–6066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, L.; Koropatnick, J.; Cherian, M.G. Metallothionein protects DNA from copper-induced but not iron-induced cleavage in vitro. Chem. Biol. Interact. 1995, 96, 143–155. [Google Scholar] [CrossRef]
- Valko, M.; Jomova, K.; Rhodes, C.J.; Kuča, K.; Musílek, K. Redox- and non-redox-metal-induced formation of free radicals and their role in human disease. Arch. Toxicol. 2016, 90, 1–37. [Google Scholar] [CrossRef] [PubMed]
- Lazo, J.S.; Pitt, B.R. Metallothioneins and cell death by anticancer drugs. Ann. Rev. Pharmacol. Toxicol. 1995, 35, 635–653. [Google Scholar] [CrossRef]
- Hidalgo, J.; Aschner, M.; Zatta, P.; Vasak, M. Roles of the metallothionein family of proteins in the central nervous system. Brain Res. Bull. 2001, 55, 133–145. [Google Scholar] [CrossRef]
- Thirumoorthy, N.; Sunder, A.S.; Kumar, K.M.; Kumar, M.S.; Ganesh, G.; Chatterjee, M. A review of metallothionein isoforms and their role in pathophysiology. World J. Surg. Oncol. 2011, 9, 54. [Google Scholar] [CrossRef] [Green Version]
- Shamsi, T.; Fatima, S. Metallothionein: Classification, biochemical features and clinical applications. J. Prot. Proteom. 2014, 5, 25–33. [Google Scholar]
- Jakovac, H.; Kezele, T.G.; Radošević-Stašić, B. Expression Profiles of Metallothionein I/II and Megalin in Cuprizone Model of De- and Remyelination. Neuroscience 2018, 388, 69–86. [Google Scholar] [CrossRef]
- Ebadi, M.; Iversen, P.L.; Hao, R.; Cerutis, D.R.; Rojas, P.; Happe, H.K.; Murrin, L.C.; Pfeiffer, R.F. Expression and regulation of brain metallothionein. Neurochem. Int. 1995, 27, 1–22. [Google Scholar] [CrossRef]
- Yamada, M.; Hayashi, S.; Hozumi, I.; Inuzuka, T.; Tsuji, S.; Takahashi, H. Subcellular localization of growth inhibitory factor in rat brain: Light and electron microscopic immunohistochemical studies. Brain Res. 1996, 735, 257–264. [Google Scholar] [CrossRef]
- Aschner, M.; Cherian, M.G.; Klaassen, C.D.; Palmiter, R.D.; Erickson, J.C.; Bush, A.I. Metallothioneins in brain—The role in physiology and pathology. Toxicol. Appl. Pharmacol. 1997, 142, 229–242. [Google Scholar] [CrossRef] [PubMed]
- Carrasco, J.; Penkowa, M.; Giralta, M.; Camats, J.; Molinero, A.; Campbell, I.L.; Palmiter, R.D.; Hidalgo, J. Role of metallothionein-III following central nervous system damage. Neurobiol. Dis. 2003, 13, 22–36. [Google Scholar] [CrossRef]
- West, A.K.; Hidalgo, J.; Eddins, D.; Levin, E.D.; Aschner, M. Metallothionein in the central nervous system: Roles in protection, regeneration and cognition. Neurotoxicology 2008, 29, 488–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coyle, P.; Philcox, J.C.; Carey, L.C.; Rofe, A.M. Metallothionein: The multipurpose protein. Cell. Mol. Life. Sci. 2002, 59, 627–647. [Google Scholar] [CrossRef] [PubMed]
- Maret, W. Redox biochemistry of mammalian metallothioneins. J. Biol. Inorg. Chem. 2011, 16, 1079–1086. [Google Scholar] [CrossRef] [PubMed]
- Mocchegiani, E.; Giacconi, R.; Cipriano, C.; Costarelli, L.; Muti, E.; Tesei, S.; Giuli, C.; Papa, R.; Marcellini, F.; Mariani, E.; et al. Zinc, metallothioneins, and longevity–effect of zinc supplementation: Zincage study. Ann. N. Y. Acad. Sci. 2007, 1119, 129–146. [Google Scholar] [CrossRef]
- Levenson, C.W.; Morris, D. Zinc and neurogenesis: Making new neurons from development to adulthood. Adv. Nutr. 2011, 2, 96–100. [Google Scholar] [CrossRef]
- Chung, R.S.; Penkowa, M.; Dittmann, J.; King, C.E.; Bartlett, C.; Asmussen, J.W.; Hidalgo, J.; Carrasco, J.; Leung, Y.K.; Walker, A.K.; et al. Redefining the role of metallothionein within the injured brain: Extracellular metallothioneins play an important role in the astrocyte-neuron response to injury. J. Biol. Chem. 2008, 283, 15349–15358. [Google Scholar] [CrossRef] [Green Version]
- Chung, R.S.; Hidalgo, J.; West, A.K. New insight into the molecular pathways of metallothionein-mediated neuroprotection and regeneration. J. Neurochem. 2008, 104, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Ambjørn, M.; Asmussen, J.W.; Lindstam, M.; Gotfryd, K.; Jacobsen, C.; Kiselyov, V.V.; Moestrup, S.K.; Penkowa, M.; Bock, E.; Berezin, V. Metallothionein and a peptide modeled after metallothionein, EmtinB, induce neuronal differentiation and survival through binding to receptors of the low-density lipoprotein receptor family. J. Neurochem. 2008, 104, 21–37. [Google Scholar] [CrossRef] [PubMed]
- Asmussen, J.W.; Sperling, M.L.V.; Penkowa, M. Intraneuronal signaling pathways of metallothionein. J. Neurosci. Res. 2009, 87, 2926–2936. [Google Scholar] [CrossRef] [PubMed]
- West, A.K.; Leung, J.Y.K.; Chung, R.S. Neuroprotection and regeneration by extracellular metallothionein via lipoprotein-receptor-related proteins. J. Biol. Inorg. Chem. 2011, 16, 1115. [Google Scholar] [CrossRef] [PubMed]
- Auderset, L.; Landowski, L.M.; Foa, L.; Young, K.M. Low density lipoprotein receptor related proteins as regulators of neural stem and progenitor cell function. Stem Cells Int. 2016, 2108495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimoda, R.; Achanzar, W.E.; Qu, W.; Nagamine, T.; Takagi, H.; Mori, M.; Waalkes, M.P. Metallothionein is a potential negative regulator of apoptosis. Toxicol. Sci. 2003, 73, 294–300. [Google Scholar] [CrossRef] [Green Version]
- Santon, A.; Albergoni, V.; Sturniolo, G.C.; Irato, P. Evaluation of MT expression and detection of apoptotic cells in LEC rat kidneys. Biochim. Biophys. Acta 2004, 1688, 223–231. [Google Scholar] [CrossRef]
- Telford, W.G.; Fraker, P.J. Preferential induction of apoptosis in mouse CD4(+)CD8(+)alphabeta- tcr(lo)CD3-epsilon(lo) thymocytes by zinc. J. Cell. Physiol. 1995, 164, 259–270. [Google Scholar] [CrossRef]
- Perry, D.K.; Smyth, M.J.; Stennicke, H.R.; Salvesen, G.S.; Duriez, P.; Poirier, G.G.; Hannun, Y.A. Zinc is a potent inhibitor of the apoptotic protease, caspase-3—A novel target for zinc in the inhibition of apoptosis. J. Biol. Chem. 1997, 272, 18530–18533. [Google Scholar] [CrossRef] [Green Version]
- Bozym, R.A.; Chimienti, F.; Giblin, L.J.; Gross, G.W.; Korichneva, I.; Li, Y.; Libert, S.; Maret, W.; Parviz, M.; Frederickson, C.J.; et al. Free zinc ions outside a narrow concentration range are toxic to a variety of cells in vitro. Exp. Biol. Med. (Maywood) 2010, 235, 741–750. [Google Scholar] [CrossRef] [Green Version]
- Cano-Gauci, D.F.; Sarkar, B. Reversible zinc exchange between metallothionein and the estrogen receptor zinc finger. FEBS Lett. 1996, 386, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Moffatt, P.; Denizeau, F. Metallothionein in physiological and physiopathological processes. Drug Metab. Rev. 1997, 29, 261–307. [Google Scholar] [CrossRef] [PubMed]
- Maret, W.; Jacob, C.; Vallee, B.L.; Fischer, E.H. Inhibitory sites in enzymes: Zinc removal and reactivation by thionein. Proc. Natl. Acad. Sci. USA 1999, 96, 1936–1940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meplan, C.; Verhaegh, G.; Richard, M.J.; Hainaut, P. Metal ions as regulators of the conformation and function of the tumour suppressor protein p53: Implications for carcinogenesis. Proc. Nutr. Soc. 1999, 58, 565–571. [Google Scholar] [CrossRef] [Green Version]
