Butyric Acid Added Apically to Intestinal Caco-2 Cells Elevates Hepatic ApoA-I Transcription and Rescues Lower ApoA-I Expression in Inflamed HepG2 Cells Co-Cultured in the Basolateral Compartment


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
2.1. Materials
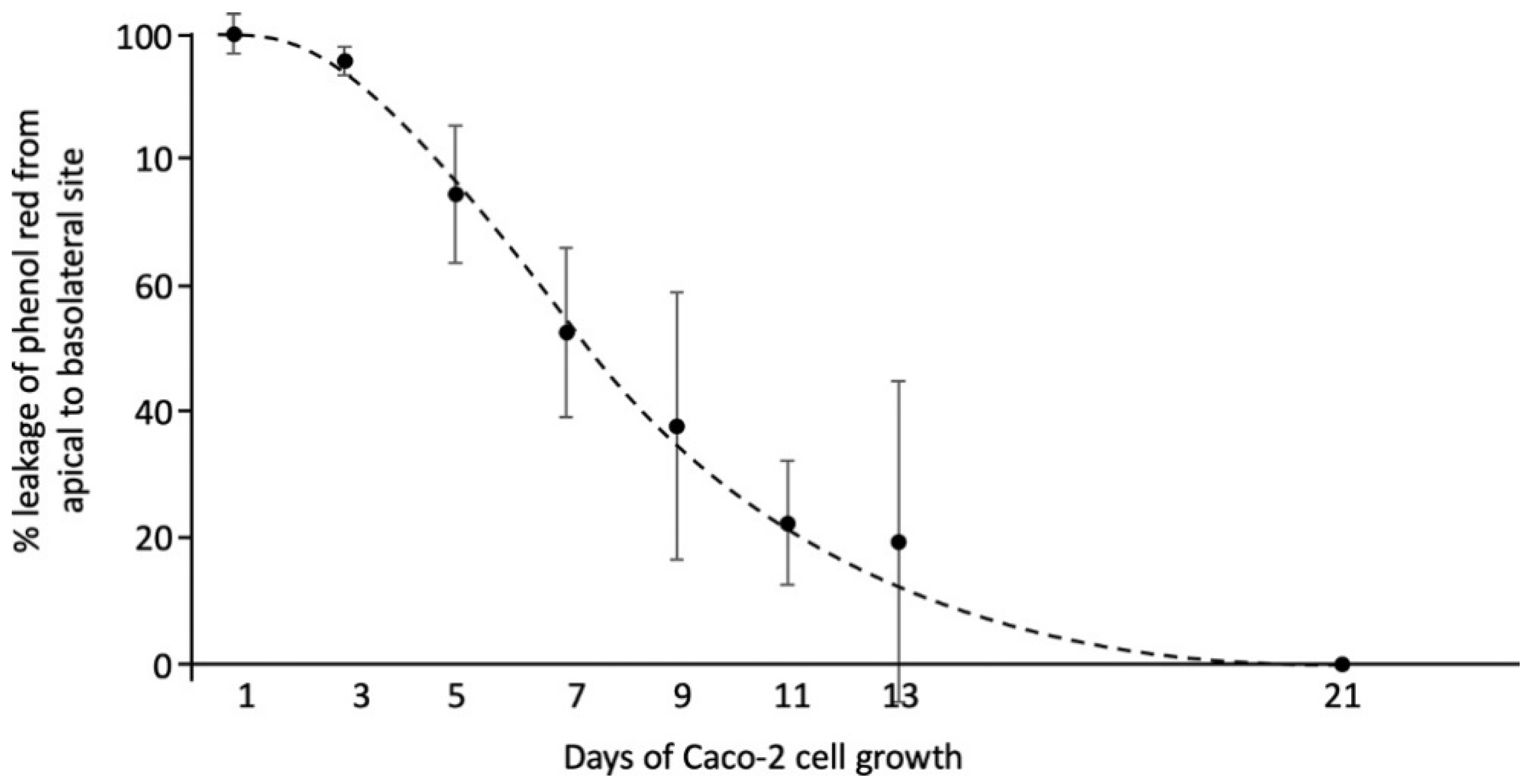
2.2. Cell Culture and C4 Treatment
2.3. Quantification of Gene mRNA Transcription
2.4. Statistical Analysis
3. Results
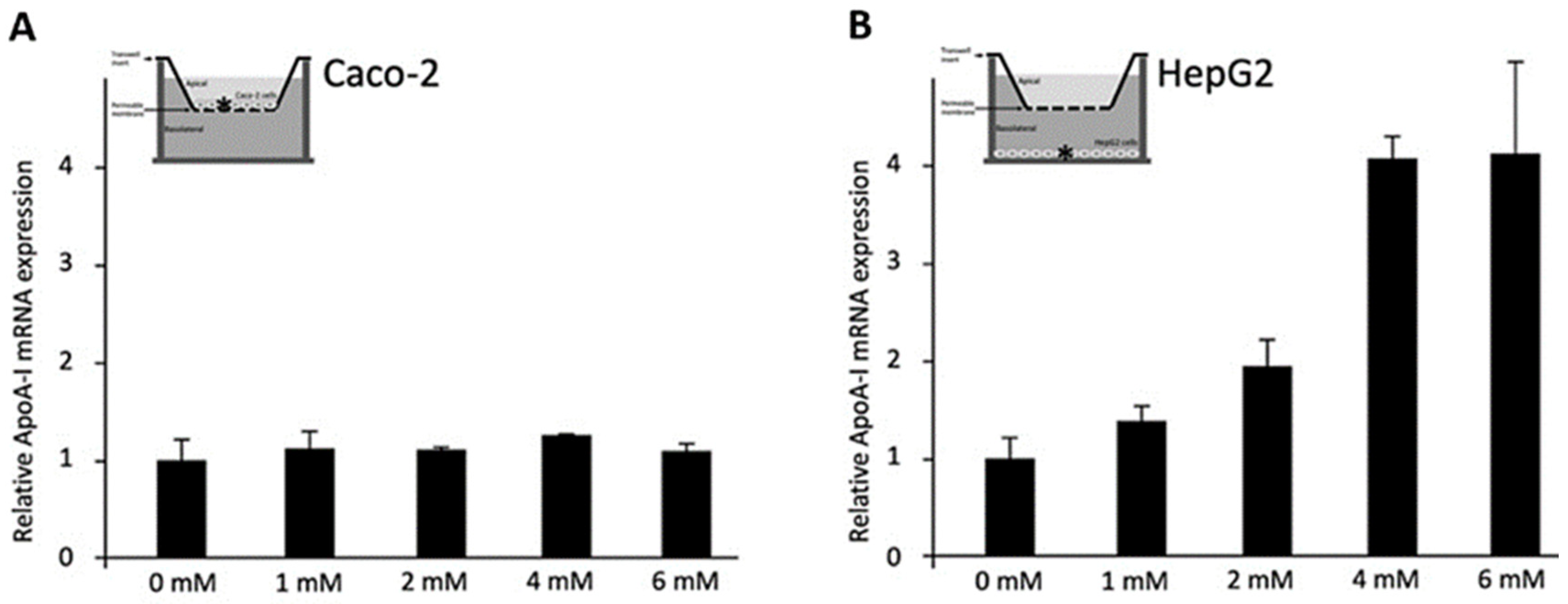
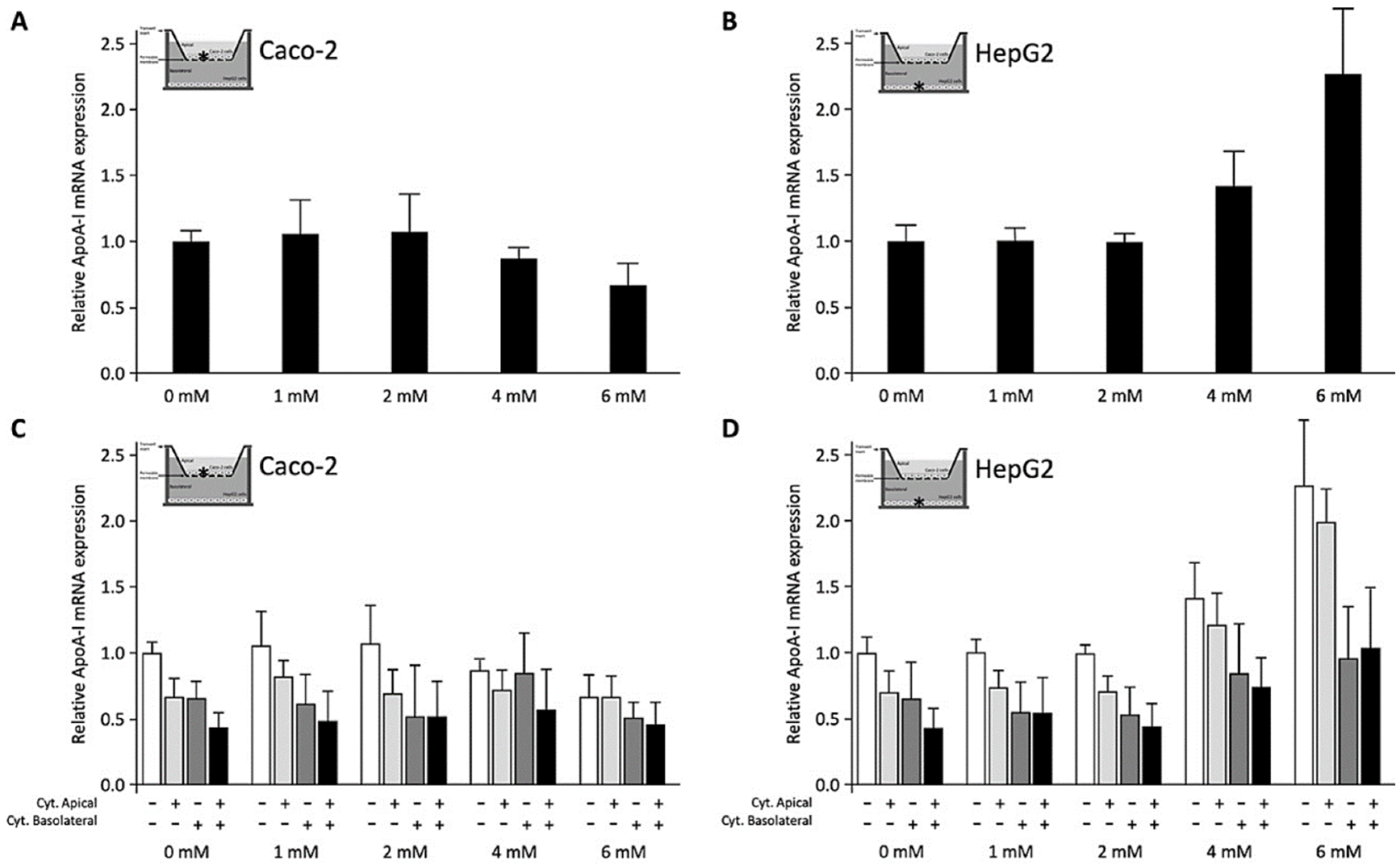
3.1. Effects of C4 on ApoA-I mRNA Expression in Caco-2 or HepG2 Cells (Conditions 1 and 2)
3.2. Effects of C4 Added to the Apical Surface of Caco-2 Cells on ApoA-I mRNA Expression in Caco-2 and HepG2 Cells (Condition 3)
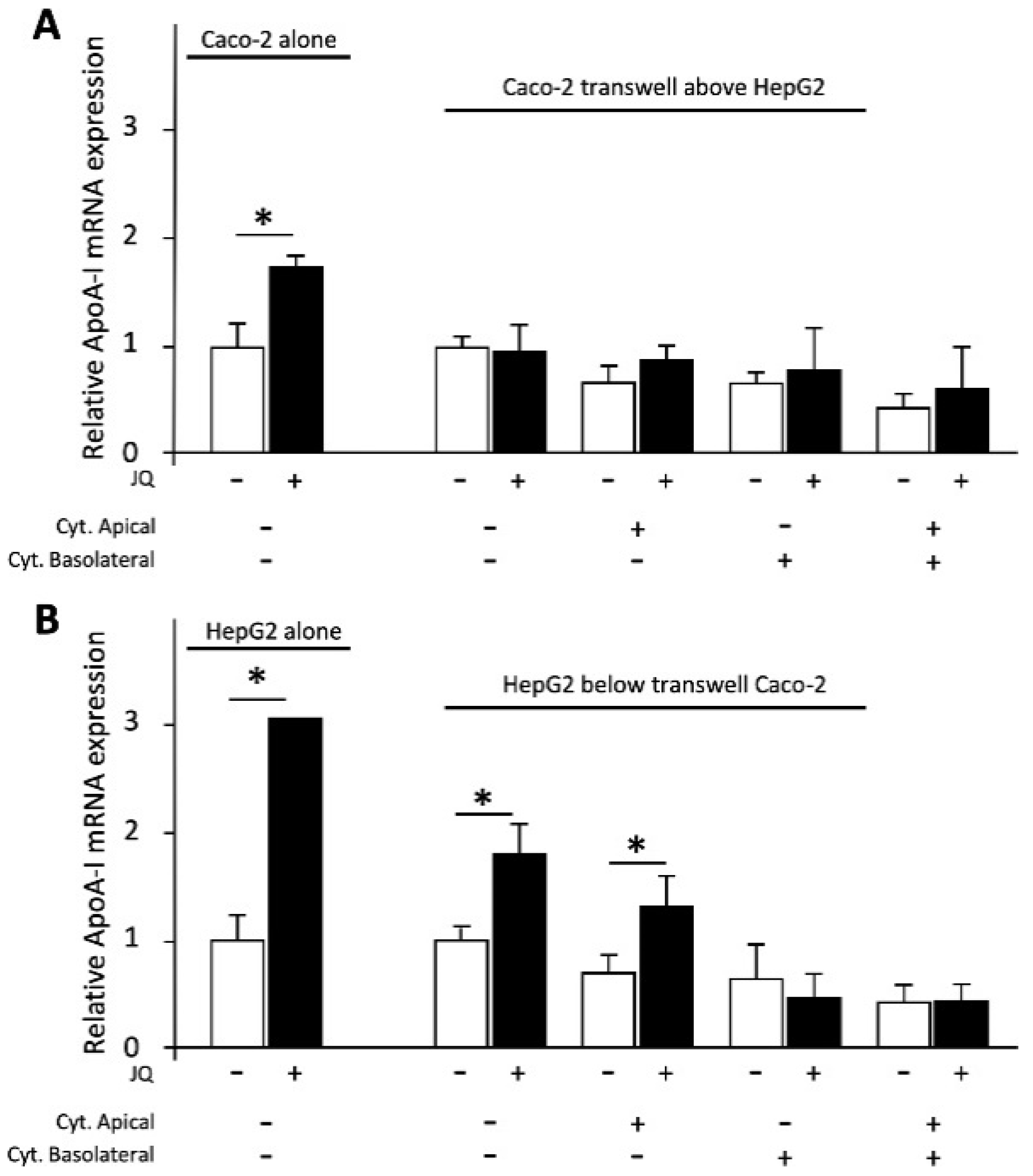
3.3. Effects of the Positive Control JQ1(+) on ApoA-I mRNA Expression in Caco-2 and HepG2 Cells
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ApoA-I | Apolipoprotein A-I |
| cDNA | Complementary DNA |
| IL-1β | Interleukin 1 beta |
| NF-κB | Nuclear factor kappa B |
| SCFAs | Short chain fatty acids |
| TNFα | Tumor necrosis factor-α |
| mRNA | messenger RNA |
References
- Kles, K.A.; Chang, E.B. Short-chain fatty acids impact on intestinal adaptation, inflammation, carcinoma, and failure. Gastroenterology 2006, 2, S100–S105. [Google Scholar] [CrossRef] [PubMed]
- Waldecker, M.; Kautenburger, T.; Daumann, H.; Veeriah, S.; Will, F.; Dietrich, H. Histone-deacetylase inhibition and butyrate formation: Fecal slurry incubations with apple pectin and apple juice extracts. Nutrition 2008, 4, 366–374. [Google Scholar] [CrossRef] [PubMed]
