


Abstract
Background/Aim: Glucose transporter 1 (GLUT1) has been demonstrated to be overexpressed in various cancer tissues and play a significant role on growth, metastasis, and apoptosis in cancer cells. This study aimed to reveal the clinical relevance of glucose transporter 1 (GLUT1) in carcinogenesis and progression on liver fluke-associated cholangiocarcinoma (CCA). Materials and Methods: Expression of GLUT1 in CCA tissues from patients, as well as from a liver fluke-induced CCA hamster model, was determined using immunohistochemistry. CCA cell lines were transfected with GLUT1 siRNA and the roles of GLUT1 on cell growth as well as migration and invasion were investigated by using a clonogenic assay and Boyden chamber assays, respectively. Results: GLUT1 was aberrantly expressed in hyperplastic/dysplastic bile ducts and CCA, but not in the normal bile ducts. High GLUT1 expression was significantly associated with non-papillary type, large tumor size, and short survival of patients. GLUT1 was expressed during cholangio-carcinogenesis and gradually increased with progression of histopathologic bile ducts. Silencing of GLUT1 expression significantly suppressed growth, migration, and invasion of CCA cell lines. Conclusion: GLUT1 plays important roles in carcinogenesis and progression of liver fluke-associated CCA. Targeting GLUT1 may be a strategy for treatment of metastasis in liver fluke-associated CCA.
The well-known aerobic glycolysis or Warburg effect (1) is a phenomenon when cancer cells reprogram their glucose metabolism via increasing glucose uptake and glycolytic activity even in the presence of oxygen. This event is one of the hallmarks for cancer to maintain energy and contribute the intermediates for de novo synthesis of lipids, nucleotides, and amino acid (2).
Cells uptake glucose via glucose transporters (GLUTs) that are a key rate-limiting factor in transporting glucose into cells and accelerate glycolysis. At present, there are 14 GLUTs (GLUT1-14 or SLC2A1-14) reported in humans (3). Among these, GLUT1 is reported to be over-expressed in various tumor tissues (4) and related to aggressive phenotypes, e.g., high proliferation rate, metastasis as well as resistances to chemo- and radio-therapy in a range of tumor entities (5, 6). Therefore, targeting GLUTs and pathways related to glucose metabolism has become a novel strategy for cancer therapy.
Cholangiocarcinoma (CCA) is the second most common liver cancer; the incidence and mortality rates of which are increasing globally (7). The incidence of CCA is particularly high in the Northeast of Thailand and countries around the Great Mae-Kong subregions (8). A known risk factor of CCA in this region is the liver fluke (Opisthorchis viverrini, Ov) infection that creates a chronic inflammation in the biliary system (8). Lack of early diagnosis and effective treatments for CCA patients, as well as the highly metastatic nature of CCA, result in a poor patient outcome. In addition, CCA has a high recurrence rate even in the operative cases with a low 5-year survival (9). A new strategic approach for a better treatment is therefore, urgently required.
Our previous studies demonstrated that up-regulation of hexokinase 2 (10) and lactate dehydrogenase A (11) were related to poor prognosis of CCA patients. These observations indicated that CCA might have a high glycolytic rate and high glucose uptake implying the high GLUT1 expression in CCA. Significant association of GLUT1 expression with aggressive potential in non-fluke related CCA has been reported by Kubo et al. in 2014 (12). As expression profiles, mutations and epigenetic landscapes (13) of fluke-related intrahepatic CCA were different from those of the non-fluke-related intrahepatic CCAs, the involvement of GLUT1 in tumor development and progression of fluke-related CCA remains to be elucidated.
The present study aimed to reveal the involvement of GLUT1 expression in carcinogenesis and progression of liver fluke-associated CCA. Therefore, GLUT1 expression was investigated in CCA tissues from a liver fluke-induced CCA hamster model. Moreover, the effects of GLUT1 knockdown on cell growth, migration, and invasion were investigated in CCA cell lines.
Materials and Methods
Patients. The study protocol was approved by The Khon Kaen University Ethics Committee for Human Research (HE591063). Paraffin-embedded liver tissues from histologically proven CCA patients (n=82) were obtained from the biobank of Cholangiocarcinoma Research Institute, Faculty of Medicine, Khon Kaen University, Thailand. All tissue samples were from Thai CCA patients who lived in the endemic area of Ov infection and had active or history of Ov infection. Informed consent was obtained from each patient.