- Meplan, C.; Richard, M.J.; Hainaut, P. Metalloregulation of the tumor suppressor protein p53: Zinc mediates the renaturation of p53 after exposure to metal chelators in vitro and in intact cells. Oncogene 2000, 19, 5227–5236. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Mageed, A.B.; Agrawal, K.C. Activation of nuclear factor kappa B: Potential role in metallothionein-mediated mitogenic response. Cancer Res. 1998, 58, 2335–2338. [Google Scholar]
- Kim, C.H.; Kim, J.H.; Lee, J.; Ahn, Y.S. Zinc-Induced NF-kappa B inhibition can be modulated by changes in the intracellular metallothionein level. Toxicol. Appl. Pharmacol. 2003, 190, 189–196. [Google Scholar] [CrossRef]
- Butcher, H.L.; Kennette, W.A.; Collins, O.; Zalups, R.K.; Koropatnick, J. Metallothionein mediates the level and activity of nuclear factor kappa B in murine fibroblasts. J. Pharmacol. Exp. Ther. 2004, 310, 589–598. [Google Scholar] [CrossRef] [Green Version]
- Tsangaris, G.T.; Vamvoukakis, J.; Politis, I.; Kattamis, A.C.; Tzortzatou-Stathopoulou, F. Metallothionein expression prevents apoptosis. II: Evaluation of the role of metallothionein expression on the chemotherapy-induced apoptosis during the treatment of acute leukemia. Anticancer Res. 2000, 20, 4407–4411. [Google Scholar]
- Si, M.; Lang, J. The roles of metallothioneins in carcinogénesis. J. Hematol. Oncol. 2018, 11, 107. [Google Scholar] [CrossRef]
- Krezel, A.; Maret, W. Different redox states of metallothionein/thionein in biological tissue. Biochem. J. 2007, 402, 551–558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maret, W. Molecular aspects of human cellular zinc homeostasis: Redox control of zinc potentials and zinc signals. Biometals 2009, 22, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Iglesias, H.; Alvarez, L.; García, M.; Petrash, C.; Sanz-Medel, A.; Coca-Prados, M. Metallothioneins (MTs) in the human eye: A perspective article on the zinc-MT redox cycle. Metallomics 2014, 6, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Sutherland, D.E.; Stillman, M.J. The “magic numbers” of metallothionein. Metallomics 2011, 3, 444–463. [Google Scholar] [CrossRef]
- Krezel, A.; Hao, Q.; Maret, W. The zinc/thiolate redox biochemistry of metallothionein and the control of zinc ion fluctuations in cell signaling. Arch. Biochem. Biophys. 2007, 463, 188–200. [Google Scholar] [CrossRef]
- Chen, Y.; Maret, W. Catalytic selenols couple the redox cycles of metallothionein and glutathione. Eur. J. Biochem. 2001, 268, 3346–3353. [Google Scholar] [CrossRef]
- Maret, W.; Krezel, A. Cellular zinc and redox buffering capacity of metallothionein/thionein in health and disease. Mol. Med. 2007, 13, 371–375. [Google Scholar] [CrossRef]
- Laukens, D.; Waeytens, A.; De Bleser, P.; Cuvelier, C.; De Vos, M. Human metallothionein expression under normal and pathological conditions: Mechanisms of gene regulation based on in silico promoter analysis. Crit. Rev. Eukaryot. Gene Expr. 2009, 19, 301–317. [Google Scholar] [CrossRef]
- Schmidt, C.J.; Hamer, D.H. Cell specificity and an effect of ras on human metallothionein gene expression. Proc. Natl. Acad. Sci. USA 1986, 83, 3346–3350. [Google Scholar] [CrossRef] [Green Version]
- Varshney, U.; Jahroudi, N.; Foster, R.; Gedamu, L. Structure, organization, and regulation of human metallothionein IF gene: Differential and cell-type-specific expression in response to heavy metals and glucocorticoids. Mol. Cell. Biol. 1986, 6, 26–37. [Google Scholar] [CrossRef] [Green Version]
- Ghoshal, K.; Jacob, S.T. Regulation of metallothionein gene expression. Prog. Nucleic. Acid. Res. Mol. Biol. 2001, 66, 357–384. [Google Scholar] [CrossRef] [PubMed]
- Haq, F.; Mahoney, M.; Koropatnick, J. Signaling events for metallothionein induction. Mutat. Res. 2003, 533, 211–226. [Google Scholar] [CrossRef] [PubMed]
- Borghesi, L.A.; Lynes, M.A. Stress proteins as agents of immunological change: Some lessons from metallothionein. Cell Stress Chaperones 1996, 1, 99–108. [Google Scholar] [CrossRef]
- Takahashi, S. Molecular functions of metallothionein and its role in hematological malignancies. J. Hematol. Oncol. 2012, 5, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakulsak, N. Metallothionein: An overview on its metal homeostatic regulation in mammals. Int. J. Morphol. 2012, 30, 1007–1012. [Google Scholar] [CrossRef] [Green Version]
- Kimura, T.; Kambe, T. The functions of metallothionein and ZIP and ZnT transporters: An overview and perspective. Int. J. Mol. Sci. 2016, 17, 336–357. [Google Scholar] [CrossRef] [Green Version]
- Westin, G.; Schaffner, W. A zinc responsive factor interacts with a metal regulated enhancer element (MRE) of the mouse metallothionein I gene. EMBO J. 1988, 7, 3763–3770. [Google Scholar] [CrossRef]
- Dalton, T.; Palmiter, R.D.; Andrews, G.K. Transcriptional induction of the mouse metallothionein I gene in hydrogen peroxide treated Hepa cells involves a composite major late transcription factor/antioxidant response element and metal response promoter elements. Nucleic Acids Res. 1994, 22, 5016–5023. [Google Scholar] [CrossRef] [Green Version]
- Murphy, B.J.; Andrews, G.K.; Bittel, D.; Discher, D.J.; McCue, J.; Green, C.J.; Yanovsky, M.; Giaccia, A.; Sutherland, R.M.; Laderoute, K.R.; et al. Activation of metallothionein gene expression by hypoxia involves metal response elements and metal transcription factor 1. Cancer Res. 1999, 59, 1315–1322. [Google Scholar]
- Günther, V.; Lindert, U.; Schaffner, W. The taste of heavy metals: Gene regulation by MTF1. Biochim. Biophys. Acta 2012, 1823, 1416–1425. [Google Scholar] [CrossRef] [Green Version]
- Andrews, G.K.; Geiser, J. Expression of the mouse metallothionein-I and-II genes provides a reproductive advantage during maternal dietary Zn deficiency. J. Nutr. 1999, 129, 1643–1648. [Google Scholar] [CrossRef] [PubMed]
- Klassen, R.B.; Crenshaw, K.; Kozyraki, R.; Verroust, P.J.; Tio, L.; Atrian, S.; Allen, P.L.; Hammond, T.G. Megalin mediates renal uptake of heavy metalmetallothionein complexes. Am. J. Physiol. Renal. Physiol. 2004, 287, F393–F403. [Google Scholar] [CrossRef]
- Samson, S.L.A.; Gedamu, L. Molecular analyses of metallothionein gene regulation. Prog. Nucleic Acid Res. Mol. Biol. 1998, 59, 257–288. [Google Scholar] [CrossRef] [PubMed]
- Andrews, G.K. Regulation of metallothionein gene expression by oxidative stress and metal ions. Biochem. Pharmacol. 2000, 59, 95–104. [Google Scholar] [CrossRef]
- Kelly, E.; Sandgren, E.; Brinster, R.; Palmiter, R. A pair of adjacent glucocorticoid response elements regulate expression of two mouse metallothionein genes. Proc. Natl. Acad. Sci. USA 1997, 94, 10045–10050. [Google Scholar] [CrossRef] [Green Version]
- Davis, S.; Cousins, R. Metallothionein expression in animals: A physiological perspective on function. J. Nutr. 2000, 130, 1085–1088. [Google Scholar] [CrossRef] [Green Version]
- Ivashkiv, L.B.; Hu, X. Signaling by STATs. Arthritis Res Ther. 2004, 6, 159–168. [Google Scholar] [CrossRef] [Green Version]
- De, S.K.; McMaster, M.T.; Andrews, G.K. Endotoxin induction of murine metallothionein gene expression. J. Biol. Chem. 1990, 265, 15267–15274. [Google Scholar] [CrossRef]