- Robles-Vera, I.; Toral, M.; de la Visitación, N.; Aguilera-Sánchez, N.; Redondo, J.M.; Duarte, J. Protective Effects of Short-Chain Fatty Acids on Endothelial Dysfunction Induced by Angiotensin II. Front. Physiol. 2020, 11, 277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Canani, R.B.; Di Costanzo, M.; Leone, L.; Pedata, M.; Meli, R.; Calignano, A. Potential beneficial effects of butyrate in intestinal and extraintestinal diseases. World J. Gastroenterol. 2011, 12, 1519. [Google Scholar] [CrossRef]
- Segain, J.; De La Blétiere, D.R.; Bourreille, A.; Leray, V.; Gervois, N.; Rosales, C. Butyrate inhibits inflammatory responses through NFκB inhibition: Implications for Crohn’s disease. Gut 2000, 3, 397–403. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; van Esch, B.C.; Wagenaar, G.T.; Garssen, J.; Folkerts, G.; Henricks, P.A. Pro-and anti-inflammatory effects of short chain fatty acids on immune and endothelial cells. Eur. J. Pharmacol. 2018, 11, 52–59. [Google Scholar] [CrossRef]
- Ohira, H.; Tsutsui, W.; Fujioka, Y. Are short chain fatty acids in gut microbiota defensive players for inflammation and atherosclerosis? J. Atheroscler. Thromb. 2017, 7, 660–672. [Google Scholar] [CrossRef] [Green Version]
- Thibault, R.; Blachier, F.; Darcy-Vrillon, B.; De Coppet, P.; Bourreille, A.; Segain, J.-P. Butyrate utilization by the colonic mucosa in inflammatory bowel diseases: A transport deficiency. Inflammatory bowel diseases. Inflamm. Bowel Dis. 2010, 4, 684–695. [Google Scholar] [CrossRef]