Tissues from Ov-induced CCA hamsters. The liver sections from Ov-induced CCA hamsters were obtained as previously described (14). Briefly, hamsters were grouped into: non-treated control, Ov infected, N-nitroso dimethylamine (NDMA) treated, and Ov + NDMA treated groups (5 animals each). Liver tissues were collected at 1, 3, and 6 months after treatment. The protocols for animal handling and treatment were approved by the Animal Ethics Committee, Khon Kaen University, Thailand (AEMDKKU1/2558).
Cell lines and cell culture. The CCA cell lines, KKU-100 and KKU-213A, were established from CCA tissues of Thai patients with Ov eggs were evident in the tumor tissues (15, 16). These two cell lines were obtained from the Japanese Collection of Research Bioresources Cell Bank, Osaka, Japan. Cells were cultured in HAM-F12 media (Gibco/BRL, Grand Island, NY, USA) supplemented with 10% fetal bovine serum, 100 U/ml penicillin and 100 mg/ml streptomycin (Gibco/BRL), at 37°C with 5% CO2.
GLUT1 expression data from the Gene Expression Profiling Interactive Analysis (GEPIA) dataset. GEPIA (http://gepia2.cancer-pku.cn/) is a web server for analyzing the RNA sequencing expression data of 9,736 tumors and 8,587 normal samples from The Cancer Genome Atlas (TCGA) and the Genotype-Tissue Expression (GTEx) projects, using a standard processing pipeline (http://gepia2.cancer-pku.cn/). The differential expression of GLUT1 in CCA and the normal bile ducts (NBD) were compared using one-way ANOVA (17).
Immunohistochemical analysis. Protein expression of GLUT1 was examined in paraffin embedded tissues from CCA patients and Ov-induced CCA hamsters by immunohistochemistry (IHC), following the standard protocol (18). The sections were incubated with 1:200 anti-GLUT1 antibody (ab40084, Abcam, Cambridge, UK) at room temperature, overnight and then with peroxidase-conjugated Envision™ secondary antibody (Dako, Carpinteria, CA, USA) for 1 h at room temperature. Positive signal was developed and observed under a light microscope (Axioscope A1, Carl Zeiss, Jena, Germany). The intensity of the signals was scored as 1=weak, 2=moderate, 3=strong and the number of positive cells were graded as 0%=negative, 1-25%=1, 26-50%=2 and 51-75%=3. The expression level of GLUT1 was semiquantitated according to the IHC scores as intensity × frequency (10) and designated as low and high according to the median.
SDS-PAGE and western blot. NP-40 lysis buffer was used for cell lysis and the Lowry method (19) was performed to determine the concentration of proteins. Proteins were separated and transferred to the Hybond™-P PVDF membrane (GE Healthcare, Buckinghamshire, UK) by wet electro-transferring using the Bolt & Mahoney buffer (20). The membrane was incubated with blocking solution (5% skim milk in PBS with 0.3% Tween-20) and then incubated with the primary antibody, 1:2,000 anti-GLUT1 (ab14683; Abcam) or 1:10,000 anti-actin (A5441; Sigma, St. Louis, MO, USA). Signals were developed with the ECL™ Prime Western Blotting Detection System (GE healthcare). ECL signals were detected with Image Quant LAS 4000 mini-image analyzer and quantitated using Image Quant™ TL analysis software (GE healthcare).
RNA extraction and real-time reverse transcriptase polymerase chain reaction. Total RNA was extracted from CCA cell lines using the trizol reagent (Invitrogen, Carlsbad, CA, USA) and reversed to cDNA using the high capacity cDNA Reverse Transcription Kit (Thermo Fisher Scientific, Vilnius, Lithuania). The PCR was performed in the Light Cycle 480® real-time PCR system (Roche Diagnostics, Mannheim, Germany) using Light Cycle 480® SYBR green I master mix (Roche Diagnostics) and GLUT1 primers: forward: 5’-CTTTGTGGCCTTCTTTGAAGT-3’, reverse: 5’-ACACAGTTGCTCCACAT-3’.