- Pfitzner, E.; Kliem, S.; Baus, D.; Litterst, C.M. The Role of STATs in Inflammation and Inflammatory Diseases. Curr Pharm Des. 2004, 10, 2839–2850. [Google Scholar] [CrossRef]
- Lee, D.K.; Carrasco, J.; Hidalgo, J.; Andrews, G.K. Identification of a signal transducer and activator of transcription (STAT) binding site in the mouse metallothionein-I promoter involved in interleukin-6-induced gene expression. Biochem. J. 1999, 337, 59–65. [Google Scholar] [CrossRef]
- Kanekiyo, M.; Itoh, N.; Kawasaki, A.; Matsuyama, A.; Matsuda, K.; Nakanishi, T.; Tanaka, K. Metallothionein modulates lipopolysaccharide-stimulated tumour necrosis factor expression in mouse peritoneal macrophages. Biochem. J. 2002, 361, 363–369. [Google Scholar] [CrossRef] [PubMed]
- Murphy, B.J.; Kimura, T.; Sato, B.G.; Shi, Y.; Andrews, G.K. Metallothionein induction by hypoxia involves cooperative interactions between metal-responsive transcription factor-1 and hypoxia-inducible transcription factor- 1alpha. Mol. Cancer Res. 2008, 6, 483–490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrews, G.K.; Lee, D.K.; Ravindra, R.; Lichtlen, P.; Sirito, M.; Sawadogo, M.; Schaffner, W. The transcription factors MTF 1 and USF1 cooperate to regulate mouse metallothionein I expression in response to the essential metal zinc in visceral endoderm cells during early development. EMBO J. 2001, 20, 1114–1122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- LaRochelle, O.; Labbe, S.; Harrisson, J.F.; Simard, C.; Tremblay, V.; St-Gelais, G.; Govindan, M.V.; Séguin, C. Nuclear factor 1 and metal transcription factor 1 synergistically activate the mouse metallothionein 1 gene in response to metal ions. J. Biol. Chem. 2008, 283, 8190–8201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imoto, A.; Okada, M.; Okazaki, T.; Kitasato, H.; Harigae, H.; Takahashi, S. Metallothionein 1 isoforms and vimentin are direct PU.1 downstream target genes in leukemia cells. J. Biol. Chem. 2010, 285, 10300–10309. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, S.; Nakano, H.; Takahashi, S. Epigenetic regulation of the metallothionein 1A promoter by PU.1 during differentiation of THP 1 cells. Biochem. Biophys. Res. Commun. 2013, 433, 349–353. [Google Scholar] [CrossRef]
- Tang, C.M.; Westling, J.; Seto, E. trans repression of the human metallothionein IIA gene promoter by PZ120, a novel 120 kilo¬dalton zinc finger protein. Mol. Cell. Biol. 1999, 19, 680–689. [Google Scholar] [CrossRef] [Green Version]
- Yin, H.; Smith, M.; Glass, J. Stable expression of C/EBPalpha in prostate cancer cells down regulates metallothionein and increases zinc induced toxicity. Prostate 2005, 62, 209–216. [Google Scholar] [CrossRef]
- Ghoshal, K.; Li, Z.; Jacob, S.T. Overexpression of the large subunit of the protein Ku suppresses metallothionein I induction by heavy metals. Proc. Natl. Acad. Sci. USA 1998, 95, 10390–10395. [Google Scholar] [CrossRef] [Green Version]
- Majumder, S.; Kutay, H.; Datta, J.; Summers, D.; Jacob, S.T.; Kalpana-Ghoshal, K. Epigenetic regulation of metallothionein-i gene expression: Differential regulation of methylated and unmethylated promoters by DNA methyltransferases and methyl CpG binding proteins. J. Cell. Biochem. 2006, 97, 1300–1316. [Google Scholar] [CrossRef]
- Ghoshal, K.; Datta, J.; Majumder, S.; Bai, S.; Dong, X.; Parthun, M.; Jacob, S.T. Inhibitors of Histone Deacetylase and DNA Methyltransferase synergistically activate the methylated metallothionein I promoter by activating the transcription factor MTF-1 and forming an open chromatin structure. Mol. Cell. Biol. 2002, 22, 8302–8319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKim, J.M.; Choudhuri, S.; Klaassen, C.D. In vitro degradation of apo-, zinc-, and cadmium metallothionein by cathepsins B, C, and D. Toxicol. Appl. Pharmacol. 1992, 116, 117–124. [Google Scholar] [CrossRef]
- Klaassen, C.D.; Choudhuri, S.; McKim, J.M.; Lehman-McKeeman, L.D.; Kershaw, W.C. In vitro and in vivo studies on the degradation of metallothionein. Environ. Health Perspect. 1994, 102, 141–146. [Google Scholar] [CrossRef]
- Shcherbik, N.; Pestov, D.G. The Impact of Oxidative Stress on Ribosomes: From Injury to Regulation. Cells 2019, 8, 1379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, A.; Kukreti, R.; Saso, L.; Kukreti, S. Oxidative Stress: A Key Modulator in Neurodegenerative Diseases. Molecules 2019, 24, 1583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bilkis, I.; Silman, I.; Weiner, L. Generation of Reactive Oxygen Species by Photosensitizers and their Modes of Action on Proteins. Curr. Med. Chem. 2018, 25, 5528–5539. [Google Scholar] [CrossRef]
- Brem, R.; Guven, M.; Karran, P. Oxidatively-generated damage to DNA and proteins mediated by photosensitized UVA. Free Radic. Biol. Med. 2017, 107, 101–109. [Google Scholar] [CrossRef] [Green Version]
- Hauck, A.K.; Huang, Y.; Hertzel, A.V.; Bernlohr, D.A. Adipose oxidative stress and protein carbonylation. J. Biol. Chem. 2019, 294, 1083–1088. [Google Scholar] [CrossRef] [Green Version]
- Kamari, F.; Hallaj, S.; Dorosti, F.; Alinezhad, F.; Taleschian-Tabrizi, N.; Farhadi, F.; Aslani, H. Phototoxicity of environmental radiations in human lens: Revisiting the pathogenesis of UV-induced cataract. Graefes Arch. Clin. Exp. Ophthalmol. 2019, 257, 2065–2077. [Google Scholar] [CrossRef]
- Truscott, R.J.W.; Friedrich, M.G. Molecular processes implicated in human age-related nuclear cataract. IOVS 2019, 60, 5007–5021. [Google Scholar] [CrossRef] [Green Version]
- Richardson, R.B.; Ainsbury, E.A.; Prescott, C.R.; Lovicu, F.J. Etiology of posterior subcapsular cataracts based on a review of risk factors including, aging, diabetes, and ionizing radiation. Int. J. Radiat. Biol. 2020, 96, 1339–1361. [Google Scholar] [CrossRef] [PubMed]
- van Kuijk, F.J. Effects of ultraviolet light on the eye: Role of protective glasses. Environ. Health Perspect. 1991, 96, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Roberts, J.E. Ocular phototoxicity. J. Photochem. Photobiol. B 2001, 64, 136–143. [Google Scholar] [CrossRef]
- Hamba, N.; Gerbi, A.; Tesfaye, S. Histopathological effects of ultraviolet radiation exposure on the ocular structures in animal studies—Literature review. Transl. Res. Anat. 2020, 100086. [Google Scholar] [CrossRef]
- Ahmad, A.; Ahsan, H. Biomarkers of inflammation and oxidative stress in ophthalmic disorders. J. Immunoass. Immunochem. 2020, 41, 257–271. [Google Scholar] [CrossRef]
- Deng, R.; Hua, X.; Li, J.; Chi, W.; Zhang, Z.; Lu, F.; Zhang, L.; Pflugfelder, S.C.; Li, D.Q. Oxidative stress markers induced by hyperosmolarity in primary human corneal epithelial cells. PLoS ONE 2015, 10, e0126561. [Google Scholar] [CrossRef] [Green Version]
- Kojima, T.; Wakamatsu, T.H.; Dogru, M.; Ogawa, Y.; Igarashi, A.; Ibrahim, O.M.; Inaba, T.; Shimizu, T.; Noda, S.; Obata, H.; et al. Age-related dysfunction of the lacrimal gland and oxidative stress: Evidence from the Cu,Zn-superoxide dismutase-1 (Sod1) knockout mice. Am. J. Pathol. 2012, 180, 1879–1896. [Google Scholar] [CrossRef]
- Ibrahim, O.M.; Dogru, M.; Matsumoto, Y.; Igarashi, A.; Kojima, T.; Wakamatsu, T.H.; Inaba, T.; Shimizu, T.; Shimazaki, J.; Tsubota, K. Oxidative stress induced age dependent meibomian gland dysfunction in Cu, Zn-superoxide dismutase-1 (Sod1) knockout mice. PLoS ONE 2014, 9, e99328. [Google Scholar] [CrossRef] [Green Version]