- Tayyeb, J.Z.; Popeijus, H.E.; Mensink, R.P.; Konings, M.C.; Mulders, K.H.; Plat, J. The effects of short-chain fatty acids on the transcription and secretion of apolipoprotein A-I in human hepatocytes in vitro. J. Cell. Biochem. 2019, 10, 17219–17227. [Google Scholar] [CrossRef] [Green Version]
- Bloemen, J.G.; Venema, K.; van de Poll, M.C.; Damink, S.W.O.; Buurman, W.A.; Dejong, C.H. Short chain fatty acids exchange across the gut and liver in humans measured at surgery. Clin. Nutr. 2009, 6, 660–672. [Google Scholar] [CrossRef]
- Den Besten, G. Elucidating the Mechanisms of Action of Short-Chain Fatty Acids: From Dietary Fiber to Host Metabolism. Ph.D. Thesis, Groningen University, Groningen, The Netherlands, 2014. [Google Scholar]
- Tayyeb, J.Z.; Popeijus, H.E.; Mensink, R.P.; Konings, M.; Mokhtar, F.B.A.; Plat, J. Short-Chain Fatty Acids (Except Hexanoic Acid) Lower NF-kB Transactivation, Which Rescues Inflammation-Induced Decreased Apolipoprotein A-I Transcription in HepG2 Cells. Int. J. Mol. Sci. 2020, 14, 5088. [Google Scholar] [CrossRef] [PubMed]
- Carpintero, R.; Pineiro, M.; Andres, M.; Iturralde, M.; Alava, M.; Heegaard, P.M. The concentration of apolipoprotein AI decreases during experimentally induced acute-phase processes in pigs. Infect. Immun. 2005, 5, 3184–3187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyazaki, A.; Sakuma, S.; Morikawa, W.; Takiue, T.; Miake, F.; Terano, T. Intravenous injection of rabbit apolipoprotein AI inhibits the progression of atherosclerosis in cholesterol-fed rabbits. Arterioscler. Thromb. Vasc. Biol. 1995, 11, 1882–1888. [Google Scholar] [CrossRef] [PubMed]