RNA silencing. GLUT1 expression in CCA cell lines was suppressed by transient transfection of GLUT1-siRNA using Lipofectamine® 2000 (Invitrogen). The siRNA sequences were as previously described (21). GLUT1-siRNA (50 pmole) was transfected into 8×104 CCA cells in a serum free-HAM-F12 media. Complete media was added after 6 h. Scrambled siRNA (SC) (Qiagen, Hilden, CA) served as a negative control.
Clonogenic assay. CCA cells were treated with siGLUT1 for 48 h and then seeded into a 24-well plate (80 cells/well). The plates were incubated at 37°C in a 5% CO2 incubator for 7 days with media being replaced on day 4. A cluster of more than 50 cells was counted as 1 colony under the microscope.
Cell migration and invasion assay. Cell migration and invasion assays were performed using 8-μm pore transwell plates (Corning, Lowell, MA, USA). The transwells were pre-coated with 40 μg Matrigel/insert (Corning) for the invasion assay. CCA cells were treated with siGLUT1 for 48 h and viable cells of 5×104 from KKU-100 and 4×104 from KKU-213A cells were seeded into the upper chamber. Cells were allowed to move or invade, 16 h for KKU-100 and 6 h for KKU-213A. The migrated or invaded cells were fixed and stained with 1:20,000 Hoechst 33342 for 15 min at room temperature (Invitrogen) and counted under a microscope.
Statistical analysis. The experiments were performed in duplicates or triplicates and repeated three independent times. The data are presented as mean±standard deviation (SD). The statistical differences between groups were compared using Pearson chi-squre, one-way ANOVA and the Student t-test. Survival analysis was performed using Kaplan-Meier method and log-rank test. All p-values<0.05 were considered significant. All analyses were performed using SPSS 17.0 software (SPSS, Chicago, IL, USA).
Results
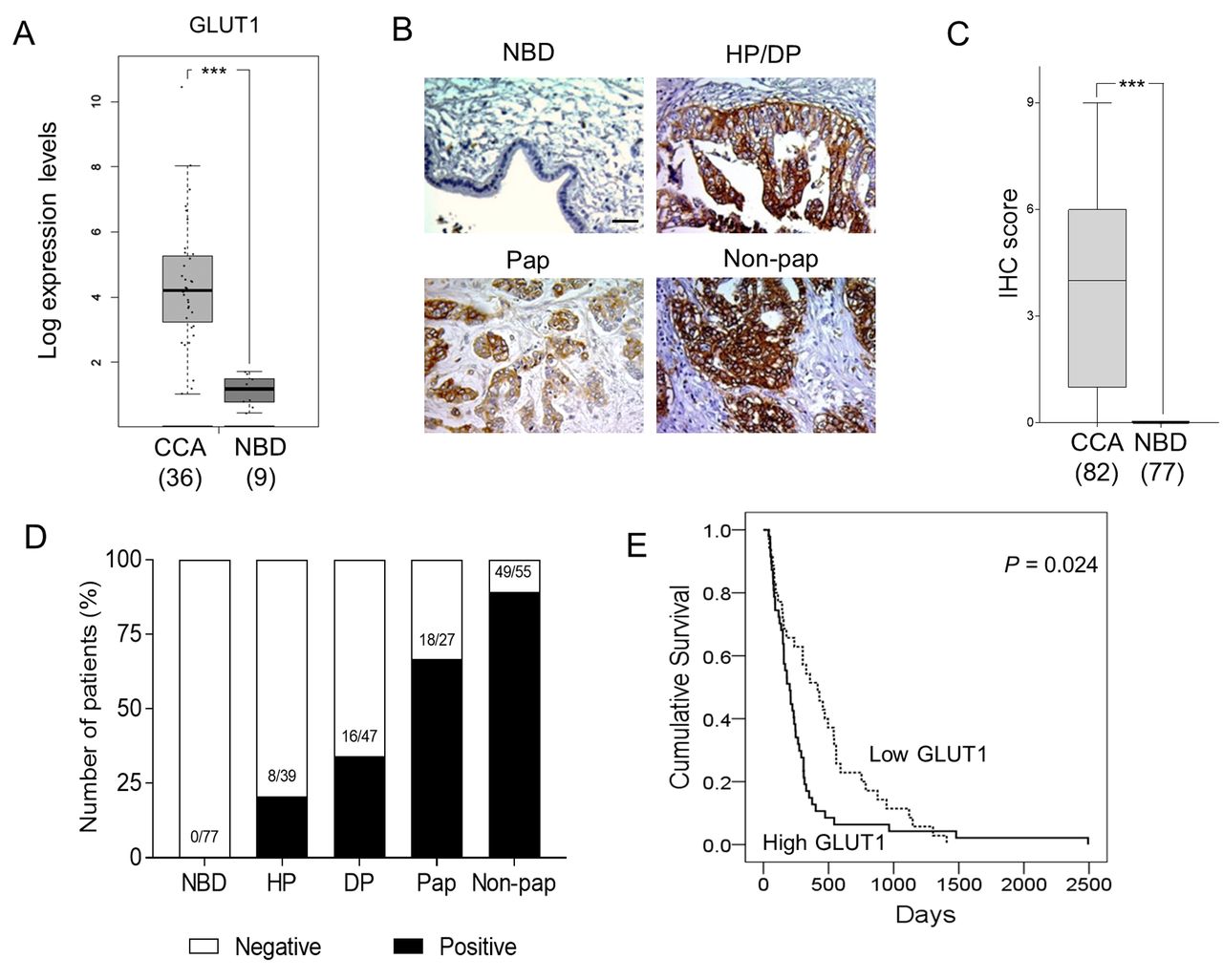
GLUT1 was highly expressed in human CCA tissue and not in normal adjacent tissue. The levels of GLUT1 mRNA in patient CCA tissues were investigated using the data source from GEPIA. The expression levels of GLUT1 mRNA were significantly higher in CCA tissues (n=36) than the adjacent normal tissues (n=9) (p<0.005; Figure 1A). Protein expression of GLUT1 was also examined in the CCA tissue sections from 82 Thai patients. A positive signal of GLUT1 was observed in both plasma membranes and cytoplasm of hyperplastic/dysplastic (HP/DP) bile ducts and CCA, but not in the NBD in adjacent non-tumor tissues (n=77; Figure 1B). The mean