- Uchino, Y.; Kawakita, T.; Miyazawa, M.; Ishii, T.; Onouchi, H.; Yasuda, K.; Ogawa, Y.; Shimmura, S.; Ishii, N.; Tsubota, K. Oxidative stress induced inflammation initiates functional decline of tear production. PLoS ONE 2012, 7, e45805. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, S.; Shibuya, M.; Nakashima, H.; Hisamura, R.; Masuda, N.; Imagawa, T.; Uehara, M.; Tsubota, K. Involvement of oxidative stress on corneal epithelial alterations in a blink-suppressed dry eye. Investig. Ophthalmol. Vis. Sci. 2007, 48, 1552–1558. [Google Scholar] [CrossRef]
- Choi, W.; Lian, C.; Ying, L.; Kim, G.E.; You, I.C.; Park, S.H.; Yoon, K.C. Expression of Lipid Peroxidation Markers in the Tear Film and Ocular Surface of Patients with Non- Sjogren Syndrome: Potential Biomarkers for Dry Eye Disease. Curr. Eye Res. 2016, 41, 1143–1149. [Google Scholar] [CrossRef]
- Perez-Garmendia, R.; Lopez de Eguileta Rodriguez, A.; Ramos-Martinez, I.; Martínez Zuñiga, N.; Gonzalez-Salinas, R.; Quiroz-Mercado, H.; Zeneto, E.; Ramírez Hernández, E.; Hernández-Zimbrón, L.F. Interplay between oxidative stress, inflammation, and amyloidosis in the anterior segment of the eye; its pathological implications. Oxid. Med. Cell. Longev. 2020, 2020, 6286105. [Google Scholar] [CrossRef] [PubMed]
- Kojima, T.; Dogru, M.; Kawashima, M.; Nakamura, S.; Tsubota, K. Advances in the diagnosis and treatment of dry eye. Prog. Retin. Eye Res. 2020, 78, 100842. [Google Scholar] [CrossRef] [PubMed]
- Arden, G.B.; Sivaprasad, S. Hypoxia and oxidative stress in the causation of diabetic retinopathy. Curr. Diabetes Rev. 2011, 7, 291–304. [Google Scholar] [CrossRef] [PubMed]
- Son, S.M. Role of vascular reactive oxygen species in development of vascular abnormalities in diabetes. Diabetes Res. Clin. Pract. 2007, 77 (Suppl. 1), S65–S70. [Google Scholar] [CrossRef] [PubMed]
- Gupta, M.M.; Chari, S. Lipid peroxidation and antioxidant status in patients with diabetic retinopathy. Indian J. Physiol. Pharmacol. 2005, 49, 187–192. [Google Scholar]
- Robison, W.G., Jr.; Jacot, J.L.; Katz, M.L.; Glover, J.P. Retinal vascular changes induced by the oxidative stress of alpha-tocopherol deficiency contrasted with diabetic microangiopathy. J. Ocul. Pharmacol. Ther. 2000, 16, 109–120. [Google Scholar] [CrossRef]
- Martín-Gallán, P.; Carrascosa, A.; Gussinyé, M.; Domínguez, C. Biomarkers of diabetes-associated oxidative stress and antioxidant status in young diabetic patients with or without subclinical complications. Free Radic. Biol. Med. 2003, 34, 1563–1574. [Google Scholar] [CrossRef]
- Chen, Q.; Tang, L.; Xin, G.; Li, S.; Ma, L.; Xu, Y.; Zhuang, M.; Xiong, Q.; Wei, Z.; Xing, Z.; et al. Oxidative stress mediated by lipid metabolism contributes to high glucose-induced senescence in retinal pigment epithelium. Free Radic. Biol. Med. 2019, 130, 48–58. [Google Scholar] [CrossRef]
- Chen, B.H.; Jiang, D.Y.; Tang, L.S. Advanced glycation end-products induce apoptosis involving the signaling pathways of oxidative stress in bovine retinal pericytes. Life Sci. 2006, 79, 1040–1048. [Google Scholar] [CrossRef]
- Kowluru, R.A.; Abbas, S.N. Diabetes-induced mitochondrial dysfunction in the retina. Investig. Ophthalmol. Vis. Sci. 2003, 44, 5327–5334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baynes, J.W. Role of oxidative stress in development of complications in diabetes. Diabetes. 1991, 40, 405–412. [Google Scholar] [CrossRef] [PubMed]
- Chan, T.C.W.; Wilkinson-Berka, J.L.; Deliyanti, D.; Hunter, D.; Fung, A.; Liew, G.; White, A. The role of reactive oxygen species in the pathogenesis and treatment of retinal diseases. Exp. Eye Res. 2020, 201, 108255. [Google Scholar] [CrossRef] [PubMed]
- Kang, Q.; Yang, C. Oxidative stress and diabetic retinopathy: Molecular mechanisms, pathogenetic role and therapeutic implications. Redox Biol. 2020, 37, 101799. [Google Scholar] [CrossRef]
- Münzel, T.; Camici, G.G.; Maack, C.; Bonetti, N.R.; Fuster, V.; Kovacic, J.C. Impact of Oxidative Stress on the Heart and Vasculature: Part 2 of a 3-Part Series. J. Am. Coll. Cardiol. 2017, 70, 212–229. [Google Scholar] [CrossRef]
- Moran, E.P.; Wang, Z.; Chen, J.; Sapieha, P.; Smith, L.E.; Mam, J.X. Neurovascular cross talk in diabetic retinopathy: Pathophysiological roles and therapeutic implications. Am. J. Physiol. Heart Circ. Physiol. 2016, 311, H738–K749. [Google Scholar] [CrossRef] [Green Version]
- Zadeh, J.K.; Ruemmler, R.; Hartmann, E.K.; Ziebart, A.; Ludwig, M.; Patzak, A.; Xia, N.; Li, H.; Pfeiffer, N.; Gericke, A. Responses of retinal arterioles and ciliary arteries in pigs with acute respiratory distress syndrome (ARDS). Exp. Eye Res. 2019, 184, 152–161. [Google Scholar] [CrossRef]
- Domènech, E.B.; Marfany, G. The Relevance of Oxidative Stress in the Pathogenesis and Therapy of Retinal Dystrophies. Antioxidants 2020, 9, 347. [Google Scholar] [CrossRef] [Green Version]
- Benoist d’Azy, C.; Pereira, B.; Chiambaretta, F.; Dutheil, F. Oxidative and Anti-Oxidative Stress Markers in Chronic Glaucoma: A Systematic Review and Meta- Analysis. PLoS ONE 2016, 11, e0166915. [Google Scholar] [CrossRef]
- Gericke, A.; Mann, C.; Zadeh, J.K.; Musayeva, A.; Wolff, I.; Wang, M.; Pfeiffer, N.; Daiber, A.; Li, H.; Xia, N.; et al. Elevated Intraocular Pressure Causes Abnormal Reactivity of Mouse Retinal Arterioles. Oxid. Med. Cell. Longev. 2019, 29, 9736047. [Google Scholar] [CrossRef] [Green Version]
- Zanon-Moreno, V.; Marco-Ventura, P.; Lleo-Perez, A.; Pons-Vazquez, S.; Garcia-Medina, J.J.; Vinuesa-Silva, I.; Moreno-Nadal, M.A.; Pinazo-Duran, M.D. Oxidative stress in primary open-angle glaucoma. J. Glaucoma 2008, 17, 263–268. [Google Scholar] [CrossRef] [PubMed]
- Tezel, G.; Yang, X.; Luo, C.; Peng, Y.; Sun, S.L.; Sun, D. Mechanisms of immune system activation in glaucoma: Oxidative stress-stimulated antigen presentation by the retina and optic nerve head glia. Invest. Ophthalmol. Vis. Sci. 2007, 48, 705–714. [Google Scholar] [CrossRef] [PubMed]
- Tezel, G.; Wax, M.B. Increased production of tumor necrosis factor-alpha by glial cells exposed to simulated ischemia or elevated hydrostatic pressure induces apoptosis in cocultured retinal ganglion cells. J. Neurosci. 2000, 20, 8693–8700. [Google Scholar] [CrossRef] [PubMed]
- Tezel, G. Oxidative stress in glaucomatous neurodegeneration: Mechanisms and consequences. Prog. Retin. Eye Res. 2006, 25, 490–513. [Google Scholar] [CrossRef] [Green Version]
- Tezel, G. The immune response in glaucoma: A perspective on the roles of oxidative stress. Exp. Eye Res. 2011, 93, 178–186. [Google Scholar] [CrossRef] [Green Version]
- Izzotti, A.; Bagnis, A.; Saccà, S.C. The role of oxidative stress in glaucoma. Mutat. Res. 2006, 612, 105–114. [Google Scholar] [CrossRef]
- Garcia-Medina, J.J.; Rubio-Velazquez, E.; Lopez-Bernal, M.D.; Cobo-Martinez, A.; Zanon-Moreno, V.; Pinazo-Duran, M.D.; del-Rio-Vellosillo, M. Glaucoma and antioxidants: Review and update. Antioxidants 2020, 9, 1031. [Google Scholar] [CrossRef]