- Shah, P.K.; Yano, J.; Reyes, O.; Chyu, K.-Y.; Kaul, S.; Bisgaier, C.L. High-Dose Recombinant Apolipoprotein A-IMilano Mobilizes Tissue Cholesterol and Rapidly Reduces Plaque Lipid and Macrophage Content in Apolipoprotein E–Deficient Mice: Potential Implications for Acute Plaque Stabilization. Circulation 2001, 25, 3047–3050. [Google Scholar] [CrossRef] [Green Version]
- Cummings, J.; Pomare, E.; Branch, W.; Naylor, C.; Macfarlane, G. Short chain fatty acids in human large intestine, portal, hepatic and venous blood. Gut 1987, 10, 1221–1227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmitt, M.G., Jr.; Soergel, K.H.; Wood, C.M. Absorption of short chain fatty acids from the human jejunum. Gastroenterology 1976, 2, 211–215. [Google Scholar] [CrossRef]
- Lammi, C.; Zanoni, C.; Ferruzza, S.; Ranaldi, G.; Sambuy, Y.; Arnoldi, A. Hypocholesterolaemic activity of lupin peptides: Investigation on the crosstalk between human enterocytes and hepatocytes using a co-culture system including Caco-2 and HepG2 cells. Nutrients 2016, 7, 437. [Google Scholar] [CrossRef] [Green Version]
- Van Breemen, R.B.; Li, Y. Caco-2 cell permeability assays to measure drug absorption. Expert Opin. Drug Metab. Toxicol. 2005, 2, 175–185. [Google Scholar] [CrossRef]
- Garcia, M.N.; Flowers, C.; Cook, J.D. The Caco-2 cell culture system can be used as a model to study food iron availability. J. Nutr. 1996, 1, 251–258. [Google Scholar] [CrossRef] [Green Version]