IHC scores of GLUT1 observed in CCA tissues were significantly higher than those of NBD (p<0.005; Figure 1C). Differential expression of GLUT1 was detected in pathologic bile ducts. The positive signals of GLUT1 were gradually increased with an increasing degree of bile duct pathology; hyperplasia (21%), dysplasia (34%), papillary type (67%) and non-papillary type CCA (89%) (Figure 1D). These results implied the involvement of GLUT1 in the development and progression of CCA in humans.

GLUT1 is overexpressed in human cholangiocarcinoma (CCA) tissues and correlates with short survival of CCA patients. Comparison of GLUT1 mRNA expressed in CCA tissues from patients and normal bile ducts (NBD) and in the adjacent tissues. Data were analyzed using TCGA data (http://gepia2.cancer-pku.cn/) (A). GLUT1 immunohistochemical (IHC) staining in NBD, hyperplasia (HP), dysplasia (DP), papillary type (Pap) and non-papillary type (Non-pap) CCAs (magnification 200×). scale bar=100 μm (B). The IHC score of GLUT1 observed in CCA tissues vs. NBD (C). Percent of GLUT1 expression in each histopatological bile duct. Numbers indicate numbers of GLUT1 positive/total number of cases (D). Patients with high GLUT1 expression had shorter survivals than those with low GLUT1 expression (E). Data in (A) and (C) are presented as mean±SD, numbers in the parentheses represent numbers of samples, ***p<0.001.
High GLUT1 expression is correlated with poor prognosis of CCA patients. To examine the clinical relevance of GLUT1 in CCA, the association of GLUT1 expression with the clinicopathological features of CCA patients was examined. CCA patients were divided according to the median of GLUT1-IHC scores into low (IHC score <4) and high (IHC score ≥4). Univariate analysis indicated that high GLUT1 expression was correlated with non-papillary type CCA and tumor size of larger than 5 cm (p=0.03 and p=0.009, respectively; Table I). Overall survival analysis using the Kaplan-Meier method showed that CCA patients with high GLUT1 expression had significantly shorter survival than those with low GLUT1 expression (p=0.024; Figure 1E). The univariate analysis using the Cox regression model demonstrated that the non-papillary type CCA (p=0.002) and high expression of GLUT1 (p=0.025) were significantly correlated with poor survival for patients with CCA. Moreover, multivariate analysis indicated that high level of GLUT1 was an independent predictor for poor survival in CCA patients (HR=1.698, p<0.029; Table II).