- Tang, B.; Li, S.; Cao, W.; Sun, X. The association of oxidative stress status with open-angle glaucoma and exfoliation glaucoma: A systematic review and meta-analysis. J. Ophthalmol. 2019, 2019, 1803619. [Google Scholar] [CrossRef] [Green Version]
- Hollyfield, J.G.; Bonilha, V.L.; Rayborn, M.E.; Yang, X.; Shadrach, K.G.; Lu, L.; Ufret, R.L.; Salomon, R.G.; Perez, V.L. Oxidative damage-induced inflammation initiates age- related macular degeneration. Nat. Med. 2008, 14, 194–198. [Google Scholar] [CrossRef]
- Jarrett, S.G.; Boulton, M.E. Consequences of oxidative stress in age-related macular degeneration. Mol. Aspects Med. 2012, 33, 399–417. [Google Scholar] [CrossRef] [Green Version]
- Klettner, A. Oxidative stress induced cellular signaling in RPE cells. Front. Biosci. 2012, 4, 392–411. [Google Scholar] [CrossRef]
- Nowak, J.Z. Oxidative stress, polyunsaturated fatty acids-derived oxidation products and bisretinoids as potential inducers of CNS diseases: Focus on age-related macular degeneration. Pharmacol. Rep. 2013, 65, 288–304. [Google Scholar] [CrossRef]
- Sparrow, J.R.; Zhou, J.; Ben-Shabat, S.; Vollmer, H.; Itagaki, Y.; Nakanishi, K. Involvement of oxidative mechanisms in blue-light-induced damage to A2E-laden RPE. Investig. Ophthalmol. Vis. Sci. 2002, 43, 1222–1227. [Google Scholar]
- Winkler, B.S.; Boulton, M.E.; Gottsch, J.D.; Sternberg, P. Oxidative damage and age-related macular degeneration. Mol. Vis. 1999, 5, 32. [Google Scholar] [PubMed]
- Ham, W.T.; Ruffolo, J.J.; Mueller, H.A.; Clarke, A.M.; Moon, M.E. Histologic analysis of photochemical lesions produced in rhesus retina by short-wave-length light. Investig. Ophthalmol. Vis. Sci. 1978, 17, 1029–1035. [Google Scholar]
- Tong, Y.; Wang, S. Not all stressors are equal: Mechanism of stressors on RPE cell degeneration. Front. Cell Dev. Biol. 2020, 8, 591067. [Google Scholar] [CrossRef]
- Somasundaran, S.; Constable, I.J.; Mellough, C.B.; Carvalho, L.S. Retinal pigment epithelium and age-related macular degeneration: A review of major disease mechanisms. Clin. Exp. Ophthalmol. 2020, 48, 1043–1056. [Google Scholar] [CrossRef]
- Duncan, K.E.; Stillman, M.J. Metal-dependent protein folding: Metallation of metallothionein. J. Inorg. Biochem. 2006, 100, 2101–2107. [Google Scholar] [CrossRef]
- Romero-Isart, N.; Vasák, M. Advances in the structure and chemistry of metallothioneins. J. Inorg. Biochem. 2002, 88, 388–396. [Google Scholar] [CrossRef]
- Oppermann, B.; Zhang, W.; Magabo, K.; Kantorow, M. Identification and spatial analysis of metallothioneins expressed by the adult human lens. Investig. Ophthalmol. Vis. Sci. 2001, 42, 188–193. [Google Scholar]
- Tate, D.J.; Miceli, M.V.; Newsome, D.A. Expression of metallothionein isoforms in human chorioretinal complex. Curr. Eye Res. 2002, 24, 12–25. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, S.; Shimazawa, M.; Hara, H. Physiological Roles of Metallothioneins in Central Nervous System Diseases. Biol. Pharm. Bull. 2018, 41, 1006–1013. [Google Scholar] [CrossRef] [PubMed]
- Thornalley, P.J.; Vasák, M. Possible role for metallothionein in protection against radiation-induced oxidative stress. Kinetics and mechanism of its reaction with superoxide and hydroxyl radicals. Biochim. Biophys. Acta 1985, 827, 36–44. [Google Scholar] [CrossRef]
- Ohashi, Y.; Dogru, M.; Tsubota, K. Laboratory findings in tear fluid analysis. Clin. Chim. Acta 2006, 369, 17–28. [Google Scholar] [CrossRef] [PubMed]
- Karnati, R.; Laurie, D.E.; Laurie, G.W. Lacritin and the Tear Proteome as Natural Replacement Therapy for Dry Eye. Exp. Eye Res. 2013, 117, 39–52. [Google Scholar] [CrossRef] [Green Version]
- Lauweryns, B.; van den Oord, J.J.; Missotten, L. The transitional zone between limbus and peripheral cornea. An immunohistochemical study. Investig. Ophthalmol. Vis. Sci. 1993, 34, 1991–1999. [Google Scholar]
- Schlötzer-Schrehardt, U.; Kruse, F.E. Identification and characterization of limbal stem cells. Exp. Eye Res. 2005, 81, 247–264. [Google Scholar] [CrossRef]
- Lu, H.; Hunt, D.M.; Ganti, R.; Davis, A.; Dutt, K.; Alam, J.; Hunt, R.C. Metallothionein protects retinal pigment epithelial cells against apoptosis and oxidative stress. Exp. Eye Res. 2002, 74, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Carnes, M.U.; Allingham, R.R.; Ashley-Koch, A.; Hauser, M.A. Transcriptome analysis of adult and fetal trabecular meshwork, cornea, and ciliary body tissues by RNA sequencing. Exp. Eye Res. 2018, 167, 91–99. [Google Scholar] [CrossRef]
- Gottsch, J.D.; Bowers, A.L.; Margulies, E.H.; Seitzman, G.D.; Kim, S.W.; Saha, S.; Jun, A.S.; Stark, W.J.; Liu, S.H. Serial analysis of gene expression in the corneal endothelium of Fuchs’ dystrophy. Investig. Ophthalmol. Vis. Sci. 2003, 44, 594–599. [Google Scholar] [CrossRef] [Green Version]
- De Roo, A.-K.; Wouters, J.; Govaere, O.; Foets, B.; van den Oord, J.J. Identification of circulating fibrocytes and dendritic derivatives in corneal endothelium of patients with Fuch’s dystrophy. Investig. Ophthalmol. Vis. Sci. 2017, 58, 670–681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azizi, B.; Ziaei, A.; Fuchsluger, T.; Schmedt, T.; Chen, Y.; Jurkunas, U.V. p53-regulated increase in oxidative-stress–induced apoptosis in Fuchs endothelial corneal dystrophy: A native tissue model. Investig. Ophthalmol. Vis. Sci. 2011, 52, 9291–9297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaluzhny, Y.; Kinuthia, M.W.; Lapointe, A.M.; Truong, T.; Klausner, M.; Hayden, P. Oxidative stress in corneal injuries of different origin: Utilization of 3D human corneal epithelial tissue model. Exp. Eye Res. 2020, 190, 107867. [Google Scholar] [CrossRef] [PubMed]
- Han, E.S.; Muller, F.L.; Perez, V.I.; Qi, W.; Liang, H.; Xi, L.; Fu, C.; Doyle, E.; Hickey, M.; Cornell, J.; et al. The in vivo gene expression signature of oxidative stress. Physiol. Genom. 2008, 34, 112–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hejtmancik, J.F.; Riazuddin, S.A.; McGreal, R.; Liu, W.; Cvekl, A.; Shiels, A. Lens Biology and Biochemistry. Prog. Mol. Biol. Transl. Sci. 2015, 134, 169–201. [Google Scholar] [CrossRef]
- Cejková, J.; Stípek, S.; Crkovská, J.; Ardan, T. Changes of superoxide dismutase, catalase and glutathione peroxidase in the corneal epithelium after UVB rays. Histochemical and biochemical study. Histol. Histopathol. 2000, 15, 1043–1050. [Google Scholar] [CrossRef]
- Fecondo, J.V.; Augusteyn, R.C. Superoxide dismutase, catalase and glutathione peroxidase in the human cataractous lens. Exp. Eye Res. 1983, 36, 15–23. [Google Scholar] [CrossRef]
- Bova, L.M.; Sweeney, M.H.; Jamie, J.F.; Truscott, R.J. Major changes in human ocular UV protection with age. Investig. Ophthalmol. Vis. Sci. 2001, 42, 200–205. [Google Scholar]
- Kantorow, M.; Kays, T.; Horwitz, J.; Huang, Q.; Sun, J.; Piatigorsky, J.; Carper, D. Differential display detects altered gene expression between cataractous and normal human lenses. Investig. Ophthalmol. Vis. Sci. 1998, 39, 2344–2354. [Google Scholar]
- Whitson, J.A.; Zhang, X.; Medvedovic, M.; Chen, J.; Wei, Z.; Monnier, V.M.; Fan, X. Transcriptome of the GSH-depleted lens reveals changes in detoxification and EMT signaling genes, transport systems, and lipid homeostasis. Investig. Ophthalmol. Vis. Sci. 2017, 58, 2666–2684. [Google Scholar] [CrossRef]