- Van der Krieken, S.E.; Popeijus, H.E.; Mensink, R.P.; Plat, J. Link between ER-stress, PPAR-alpha activation, and BET inhibition in relation to apolipoprotein A-I transcription in HepG2 cells. J. Cell. Biochem. 2017, 8, 2161–2167. [Google Scholar] [CrossRef]
- Wu, A.L.; Windmueller, H.G. Relative contributions by liver and intestine to individual plasma apolipoproteins in the rat. J. Biol. Chem. 1979, 15, 7316–7322. [Google Scholar] [CrossRef]
- Peng, L.; He, Z.; Chen, W.; Holzman, I.R.; Lin, J. Effects of butyrate on intestinal barrier function in a Caco-2 cell monolayer model of intestinal barrier. Pediatr. Res. 2007, 1, 37–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glickman, R.M.; Green, P.H. The intestine as a source of apolipoprotein A1. Proc. Natl. Acad. Sci. USA 1977, 6, 2569–2573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neis, E.P.; van Eijk, H.M.; Lenaerts, K.; Damink, S.W.O.; Blaak, E.E.; Dejong, C.H. Distal versus proximal intestinal short-chain fatty acid release in man. Gut 2019, 4, 764–765. [Google Scholar] [CrossRef] [PubMed]
- Parada Venegas, D.; De la Fuente, M.K.; Landskron, G.; González, M.J.; Quera, R.; Dijkstra, G. Short Chain Fatty Acids (SCFAs)-mediated gut epithelial and immune regulation and its relevance for inflammatory bowel diseases. Front. Immunol. 2019, 10, 277. [Google Scholar] [CrossRef] [Green Version]
- Albillos, A.; de Gottardi, A.; Rescigno, M. The gut-liver axis in liver disease: Pathophysiological basis for therapy. J. Hepatol. 2020, 3, 558–577. [Google Scholar] [CrossRef] [Green Version]