Univariate analysis demonstrates the association of Glucose transporter 1 (GLUT1) expression and clinicopathologic characteristics of cholangiocarcinoma (CCA) patients.
Multivariate analysis for overall survival using a Cox proportional hazards regression model.
Up-regulation of GLUT1 is associated with carcinogenesis of CCA. To investigate the association of GLUT1 with carcinogenesis of CCA, GLUT1 expression was evaluated in the liver sections of Ov-induced CCA hamsters collected at 1, 3, and 6 months post-treatment. There were three control groups in this model; the non-treated, Ov infected, and NDMA treated groups. CCA was induced in the Ov + NDMA group. There was no CCA development in the control groups at all treatment periods, whereas CCA was developed in the Ov + NDMA group observed at 3 and 6 month-periods. The IHC results revealed that no GLUT1 signal was observed in the NBD and hepatocytes from all animal groups at every time point (Figure 2A). Most of the hyperproliferative bile ducts from Ov-infected and NDMA treated groups were negative for GLUT1. In contrast, GLUT1 expression was detected in the hyperproliferative bile ducts of the Ov + NDMA treated group as early as 1-month of CCA induction. Membranous staining with moderate to high expression levels of GLUT1 were observed in almost all CCA tissues developed at 3- and 6-months post-treatment (Figure 2A-B). The aberrant expression of GLUT1 in CCA tissues from Ov-induced hamsters indicates the involvement of GLUT1 in CCA development.

GLUT1 expression in the liver sections from the Ov-induced cholangiocarcinoma (CCA) hamster model. GLUT1 was highly expressed in the Ov + NDMA-induced CCA group, but not in the control groups (A). 200× magnification; scale bar=100 μm. The immunohistochemistry (IHC) scores from each histopathologic bile duct from all groups of animals were determined and compared (B). Number=number of bile ducts with positive GLUT1/total. NBD, normal bile duct; HP, hyperplasia; DP, dysplasia; M, month.
GLUT1 expression is associated with progressive phenotypes of CCA cell lines. Two CCA cell lines established from primary CCA tissues of two Thai patients in whom liver-fluke eggs were evident in tumor tissues of KKU-100 and KKU213A (15, 16), were used in this study. Silencing of GLUT1 expression using a specific siRNA towards GLUT1 (siGLUT1) was used to demonstrate the connection of GLUT1 with progressive phenotypes of CCA cell lines. Compared with the scramble control (SC) cells, CCA cells treated with siGLUT1 for 48 h effectively suppressed the expression of GLUT1 in both cell lines to 10-12% of RNA levels and 27-30% of protein levels (Figures 3A-B). The effect of GLUT1 on cells growth was investigated using a colony formation assay. Suppression of GLUT1 expression for 48 h significantly reduced the number of colony formations of KKU-100 to 44% and KKU-213A to 75% of the control cells (p<0.001, Figure 3C).