- Spector, A. Oxidative stress-induced cataract: Mechanism of action. FASEB J. 1995, 9, 1173–1182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hawse, J.R.; Hejtmancik, J.F.; Huang, Q.; Sheets, N.L.; Hosack, D.A.; Lempicki, R.A.; Horwitz, J.; Kantorow, M. Identification and functional clustering of global gene expression differences between human age-related cataract and clear lenses. Mol. Vis. 2003, 9, 515–537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hawse, J.R.; Padgaonkar, V.A.; Leverenz, V.R.; Pelliccia, S.E.; Kantorow, M.; Giblin, F.J. The role of metallothionein IIa in defending lens epithelial cells against cadmium and TBHP induced oxidative stress. Mol. Vis. 2006, 12, 342–349. [Google Scholar] [PubMed]
- Saito, T.; Tezuka, T.; Konno, R.; Fujii, N. Protective effects of metallothionein I and II against metal- and ultraviolet radiation-induced damage in cultured lens epithelial cells. Jpn. J. Ophthalmol. 2010, 54, 486–493. [Google Scholar] [CrossRef]
- González-Iglesias, H.; Petrash, C.; Rodríguez-Menéndez, S.; García, M.; Álvarez, L.; Fernández-Vega-Cueto, L.; Fernández, B.; Pereiro, R.; Sanz-Medel, A.; Coca-Prados, M. Quantitative distribution of Zn, Fe and Cu in the human lens and study of the Zn–metallothionein redox system in cultured lens epithelial cells by elemental MS. J. Anal. At. Spectrom. 2017, 32, 1746–1756. [Google Scholar] [CrossRef]
- Bazan, N.G. The metabolism of omega-3 polyunsaturated fatty acids in the eye: The possible role of docosahexaenoic acid and docosanoids in retinal physiology and ocular pathology. Prog. Clin. Biol. Res. 1989, 312, 95–112. [Google Scholar]
- Dargel, R. Lipid peroxidation—A common pathogenetic mechanism? Exp. Toxicol. Pathol. 1992, 44, 169–181. [Google Scholar] [CrossRef]
- Glickman, R.D. Phototoxicity to the retina: Mechanisms of damage. Int. J. Toxicol. 2002, 21, 473–490. [Google Scholar] [CrossRef]
- van Reyk, D.M.; Gillies, M.C.; Davies, M.J. The retina: Oxidative stress and diabetes. Redox Rep. 2003, 8, 187–192. [Google Scholar] [CrossRef]
- Tate, D.J.; Newsome, D.A.; Oliver, P.D. Metallothionein shows an age-related decrease in human macular retinal pigment epithelium. Investig. Ophthalmol. Vis. Sci. 1993, 34, 2348–2351. [Google Scholar]
- Heesterbeek, T.J.; Lorés-Motta, L.; Hoyng, C.B.; Lechanteur, Y.T.E.; den Hollander, A.I. Risk factors for progression of age-related macular degeneration. Ophthalmic. Physiol. Opt. 2020, 40, 140–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liles, M.R.; Newsome, D.A.; Oliver, P.D. Antioxidant enzymes in the aging human retinal pigment epithelium. Arch. Ophthalmol. 1991, 109, 1285–1288. [Google Scholar] [CrossRef] [PubMed]
- Newsome, D.A.; Miceli, M.V.; Tate, D.J.; Alcock, N.W.; Oliver, P.D. Zinc content of human retinal pigment epithelium decreases with age and macular degeneration, but superoxide dismutase activity increases. J. Trace Elem. Exp. Med. 1995, 8, 193–199. [Google Scholar] [CrossRef]
- Decanini, A.; Nordgaard, C.L.; Feng, X.; Ferrington, D.A.; Olsen, T.W. Changes in select redox proteins of the retinal pigment epithelium in age-related macular degeneration. Am. J. Ophthalmol. 2008, 143, 607–615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yi, W.; Lu, Y.; Zhong, S.; Zhang, M.; Sun, L.; Dong, H.; Wang, M.; Wei, M.; Xie, H.; Qu, H.; et al. A single-cell transcriptome atlas of the aging human and macaque retina. Natl. Sci. Rev. 2020. [Google Scholar] [CrossRef]
- Voigt, A.P.; Mullin, N.K.; Stone, E.M.; Tucker, B.A.; Scheetz, T.E.; Mullins, R.F. Single-cell RNA sequencing in vision research: Insights into human retinal health and disease. Prog. Retin. Eye Res. 2020, 100934. [Google Scholar] [CrossRef]
- Nicolas, M.G.; Fujiki, K.; Murayama, K.; Suzuki, M.T.; Shindo, N.; Hotta, Y.; Iwata, F.; Fujimura, T.; Yoshikawa, Y.; Cho, F.; et al. Studies on the mechanism of early onset macular degeneration in cynomolgus monkeys. II. Suppression of metallothionein synthesis in the retina in oxidative stress. Exp. Eye Res. 1996, 62, 399–408. [Google Scholar] [CrossRef]
- Miceli, M.V.; Tate, D.J.; Alcock, N.W.; Newsome, D.A. Zinc deficiency and oxidative stress in the retina of pigmented rats. Investig. Ophthalmol. Vis. Sci. 1999, 40, 1238–1244. [Google Scholar]
- Noell, W.K.; Organisciak, D.T.; Ando, H.; Braniecki, M.A.; Durlin, C. Ascorbate and dietary protective mechanisms in retinal light damage of rats: Electrophysiological, histological and DNA measurements. Prog. Clin. Biol. Res. 1987, 247, 469–483. [Google Scholar]
- Truscott, R.J. Age-related nuclear cataract-oxidation is the key. Exp. Eye Res. 2005, 80, 709–725. [Google Scholar] [CrossRef] [PubMed]
- Siu, T.L.; Morley, J.W.; Coroneo, M.T. Toxicology of the retina: Advances in understanding the defence mechanisms and pathogenesis of drug- and light-induced retinopathy. Clin. Exp. Ophthalmol. 2008, 36, 176–185. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Tower, J. Effects of light on aging and longevity. Ageing Res. Rev. 2019, 53, 100913. [Google Scholar] [CrossRef] [PubMed]
- Glickman, R.D. The origin of photo-oxidative stress in the aging eye. Prog. Brain Res. 2001, 131, 699–712. [Google Scholar] [CrossRef] [PubMed]
- Baksheeva, V.E.; Tiulina, V.V.; Tikhomirova, N.K.; Gancharova, O.S.; Komarov, S.V.; Philippov, P.P.; Zamyatnin, A.A., Jr.; Senin, I.I.; Zernii, E.Y. Supression of light-induced oxidative stress in the retina by mitochondria-targeted antioxidant. Antioxidants 2019, 8, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Wu, W.; Dentchev, T.; Zeng, Y.; Wang, J.; Tsui, I.; Tobias, J.W.; Bennett, J.; Baldwin, D.; Dunaief, J.L. Light damage induced changes in mouse retinal gene expression. Exp. Eye Res. 2004, 79, 239–247. [Google Scholar] [CrossRef]
- Chen, L.; Wu, W.; Dentchev, T.; Wong, R.; Dunaief, J.L. Increased metallothionein in light damaged mouse retinas. Exp. Eye Res. 2004, 79, 287–293. [Google Scholar] [CrossRef]
- Natoli, R.; Zhu, Y.; Valter, K.; Bisti, S.; Eells, J.; Stone, J. Gene and noncoding RNA regulation underlying photoreceptor protection: Microarray study of dietary antioxidant saffron and photobiomodulation in rat retina. Mol. Vis. 2010, 16, 1801–1822. [Google Scholar]
- Tsuruma, K.; Shimazaki, H.; Ohno, Y.; Inoue, Y.; Honda, A.; Imai, S.; Lee, J.; Shimazawa, M.; Satoh, M.; Hara, H. Metallothionein-III deficiency exacerbates light-induced retinal degeneration. Investig. Ophthalmol. Vis. Sci. 2012, 53, 7896–7903. [Google Scholar] [CrossRef]
- Michalska, A.E.; Choo, K.H. Targeting and germ-line transmission of a null mutation at the metallothionein I and II loci in mouse. Proc. Natl. Acad. Sci. USA 1993, 90, 8088–8092. [Google Scholar] [CrossRef] [Green Version]
- Sato, M.; Bremner, I. Oxygen free radicals and metallothionein. Free Radic. Biol. Med. 1993, 14, 325–337. [Google Scholar] [CrossRef]
- Masters, B.A.; Kelly, E.J.; Quaife, C.J.; Brinster, R.L.; Palmiter, R.D. Targeted disruption of metallothionein I and II genes increases sensitivity to cadmium. Proc. Natl. Acad. Sci. USA 1994, 91, 584–588. [Google Scholar] [CrossRef] [Green Version]
- Koh, J.Y.; Lee, S.J. Metallothionein-3 as a multifunctional player in the control of cellular processes and diseases. Mol. Brain 2020, 13, 116. [Google Scholar] [CrossRef]