- Trapecar, M.; Communal, C.; Velazquez, J.; Maass, C.A.; Huang, Y.-J.; Schneider, K. Gut-Liver physiomimetics reveal paradoxical modulation of IBD-related inflammation by short-chain fatty acids. Cell Syst. 2020, 3, 223–239. [Google Scholar] [CrossRef]
- Kan, H.-Y.; Georgopoulos, S.; Zannis, V. A hormone response element in the human apolipoprotein CIII (ApoCIII) enhancer is essential for intestinal expression of the ApoA-I and ApoCIII genes and contributes to the hepatic expression of the two linked genes in transgenic mice. J. Biol. Chem. 2000, 39, 30423–30431. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tayyeb, J.Z.; Popeijus, H.E.; Mensink, R.P.; Plat, J. Butyric Acid Added Apically to Intestinal Caco-2 Cells Elevates Hepatic ApoA-I Transcription and Rescues Lower ApoA-I Expression in Inflamed HepG2 Cells Co-Cultured in the Basolateral Compartment. Biomolecules 2021, 11, 71. https://doi.org/10.3390/biom11010071
Tayyeb JZ, Popeijus HE, Mensink RP, Plat J. Butyric Acid Added Apically to Intestinal Caco-2 Cells Elevates Hepatic ApoA-I Transcription and Rescues Lower ApoA-I Expression in Inflamed HepG2 Cells Co-Cultured in the Basolateral Compartment. Biomolecules. 2021; 11(1):71. https://doi.org/10.3390/biom11010071
Chicago/Turabian StyleTayyeb, Jehad Z., Herman E. Popeijus, Ronald P. Mensink, and Jogchum Plat. 2021. "Butyric Acid Added Apically to Intestinal Caco-2 Cells Elevates Hepatic ApoA-I Transcription and Rescues Lower ApoA-I Expression in Inflamed HepG2 Cells Co-Cultured in the Basolateral Compartment" Biomolecules 11, no. 1: 71. https://doi.org/10.3390/biom11010071