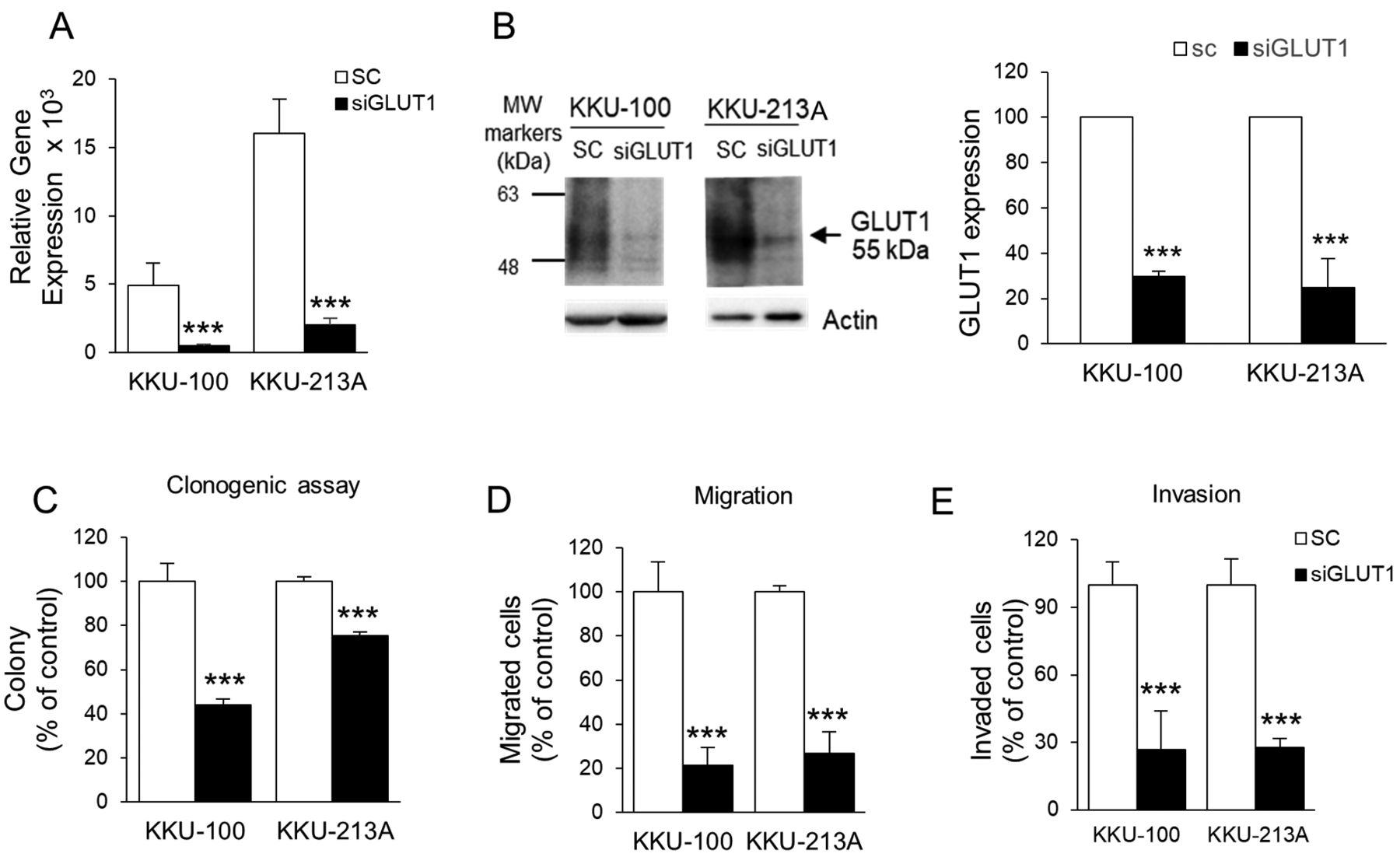
Expression of GLUT1 associated with aggressiveness of cholangiocarcinoma (CCA) cell lines. Two CCA cell lines, KKU-100 and KKU-213A, were treated with siGLUT1 for 48 h. Silencing of GLUT1 expression with siGLUT1 significantly decreased levels of GLUT1 mRNA (A) and GLUT1 protein (B) in CCA cell lines. Colony formation (C), migration (D) and invasion (E) of siGLUT1-treated cells were examined, compared to the scramble control cells (SC). MW, molecular weight, ***p<0.001.
The association of GLUT1 with migration and invasion abilities of CCA cells were next determined using Boyden chamber assays. Cells were treated with siGLUT1 for 48 h and harvested for the migration and invasion assays. siGLUT1 treatment significantly reduced the number of migrating cells of both cell lines to approximately 20-30% of SC cells (p<0.001; Figure 3E). Similar results were observed for the invasion assay. Suppression of GLUT1 expression significantly inhibited invasion ability of both CCA cells to approximately 25% of the control cells (p<0.001; Figure 3F).
Discussion
The expression of GLUT1 in association with progression of CCA has been previously demonstrated in non-fluke related CCA patients and cell lines (HuCCT1 and RBE cells) (12, 22). This study is the first to demonstrate the clinical relevance of GLUT1 expression in CCA tissues and its association with CCA development and an aggressive phenotype of CCA in liver fluke-related CCA. Knock-down of GLUT1 using GLUT1-siRNA suppressed the aggressiveness of CCA cells, suggesting GLUT1 as a potential therapeutic target for CCA.