- Suemori, S.; Shimazawa, M.; Kawase, K.; Satoh, M.; Nagase, H.; Yamamoto, T.; Hara, H. Metallothionein, an endogenous antioxidant, protects against retinal neuron damage in mice. Investig. Ophthalmol. Vis. Sci. 2006, 47, 3975–3982. [Google Scholar] [CrossRef] [Green Version]
- Choi, D.W. Excitotoxic cell death. J. Neurobiol. 1992, 23, 1261–1276. [Google Scholar] [CrossRef]
- Salt, T.E. Handbook of Neurotoxicity; Kostrzewa, R.M., Ed.; Springer: New York, NY, USA, 2014; pp. 1273–1285. [Google Scholar]
- Wallin, C.; Weber, S.G.; Sandberg, M. Glutathione efflux induced by NMDA and kainate: Implications in neurotoxicity? J. Neurochem. 1999, 73, 1566–1572. [Google Scholar] [CrossRef] [PubMed]
- Wallin, C.; Abbas, A.-K.; Tranberg, M.; Weber, S.G.; Wigström, H.; Sandberg, M. Searching for mechanisms of N-Methyl-D-Aspartate-induced glutathione efflux in organotypic hippocampal cultures. Neurochem. Res. 2003, 28, 281–291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karasawa, M.; Hosoi, J.; Hashiba, H.; Nose, K.; Tohyama, C.; Abe, E.; Suda, T.; Kuroki, T. Regulation of metallothionein gene expression by 1 alpha,25-dihydroxyvitamin D3 in cultured cells and in mice. Proc. Natl. Acad. Sci. USA 1987, 84, 8810–8813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tate, D.J.; Miceli, M.V.; Newsome, D.A. Zinc protects against oxidative damage in cultured human retinal pigment epithelial cells. Free Radic. Biol. Med. 1999, 26, 704–713. [Google Scholar] [CrossRef]
- Newsome, D.A.; Miceli, M.V.; Liles, M.R.; Tate, D.J.; Oliver, P.D. Antioxidants in the retinal pigment epithelium. Prog. Retinal Eye Res. 1994, 13, 101–123. [Google Scholar] [CrossRef]
- Chen, R.W.; Vasey, E.J.; Whanger, P.D. Accumulation and depletion of zinc in rat liver and kidney metallothionens. J. Nutr. 1977, 107, 805–813. [Google Scholar] [CrossRef] [PubMed]
- Oliver, P.D.; Tate, D.J.; Newsome, D.A. Metallothionein in human retinal pigment epithelial cells: Expression, induction and zinc uptake. Curr. Eye Res. 1992, 11, 183–188. [Google Scholar] [CrossRef] [PubMed]
- Tate, D.J.; Newsome, D.A. A novel zinc compound (zinc monocysteine) enhances the antioxidant capacity of human retinal pigment epithelial cells. Curr Eye Res. 2006, 31, 675–683. [Google Scholar] [CrossRef] [PubMed]
- Pabis, K.; Gundacker, C.; Giacconi, R.; Basso, A.; Costarelli, L.; Piacenza, F.; Strizzi, S.; Provinciali, M.; Malavolta, M. Zinc supplementation can reduce accumulation of cadmium in aged metallothionein transgenic mice. Chemosphere 2018, 211, 855–860. [Google Scholar] [CrossRef] [PubMed]
- Scimone, C.; Alibrandi, S.; Scalinci, S.Z.; Trovato Battagliola, E.; D’Angelo, R.; Sidoti, A.; Donato, L. Expression of Pro-Angiogenic Markers Is Enhanced by Blue Light in Human RPE Cells. Antioxidants 2020, 9, 1154. [Google Scholar] [CrossRef] [PubMed]
- Donato, L.; D’Angelo, R.; Alibrandi, S.; Rinaldi, C.; Sidoti, A.; Scimone, C. Effects of A2E-Induced Oxidative Stress on Retinal Epithelial Cells: New Insights on Differential Gene Response and Retinal Dystrophies. Antioxidants 2020, 9, 307. [Google Scholar] [CrossRef] [Green Version]
- Bailey, T.A.; Kanuga, N.; Romero, I.A.; Greenwood, J.; Luthert, P.J.; Cheetham, M.E. Oxidative stress affects the junctional integrity of retinal pigment epithelial cells. Investig. Ophthalmol. Vis. Sci. 2004, 45, 675–684. [Google Scholar] [CrossRef] [Green Version]
- Karlsson, M.; Kurz, T.; Brunk, U.T.; Nilsson, S.E.; Frennesson, C.I. What does the commonly used DCF test for oxidative stress really show? Biochem. J. 2010, 428, 183–190. [Google Scholar] [CrossRef] [Green Version]
- Zareba, M.; Raciti, M.W.; Henry, M.M.; Sarna, T.; Burke, J.M. Oxidative stress in ARPE-19 cultures: Do melanosomes confer cytoprotection? Free Radic. Biol. Med. 2006, 40, 87–100. [Google Scholar] [CrossRef]
- Kurz, T.; Karlsson, M.; Brunk, U.T.; Nilsson, S.E.; Frennesson, C. ARPE-19 retinal pigment epithelial cells are highly resistant to oxidative stress and exercise strict control over their lysosomal redox-active iron. Autophagy 2009, 5, 494–501. [Google Scholar] [CrossRef]
- Karlsson, M.; Frennesson, C.; Gustafsson, T.; Brunk, U.T.; Nilsson, S.E.; Kurz, T. Autophagy of iron-binding proteins may contribute to the oxidative stress resistance of ARPE-19 cells. Exp. Eye Res. 2013, 116, 359–365. [Google Scholar] [CrossRef]
- Karlsson, M.; Kurz, T. Attenuation of iron-binding proteins in ARPE-19 cells reduces their resistance to oxidative stress. Acta Ophthalmol. 2016, 94, 556–564. [Google Scholar] [CrossRef] [PubMed]
- Creagh, E.M.; Carmody, R.J.; Cotter, T.G. Heat shock protein 70 inhibits caspase- dependent and -independent apoptosis in Jurkat T cells. Exp. Cell. Res. 2000, 257, 58–66. [Google Scholar] [CrossRef] [PubMed]
- Martindale, J.L.; Holbrook, N.J. Cellular response to oxidative stress: Signaling for suicide and survival. J. Cell. Physiol. 2002, 192, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Baird, S.K.; Kurz, T.; Brunk, U.T. Metallothionein protects against oxidative stress-induced lysosomal destabilization. Biochem J. 2006, 394, 275–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doulias, P.T.; Kotoglou, P.; Tenopoulou, M.; Keramisanou, D.; Tzavaras, T.; Brunk, U.; Galaris, D.; Angelidis, C. Involvement of heat shock protein-70 in the mechanism of hydrogen peroxide-induced DNA damage: The role of lysosomes and iron. Free Radic. Biol. Med. 2007, 42, 567–577. [Google Scholar] [CrossRef]
- Kurz, T.; Gustafsson, B.; Brunk, U.T. Cell sensitivity to oxidative stress is influenced by ferritin autophagy. Free Radic. Biol. Med. 2011, 50, 1647–1658. [Google Scholar] [CrossRef] [Green Version]
- Choudhary, S.; Xiao, T.; Srivastava, S.; Zhang, W.; Chan, L.L.; Vergara, L.A.; Van Kuijk, F.J.; Ansari, N.H. Toxicity and detoxification of lipid-derived aldehydes in cultured retinal pigmented epithelial cells. Toxicol. Appl. Pharmacol. 2005, 204, 122–134. [Google Scholar] [CrossRef] [PubMed]
- Huang, I.Y.; Yoshida, A.; Tsunoo, H.; Nakajima, H. Mouse liver metallothioneins. Complete amino acid sequence of metallothionein-I. J. Biol. Chem. 1977, 252, 8217–8221. [Google Scholar] [CrossRef]
- Yoshida, A.; Kaplan, B.E.; Kimura, M. Metal-binding and detoxification effect of synthetic oligopeptides containing three cysteinyl residues. Proc. Natl. Acad. Sci. USA 1979, 76, 486–490. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Wang, L.; Gu, S.; Yu, Y.; Huang, H.; Mo, K.; Xu, H.; Zeng, F.; Xiao, Y.; Peng, L.; et al. D609 protects retinal pigmented epithelium as a potential therapy for age-related macular degeneration. Signal Transduct. Target Ther. 2020, 5, 20. [Google Scholar] [CrossRef] [Green Version]
- Amorati, R.; Valgimigli, L. Advantages and limitations of common testing methods for antioxidants. Free Radic. Res. 2015, 49, 633–649. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, C.M.; Biswal, M.R.; Li, H.; Han, P.; Ildefonso, C.J.; Lewin, A.S. Repurposing an orally available drug for the treatment of geographic atrophy. Mol. Vis. 2016, 22, 294–310. [Google Scholar] [PubMed]
- Castello, P.R.; Drechsel, D.A.; Patel, M. Mitochondria are a major source of paraquat-induced reactive oxygen species production in the brain. J. Biol. Chem. 2007, 282, 14186–14193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biswal, M.R.; Ahmed, C.M.; Ildefonso, C.J.; Han, P.; Li, H.; Jivanji, H.; Mao, H.; Lewin, A.S. Systemic treatment with a 5HT1a agonist induces anti-oxidant protection and preserves the retina from mitochondrial oxidative stress. Exp. Eye Res. 2015, 140, 94–105. [Google Scholar] [CrossRef] [Green Version]
- Ramos, A.J.; Rubio, M.D.; Defagot, C.; Hischberg, L.; Villar, M.J.; Brusco, A. The 5HT1A receptor agonist, 8-OH-DPAT, protects neurons and reduces astroglial reaction after ischemic damage caused by cortical devascularization. Brain Res. 2004, 1030, 201–220. [Google Scholar] [CrossRef] [PubMed]
- Miyazaki, I.; Asanuma, M.; Murakami, S.; Takeshima, M.; Torigoe, N.; Kitamura, Y.; Miyoshi, K. Targeting 5-HT(1A) receptors in astrocytes to protect dopaminergic neurons in Parkinsonian models. Neurobiol. Dis. 2013, 59, 244–256. [Google Scholar] [CrossRef]
- Rodríguez-Menéndez, S.; Fernández, B.; García, M.; Álvarez, L.; Fernández, M.L.; Sanz-Medel, A.; Coca-Prados, M.; Pereiro, R.; González-Iglesias, H. Quantitative study of zinc and metallothioneins in the human retina and RPE cells by mass spectrometry-based methodologies. Talanta 2018, 178, 222–230. [Google Scholar] [CrossRef]
- Rodríguez-Menéndez, S.; García, M.; Fernández, B.; Álvarez, L.; Fernández-Vega-Cueto, A.; Coca-Prados, M.; Pereiro, R.; González-Iglesias, H. The Zinc-Metallothionein Redox System Reduces Oxidative Stress in Retinal Pigment Epithelial Cells. Nutrients 2018, 10, 1874. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Schematic view of the human eye anatomy and representation of the absorption of UV radiation by the main ocular barriers.
Figure 1.
Schematic view of the human eye anatomy and representation of the absorption of UV radiation by the main ocular barriers.

Figure 2.
Hierarchical cluster and heat map (whole-genome expression microarrays) of the main antioxidant enzymes in human ocular tissues. The row at the top shows the clustering information in the form of a dendogram and the similarity relationships among the genes and tissues: cornea (n = 11), trabecular meshwork (TM; n = 9), CB (n = 12), sclera (n = 7), iris (11), RPE (n = 8), retina (n = 12) and lens (n = 10). The column at the left of the heat map shows four clusters (A to D), each with antioxidant enzymes expressed at different abundance. Mean values of 6.6–14.9 from biological replicas per tissue are indicated according to the log2 scale, in arbitrary units, depicted at the bottom. Hierarchical cluster and heat maps were created using Illumina BeadChip array platform (HumanHT-12 v4.0 Expression BeadChip Kit, Illumina, San Diego, CA, USA) and ArrayStar software, version 4 (DNASTAR, Inc, Madison, WI, USA) according to Alvarez et al., 2012 [15]. n: number of samples per tissue.
Figure 2.
Hierarchical cluster and heat map (whole-genome expression microarrays) of the main antioxidant enzymes in human ocular tissues. The row at the top shows the clustering information in the form of a dendogram and the similarity relationships among the genes and tissues: cornea (n = 11), trabecular meshwork (TM; n = 9), CB (n = 12), sclera (n = 7), iris (11), RPE (n = 8), retina (n = 12) and lens (n = 10). The column at the left of the heat map shows four clusters (A to D), each with antioxidant enzymes expressed at different abundance. Mean values of 6.6–14.9 from biological replicas per tissue are indicated according to the log2 scale, in arbitrary units, depicted at the bottom. Hierarchical cluster and heat maps were created using Illumina BeadChip array platform (HumanHT-12 v4.0 Expression BeadChip Kit, Illumina, San Diego, CA, USA) and ArrayStar software, version 4 (DNASTAR, Inc, Madison, WI, USA) according to Alvarez et al., 2012 [15]. n: number of samples per tissue.

Figure 3.
(a) Mammalian MTs contain 60–68 amino acids, 20 of which are cysteines that bind seven divalent metal ions in two metal/thiolate clusters, the α- and β-metal-binding domains. Eleven cysteines bind four zinc ions in the C-terminal α-cluster, whereas nine cysteines bind three zinc ions in the N-terminal β-cluster. (b) Zinc–metallothionein redox cycle: zinc bound to MT (Zn–MT) is released under physiological oxidative conditions, forming MT-disulfide (thionin), process boosted by free radicals derived from nitric oxide, ROS and oxidized glutathione. MT-disulfide may be degraded or reduced to MT-thiol (thionein) in the presence of a selenium-derived catalyst. MT-thiol binds zinc to form the thermodynamically stable Zn–MT system. Adapted from Gonzalez-Iglesias et al., 2014 [138].
Figure 3.
(a) Mammalian MTs contain 60–68 amino acids, 20 of which are cysteines that bind seven divalent metal ions in two metal/thiolate clusters, the α- and β-metal-binding domains. Eleven cysteines bind four zinc ions in the C-terminal α-cluster, whereas nine cysteines bind three zinc ions in the N-terminal β-cluster. (b) Zinc–metallothionein redox cycle: zinc bound to MT (Zn–MT) is released under physiological oxidative conditions, forming MT-disulfide (thionin), process boosted by free radicals derived from nitric oxide, ROS and oxidized glutathione. MT-disulfide may be degraded or reduced to MT-thiol (thionein) in the presence of a selenium-derived catalyst. MT-thiol binds zinc to form the thermodynamically stable Zn–MT system. Adapted from Gonzalez-Iglesias et al., 2014 [138].

Figure 4.
Gene expression profiling of MT1 (MT1A, MT1B, MT1E, MT1F, MT1G, MT1H, MT1M AND MT1X), MT2 (MT2A) and MT3 isoforms in human eye tissues, according to Alvarez et al., 2012 [15] and adapted, expressed in arbitrary units (AU) after normalization with internal controls. Scale bars of Y-axis range from 0 to 35,000 AU for each eye tissue, with the exception of lens, which Y-axis scale bar ranges from 0 to 90,000 AU.
Figure 4.
Gene expression profiling of MT1 (MT1A, MT1B, MT1E, MT1F, MT1G, MT1H, MT1M AND MT1X), MT2 (MT2A) and MT3 isoforms in human eye tissues, according to Alvarez et al., 2012 [15] and adapted, expressed in arbitrary units (AU) after normalization with internal controls. Scale bars of Y-axis range from 0 to 35,000 AU for each eye tissue, with the exception of lens, which Y-axis scale bar ranges from 0 to 90,000 AU.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Álvarez-Barrios, A.; Álvarez, L.; García, M.; Artime, E.; Pereiro, R.; González-Iglesias, H. Antioxidant Defenses in the Human Eye: A Focus on Metallothioneins. Antioxidants 2021, 10, 89. https://doi.org/10.3390/antiox10010089
AMA Style
Álvarez-Barrios A, Álvarez L, García M, Artime E, Pereiro R, González-Iglesias H. Antioxidant Defenses in the Human Eye: A Focus on Metallothioneins. Antioxidants. 2021; 10(1):89. https://doi.org/10.3390/antiox10010089
Chicago/Turabian StyleÁlvarez-Barrios, Ana, Lydia Álvarez, Montserrat García, Enol Artime, Rosario Pereiro, and Héctor González-Iglesias. 2021. "Antioxidant Defenses in the Human Eye: A Focus on Metallothioneins" Antioxidants 10, no. 1: 89. https://doi.org/10.3390/antiox10010089
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.