The clinical relevance of GLUT1 and poor prognosis of CCA patients was emphasized in this study. High expression of GLUT1 was shown to be related with the aggressive non-papillary type CCA, large tumor size, and shorter survival of patients. Moreover, GLUT1 expression was an independent prognostic factor for CCA patients in this cohort. The association of GLUT1 expression with poor prognosis has been repeatedly reported in many cancer types including CCA (12, 22). This collective evidence indicated the involvement of GLUT1 in worse prognosis of CCA patients regardless of the etiology of the disease.
The gradual increasing of GLUT1-positive signals with the increasing degree of bile duct abnormality (hyperplasia, dysplasia and CCA) found in the hamster model, indicated the association of GLUT1 with CCA progression. This association was also observed in CCA tissues from patients. Positive GLUT1 signals were gradually detected in dysplastic lesions and invasive CCA. In addition, high GLUT1 expression was found in the advanced non-papillary type and larger tumor sizes of CCA. The increasing expression of GLUT1 during progression of cancer may be an essential characteristic of cancer as this event was also observed in many types of cancer, such as cervical cancer (23), hepatoma (24), and oral tongue squamous cell carcinomas (25).
The association of GLUT1 with progression of CCA was demonstrated in two CCA cell lines, KKU-100 and KKU-213A. Silencing of GLUT1 expression using siGLUT1 significantly suppressed growth, migration and invasion of these cell lines. A similar observation has also been reported in non-fluke related CCA, in vitro and in vivo (12, 22). Specifically, Kubo et al. (12) demonstrated that suppression of GLUT1 using siRNA suppressed migration of CCA cells. In addition, Tiemin et al. showed that down-regulation of GLUT1 expression using shRNA or micro-RNA (miR-148a) specifically to GLUT1 could significantly reduce the aggressiveness of CCA cells via the reduction of cyclin D1 leading to G1 to S arrest and suppression of MMP2 expression (22). Moreover, suppression of GLUT1 could substantially sensitize CCA cells to gemcitabine in vitro and in vivo (22).
Increased aerobic glycolysis and glucose uptake in cancer cells has been attributed primarily to the up-regulation of GLUT1; hence, targeting GLUT1 as an anti-cancer strategy is well recognized (26). Liu et al. have demonstrated that treatment with WZB117, a synthetic small-molecule GLUT1 inhibitor, could reduce cancer cell growth, migration, and invasion of several cancer cell lines, in vitro and in vivo (26). Lung cancer cells treated with WZB117 had decreased intracellular ATP and glycolytic enzymes resulting in cell-cycle arrest and apoptosis (26). Additionally, it was shown that targeting GLUT1 as an anti-tumor therapy was more effective in inhibiting cancer cells than their non-cancerous cell counterparts and showed a synergistic effect when used in combination with cisplatin or paclitaxel (27).
Conclusion
High expression of GLUT1 was associated with carcinogenesis and progression of CCA, as well as with poor patient outcomes. Decreased GLUT1 expression using siGLUT1 significantly reduced aggressiveness of CCA cell lines, suggesting GLUT1 as a potential therapeutic target of CCA.
Acknowledgements
This project was supported by the RGJ-PhD Program, TRF (PHD/0100/2560) to UT and SW; the co-funding of Khon Kaen University to RGJ (2561); the research grant from the Faculty of Medicine, (IN61328) and Cholangiocarcinoma Research Institute, Khon Kaen University to UT (CARI 01/2560). The authors would like to thank Prof. James A Will for editing this manuscript via the Faculty of Medicine Publication Clinic, KKU.
Footnotes
Authors’ Contributions
UT performed the experiments, data analysis and drafted the manuscript. SS examined the immunohistochemistry and analysis. PS performed cell cultures. UC participated in the design of animal study. SW and WS conceived the study and performed analysis and data interpretation. SW supervised the study and revised the manuscript. All authors participated in the interpretation of the studies and review of the manuscript.
This article is freely accessible online.
Conflicts of Interest
The Authors declare no conflicts of interest.
- Received August 1, 2020.
- Revision received October 9, 2020.
- Accepted October 12, 2020.
- Copyright© 2021, International Institute of Anticancer Research (Dr. George J. Delinasios), All rights reserved




{kind=link}
{kind=link}
{kind=link}